
Extracellular Vesicles in the Fungi Kingdom
by
 and
and
Marc Liebana-Jordan
1,†, 
Bruno Brotons
1,†,
Juan Manuel Falcon-Perez
1,2,3,* and
Esperanza Gonzalez
1,* 1
Exosomes Laboratory, Center for Cooperative Research in Biosciences (CIC bioGUNE), Basque Research and Technology Alliance (BRTA), 48160 Derio, Spain
2
Centro de Investigación Biomédica en Red de Enfermedades Hepáticas y Digestivas (CIBERehd), 28029 Madrid, Spain
3
IKERBASQUE Basque Foundation for Science, 48009 Bilbao, Spain
*
Authors to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Int. J. Mol. Sci. 2021, 22(13), 7221; https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22137221
Submission received: 7 May 2021
/
Revised: 14 June 2021
/
Accepted: 28 June 2021
/
Published: 5 July 2021
(This article belongs to the Special Issue Extracellular Vesicles in Phylogeny)
Abstract
:Extracellular vesicles (EVs) are membranous, rounded vesicles released by prokaryotic and eukaryotic cells in their normal and pathophysiological states. These vesicles form a network of intercellular communication as they can transfer cell- and function-specific information (lipids, proteins and nucleic acids) to different cells and thus alter their function. Fungi are not an exception; they also release EVs to the extracellular space. The vesicles can also be retained in the periplasm as periplasmic vesicles (PVs) and the cell wall. Such fungal vesicles play various specific roles in the lives of these organisms. They are involved in creating wall architecture and maintaining its integrity, supporting cell isolation and defence against the environment. In the case of pathogenic strains, they might take part in the interactions with the host and affect the infection outcomes. The economic importance of fungi in manufacturing high-quality nutritional and pharmaceutical products and in remediation is considerable. The analysis of fungal EVs opens new horizons for diagnosing fungal infections and developing vaccines against mycoses and novel applications of nanotherapy and sensors in industrial processes.
1. General Concepts on Extracellular Vesicles
Extracellular vesicles (EVs) are membranous structures released by prokaryotic and eukaryotic cells during normal physiological functioning, whose secretion is altered in pathophysiological states. The biological importance of these vesicles relies on their capacity to mediate in intercellular communication by transferring information between cells [1].
The EVs have been widely characterized in mammalian cells and can be broadly divided into three main categories. The first type, exosomes, range from ~40 to 160 nm in diameter and are of endosomal origin. They are formed by the fusion of multivesicular bodies (MVBs) with the plasma membrane. After fusion, the intraluminal vesicles (ILVs) conforming MVBs are released from the cells, becoming exosomes. The second group, ectosomes, includes microvesicles, microparticles and large vesicles from ~50 nm to 1 µm in diameter. These vesicles pinch off the surface of the plasma membrane via outward budding. Finally, apoptotic bodies, ranging from 50 nm to 5 µm in diameter, are formed during plasma membrane blebbing in apoptosis. It has been widely accepted that these three EV subgroups have their specific features; however, there is no consensus on particular distinguishing markers [2].
Over the past few years, EVs have been isolated from nearly all cell types and biological fluids [3]. Their characteristic bilayer membrane, enriched in sphingomyelin, cholesterol, phosphatidylserine (PS) and glycosphingolipids, contributes to their stability in different extracellular environments, making them excellent shuttles for various molecules [1]. The EVs can carry many different macromolecules such as proteins, nucleic acids (both RNA and DNA), carbohydrates and metabolites that can act as signals [1]. The vesicles transport such macromolecules across biological barriers to the neighbouring and remote cells in specific organs or tissues via circulatory systems (blood, lymph). Thus, they can affect the function of recipient cells in multicellular organisms. The size, membrane composition and content of EVs are highly heterogeneous. They are dynamic and strongly dependent on the source cell, cell topography, state and environmental conditions. All these factors determine their functionality and target specificity. The same cell type may secrete different types of vesicles, depending on these factors [4,5].
Despite all the research already conducted in the field, the physiological purpose of generating these EVs remains largely unknown and needs further investigation. Most of the available information on their biosynthesis, composition and function comes from mammalian systems, presenting the EVs as novel players in cell communication and signalling as mentioned above. However, EVs released by other multicellular and unicellular organisms and cross-kingdom EV-based communication have also been described. For example, human–fungus and plant–fungus interactions have been reported. It has been suggested that fungal virulence is enhanced by EVs containing small RNA (sRNA). In turn, the plants can secrete EVs carrying sRNA to silence the fungal infection [6].
This review describes and discusses the properties of fungal EVs. Their functionality and the still unanswered questions are presented, including the cellular origin and biogenesis of EVs, transit mechanisms through the fungal cell wall and the importance and packaging of the cargo.
2. History of Fungal EVs
Fungi form a diverse kingdom of eukaryotic organisms ranging from yeasts and moulds to mushrooms. Some species fill biological niches of basic decomposers, play a key role in recycling organic matter and others, such as many unicellular yeasts and filamentous fungi, are human, animal or plant pathogens. They can have a considerable impact on human lives. Widening our understanding of fungal biology is necessary to improve the treatment and prevention of fungal diseases. The emerging antifungal drug resistance may bring considerable epidemiological risks [7]. Only an effective control of pathogenic fungi that infect livestock and agricultural holdings can avoid substantial economic losses. However, fungi are also exploited in biotechnological processes and the food industry; thus, understanding their biology will help optimise the processes and prevent problems in end-product manufacturing.
Unicellular fungi such as yeasts are key models in science since, as simple eukaryotes, they can reflect the basic cellular physiology and molecular biology of more complex eukaryotic organisms. In similarity with the bacterial and plant cells [6], fungal cells are protected by a thick, well-built cell wall composed of glycoproteins and carbohydrate polymers, with the kingdom-defining polysaccharide chitin [8]. This barrier protects them from the external environment and limits the exchange and communication in both directions. In this scenario, intercellular communication must be conducted, at least in part, via EVs (like in mammals).
Research on fungal EVs is still in its infancy. They were first observed in the early 1970s, using the freeze-etching technique [9]. The interest in these vesicles later declined and reawakened at the beginning of the 21st century. A decade ago, fungal EVs were finally described and characterised in the study of macromolecules crossing the cell wall in the pathogen Cryptococcus neoformans [10]. This yeast secretes EVs containing glucuronoxylomannan (GXM), a component of the cryptococcal capsule and key lipids such as glucosylceramide and sterols. Since then, the EVs have been characterised in several other fungal species, both pathogenic and non-pathogenic (Figure 1) [11,12,13,14,15].
In addition, C. neoformans, EVs from other pathogenic fungi have been studied. Some of these fungi are species with particular prevalence in tropical and subtropical regions, such as Histoplasma capsulatum [16] and Paracoccidioides brasiliensis [17]. Others were the species with worldwide prevalence, Malassezia sympodialis [18] and the opportunist Candida albicans [19]. Recently, the opportunist Exophiala dermatitidis [20] and Cryptococcus gattii [21] have also attracted attention in the EV field, as well as Sporothrix brasiliensis [22], responsible for the emergent zoonosis in Brasil. EV secretion has also been investigated in fungus species with industrial interest. One example is C. albicans due to its role in the heavy metal accumulation and its possible application in remediation. Saccharomyces cerevisiae [11,23] and Pichia fermentans (used in the production of bread, beer and wine) and other less known fungi such as Torulaspora delbrueckii, Candida sake, Hanseniaspora uvarum, Metschnikowia pulcherrima, and Lachancea thermotolerans have been explored [24]. Other fungi with economic impact (infecting crops) like Fusarium oxysporum (affecting banana trees) have also been examined (Figure 1).
3. Characteristics and Composition of Fungal EVs
In terms of molecular content, fungal EVs show strong similarities to mammalian EVs [25]. In both cases, the EVs transport several RNA species belonging to a wide range of functional categories, including messenger RNA (mRNA). They transfer several types of non-coding RNAs, such as transfer RNA (tRNA), ribosomal RNA (rRNA) and small non-coding RNAs like micro-RNA (miRNA), small nuclear RNA (snRNA) and small nucleolar RNA (snoRNA), among others (Table 1). The EVs are efficient vehicles for cell–to–cell transport of RNA since their structure protects the RNA from RNase degradation. Thus, this type of intercellular communication might affect the regulation of gene expression and alter important processes such as infection persistence or suppression of virulence factors [21,26].
Proteomic characterisation of EVs has detected proteins representing almost all functional categories, from pathogenesis and immune response to nutrition, metabolism, signalling and trafficking. These proteins come from different cellular compartments, such as the cell wall, plasma membrane, cytoplasm, mitochondria, vacuoles and even the nucleus (Table 1). Some of them are the proteins common in several fungus species or have orthologs in other species [15]. There are also some species-specific proteins, e.g., several septins found in P. brasiliensis EVs [14].
Lipid profiles of EVs from several fungus species, C. neoformans, H. capsulatum, P. brasiliensis, C. albicans and C. gattii, have also been characterised using different techniques such as thin-layer chromatography, electrospray ionisation or gas-chromatography-mass spectrometry. The analysis has shown some peculiarities in the membrane of these EVs (Table 2). The main sterol-derivatives detected are ergosterol [10,17,19,27] and lanosterol [17,19,27], whereas the most abundant neutral glycosphingolipid (GSL) in fungi is glucosylceramide (GlcCer) [10,19,27]. Moreover, several types of monohexosylceramide have also been found in different strains of P. brasiliensis [28]. Finally, there is an abundance of phospholipids such as phosphatidylethanolamine (PE), phosphatidylserine (PS), phosphatidylcholine (PC), phosphatidic acid (PA), phosphatidylinositol (PI) and phosphatidylglycerol (PG) [16,27,29]. The main fatty acids encountered are linoleic, oleic, palmitic, stearic and pentadecanoic acid [27,29].
Thus, the fungal EVs carry many common but also some kingdom-specific components. It should be noted that the composition of EVs from fungi depends on the species, cellular and environmental conditions and the experimental scenario. Such a composition makes EVs the active machines they are.
4. Biogenesis of Fungal EVs
4.1. Etiology of Fungal EVs
Both the EVs and periplasmic vesicles (PVs) have been described in fungi; however, it is still unclear whether the vesicles observed inside and outside the cell walls are the same. In S. cerevisiae, both types have similar rounded shape and size and similar protein composition [23,37]. PVs comprise two populations whose biogenesis is orchestrated by the nutrient environment. In particular, the subpopulation comprising the smallest vesicles along with their gluconeogenic enzymes content increase in response to glucose starvation [23]. However, the vesicles found in the extracellular space (EVs) have an ovoid or rounded shape in all species studied so far. They are mostly from 30 to 300 nm in size, although smaller and larger EVs are also detected. Like in PVs, their secretion is also affected by nutrient accessibility and growth conditions [31]. These findings support the idea that the nutritional environment can determine the profile of EVs secreted by a certain group of cells, reflecting an adaptation response to the medium. Indeed, the fungi can secrete EVs when cultured not only in liquid but also in solid media [32] and while forming biofilms [39,40]. In C. albicans, the EVs produced inside the biofilm are different from those formed by the free-living cells, both in size and composition [39] (Table 3).
4.2. Crossing External Barriers
The cell wall acts as a barrier in the final steps of EV release (Figure 2). C. neoformans and S. cerevisiae mutants with defects in cell wall remodelling and dynamics (such as chs3 and fks1 [49]) as well as chs1 mutant in S. cerevisiae [44] show increased EV release, reflecting their weakened cell-wall phenotype. Caspofungin, an inhibitor of Fks1, can mimic this phenotype [44]. Turgor pressure could force the vesicles through the cell wall, given the viscoelastic properties of this barrier observed in C. albicans and C. neoformans [50].
In capsule-forming species like C. neoformans, the capsule will also work as a barrier in the EV release. An increase in EV release and virulence factors has been observed in the acapsular mutant Δcap67 [10,32]. The elevated laccase and acid phosphatase virulence factors in the cell surfaces of Δcap10 mutants might also be associated with increased EV secretion [51].
4.3. Trafficking EVs
Proteomic and in silico studies of many fungal species had provided evidence on trafficking pathways involved in EVs biogenesis even before any experimental proof of their existence was obtained. Prediction of signal peptide cleavage sites in S. cerevisiae shows that some EV proteins target the ER and the conventional secretory pathway (involving the Golgi apparatus and the plasma membrane). Moreover, the GPI-anchored proteins are 4-fold enriched in the EVs compared to the genome background. This difference in their abundance suggests that GPI-anchor modifications tag the proteins to be transported in the vesicles [11]. Finally, S. cerevisiae EVs are also enriched in proteins containing AAA domains, which are implicated in membrane fusion, indicating a role of the membrane system in their biogenesis [44]. In the case of P. brasiliensis, 61% of the EV proteins have been rated as secretory, although only 10% have a predicted signal peptide. This result supports the hypothesis proposing various ways for proteins to reach or incorporate into the EVs. Regarding the putative GPI-anchored proteins, only 0.98% are represented in the EVs from P. brasiliensis [14]. In C. albicans, predicted signal peptides are detected in similar proportions, in 60% of proteins, of which, almost 20% are putative GPI-anchored proteins. The remaining 40% of proteins are not predicted to be secreted [38].
Poor understanding of the mechanisms of EV biogenesis is one of the main factors impeding the study of these vesicles, their associated pathways and regulators. Apart from in silico strategies, genetic studies have brought the most informative results. After many years invested in studying fungal EVs, the genetic and molecular evidence has shown, like in mammalians [52], the involvement of both conventional secretory pathway (endoplasmic reticulum (ER)–Golgi Apparatus (GA)–exocyst–plasma membrane axis) and the ESCRT-mediated MVB pathway (early-sorting-endosomes (ESEs)–late-sorting endosomes (LSEs)–MVBs pathway) in fungal EV biogenesis and cargo loading. There are also some additional "alternative" routes and regulators [53] (Figure 2). However, the relative contribution and biological significance of these pathways remain unclear. Mutations in the regulators of both conventional secretory and MVB pathways affect fungal EV composition and release kinetics (Table 4). Nevertheless, none of these mutants shows a complete abrogation of EV formation, suggesting that other pathways are implicated in vesicle generation.
4.3.1. Conventional Pathway
In the conventional secretory pathway, transport protein Sec1, essential for the stimulation of membrane fusion, concentrates at the secretion sites. Sec4, a GTP-binding protein, regulates polarised delivery of vesicles to the exocyst at the plasma membrane. Transport protein Bos1, localised at the ER membrane, is necessary for the correct flow of vesicular transport from the ER to the Golgi. Mutations in the corresponding genes of S. cerevisiae affect the release of EVs [11] (Figure 2). In sec4 mutants, EV secretion is decreased and delayed. The population size distribution changes: sec4 mutant secretes vesicles that are distributed in two populations, from 80 to 120 and 400 to 550 nm (normally, the sizes range from 100 to 200 nm). Mutations in SEC1, SEC4 and BOS1 also alter protein abundance in the EVs without changing their composition. Mutations in SEC1 and SEC4 cause a decrease in the EV sterol content, accompanied by an intracellular accumulation of sterols [11] (Table 4).
The exocyst, an octameric protein complex, is involved in the final step of the conventional secretory pathway, necessary for the polarised fusion of the exocytic vesicles with the plasma membrane (Figure 2). In C. neoformans, a knockdown of the SEC6 component suppresses the EVs release and results in an accumulation of large cytoplasmic vesicles and 100 nm vesicles in the cellular bud necks. This is accompanied by a reduction in laccase and urease activity and in the levels of glucan cell-wall polysaccharide. In contrast, other virulence factors such as extracellular phospholipase activity and capsule production are unaffected [54] (Table 4). This duality of SEC6 knockdown on virulence factors secretion suggests the existence of different secretory pathways that, in turns, can be associated with EVs or not.
4.3.2. GRASPs, Autophagy, IVCs and Endocytosis
The Golgi reassembly and stacking proteins (GRASP) tether the vesicles destined to fuse with the Golgi apparatus and sustain the Golgi structure to maintain the flow of proteins among the different cisternae [58,59] (Figure 2). Deleting the single ortholog of C. neoformans affects the EV release and sizes higher than 300 nm are no longer observed. Moreover, some of the EV mRNAs are enriched [35,55]. When GRASP deletion, GXM polysaccharide content is reduced and, consequently, the capsule formation [35,55]. As already mentioned for the sec1 and sec4 mutants [11], the EVs in the yeast lacking GRASP show reduced sterol levels; this is accompanied by intracellular sterol accumulation (Table 4). Autophagosomes have related to GRASP unconventional secretory pathway [60]; autophagy regulator Atg7 affects both physiological and pathogenic mechanisms in C. neoformans [61]. In agreement with this, atg7 mutant is also affected in EV release and content [35].
The intracellular vesicle clusters (IVCs) are apparently not associated with the conventional secretory pathway [62]. However, as some of the IVCs resident proteins in S. cerevisiae, such as gluconeogenic enzymes FBPase, Gapdh and Vid24p, are associated with EVs, the role of this compartment in PVs biogenesis has been examined. Nevertheless, reducing the IVCs activity does not affect the number of EVs in the periplasmic space [62].
The response of PVs to glucose starvation is regulated by endocytosis, involving the actin cytoskeleton-regulatory complex protein End3. This is accomplished through the sequential recruitment at endocytic sites of the proteins driving the cargo sorting, membrane invagination and vesicle release [23]. This mechanism has not been explored in the EVs; its further examination might shed light on the similarities between the periplasmic and extracellular vesicles (Table 4).
4.3.3. The Non-Conventional Secretory Pathway
In the non-conventional pathway involving the MVBs (Figure 2), Vps27 protein (a member of the ESCRT-0 complex) is required for sorting ubiquitinated membrane proteins into intraluminal vesicles before vacuolar degradation. Vps23 is a core component of the ESCRT-I complex, associated with the ubiquitin-dependent sorting of proteins into the endosome. Vps36, a member of the ESCRT-II complex, is involved in interactions with the ESCRT-I. Snf7/Vps32 and Vps2 are the essential members of the ESCRT-III and take part in the sorting of transmembrane proteins into the MVB pathway. Finally, Bro1 is a cytoplasmic class E VPS factor coordinating deubiquitination, recruited to MVBs by interacting with the ESCRT-III subunit Snf7/Vps32 [63].
In C. neoformans vps27(0), an accumulation of MVBs and vacuole fragmentation have been observed, while the cytoskeleton is not affected. The EVs in this mutant are larger (diameter over 200 nm) but their lipid composition is unaltered [49]. In S. cerevisiae, vps23 (I) and vps36 (II) knockouts result in a significant increase in the proportion of large EVs (150–500 nm) and a reduction in the relative abundance of vesicles between 30 and 150 nm in diameter [44]. Mutant strains vps23 (I), vps 36 (II), snf7/vps32 (III) and vps2 (III) produce EVs with lower protein content than the vesicles from wild type cells and vps27 and bro1 mutants [11,44]. Similarly, the vps23 (I), vps 36 (II) and vps2 (III) mutations alter the EV protein composition, while the vps27 (0) and bro1 mutants do not affect it [44]. Notably, the EVs from mutants of ESCRT components are enriched in proteins containing AAA domains, which are implicated in membrane fusion [11] (Table 4).
Interestingly, the EVs from S. cerevisiae vps23 (I) and vps36 (II) mutants are gained in cell-wall related proteins such as chitin synthase Chs3. Increased levels of the subunit of the major 1,3-beta-glucan synthase Fks1 have been found not only in the EVs from vps23 (I) and vps36 (II) but also from vps 2 (III) mutants [44]. In C. neoformans vps27 mutant, the capsule is reduced and virulence factors, like laccase, are accumulated mostly in EVs rather than in the cell walls. Surprisingly, the LAC1 gene that codify for laccase is downregulated to compensate for trafficking defects. In this C. neoformans vps27, a reduction in urease activity and alterations in melanin production is also produced [49]. In C. neoformans, the absence of cyclin Cln1 results in an increase in GXM deposition and an apparent increase in the EV release [57]. Intriguingly, the absence of laccase activity in the cells is accompanied by LAC1 downregulation [64], such as in C. neoformans vps27, suggesting a feasible trafficking of this enzyme to EVs and a connection between cyclin Cln1 and the non-conventional secretory pathway.
4.4. Lipid Homeostasis
Lipid asymmetry is important for cellular homeostasis, including regulation of membrane curvature and vesicle trafficking (Figure 2). Enzymes such as flippases, floppases and scramblases are essential for the translocation of phospholipids and their proper disposition [65,66]. Aminophospholipid translocases (APTs or APLTs) flip specifically phosphatidylserine (PS) and phosphatidylethanolamine (PE) from the external face to the inner side of the membrane [67]. In S. cerevisiae, five APTs have been described and characterised as components of the Golgi apparatus (Drs2p, Dnf1p, Dnf2p, Dnf3p and Neo1p); they are all involved in vesicular traffic [68]. Indeed, in C. neoformans, apt1 mutants show Golgi structures concentrated in the centre of the cell instead of the typical peripheral distribution, aberrant vacuole morphology and formation of gigantic multivesicular body (MVB)-like compartments [69]. Interestingly, the size of EVs secreted in this mutant is also altered. C. neoformans secretes two populations of EVs, from 10 to 150 nm and from 400 to 1000 nm in diameter. In apt1 mutants, the largest EVs are distributed in a narrower range of 400 to 600 nm. Moreover, defective intracellular GXM synthesis in mutant cells give rise to a significantly lower content of GXM polysaccharide in EVs, which results in an attenuated virulence and defective oxidative stress response in the host [61,69]. The absence of the scramblase Aim25 also results in disorganised membranes; the cells lack the typical vacuoles and show aberrant membranous structures [32]. In C. gattii, knockout mutation in Aim25 decreases the production of EVs. The EVs from the aim25 mutant are also larger than normal and their RNA content is altered. Physicochemical properties are also altered, resulting in a more efficient release of GXM from EVs to the capsule, thus increasing its dimensions. The data described above demonstrate the importance of phospholipid translocation in membrane organisation and EV secretion (Table 4).
It must be noted that not only the translocation of the phospholipids but also their synthesis is essential for the biogenesis of the EVs. Phosphatidylserine synthase (Cho1) and phosphatidylserine decarboxylases (Psd1 and Psd2) are involved in phosphatidylserine and phosphatidylethanolamine biosynthesis, respectively [70]. In C. albicans, a double mutant psd1/psd2 produces much larger EVs. In cho1 and psd1/psd2 mutants, the EV phosphatidylcholine and protein contents are distorted. Several EV-associated virulence factors are absent in this scenario, such as phospholipase Plb3 and adhesin Sim1 in cho1 mutant and proteinase Prd1 in the double mutant psd1/psd2 [29] (Table 4).
4.5. Other Players in EV Biogenesis
Other proteins also are implicated in EV biogenesis, involving mechanisms poorly understood. Decreased expression of a 14-3-3 protein, normally abundant in cryptococcal EVs, leads to reduced EV release [71]. Mutant phenotype also involves lighter and shorter capsule, lower EV GXM, protein content and activity of laccase and acid phosphatase [72]. Blocking of Hsp60 by monoclonal antibodies in H. capsulatum leads to fluctuations in EVs size, release and protein loading [43,73].
5. Functionality and Biological Implications of Fungal EVs
The functions of fungal EVs comprise a wide spectrum of responses to internal and external stimuli. These responses must be understood not only in the context of individual cells but also in the process of coordinating fungal activity in a community (Figure 3).
5.1. Protective and Defensive Structures: Capsule, Cell Wall and Matrix
The first study reporting EVs in a fungal experimental model (C. neoformans) demonstrated the presence of glucuronoxylomannan (GXM), the major component of the cryptococcal capsule. The GXM has been detected in the lumen of the vesicles found in the extracellular space and those attached to the cell wall. GXM is transferred in these vesicles from the cytoplasm to the cell wall. The vesicles are released from the cells and then taken up by the producers themselves and neighbouring cells, aiding in the capsule assembly and maintenance [10,32]. In agreement with this, enhanced EV release could be responsible for the increased GXM deposition that results in enlarged capsule phenotype in the C. neoformans cln1 mutant [57].
Cell wall-remodelling enzymes are a part of EV cargo in S. cerevisiae (Fks1 and Chs3) [44], H. capsulatum (Chi1) [16] and Aspergillus fumigatus (Fks1, Gel and Bgt) as well [74]. The EVs from P. brasiliensis also carry glycoconjugates containing α-galactosyl epitopes. These glycoconjugates are stored in intracellular vacuoles and might travel to the cell wall utilising the MVB pathway [17]. Thus, the available data indicate that EVs have a role in cell wall homeostasis. Indeed, mutating components of ESCRT machinery in S. cerevisiae reduces the release of EVs but increases their Fks1 and Chs3 levels and stimulates cell wall synthesis [44].
It has been proposed that the EVs might take part in matrix biogenesis, development and maintenance [39]. C. albicans growing in biofilm communities secretes EVs that carry proteins involved in extracellular matrix structure and biogenesis, specifically polysaccharide modification enzymes and polysaccharides (i.e., mannan and glucan).
5.2. Pathogenesis
Cell wall and capsule structures protect the cell from adverse environmental conditions and host immune agents; they are considered the primary pathogenic elements for most fungal species as they ensure their survival [44]. Thus, the participation of fungal EVs in the biogenesis and maintenance of the capsule and the cell wall may be considered a pathogenic mechanism itself. In S. cerevisiae, EVs from mutants in ESCRT machinery rescue the cells from antifungal treatment because of their activity in cell wall synthesis [44]. In C. neoformans [10] and C. gattii [21,32], the EV GXM-related capsule synthesis constitutes a protective rather than a virulent mechanism, allowing the yeast to survive in the cells of its usual environmental predator, the amoeba Acanthamoeba castellanii [75]. When the release of EVs is blocked in C. albicans forming biofilm-protected communities, a strong sensitivity to several antifungal drugs is observed as the matrix synthesis fails [39].
First reviews of fungal EV studies have classified them as virulence bags. The concept has its origin in early proteomic studies of EVs from C. neoformans and H. capsulatum [76], in which virulence factors such as chaperones (e.g., Hsp60 precursors, Hsp70 and Hsp30), catalase B and superoxide dismutase have been identified. The enzymes laccase, urease, phosphatase and catalase were shown to be active [13]. Since then, other molecules involved in the pathogenicity of other fungi species have been described, such as melanin in E. dermatitidis [20], GlcCer and sterols [10,17,19,27]. GlcCer plays an important role in fungal cell division, alkaline tolerance, hyphal formation and spore germination, being it is a key regulator of pathogenicity [28]. In the C. neoformans cln1 mutant, boosting in EV release is accompanied by an increase in their sterol content and strengthened virulence [57]. This strongly suggests an active role for the EVs in the virulence regulation (Table 1 and Table 2).
On a more physiological level, E. dermatitidis-derived melanised EVs may mediate the neurotoxicity associated with this neurotropic black yeast [20]. In Zymoseptoria tritici, a wheat pathogen, the EVs are involved in switching from a yeast-like form of growth to a filamentous mode, responsible for pathogenicity in plants [77]. In S. cerevisiae, both the PVs and are involved in prion transmission. Interestingly, both native and aggregated states of the fungal prion Sup35p can be EV-packaged, maintaining their infectivity [37,47]. Moreover, EVs from fungi pathogens infecting plants, as Fusarium oxysporum, are able to induce a phytotoxic response [36].
5.3. Immunomodulation
There has been a lot of interest in the ability of EVs to regulate the host immune system; many cases of immunogenic molecules found in the EVs have been described (Table 1 and Table 2). Some EV-associated molecules (e.g., S. brasiliensis [22]) can induce immunological responses and can become allergens, as reported for M. sympodialis [18,42]. P. brasiliensis secretes the immunogen alpha-linked galactopyranosyl (α-Gal) associated with EVs [17]. Lipids such as GlcCer [10,19,27], which exhibit a characteristic and specific structure in comparison with the form found in mammals and plants and ergosterol [10,17,19,27] are also immunomodulators [28]. Hgt1p, a high-affinity glucose transporter from C. albicans EVs, can also act as a complement factor H (FH) binding molecule, mimicking human cells and, thus, evading the immune response and resulting in increased virulence [46].
Several types of immune cells can be modulated by the yeast EVs. Some early studies have shown that C. neoformans-derived EVs can alter the functionality of murine macrophages in vitro, achieving cross-kingdom communication and modulation of host cell response during infection [78]. In another example of such interactions, P. brasiliensis EVs can induce M1 polarisation of murine macrophages [79]. This triggers the production of proinflammatory mediators (e.g., NO, IL-12, IL-6, TNF-α) and an increase in the fungicidal activity, helping the host to limit the infection. In contrast, H. capsulatum EVs inhibit phagocytosis in macrophages as well as intracellular yeast killing by these innate immunity cells, thus promoting survival of the fungus and persisting infection. This effect has been observed during a low-level immune response simulated in vitro, suggesting a bidirectional communication between the host and pathogen cells [73].
The activity of other immune cells, such as monocytes, can also be modulated by fungal EVs [42]. Dendritic cells (DCs) stimulated with S. brasiliensis EVs increase phagocytosis and cytokine production. However, in contrast to EVs from C. neoformans [78] and P. brasiliensis [79], this results in a rise in the fungal load. In vivo assays have confirmed that, effectively, the EVs enhance fungal burden and skin lesions in murine models [22]. Other evidence of immune system modulation has been obtained for the skin-colonising commensal yeast M. sympodialis. The EVs from this fungus (designated MalaEx) can activate keratinocytes by augmenting the ICAM-1 expression [48] and IL-4 and TNF-α responses in DCs [18].
The MalaEx might have a role in atopic eczema. The MalaEx EVs release digestive enzymes to break down sebaceous lipids on the host skin, producing irritant unsaturated free fatty acids. This activity can lead to various skin disorders associated with this lipid-dependent yeast genus [42]. Similarly, the EVs derived from Trichophyton interdigitale can induce proinflammatory cytokine release (e.g., NO) from macrophages and keratinocytes. These observations suggest that fungal EVs from sources other than the yeast cells can also have some pathogenic functions [80].
Interestingly, EVs from C. gattii play an active role in the division of labour of this pathogen. During infection by outbreak strains of C. gattii, the host reactive oxygen species induce tubular mitochondrial phenotype in some of the phagocyted fungal population. These altered population protect the normal cells, fostering their intracellular proliferation, leading to enhanced pathogenesis within the outbreak lineage [81]. EVs from virulent cells can diffuse over significant distances, become rapidly internalised by macrophages and then trigger increased rates of proliferation in cryptococci residing within the phagosome [21].
5.4. Host Modulation of Fungi Pathogenicity
Host can also modulate the functionality and pathogenicity of yeast EVs via the immune system. Exposing H. capsulatum to monoclonal antibodies significantly alters its EV characteristics, e.g., their size, protein loading and activity of some enzymes related to virulence, such as laccase, catalase and phosphatase. This can decrease the pathogen virulence and change the outcome of infection in mice [43]. However, pathogen modulation is not exclusive to mammals. Regente et al. have demonstrated that sunflower EVs contain defensive agents that promote morphological changes and cell death in the phytopathogenic fungus Sclerotinia sclerotiorum [82].
In this scenario, the EVs must be stable to be functionally successful. Ergosterol is a pivotal component of fungal EVs since serum albumin can bind to it, destabilising the EV membrane. In the course of infection, albumin can compromise the integrity of the fungi EVs, causing their rupture and the consequent release of potential immunomodulators [83].
5.5. Nutrition and Environmental Sensing
Proteomic studies of fungal periplasmic and extracellular vesicles have demonstrated they carry proteins related to nutrient exploitation, from glucose to phosphate to iron (Table 1), indicating that such vesicles might take part in environmental sensing. In S. cerevisiae, small PVs carrying the gluconeogenic enzymes FBPase, Pck1p, Mdh2 and Icl1p are secreted under conditions of glucose starvation and internalised in the presence of glucose [23,84]. Thus, the export of prion-like proteins in PVs is dependent on glucose availability [37]. Interestingly, the EVs from C. albicans carry Hgt1p, the high-affinity glucose transporter; this might be associated with glucose sensing in the extracellular environment and, in turn, with induction of Hgt1p synthesis and its transport to the plasma membrane [46].
Furthermore, the EVs change their composition depending on growth conditions, supporting that they might participate in nutrient sensing and adaptation to the medium. Some of the many examples of research in this field are the study of C. albicans in biofilm culture [39,40], C. gattii in solid media [32], P. fermentans in two liquid formulas [31] and more recently, an exhaustive study of H. capsulatum cultured under four different conditions [85].
6. EVs Isolation and Characterization Procedures
Environmental conditions affect the production of fungal PVs and EVs, which is species-, strain- and application-dependent. The variables to be considered are the temperature, timing, nutritional conditions, shaking of yeast cultures, liquid or solid media or biofilm format (summarised in Table 3).
For purification of EVs, most authors follow the gold standard established for mammals, based on differential centrifugation and concentration by ultracentrifugation. As the first step, the general protocol recommends low-speed centrifugation to remove the whole cells and small debris. This consists of first centrifugation at 4000× g for 15 min at 4 °C, followed by the collection of the supernatant and further centrifugation for 15–30 min at 15,000× g at 4 °C to remove or separate large vesicles and aggregates (Step 1). The supernatant is usually concentrated by ultrafiltration (20-fold concentration using a cut-off of 100 kDa) (Step 2). Sometimes, further serial centrifugations are performed, as in Step 1 and the supernatant filtered through filters with a variable pore size (from 0.2 to 0.8 µm) (Step 3). Finally, several serial ultracentrifugation steps (100,000× g for 1 h at 4 °C) are executed to precipitate and wash the EVs [Step 4]. All such centrifugation-based procedures found in the literature are modifications of the method used by Rodrigues et al., 2007, for the first-ever fungal EV purification. They are now thoroughly adapted to the specific needs.
If high-purity EVs are required, a density gradient (sucrose or iodixanol) [10,18] or size exclusion chromatography can be performed [86]. A noteworthy option for specific subpopulations management is the immunocapture method [10,18]. Generally, this is limited by the shortage of surface markers. However, a recent study has reported that Sur7 and Evp1, claudin-like proteins from the Sur7 family, might be useful as EV surface markers in fungi [40]. These methods have similar limitations for the fungal and mammalian EV sources; it is not possible to separate the heterogeneous EV populations from non-vesicular contaminants.
Characterisation of isolated EVs is usually conducted employing transmission electron microscopy (TEM), nanoparticle tracking analysis (NTA) and immunoblotting (WB). TEM is used to visualise the EVs, their spatial distribution and shape. The cryo-EM method gives excellent results since the cells and EVs retain their innate state and structure details. NTA quantifies the EVs in the sample and obtains their size distribution within the population, similarly to the dynamic light scattering technique (DLS). The WB allows protein detection by using specific antibodies. In addition, WB, other immunodetection methods can be used, such as immuno-gold labelling combined with electron microscopy and antibody-coupled bead flow cytometry analysis. These techniques can be combined with other methods mentioned above to widen the spectrum of variables that can be studied and obtain more reliable results [87,88].
7. Discussion
Fungi are of great interest in many different areas, from biological research to human, animal and plant health and various industrial fields. The yeasts have been widely used as the eukaryotic model because their cellular activities and mechanisms are common or closely related to the organisms placed higher on the evolutionary ladder. Recently, extracellular vesicles have emerged as a novel intercellular communication model. They have been discovered in mammals, but all kingdoms and species studied so far have shown similar cell–to–cell communication features. As discussed here, all fungi species, whether pathogenic (such as C. neoformans, H. capsulatum, P. brasiliensis, M. sympodialis, C. albicans, S. brasiliensis, C. gattii, E. dermatitis or F. oxysporum) or industrially relevant (e.g., S. cerevisiae and P.fermentans), secrete vesicles of this type (Figure 1). As expected, the EVs in the fungi are involved in the delivery of a wide variety of molecules to the extracellular milieu, such as proteins, nucleic acids, lipids and polysaccharides related to EV functionality (Table 1 and Table 2). Thus, in a natural environment, intercellular communication via the EVs may help these organisms to survive under cell wall-damaging conditions (such as saline stress), conduct nutritional sensing or maintain their virulence (Figure 3).
In the fungi, like in mammals, both conventional ER–GA–plasma membrane secretory axis and the endocytic secretory pathway are involved in the EV biogenesis (Table 4). In fact, some secretory components are highly conserved throughout the Eukaryota domain. In the yeast, the non-classical components such as Golgi reassembly and stacking proteins (GRASP) and autophagosome have also been associated with EV secretion. The mechanisms involved in the organisation and homeostasis of lipids have a key role. This highlights the underlying complexity of the regulation of EV secretion. In S. cerevisiae, some vesicles located in the periplasmic region have been described as PVs; however, they might have been the EVs on their way to the extracellular space (Figure 2). It is still unknown whether the EV secretion could be switched off and what would be the key factor in such a process. Clearly, there are many details of EV biogenesis, identification and secretion that must be investigated and understood.
8. Current Limitations
Although our understanding of fungal EV biology is advanced, we are still hampered by several limitations that need to be addressed. Some of them are common to all biological systems. For example, the physiological relevance of many studies might be questioned as the stoichiometry of EV release remains elusive. Experimental analysis normally does not reflect whether EVs are secreted constitutively under real-life conditions or not. Moreover, none of the currently available protocols can isolate pure EVs or even properly separate the subtypes. Distinguishing true vesicles from cell artefacts is often difficult and reliable EV markers are scarce. Further research is necessary to overcome these difficulties.
9. Future Perspectives
The EV research is strongly stimulated by the potential of these vesicles as diagnostic and therapeutic tools. As the EVs can be isolated from biological fluids, they make excellent targets for future liquid biopsy strategies. The components of fungal EVs could be used to diagnose both simple and complex infections or those associated with other conditions. The infection progression also could be monitored and used to make therapeutic decisions. Moreover, fungal EVs can modulate the immune system of the host and affect the disease progression. In this field, the utilisation of fungal EVs in biodelivery-based therapies and vaccine technologies looks very promising. Some bacterial EVs have been already evaluated as adjuvants in a meningococcal vaccine [89]. In the clinical field, fungal EVs could also be used to produce various vaccines (e.g., against mycoses).
Fungal EVs might also become an asset as biosensors. It is believed that in biofilm formation, the EVs can manage both intra- and inter-species communication; they are crucial in maintaining biofilm communities. They could act as radars of opportunist biofilm formation and response to antifungal treatments and in beer or wine fermentation. They might also be used for sensing contamination or monitoring industrial processes such as soil and water remediation.
Both the basic research into the EVs and their clinical applications would benefit from the development of reference materials (RMs) for EV study. Synthetics monodisperse or polydisperse nanoparticles are used as RMs for EV analysis employing the NTA, DLS or flow cytometry. However, RMs with biochemical and physicochemical resemblance to natural EVs would allow perfect calibration in nano-technique procedures; fungal EVs would be an excellent source of such components.
Nevertheless, we should not forget that basic research in this field is still needed to overcome the current limitations. Moreover, improving our understanding of the EV biogenesis and function will help to identify the EV-dependent mechanisms and use this knowledge in future applications. For example, analysing the role of fungal EVs in the division of labour in C. gattii might elucidate the infection mechanisms and help us tackle similar pathologies.
Author Contributions
Study concept and design: E.G. and J.M.F.-P. Acquisition of data: E.G., M.L.-J. and B.B. Analysis and interpretation of data: E.G. and J.M.F.-P. Drafting of the manuscript: E.G., M.L.-J. and B.B. Critical revision of the manuscript for important intellectual content: E.G. and J.M.F.-P. Study supervision and funding acquisition: J.M.F.-P. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the project RTI2018-094969-B-I00 (MCIU/AEI/FEDER, UE) and Excellence Severo Ochoa Grant (SEV-2016-0644) and by the European Union’s Horizon 2020 research (860303).
Acknowledgments
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| EVs | Extracellular vesicles |
| MVs | Microvesicles |
| PVs | Periplasmic vesicles |
| ILVs | Intraluminar vesicles |
| MVBs | Multivesicular bodies |
| EE | early endosomes |
| LE | late endosomes |
| VC | vacuole |
| CW | cell wall |
| CAP | capsule |
| PM | plasmamembrane |
| PP | periplasm |
| ER | endoplasmic reticulum |
| GA | Golgi apparatus |
| GRASP | Golgi Reassembly Stacking Protein |
| IVCs | Intracellular vesicle clusters |
| Nu | nucleus |
| GPI-AP | Glycosylphosphatidylinositol (GPI)-anchored proteins |
| AAA | ATPases Associated with diverse cellular Activities |
| GXM | Glucuronoxylomannan |
| sRNA | small RNA |
| tRNA | transfer RNA |
| rRNA | ribosomal RNA |
| miRNA | micro RNA |
| snRNA | small nuclear RNA |
| snoRNA | small nucleolar RNA |
| GSL | neutral glycosphingolipid |
| GlcCer | glucosylceramide |
| PE | Phosphatidylethanolamine |
| PS | Phosphatidylserine |
| PC | Phosphatidylcholine |
| PA | Phosphatidic acid |
| PI | Phosphatidylinositol |
| PG | Phosphatidylglycerol |
References
- Yáñez-Mó, M.; Siljander, P.R.; Andreu, Z.; Zavec, A.B.; Borràs, F.E.; Buzas, E.I.; Buzas, K.; Casal, E.; Cappello, F.; Carvalho, J.; et al. Biological properties of extracellular vesicles and their physiological functions. J. Extracell. Vesicles 2015, 4, 27066. [Google Scholar] [CrossRef] [Green Version]
- Kalra, H.; Adda, C.G.; Liem, M.; Ang, C.S.; Mechler, A.; Simpson, R.J.; Hulett, M.D.; Mathivanan, S. Comparative proteomics evaluation of plasma exosome isolation techniques and assessment of the stability of exosomes in normal human blood plasma. Proteomics 2013, 13, 3354–3364. [Google Scholar] [CrossRef]
- Keller, S.; Ridinger, J.; Rupp, A.K.; Janssen, J.W.; Altevogt, P. Body fluid derived exosomes as a novel template for clinical diagnostics. J. Transl. Med. 2011, 9, 86. [Google Scholar] [CrossRef] [Green Version]
- Tauro, B.J.; Greening, D.W.; Mathias, R.A.; Mathivanan, S.; Ji, H.; Simpson, R.J. Two distinct populations of exosomes are released from LIM1863 colon carcinoma cell-derived organoids. Mol. Cell. Proteom. 2013, 12, 587–598. [Google Scholar] [CrossRef] [Green Version]
- Pallet, N.; Sirois, I.; Bell, C.; Hanafi, L.A.; Hamelin, K.; Dieudé, M.; Rondeau, C.; Thibault, P.; Desjardins, M.; Hebert, M.J. A comprehensive characterization of membrane vesicles released by autophagic human endothelial cells. Proteomics 2013, 13, 1108–1120. [Google Scholar] [CrossRef]
- Samuel, M.; Bleackley, M.; Anderson, M.; Mathivanan, S. Extracellular vesicles including exosomes in cross kingdom regulation: A viewpoint from plant-fungal interactions. Front. Plant Sci. 2015, 6, 766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Toledo Martins, S.; Szwarc, P.; Goldenberg, S.; Alves, L.R. Extracellular Vesicles in Fungi: Composition and Functions. Curr. Top. Microbiol. Immunol. 2019, 422, 45–59. [Google Scholar] [CrossRef] [PubMed]
- Bleackley, M.R.; Dawson, C.S.; Anderson, M.A. Fungal Extracellular Vesicles with a Focus on Proteomic Analysis. Proteomics 2019, 19, e1800232. [Google Scholar] [CrossRef] [PubMed]
- Takeo, K.; Uesaka, I.; Uehira, K.; Nishiura, M. Fine structure of Cryptococcus neoformans grown in vivo as observed by freeze-etching. J. Bacteriol. 1973, 113, 1449–1454. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, M.L.; Nimrichter, L.; Oliveira, D.L.; Frases, S.; Miranda, K.; Zaragoza, O.; Alvarez, M.; Nakouzi, A.; Feldmesser, M.; Casadevall, A. Vesicular polysaccharide export in Cryptococcus neoformans is a eukaryotic solution to the problem of fungal trans-cell wall transport. Eukaryot. Cell 2007, 6, 48–59. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, D.L.; Nakayasu, E.S.; Joffe, L.S.; Guimarães, A.J.; Sobreira, T.J.; Nosanchuk, J.D.; Cordero, R.J.; Frases, S.; Casadevall, A.; Almeida, I.C.; et al. Characterization of yeast extracellular vesicles: Evidence for the participation of different pathways of cellular traffic in vesicle biogenesis. PLoS ONE 2010, 5, e11113. [Google Scholar] [CrossRef] [Green Version]
- Wolf, J.M.; Espadas-Moreno, J.; Luque-Garcia, J.L.; Casadevall, A. Interaction of Cryptococcus neoformans extracellular vesicles with the cell wall. Eukaryot. Cell 2014, 13, 1484–1493. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, M.L.; Nakayasu, E.S.; Oliveira, D.L.; Nimrichter, L.; Nosanchuk, J.D.; Almeida, I.C.; Casadevall, A. Extracellular vesicles produced by Cryptococcus neoformans contain protein components associated with virulence. Eukaryot. Cell 2008, 7, 58–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vallejo, M.C.; Nakayasu, E.S.; Matsuo, A.L.; Sobreira, T.J.; Longo, L.V.; Ganiko, L.; Almeida, I.C.; Puccia, R. Vesicle and vesicle-free extracellular proteome of Paracoccidioides brasiliensis: Comparative analysis with other pathogenic fungi. J. Proteome Res. 2012, 11, 1676–1685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodrigues, M.L.; Nakayasu, E.S.; Almeida, I.C.; Nimrichter, L. The impact of proteomics on the understanding of functions and biogenesis of fungal extracellular vesicles. J. Proteom. 2014, 97, 177–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Albuquerque, P.C.; Nakayasu, E.S.; Rodrigues, M.L.; Frases, S.; Casadevall, A.; Zancope-Oliveira, R.M.; Almeida, I.C.; Nosanchuk, J.D. Vesicular transport in Histoplasma capsulatum: An effective mechanism for trans-cell wall transfer of proteins and lipids in ascomycetes. Cell. Microbiol. 2008, 10, 1695–1710. [Google Scholar] [CrossRef] [Green Version]
- Vallejo, M.C.; Matsuo, A.L.; Ganiko, L.; Medeiros, L.C.; Miranda, K.; Silva, L.S.; Freymüller-Haapalainen, E.; Sinigaglia-Coimbra, R.; Almeida, I.C.; Puccia, R. The pathogenic fungus Paracoccidioides brasiliensis exports extracellular vesicles containing highly immunogenic α-Galactosyl epitopes. Eukaryot. Cell 2011, 10, 343–351. [Google Scholar] [CrossRef] [Green Version]
- Gehrmann, U.; Qazi, K.R.; Johansson, C.; Hultenby, K.; Karlsson, M.; Lundeberg, L.; Gabrielsson, S.; Scheynius, A. Nanovesicles from Malassezia sympodialis and host exosomes induce cytokine responses--novel mechanisms for host-microbe interactions in atopic eczema. PLoS ONE 2011, 6, e21480. [Google Scholar] [CrossRef] [Green Version]
- Vargas, G.; Rocha, J.D.; Oliveira, D.L.; Albuquerque, P.C.; Frases, S.; Santos, S.S.; Nosanchuk, J.D.; Gomes, A.M.; Medeiros, L.C.; Miranda, K.; et al. Compositional and immunobiological analyses of extracellular vesicles released by Candida albicans. Cell. Microbiol. 2015, 17, 389–407. [Google Scholar] [CrossRef]
- Lavrin, T.; Konte, T.; Kostanjšek, R.; Sitar, S.; Sepčič, K.; Prpar Mihevc, S.; Žagar, E.; Župunski, V.; Lenassi, M.; Rogelj, B.; et al. The Neurotropic Black Yeast Exophiala dermatitidis Induces Neurocytotoxicity in Neuroblastoma Cells and Progressive Cell Death. Cells 2020, 9, 963. [Google Scholar] [CrossRef] [Green Version]
- Bielska, E.; Sisquella, M.A.; Aldeieg, M.; Birch, C.; O’Donoghue, E.J.; May, R.C. Pathogen-derived extracellular vesicles mediate virulence in the fatal human pathogen Cryptococcus gattii. Nat. Commun. 2018, 9, 1556. [Google Scholar] [CrossRef] [Green Version]
- Ikeda, M.A.K.; de Almeida, J.R.F.; Jannuzzi, G.P.; Cronemberger-Andrade, A.; Torrecilhas, A.C.T.; Moretti, N.S.; da Cunha, J.P.C.; de Almeida, S.R.; Ferreira, K.S. Extracellular Vesicles from Sporothrix brasiliensis Are an Important Virulence Factor That Induce an Increase in Fungal Burden in Experimental Sporotrichosis. Front. Microbiol. 2018, 9, 2286. [Google Scholar] [CrossRef] [PubMed]
- Giardina, B.J.; Stein, K.; Chiang, H.L. The endocytosis gene END3 is essential for the glucose-induced rapid decline of small vesicles in the extracellular fraction in Saccharomyces cerevisiae. J. Extracell. Vesicles 2014, 3, 23497. [Google Scholar] [CrossRef] [Green Version]
- Mencher, A.; Morales, P.; Valero, E.; Tronchoni, J.; Patil, K.R.; Gonzalez, R. Proteomic characterization of extracellular vesicles produced by several wine yeast species. Microb. Biotechnol. 2020, 13, 1581–1596. [Google Scholar] [CrossRef] [PubMed]
- Wolf, J.M.; Casadevall, A. Challenges posed by extracellular vesicles from eukaryotic microbes. Curr. Opin. Microbiol. 2014, 22, 73–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peres da Silva, R.; Longo, L.G.V.; Cunha, J.P.C.D.; Sobreira, T.J.P.; Rodrigues, M.L.; Faoro, H.; Goldenberg, S.; Alves, L.R.; Puccia, R. Comparison of the RNA Content of Extracellular Vesicles Derived from. Cells 2019, 8, 765. [Google Scholar] [CrossRef] [Green Version]
- Vallejo, M.C.; Nakayasu, E.S.; Longo, L.V.; Ganiko, L.; Lopes, F.G.; Matsuo, A.L.; Almeida, I.C.; Puccia, R. Lipidomic analysis of extracellular vesicles from the pathogenic phase of Paracoccidioides brasiliensis. PLoS ONE 2012, 7, e39463. [Google Scholar] [CrossRef]
- Van der Grein, S.G.; Nolte-’t Hoen, E.N. “Small Talk” in the Innate Immune System via RNA-Containing Extracellular Vesicles. Front. Immunol. 2014, 5, 542. [Google Scholar] [CrossRef] [Green Version]
- Wolf, J.M.; Espadas, J.; Luque-Garcia, J.; Reynolds, T.; Casadevall, A. Lipid Biosynthetic Genes Affect Candida albicans Extracellular Vesicle Morphology, Cargo, and Immunostimulatory Properties. Eukaryot. Cell 2015, 14, 745–754. [Google Scholar] [CrossRef] [Green Version]
- Nicola, A.M.; Frases, S.; Casadevall, A. Lipophilic dye staining of Cryptococcus neoformans extracellular vesicles and capsule. Eukaryot. Cell 2009, 8, 1373–1380. [Google Scholar] [CrossRef] [Green Version]
- Leone, F.; Bellani, L.; Muccifora, S.; Giorgetti, L.; Bongioanni, P.; Simili, M.; Maserti, B.; Del Carratore, R. Analysis of extracellular vesicles produced in the biofilm by the dimorphic yeast Pichia fermentans. J. Cell. Physiol. 2018, 233, 2759–2767. [Google Scholar] [CrossRef]
- Reis, F.C.G.; Borges, B.S.; Jozefowicz, L.J.; Sena, B.A.G.; Garcia, A.W.A.; Medeiros, L.C.; Martins, S.T.; Honorato, L.; Schrank, A.; Vainstein, M.H.; et al. A Novel Protocol for the Isolation of Fungal Extracellular Vesicles Reveals the Participation of a Putative Scramblase in Polysaccharide Export and Capsule Construction in Cryptococcus gattii. mSphere 2019, 4, e00080-19. [Google Scholar] [CrossRef] [Green Version]
- Peres da Silva, R.; Puccia, R.; Rodrigues, M.L.; Oliveira, D.L.; Joffe, L.S.; César, G.V.; Nimrichter, L.; Goldenberg, S.; Alves, L.R. Extracellular vesicle-mediated export of fungal RNA. Sci. Rep. 2015, 5, 7763. [Google Scholar] [CrossRef] [Green Version]
- Rayner, S.; Bruhn, S.; Vallhov, H.; Andersson, A.; Billmyre, R.B.; Scheynius, A. Identification of small RNAs in extracellular vesicles from the commensal yeast Malassezia sympodialis. Sci. Rep. 2017, 7, 39742. [Google Scholar] [CrossRef] [Green Version]
- Peres da Silva, R.; Martins, S.T.; Rizzo, J.; Dos Reis, F.C.G.; Joffe, L.S.; Vainstein, M.; Kmetzsch, L.; Oliveira, D.L.; Puccia, R.; Goldenberg, S.; et al. Golgi Reassembly and Stacking Protein (GRASP) Participates in Vesicle-Mediated RNA Export in Cryptococcus Neoformans. Genes 2018, 9, 400. [Google Scholar] [CrossRef] [Green Version]
- Bleackley, M.R.; Samuel, M.; Garcia-Ceron, D.; McKenna, J.A.; Lowe, R.G.T.; Pathan, M.; Zhao, K.; Ang, C.S.; Mathivanan, S.; Anderson, M.A. Extracellular Vesicles from the Cotton Pathogen Fusarium oxysporum f. sp. vasinfectum Induce a Phytotoxic Response in Plants. Front. Plant Sci. 2019, 10, 1610. [Google Scholar] [CrossRef] [Green Version]
- Kabani, M.; Pilard, M.; Melki, R. Glucose availability dictates the export of the soluble and prion forms of Sup35p via periplasmic or extracellular vesicles. Mol. Microbiol. 2020, 114, 322–332. [Google Scholar] [CrossRef]
- Gil-Bona, A.; Llama-Palacios, A.; Parra, C.M.; Vivanco, F.; Nombela, C.; Monteoliva, L.; Gil, C. Proteomics unravels extracellular vesicles as carriers of classical cytoplasmic proteins in Candida albicans. J. Proteome Res. 2015, 14, 142–153. [Google Scholar] [CrossRef]
- Zarnowski, R.; Sanchez, H.; Covelli, A.S.; Dominguez, E.; Jaromin, A.; Bernhardt, J.; Mitchell, K.F.; Heiss, C.; Azadi, P.; Mitchell, A.; et al. Candida albicans biofilm-induced vesicles confer drug resistance through matrix biogenesis. PLoS Biol. 2018, 16, e2006872. [Google Scholar] [CrossRef]
- Dawson, C.S.; Garcia-Ceron, D.; Rajapaksha, H.; Faou, P.; Bleackley, M.R.; Anderson, M.A. Protein markers for Candida albicans EVs include claudin-like Sur7 family proteins. J. Extracell. Vesicles 2020, 9, 1750810. [Google Scholar] [CrossRef] [Green Version]
- Konečná, K.; Klimentová, J.; Benada, O.; Němečková, I.; Janďourek, O.; Jílek, P.; Vejsová, M. A comparative analysis of protein virulence factors released via extracellular vesicles in two Candida albicans strains cultivated in a nutrient-limited medium. Microb. Pathog. 2019, 136, 103666. [Google Scholar] [CrossRef]
- Johansson, H.J.; Vallhov, H.; Holm, T.; Gehrmann, U.; Andersson, A.; Johansson, C.; Blom, H.; Carroni, M.; Lehtiö, J.; Scheynius, A. Extracellular nanovesicles released from the commensal yeast Malassezia sympodialis are enriched in allergens and interact with cells in human skin. Sci. Rep. 2018, 8, 9182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matos Baltazar, L.; Nakayasu, E.S.; Sobreira, T.J.; Choi, H.; Casadevall, A.; Nimrichter, L.; Nosanchuk, J.D. Antibody Binding Alters the Characteristics and Contents of Extracellular Vesicles Released by Histoplasma capsulatum. mSphere 2016, 1, e00085-15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, K.; Bleackley, M.; Chisanga, D.; Gangoda, L.; Fonseka, P.; Liem, M.; Kalra, H.; Al Saffar, H.; Keerthikumar, S.; Ang, C.S.; et al. Extracellular vesicles secreted by Saccharomyces cerevisiae are involved in cell wall remodelling. Commun. Biol. 2019, 2, 305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stein, K.; Winters, C.; Chiang, H.L. Vps15p regulates the distribution of cup-shaped organelles containing the major eisosome protein Pil1p to the extracellular fraction required for endocytosis of extracellular vesicles carrying metabolic enzymes. Biol. Cell 2017, 109, 190–209. [Google Scholar] [CrossRef]
- Kenno, S.; Speth, C.; Rambach, G.; Binder, U.; Chatterjee, S.; Caramalho, R.; Haas, H.; Lass-Flörl, C.; Shaughnessy, J.; Ram, S.; et al. Candida albicans Factor H Binding Molecule Hgt1p—A Low Glucose-Induced Transmembrane Protein Is Trafficked to the Cell Wall and Impairs Phagocytosis and Killing by Human Neutrophils. Front. Microbiol. 2018, 9, 3319. [Google Scholar] [CrossRef] [PubMed]
- Kabani, M.; Melki, R. Sup35p in Its Soluble and Prion States Is Packaged inside Extracellular Vesicles. mBio 2015, 6, e01017-15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vallhov, H.; Johansson, C.; Veerman, R.E.; Scheynius, A. Extracellular Vesicles Released from the Skin Commensal Yeast Malassezia sympodialis Activate Human Primary Keratinocytes. Front. Cell. Infect. Microbiol. 2020, 10, 6. [Google Scholar] [CrossRef] [Green Version]
- Park, Y.D.; Chen, S.H.; Camacho, E.; Casadevall, A.; Williamson, P.R. Role of the ESCRT Pathway in Laccase Trafficking and Virulence of Cryptococcus neoformans. Infect. Immun. 2020, 88, e00954-19. [Google Scholar] [CrossRef]
- Walker, L.; Sood, P.; Lenardon, M.D.; Milne, G.; Olson, J.; Jensen, G.; Wolf, J.; Casadevall, A.; Adler-Moore, J.; Gow, N.A.R. The Viscoelastic Properties of the Fungal Cell Wall Allow Traffic of AmBisome as Intact Liposome Vesicles. mBio 2018, 9, e02383-17. [Google Scholar] [CrossRef] [Green Version]
- Tefsen, B.; Grijpstra, J.; Ordonez, S.; Lammers, M.; Van Die, I.; de Cock, H. Deletion of the CAP10 gene of Cryptococcus neoformans results in a pleiotropic phenotype with changes in expression of virulence factors. Res. Microbiol. 2014, 165, 399–410. [Google Scholar] [CrossRef]
- Latifkar, A.; Hur, Y.H.; Sanchez, J.C.; Cerione, R.A.; Antonyak, M.A. New insights into extracellular vesicle biogenesis and function. J. Cell Sci. 2019, 132, jcs222406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliveira, D.L.; Rizzo, J.; Joffe, L.S.; Godinho, R.M.; Rodrigues, M.L. Where do they come from and where do they go: Candidates for regulating extracellular vesicle formation in fungi. Int. J. Mol. Sci. 2013, 14, 9581–9603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panepinto, J.; Komperda, K.; Frases, S.; Park, Y.D.; Djordjevic, J.T.; Casadevall, A.; Williamson, P.R. Sec6-dependent sorting of fungal extracellular exosomes and laccase of Cryptococcus neoformans. Mol. Microbiol. 2009, 71, 1165–1176. [Google Scholar] [CrossRef]
- Kmetzsch, L.; Joffe, L.S.; Staats, C.C.; de Oliveira, D.L.; Fonseca, F.L.; Cordero, R.J.; Casadevall, A.; Nimrichter, L.; Schrank, A.; Vainstein, M.H.; et al. Role for Golgi reassembly and stacking protein (GRASP) in polysaccharide secretion and fungal virulence. Mol. Microbiol. 2011, 81, 206–218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rizzo, J.; Oliveira, D.L.; Joffe, L.S.; Hu, G.; Gazos-Lopes, F.; Fonseca, F.L.; Almeida, I.C.; Frases, S.; Kronstad, J.W.; Rodrigues, M.L. Role of the Apt1 protein in polysaccharide secretion by Cryptococcus neoformans. Eukaryot. Cell 2014, 13, 715–726. [Google Scholar] [CrossRef] [Green Version]
- García-Rodas, R.; Cordero, R.J.; Trevijano-Contador, N.; Janbon, G.; Moyrand, F.; Casadevall, A.; Zaragoza, O. Capsule growth in Cryptococcus neoformans is coordinated with cell cycle progression. mBio 2014, 5, e00945-14. [Google Scholar] [CrossRef] [Green Version]
- Shorter, J.; Watson, R.; Giannakou, M.E.; Clarke, M.; Warren, G.; Barr, F.A. GRASP55, a second mammalian GRASP protein involved in the stacking of Golgi cisternae in a cell-free system. EMBO J. 1999, 18, 4949–4960. [Google Scholar] [CrossRef] [Green Version]
- Kinseth, M.A.; Anjard, C.; Fuller, D.; Guizzunti, G.; Loomis, W.F.; Malhotra, V. The Golgi-associated protein GRASP is required for unconventional protein secretion during development. Cell 2007, 130, 524–534. [Google Scholar] [CrossRef] [Green Version]
- Duran, J.M.; Anjard, C.; Stefan, C.; Loomis, W.F.; Malhotra, V. Unconventional secretion of Acb1 is mediated by autophagosomes. J. Cell Biol. 2010, 188, 527–536. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, D.L.; Fonseca, F.L.; Zamith-Miranda, D.; Nimrichter, L.; Rodrigues, J.; Pereira, M.D.; Reuwsaat, J.C.; Schrank, A.; Staats, C.; Kmetzsch, L.; et al. The putative autophagy regulator Atg7 affects the physiology and pathogenic mechanisms of Cryptococcus neoformans. Future Microbiol. 2016, 11, 1405–1419. [Google Scholar] [CrossRef]
- Winters, C.M.; Hong-Brown, L.Q.; Chiang, H.L. Intracellular vesicle clusters are organelles that synthesize extracellular vesicle-associated cargo proteins in yeast. J. Biol. Chem. 2020, 295, 2650–2663. [Google Scholar] [CrossRef]
- Remec Pavlin, M.; Hurley, J.H. The ESCRTs—Converging on mechanism. J. Cell Sci. 2020, 133, jcs240333. [Google Scholar] [CrossRef]
- García-Rodas, R.; Trevijano-Contador, N.; Román, E.; Janbon, G.; Moyrand, F.; Pla, J.; Casadevall, A.; Zaragoza, O. Role of Cln1 during melanization of Cryptococcus neoformans. Front. Microbiol. 2015, 6, 798. [Google Scholar] [CrossRef] [Green Version]
- Contreras, F.X.; Sánchez-Magraner, L.; Alonso, A.; Goñi, F.M. Transbilayer (flip-flop) lipid motion and lipid scrambling in membranes. FEBS Lett. 2010, 584, 1779–1786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pomorski, T.G.; Menon, A.K. Lipid somersaults: Uncovering the mechanisms of protein-mediated lipid flipping. Prog. Lipid Res. 2016, 64, 69–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Auland, M.E.; Roufogalis, B.D.; Devaux, P.F.; Zachowski, A. Reconstitution of ATP-dependent aminophospholipid translocation in proteoliposomes. Proc. Natl. Acad. Sci. USA 1994, 91, 10938–10942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scherl, A.; Couté, Y.; Déon, C.; Callé, A.; Kindbeiter, K.; Sanchez, J.C.; Greco, A.; Hochstrasser, D.; Diaz, J.J. Functional proteomic analysis of human nucleolus. Mol. Biol. Cell 2002, 13, 4100–4109. [Google Scholar] [CrossRef] [PubMed]
- Rizzo, J.; Colombo, A.C.; Zamith-Miranda, D.; Silva, V.K.A.; Allegood, J.C.; Casadevall, A.; Del Poeta, M.; Nosanchuk, J.D.; Kronstad, J.W.; Rodrigues, M.L. The putative flippase Apt1 is required for intracellular membrane architecture and biosynthesis of polysaccharide and lipids in Cryptococcus neoformans. Biochim. Biophys. Acta 2018, 1865, 532–541. [Google Scholar] [CrossRef]
- Chen, Y.L.; Montedonico, A.E.; Kauffman, S.; Dunlap, J.R.; Menn, F.M.; Reynolds, T.B. Phosphatidylserine synthase and phosphatidylserine decarboxylase are essential for cell wall integrity and virulence in Candida albicans. Mol. Microbiol. 2010, 75, 1112–1132. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.H.; Wu, C.H.; Chang, Y.C.; Kwon-Chung, K.J.; Brown, R.J.; Jong, A. Cryptococcus neoformans-derived microvesicles enhance the pathogenesis of fungal brain infection. PLoS ONE 2012, 7, e48570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Chang, Y.C.; Wu, C.H.; Liu, J.; Kwon-Chung, K.J.; Huang, S.H.; Shimada, H.; Fante, R.; Fu, X.; Jong, A. The 14-3-3 Gene Function of Cryptococcus neoformans Is Required for its Growth and Virulence. J. Microbiol. Biotechnol. 2016, 26, 918–927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baltazar, L.M.; Zamith-Miranda, D.; Burnet, M.C.; Choi, H.; Nimrichter, L.; Nakayasu, E.S.; Nosanchuk, J.D. Concentration-dependent protein loading of extracellular vesicles released by Histoplasma capsulatum after antibody treatment and its modulatory action upon macrophages. Sci. Rep. 2018, 8, 8065. [Google Scholar] [CrossRef]
- Rizzo, J.; Chaze, T.; Miranda, K.; Roberson, R.W.; Gorgette, O.; Nimrichter, L.; Matondo, M.; Latgé, J.P.; Beauvais, A.; Rodrigues, M.L. Characterization of Extracellular Vesicles Produced by Aspergillus fumigatus Protoplasts. mSphere 2020, 5, e00476-20. [Google Scholar] [CrossRef] [PubMed]
- Rizzo, J.; Albuquerque, P.C.; Wolf, J.M.; Nascimento, R.; Pereira, M.D.; Nosanchuk, J.D.; Rodrigues, M.L. Analysis of multiple components involved in the interaction between Cryptococcus neoformans and Acanthamoeba castellanii. Fungal Biol. 2017, 121, 602–614. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, M.L.; Nimrichter, L.; Oliveira, D.L.; Nosanchuk, J.D.; Casadevall, A. Vesicular Trans-Cell Wall Transport in Fungi: A Mechanism for the Delivery of Virulence-Associated Macromolecules? Lipid Insights 2008, 2, 27–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hill, E.H.; Solomon, P.S. Extracellular vesicles from the apoplastic fungal wheat pathogen. Fungal Biol. Biotechnol. 2020, 7, 13. [Google Scholar] [CrossRef]
- Oliveira, D.L.; Freire-de-Lima, C.G.; Nosanchuk, J.D.; Casadevall, A.; Rodrigues, M.L.; Nimrichter, L. Extracellular vesicles from Cryptococcus neoformans modulate macrophage functions. Infect. Immun. 2010, 78, 1601–1609. [Google Scholar] [CrossRef] [Green Version]
- da Silva, T.A.; Roque-Barreira, M.C.; Casadevall, A.; Almeida, F. Extracellular vesicles from Paracoccidioides brasiliensis induced M1 polarization in vitro. Sci. Rep. 2016, 6, 35867. [Google Scholar] [CrossRef] [Green Version]
- Bitencourt, T.A.; Rezende, C.P.; Quaresemin, N.R.; Moreno, P.; Hatanaka, O.; Rossi, A.; Martinez-Rossi, N.M.; Almeida, F. Extracellular Vesicles from the Dermatophyte. Front. Immunol. 2018, 9, 2343. [Google Scholar] [CrossRef] [Green Version]
- Voelz, K.; Johnston, S.A.; Smith, L.M.; Hall, R.A.; Idnurm, A.; May, R.C. ’Division of labour’ in response to host oxidative burst drives a fatal Cryptococcus gattii outbreak. Nat. Commun. 2014, 5, 5194. [Google Scholar] [CrossRef] [PubMed]
- Regente, M.; Pinedo, M.; San Clemente, H.; Balliau, T.; Jamet, E.; de la Canal, L. Plant extracellular vesicles are incorporated by a fungal pathogen and inhibit its growth. J. Exp. Bot. 2017, 68, 5485–5495. [Google Scholar] [CrossRef] [PubMed]
- Wolf, J.M.; Rivera, J.; Casadevall, A. Serum albumin disrupts Cryptococcus neoformans and Bacillus anthracis extracellular vesicles. Cell. Microbiol. 2012, 14, 762–773. [Google Scholar] [CrossRef]
- Stein, K.; Chiang, H.L. Exocytosis and Endocytosis of Small Vesicles across the Plasma Membrane in Saccharomyces cerevisiae. Membranes 2014, 4, 608–629. [Google Scholar] [CrossRef] [Green Version]
- Cleare, L.G.; Zamith, D.; Heyman, H.M.; Couvillion, S.P.; Nimrichter, L.; Rodrigues, M.L.; Nakayasu, E.S.; Nosanchuk, J.D. Media matters! Alterations in the loading and release of Histoplasma capsulatum extracellular vesicles in response to different nutritional milieus. Cell. Microbiol. 2020, 22, e13217. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Ceron, D.; Dawson, C.S.; Faou, P.; Bleackley, M.R.; Anderson, M.A. Size-exclusion chromatography allows the isolation of EVs from the filamentous fungal plant pathogen Fusarium oxysporum f. sp. vasinfectum (Fov). Proteomics 2021, e2000240. [Google Scholar] [CrossRef]
- Rodrigues, M.L.; Oliveira, D.L.; Vargas, G.; Girard-Dias, W.; Franzen, A.J.; Frasés, S.; Miranda, K.; Nimrichter, L. Analysis of Yeast Extracellular Vesicles. Methods Mol. Biol. 2016, 1459, 175–190. [Google Scholar] [CrossRef]
- Liangsupree, T.; Multia, E.; Riekkola, M.L. Modern isolation and separation techniques for extracellular vesicles. J. Chromatogr. A 2021, 1636, 461773. [Google Scholar] [CrossRef]
- Choi, C.W.; Park, E.C.; Yun, S.H.; Lee, S.Y.; Kim, S.I.; Kim, G.H. Potential Usefulness of. J. Immunol. Res. 2017, 2017, 7931982. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Summary of the research into fungal EVs. Research activity for each species is displayed (A) over time or (B) according to the number of articles published. In both panels, human pathogen species are represented in grey and those with industrial impact in black.
Figure 1.
Summary of the research into fungal EVs. Research activity for each species is displayed (A) over time or (B) according to the number of articles published. In both panels, human pathogen species are represented in grey and those with industrial impact in black.

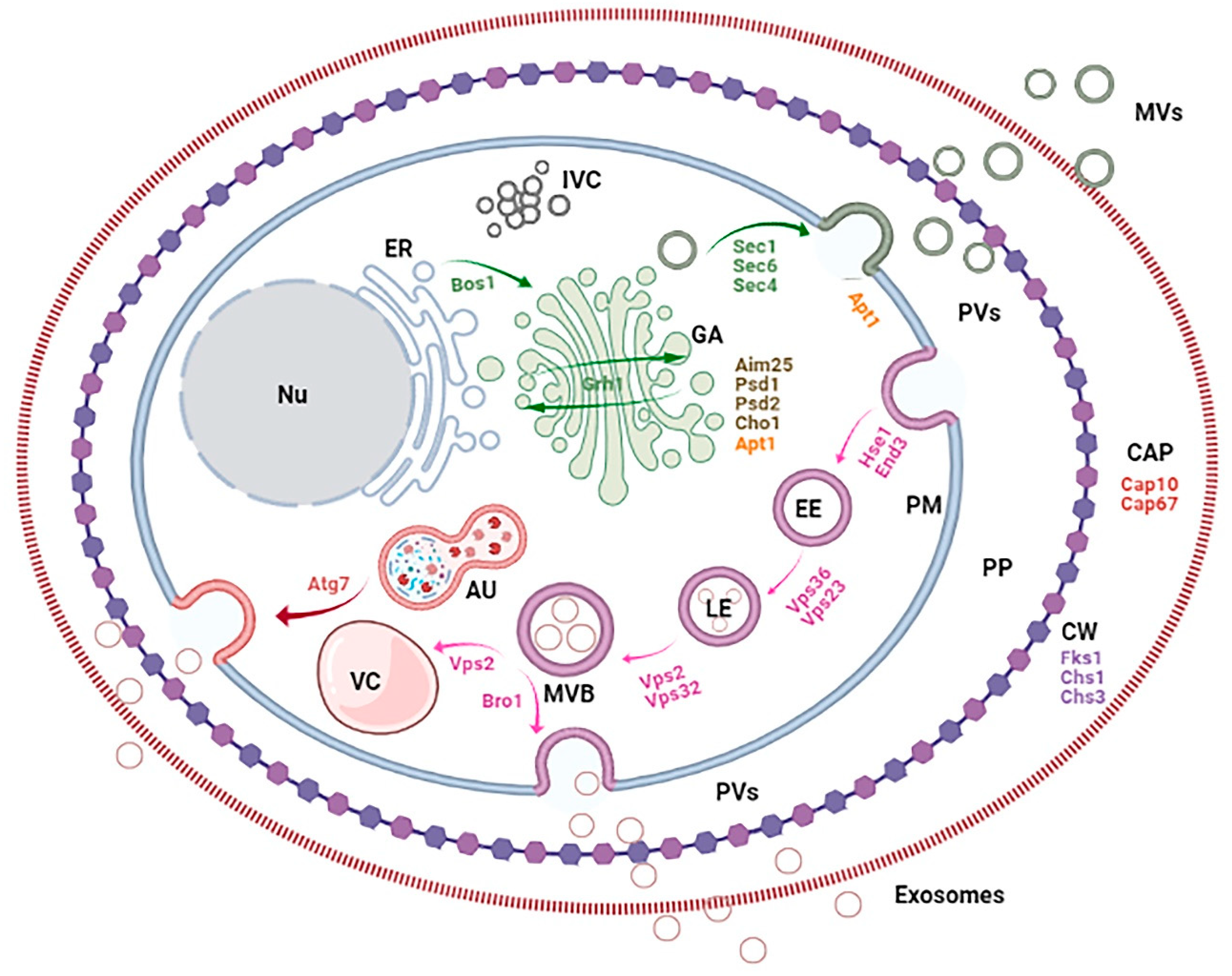
Figure 2.
Fungal cell compartments, pathways and genes involved in EV biogenesis. The figure shows the conventional secretory axis (ER: endoplasmic reticulum; GA: Golgi apparatus; IVC: Intracellular vesicle clusters) and endocytic secretory pathway (EE: early endosomes; LE: late endosomes; MVB: multivesicular bodies; VC: vacuole), autophagosome (AU), cell barriers (PM: plasma membrane; PP: periplasm; CW: cell wall; CAP: capsule), vesicles (PVs: periplasmic vesicles and EVs, including MVs: microvesicles and exosomes) and nucleus (Nu).
Figure 2.
Fungal cell compartments, pathways and genes involved in EV biogenesis. The figure shows the conventional secretory axis (ER: endoplasmic reticulum; GA: Golgi apparatus; IVC: Intracellular vesicle clusters) and endocytic secretory pathway (EE: early endosomes; LE: late endosomes; MVB: multivesicular bodies; VC: vacuole), autophagosome (AU), cell barriers (PM: plasma membrane; PP: periplasm; CW: cell wall; CAP: capsule), vesicles (PVs: periplasmic vesicles and EVs, including MVs: microvesicles and exosomes) and nucleus (Nu).

Figure 3.
Functions of fungal EVs. Descriptions of the functions (Capsule, cell wall and matrix synthesis; Environmental sensing; Virulence and Immunomodulation) are shown at the top of the figure and the corresponding responsible EV components at the bottom.
Figure 3.
Functions of fungal EVs. Descriptions of the functions (Capsule, cell wall and matrix synthesis; Environmental sensing; Virulence and Immunomodulation) are shown at the top of the figure and the corresponding responsible EV components at the bottom.


{kind=link}
{kind=link}
{kind=link}
Table 1.
Protein and nucleic acid composition of EVs in fungi.
| Specie | Nucleic Acids | Proteins | References |
|---|---|---|---|
| Cryptococcus neoformans | Categories
| [10,13,30,33,35] | |
| Histoplasma capsulatum | [16,43] | ||
| Paracoccidioides brasiliensis | [14,17,27,33] | ||
| Malassezia sympodialis | [18,34,42] | ||
| Candida albicans | [19,29,33,38,39,40,41,46] | ||
| Sporothrix brasiliensis | [22] | ||
| Cryptococcus gattii | [32] | ||
| Exophiala dermatitidis | [20] | ||
| Saccharomyces cerevisiae | [23,37,45] * [11,33,37,44,47] | ||
| Pichia fermetans | [31] | ||
| Fusarium oxysporum | [36] |
* periplasmic vesicles (PVs).
Table 2.
Kingdom specific component of fungal EVs.
| Specie | Carbohydrates | Lipids | Others | References |
|---|---|---|---|---|
| Cryptococcus neoformans | Polysaccharide
| Categories
| [10] | |
| Histoplasma capsulatum | [16,43] | |||
| Paracoccidioides brasiliensis | Polysaccharide
| [17,27] | ||
| Candida albicans | Polysaccharide(biofilm) | [19,29,39] | ||
| Cryptococcus gattii | Polysaccharide
| [21,32] | ||
| Malassezia sympodialis | Allergens | [18,42] | ||
| Sporothrix brasiliensis | Detected by antiserum | [22] | ||
| Exophiala dermatitidis | Melanin | [20] | ||
| Fusarium oxysporum | Detected by antiserum | [36] |
Table 3.
Morphology and profile of fungal EVs.
| Specie | Strains | EVs Physic Characteristic | Growth Conditions | References |
|---|---|---|---|---|
| Cryptococcus neoformans | ATCC 24067 (serotype D) [10] HEC3393 (serotype A) [10,33] H99 (serotype A) [10,13,30,33,35] B3501 (serotype D) [30,33] Cap67 (B3501 acapsular) [10,13,30,33] 2E-TUC (serotype D) [13] 2E-TU (2E-TUC LAC1-) [13] 24067 (serotype D) [30] | 1–2 L, minimal medium, 2–3 days (stationary phase), 30 °C, shaking. | [10,13,30,33,35] | |
| Histoplasma capsulatum | G217B (ATCC 26032) [16,43] |
| 0.5 L, Ham’s F-12/Glc medium, 2–7 days, 37 °C, shaking [16]. or 50 mL final, Ham’s F-12/Glc medium, refresh with 10 mL each 2 days for log phase maintenance (3 times, 7 days total), 37 °C, shaking [43]. | [16,43] |
| Paracoccidioides brasiliensis | Pb18 (group S1) [14,17,27,33] Pb3 (group PS2) [17,27] | 0.5 L, Ham’s F-12/Glc medium, 4 – 5 + 2 days, 36 °C, shaking. | [14,17,27,33] | |
| Malassezia sympodialis | ATCC 42132 [18,34,42] |
| 2 × 106–60 × 106 cells/mL, 0.3 L, RPMI-1640 or Dixon-MES medium, 2–3 days, 37 °C or 32 °C, shaking. | [18,34,42,48] |
| Candida albicans | 11 [19,33] ATCC 90028 [19,40,41] ATCC SC5314 [19,29,38] SN152 [39,46] Clinical isolated [41] DAY286 [40] ATCC 10231 [40] |
| Sabouraud medium, 48 h [19,33], or SD medium, 16 h, OD600 4 [38], 30 °C [19,38], or 0.1 L, YPD, 72 h, 37 °C [29], or OD600 0.1, Sabouraud medium, 22 h, 36 °C–37 °C [29], shaking in all cases. or RPMI 1640-MOPS (planktonic [39] and biofilm), 48–54 h, 37 °C [39,40]. or OD600 0.2, 0.1–0.3 L (1 L biofilm), YPD 0.1, 0.3 or 2% Glc, 20–48 h, 30 °C, shaking [40,46]. | [19,29,33,38,39,40,41,46] |
| Sporothrix brasiliensis | 5110 (ATCC MYA-4823) [22] |
| BHI medium, 6 days, 37 °C, shaking. | [22] |
| Cryptococcus gattii | R265 [32] |
| 0.5 mL YPD [21] or confluent YPD or Sabouraud’s medium (agar) [32], 24–72 h, 25 °C, shaking when liquid. | [32] |
| Exophiala dermatitidis | EXF-10123 [20] |
| 0.4 L, OD600 0.2, YNB medium, 15 h, 37 °C, shaking | [20] |
| Saccharomyces cerevisiae | BY4742 [23] BY4741 [44,45] RSY225 [11] BY4741 [11] RSY113 [11] SEY6210 [11,33] 74-D694 ± strong ([PSI+]S) or weak ([PSI+]W) Sup35p prion variants, or ± GFP-tagged Sup35p [37,47] | Periplasmic vesicles
| 40 mL YPKG 0.5% Glc 1–3 d + YPD 2% Glc, 30 min—15 h, 37 °C, shaking [23] *. or 1–2 L, OD600 0.1, YPDA 0.5% or 2% Glc, 3 d, 30 °C, shaking [37] **. or Sabouraud dextrose broth, 18–72 h, 25–37 °C, shaking [11] or 2 L, OD600 ~ 0.1, YPDA, 24 h, 30°C, shaking (final OD600 nm ~ 5–6) [47]. or OD600 0.2, YPD, 18 h, 30 °C, shaking[44]. | [23,37,45] * [11,33,37,44,47] |
| Pichia fermetans | Lodder [31] |
| 5–8 mL, YCU or YCM medium, 48 h, 28 °C, static. | [31] |
| Fusarium oxysporum | VCG01111 [36] |
| Liquid cultures were grown in half-strength potato dextrose broth (1/2 PDB), 72 h, 25 °C, shaking. | [36] |
Shape and distribution of EV population according to specie, strain and growth conditions. * periplasmic vesicles (PVs), ** both periplasmic (PVs) and extracellular vesicles (EVs).
Table 4.
Genes involved in biogenesis of fungal EVs.
| Gen | Mammalian Ortholog | Function | Phenotype in the Mutant | References |
|---|---|---|---|---|
| FKS1 | No cell-wall in mammals. | Catalytic subunit of 1,3-beta-D-glucan synthase. Cell wall remodeling. |
| [44] |
| CHS1 | No cell-wall in mammals. | Chitin synthase I; Cell wall remodeling and mitotic division septum formation. |
| [44] |
| CHS3 | No cell-wall in mammals. | Chitin synthase III. Cell wall remodeling. | [44] | |
| BOS1 | AtMEMB11/ AtMEMB12 | v-Snare. Localized to the ER membrane, necessary for vesicular transport from ER to GA. |
| [11] |
| SEC1 | STXBP1 | Localized to sites of secretion. Involved in docking and fusion of exocytic stimulating membrane fusion. |
| [11] |
| SEC4 | RAB8B | Regulation of polarized delivery of vesicles to the exocyst. |
| [11] |
| SEC6 | EXOC3 | Exocyst complex subunit. Mediation of polarized targeting and tethering of vesicles from GA to active sites of exocytosis at the plasma membrane. |
| [54] |
| VPS27 (HSE1) | STAM2 | ESCRT-0 complex. Sorting of ubiquitinated membrane proteins into intralumenal vesicles prior to vacuolar degradation and recycling. |
| [44,49] |
| VPS23 (STP22) | TSG101 | ESCRT-I complex. Sorting of ubiquitin- proteins into the endosome. |
| [11,44] |
| VPS36 | EAP45 | ESCRT-II complex. Interaction with ESCRT-I to sort ubiquitin- proteins into the endosome. | [44] | |
| VPS2 (DID4) | CHMP2A | ESCRT-III complex. Endocytosis, sorting of integral membrane proteins into lumenal vesicles of MVBs and vacuolar enzymes. |
| [44] |
| VPS32 (SNF7) | CHMP4A/B | ESCRT-III complex. Sorting from cytoplasm to endosomes and of transmembrane proteins into MVBs. |
| [44] |
| BRO1 | Alix/AIP1 | Cytoplasmic class E VPS factor. Coordination of deubiquitination in MVBs by recruiting Doa4p to endosomes. |
| [44] |
| GRH1(GRASP) | GRASP65 | GA trafficking and autophagosome formation. |
| [35,55] |
| END3 | EPS15 | Endocytosis, actin cytoskeletal organization and cell wall morphogenesis. Vacuolar degradation of plasma membrane proteins. |
| [23] * |
| DRS2(APT1) | ATP8B1 | Putative flippase. Fusion events at GA and plasma membrane. |
| [56] |
| AIM25 | PLSCR2; PLSCR4; PLSCR1 | Putative scramblase. Regulator of secretion and target for several antifungals. |
| [32] |
| CHO1 | PTDSS1/PTDSS2 | Phosphatidylserine synthase. Localized to the mitochondrial outer membrane. Phospholipid biosynthesis |
| [29] |
| PSD1 | PISD | Phosphatidylserine decarboxylase. Localized to the mitochondrial inner membrane. Phospholipid metabolism and inter-organelle trafficking of phosphatidylserine. | [29] | |
| PSD2 | PISD | Phosphatidylserine decarboxylase. Localized to GA, endosomes and vacuole. Regulation of phospholipids in compartments that will eventually give rise to the vacuole. | [29] | |
| CLN1 | CCNA2; CCNB1; CCNC; CCND1; CCNE1 | Cyclin-dependent protein kinase (CDK) regulatory subunit. Regulation of cell cycle. |
| [57] |
* periplasmic vesicles (PVs).
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Liebana-Jordan, M.; Brotons, B.; Falcon-Perez, J.M.; Gonzalez, E. Extracellular Vesicles in the Fungi Kingdom. Int. J. Mol. Sci. 2021, 22, 7221. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22137221
AMA Style
Liebana-Jordan M, Brotons B, Falcon-Perez JM, Gonzalez E. Extracellular Vesicles in the Fungi Kingdom. International Journal of Molecular Sciences. 2021; 22(13):7221. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22137221
Chicago/Turabian StyleLiebana-Jordan, Marc, Bruno Brotons, Juan Manuel Falcon-Perez, and Esperanza Gonzalez. 2021. "Extracellular Vesicles in the Fungi Kingdom" International Journal of Molecular Sciences 22, no. 13: 7221. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22137221
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.