
Citric Acid-Mediated Abiotic Stress Tolerance in Plants

 ,
,  ,
, 
 , , , ,
, , , ,  ,
,  , , and
, , and
Abstract
:1. Introduction
2. Effects of Abiotic Stress on Endogenous CA Levels
3. Exogenous CA for Mitigation of Abiotic Stress
3.1. Salinity Stress
3.2. Drought Stress
3.3. Temperature Stress
3.4. Alkalinity Stress
3.5. Heavy Metal Stress
4. Mechanisms of CA-Mediated Abiotic Stress Tolerance
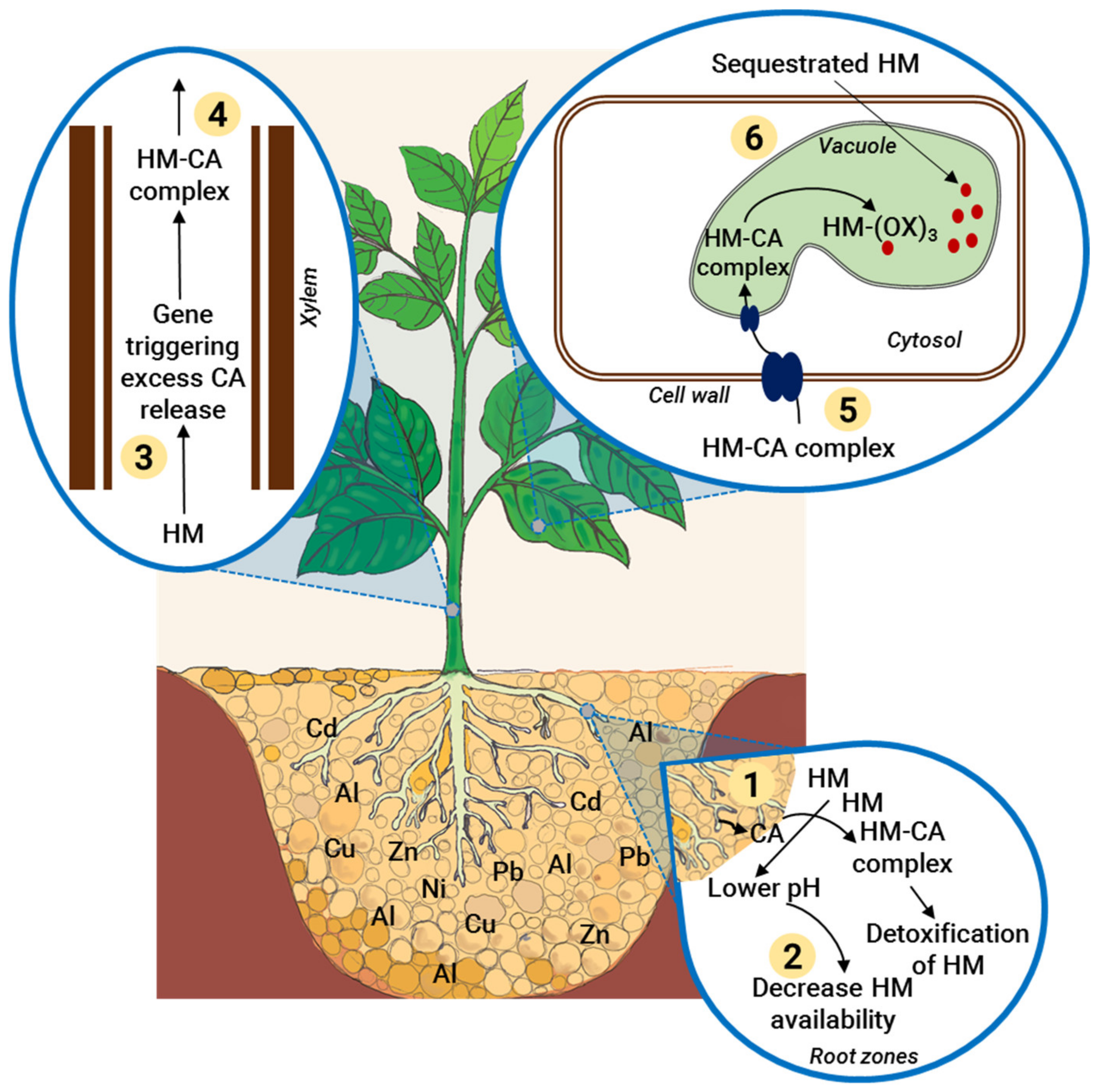
4.1. Regulation of Heavy Metal Uptake and Sequestration
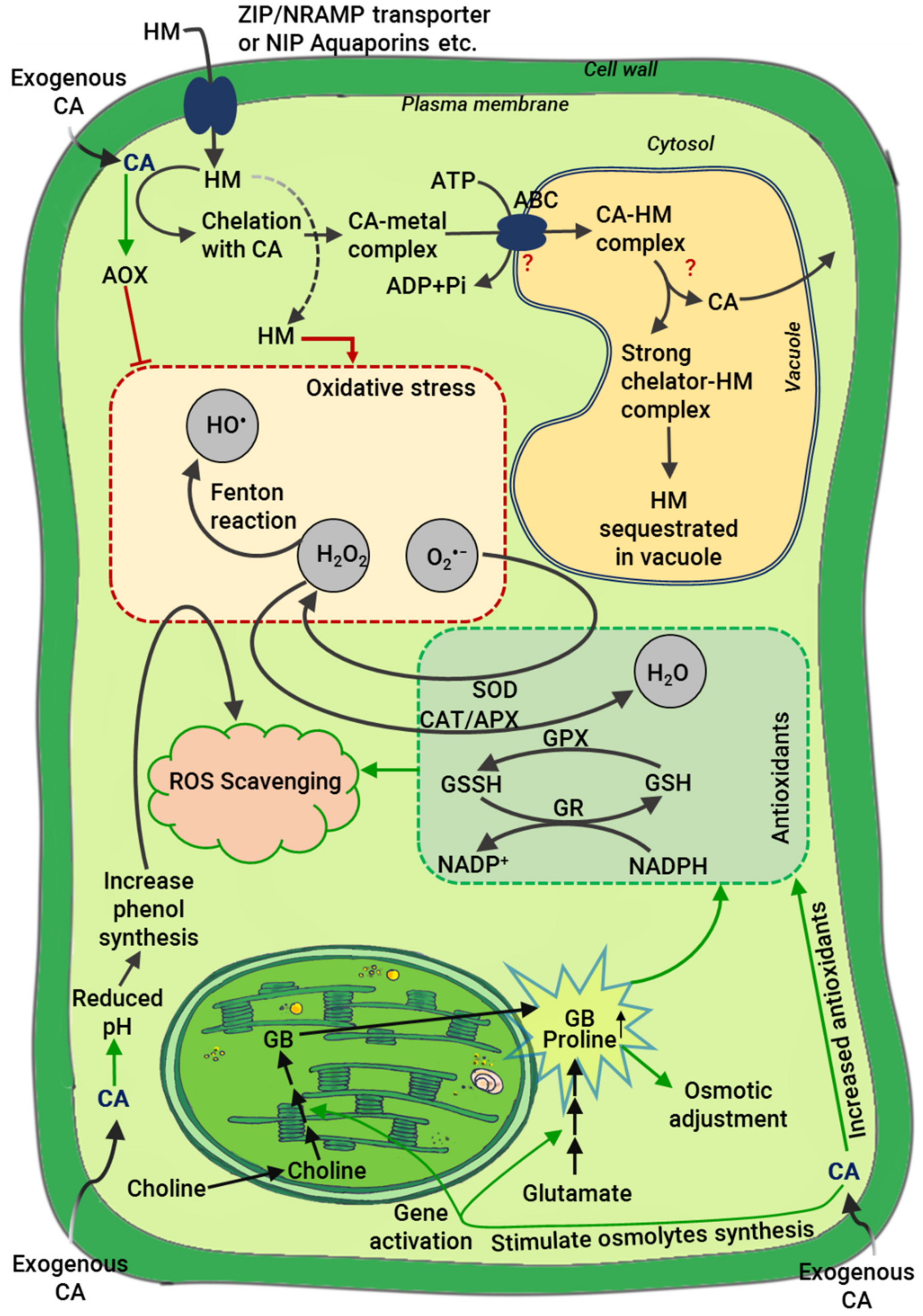
4.2. Regulation of ROS and Antioxidants
4.3. Regulation of Osmoregulators and Secondary Metabolites
5. Genetic Engineering for CA-Mediated Abiotic Stress Tolerance
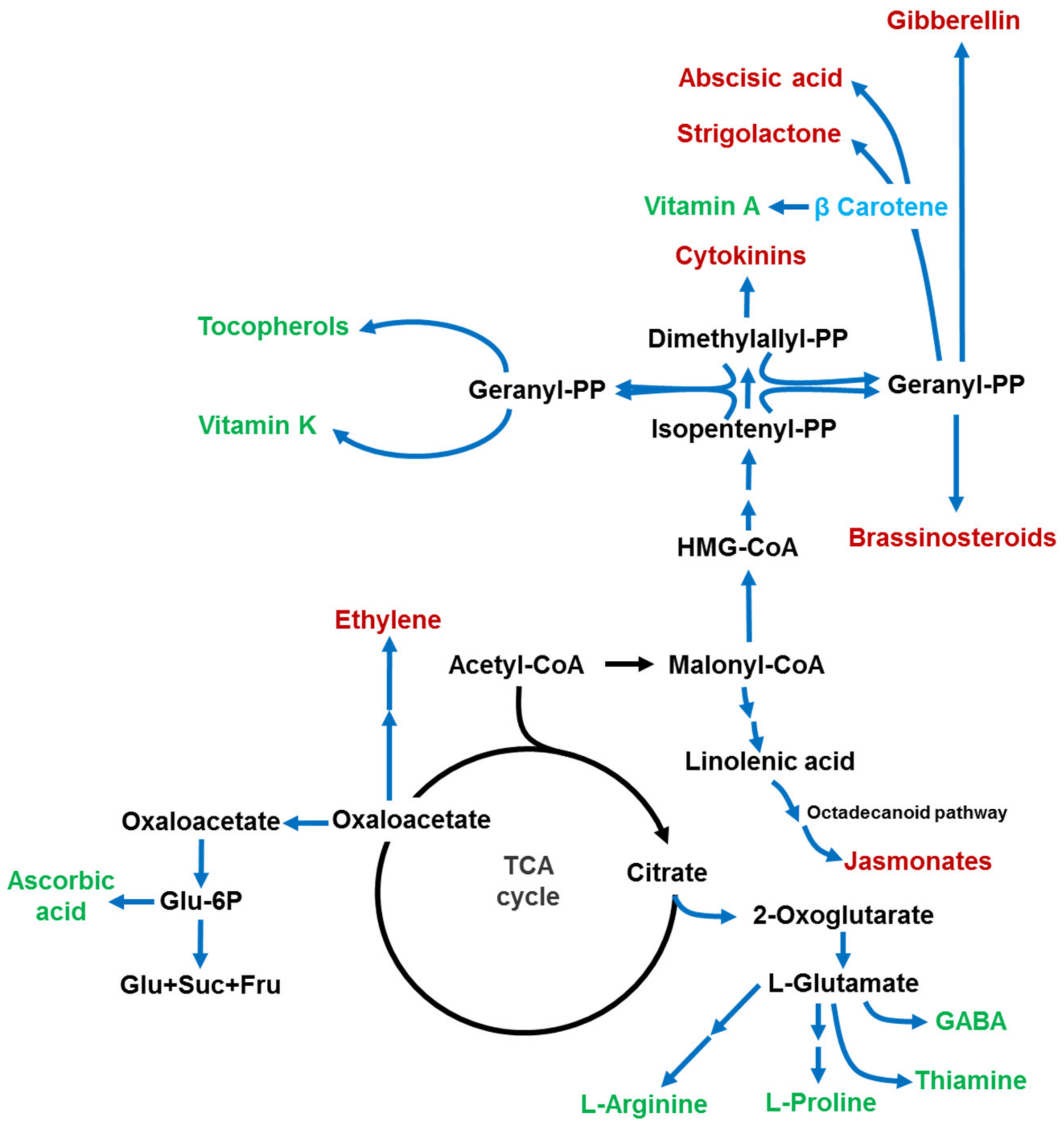
6. Metabolism of CA and Its Role in the Biosynthesis of Secondary Metabolites, Signaling Molecules, and Phytohormones
7. Conclusions and Future Perspective
- Enhances growth, photosynthesis, and many physio-biochemical parameters that promote crop productivity under abiotic stress conditions.
- Alleviates the abiotic stress-induced osmotic imbalance by increasing osmoregulators and protecting membranes from damage.
- Reduces the severity of oxidative stress by upregulating non-enzymatic and enzymatic antioxidants.
- Accelerates the HM stress tolerance of plants by chelating and sequestering HMs and improves HM phytoextraction from HM-polluted soils.
- Provides the substrate for a wide variety of metabolic pathways synthesizing stress protectant metabolites including phytohormones, amino acids, organic acids, and fatty acids.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yadav, S.; Modi, P.; Dave, A.; Vijapura, A.; Patel, D.; Patel, M. Effect of Abiotic Stress on Crops. In Sustainable Crop Production; Hasanuzzaman, M., Filho, M.C.M.T., Fujita, M., Nogueira, T.A.R., Eds.; IntechOpen Publishing, 2020; Available online: https://www.intechopen.com/books/sustainable-crop-production/effect-of-abiotic-stress-on-crops (accessed on 17 June 2020).
- Wang, Y.; Frei, M. Stressed Food–The Impact of Abiotic Environmental Stresses on Crop Quality. Agric. Ecosyst. Environ. 2011, 141, 271–286. [Google Scholar] [CrossRef]
- Dos Reis, S.P.; Lima, A.M.; De Souza, C.R.B. Recent Molecular Advances on Downstream Plant Responses to Abiotic Stress. Int. J. Mol. Sci. 2012, 13, 8628–8647. [Google Scholar] [CrossRef]
- Tuteja, N.; Sopory, S.K. Chemical Signaling under Abiotic Stress Environment in Plants. Plant Signal. Behav. 2008, 3, 525–536. [Google Scholar] [CrossRef] [Green Version]
- Gill, S.S.; Tuteja, N. Reactive Oxygen Species and Antioxidant Machinery in Abiotic Stress Tolerance in Crop Plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Trejo-Tellez, L.; Gomez-Merino, F.; Schmitt, J. Citric Acid: Biosynthesis, Properties and Applications on Higher Plants; DA Vargas and JV Medina Nova Science Publishers, Inc.: New York, NY, USA, 2012; pp. 43–70. [Google Scholar]
- Khatun, M.R.; Mukta, R.H.; Islam, M.A.; Huda, A.N. Insight into Citric Acid-Induced Chromium Detoxification in Rice (Oryza Sativa. L). Int. J. Phytoremediat. 2019, 21, 1234–1240. [Google Scholar] [CrossRef]
- Hussain, S.B.; Shi, C.-Y.; Guo, L.-X.; Kamran, H.M.; Sadka, A.; Liu, Y.-Z. Recent Advances in the Regulation of Citric Acid Metabolism in Citrus Fruit. Crit. Rev. Plant Sci. 2017, 36, 241–256. [Google Scholar] [CrossRef]
- Guo, L.-X.; Shi, C.-Y.; Liu, X.; Ning, D.-Y.; Jing, L.-F.; Yang, H.; Liu, Y.-Z. Citrate Accumulation-Related Gene Expression and/or Enzyme Activity Analysis Combined with Metabolomics Provide a Novel Insight for an Orange Mutant. Sci. Rep. 2016, 6, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Etienne, A.; Génard, M.; Lobit, P.; Mbeguié-A-Mbéguié, D.; Bugaud, C. What Controls Fleshy Fruit Acidity? A Review of Malate and Citrate Accumulation in Fruit Cells. J. Exp. Bot. 2013, 64, 1451–1469. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Bucio, J.; de la Vega, O.M.; Guevara-Garcia, A.; Herrera-Estrella, L. Enhanced Phosphorus Uptake in Transgenic Tobacco Plants That Overproduce Citrate. Nat. Biotechnol. 2000, 18, 450–453. [Google Scholar] [CrossRef]
- Eidyan, B.; Hadavi, E.; Moalemi, N. Pre-Harvest Foliar Application of Iron Sulfate and Citric Acid Combined with Urea Fertigation Affects Growth and Vase Life of Tuberose (Polianthes Tuberosa L.) ‘Por-Par’. Hortic. Environ. Biotechnol. 2014, 55, 9–13. [Google Scholar] [CrossRef]
- Darandeh, N.; Hadavi, E. Effect of Pre-Harvest Foliar Application of Citric Acid and Malic Acid on Chlorophyll Content and Post-Harvest Vase Life of Lilium Cv. Brunello. Front. Plant Sci. 2012, 2, 106. [Google Scholar] [CrossRef] [Green Version]
- El-Tohamy, W.; El-Abagy, H.; Badr, M.; Gruda, N. Drought Tolerance and Water Status of Bean Plants (Phaseolus Vulgaris L.) as Affected by Citric Acid Application. J. Appl. Bot. Food Qual. 2013, 86. [Google Scholar] [CrossRef]
- Gebaly, S.G.; Ahmed, F.M.; Namich, A.A. Effect of Spraying Some Organic, Amino Acids and Potassium Citrate on Alleviation of Drought Stress in Cotton Plant. J. Plant Prod. 2013, 4, 1369–1381. [Google Scholar] [CrossRef]
- Ehsan, S.; Ali, S.; Noureen, S.; Mahmood, K.; Farid, M.; Ishaque, W.; Shakoor, M.B.; Rizwan, M. Citric Acid Assisted Phytoremediation of Cadmium by Brassica Napus L. Ecotoxicol. Environ. Saf. 2014, 106, 164–172. [Google Scholar] [CrossRef]
- Hu, L.; Zhang, Z.; Xiang, Z.; Yang, Z. Exogenous Application of Citric Acid Ameliorates the Adverse Effect of Heat Stress in Tall Fescue (Lolium Arundinaceum). Front. Plant Sci. 2016, 7, 179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Beltagi, H.S.; Ahmed, S.H.; Namich, A.A.M.; Abdel-Sattar, R.R. Effect of Salicylic Acid and Potassium Citrate on Cotton Plant under Salt Stress. Fresen. Environ. Bull. 2017, 26, 1091–1100. [Google Scholar]
- Shi, D.; Sheng, Y. Effect of Various Salt–Alkaline Mixed Stress Conditions on Sunflower Seedlings and Analysis of Their Stress Factors. Environ. Exp. Bot. 2005, 54, 8–21. [Google Scholar] [CrossRef]
- Abbas, G.; Saqib, M.; Akhtar, J.; Murtaza, G.; Shahid, M. Effect of Salinity on Rhizosphere Acidification and Antioxidant Activity of Two Acacia Species. Can. J. For. Res. 2015, 45, 124–129. [Google Scholar] [CrossRef]
- Kang, S.-M.; Shahzad, R.; Bilal, S.; Khan, A.L.; Park, Y.-G.; Lee, K.-E.; Asaf, S.; Khan, M.A.; Lee, I.-J. Indole-3-Acetic-Acid and ACC Deaminase Producing Leclercia Adecarboxylata MO1 Improves Solanum Lycopersicum L. Growth and Salinity Stress Tolerance by Endogenous Secondary Metabolites Regulation. BMC Microbiol. 2019, 19, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Mickky, B.M.; Abbas, M.A.; Sameh, N.M. Morpho-Physiological Status of Fenugreek Seedlings under NaCl Stress. J. King Saud Univ. Sci. 2019, 31, 1276–1282. [Google Scholar] [CrossRef]
- Timpa, J.D.; Burke, J.J.; Quisenberry, J.E.; Wendt, C.W. Effects of Water Stress on the Organic Acid and Carbohydrate Compositions of Cotton Plants. Plant Physiol. 1986, 82, 724–728. [Google Scholar] [CrossRef] [Green Version]
- Franco, A.; Ball, E.; Lüttge, U. Differential Effects of Drought and Light Levels on Accumulation of Citric and Malic Acids during CAM in Clusia. Plant Cell Environ. 1992, 15, 821–829. [Google Scholar] [CrossRef]
- Herppich, W.; Peckmann, K. Responses of Gas Exchange, Photosynthesis, Nocturnal Acid Accumulation and Water Relations of Aptenia Cordifolia to Short-Term Drought and Rewatering. J. Plant Physiol. 1997, 150, 467–474. [Google Scholar] [CrossRef]
- Bethke, P.C.; Sabba, R.; Bussan, A.J. Tuber Water and Pressure Potentials Decrease and Sucrose Contents Increase in Response to Moderate Drought and Heat Stress. Am. J. Potato Res. 2009, 86, 519–532. [Google Scholar] [CrossRef]
- Nahar, K.; Ullah, S.M. Drought Stress Effects on Plant Water Relations, Growth, Fruit Quality and Osmotic Adjustment of Tomato (Solanum Lycopersicum) under Subtropical Condition. Asian J. Agric. Hortic. Res. 2018, 1–14. [Google Scholar] [CrossRef]
- Du, H.; Wang, Z.; Yu, W.; Liu, Y.; Huang, B. Differential Metabolic Responses of Perennial Grass Cynodon Transvaalensis× Cynodon Dactylon (C4) and Poa Pratensis (C3) to Heat Stress. Physiol. Plant. 2011, 141, 251–264. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Du, H.; Xu, M.; Huang, B. Metabolic Responses to Heat Stress under Elevated Atmospheric CO2 Concentration in a Cool-Season Grass Species. J. Am. Soc. Hortic. Sci. 2012, 137, 221–228. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Zhu, Y.; Ge, Q.; Li, Y.; Sun, J.; Zhang, Y.; Liu, X. Comparative Physiological Responses of Solanum Nigrum and Solanum Torvum to Cadmium Stress. New Phytol. 2012, 196, 125–138. [Google Scholar] [CrossRef]
- Mnasri, M.; Ghabriche, R.; Fourati, E.; Zaier, H.; Sabally, K.; Barrington, S.; Lutts, S.; Abdelly, C.; Ghnaya, T. Cd and Ni Transport and Accumulation in the Halophyte Sesuvium Portulacastrum: Implication of Organic Acids in These Processes. Front. Plant Sci. 2015, 6, 156. [Google Scholar] [CrossRef] [Green Version]
- Pietrini, F.; Iori, V.; Cheremisina, A.; Shevyakova, N.I.; Radyukina, N.; Kuznetsov, V.V.; Zacchini, M. Evaluation of Nickel Tolerance in Amaranthus Paniculatus L. Plants by Measuring Photosynthesis, Oxidative Status, Antioxidative Response and Metal-Binding Molecule Content. Environ. Sci. Pollut. Res. 2015, 22, 482–494. [Google Scholar] [CrossRef] [PubMed]
- Kaur, R.; Yadav, P.; Thukral, A.K.; Sharma, A.; Bhardwaj, R.; Alyemeni, M.N.; Wijaya, L.; Ahmad, P. Castasterone and Citric Acid Supplementation Alleviates Cadmium Toxicity by Modifying Antioxidants and Organic Acids in Brassica Juncea. J. Plant Growth Regul. 2018, 37, 286–299. [Google Scholar] [CrossRef]
- Zeng, F.; Chen, S.; Miao, Y.; Wu, F.; Zhang, G. Changes of Organic Acid Exudation and Rhizosphere PH in Rice Plants under Chromium Stress. Environ. Pollut. 2008, 155, 284–289. [Google Scholar] [CrossRef]
- Ma, J.F.; Zheng, S.J.; Matsumoto, H. Specific Secretion of Citric Acid Induced by Al Stress in Cassia Tora L. Plant Cell Physiol. 1997, 38, 1019–1025. [Google Scholar] [CrossRef]
- Ishikawa, S.; Wagatsuma, T.; Sasaki, R.; Ofei-Manu, P. Comparison of the Amount of Citric and Malic Acids in Al Media of Seven Plant Species and Two Cultivars Each in Five Plant Species. Soil Sci. Plant Nutr. 2000, 46, 751–758. [Google Scholar] [CrossRef]
- Li, X.F.; Ma, J.F.; Matsumoto, H. Pattern of Aluminum-Induced Secretion of Organic Acids Differs between Rye and Wheat. Plant Physiol. 2000, 123, 1537–1544. [Google Scholar] [CrossRef] [Green Version]
- Li, X.F.; Ma, J.F.; Matsumoto, H. Aluminum-Induced Secretion of Both Citrate and Malate in Rye. Plant Soil 2002, 242, 235–243. [Google Scholar]
- Kidd, P.; Llugany, M.; Poschenrieder, C.; Gunse, B.; Barcelo, J. The Role of Root Exudates in Aluminium Resistance and Silicon-induced Amelioration of Aluminium Toxicity in Three Varieties of Maize (Zea Mays L.). J. Exp. Bot. 2001, 52, 1339–1352. [Google Scholar] [PubMed]
- Farid, M.; Ali, S.; Saeed, R.; Rizwan, M.; Bukhari, S.A.H.; Abbasi, G.H.; Hussain, A.; Ali, B.; Zamir, M.S.I.; Ahmad, I. Combined Application of Citric Acid and 5-Aminolevulinic Acid Improved Biomass, Photosynthesis and Gas Exchange Attributes of Sunflower (Helianthus Annuus L.) Grown on Chromium Contaminated Soil. Int. J. Phytoremediat. 2019, 21, 760–767. [Google Scholar] [CrossRef] [PubMed]
- Tahjib-Ul-Arif, M.; Al Mamun Sohag, A.; Mostofa, M.G.; Polash, M.A.S.; Mahamud, A.S.U.; Afrin, S.; Hossain, A.; Hossain, M.A.; Murata, Y.; Phan Tran, L. Comparative Effects of Ascobin and Glutathione on Copper Homeostasis and Oxidative Stress Metabolism in Mitigation of Copper Toxicity in Rice. Plant Biol. 2020. [Google Scholar] [CrossRef]
- Zanotti, R.F.; Lopes, J.C.; Motta, L.B.; de Freitas, A.R.; Mengarda, L.H.G. Tolerance Induction to Saline Stress in Papaya Seeds Treated with Potassium Nitrate and Sildenafil Citrate. Semin. Ciências Agrárias 2013, 1, 3669–3673. [Google Scholar] [CrossRef]
- El-Hawary, M.; Nashed, M.E. Effect of Foliar Application by Some Antioxidants on Growth and Productivity of Maize under Saline Soil Conditions. J. Plant Prod. 2019, 10, 93–99. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, A.; Talaat, I.; Khalid, K. Citric Acid Affects Melissa Officinalis L. Essential Oil under Saline Soil. Asian J. Crop. Sci. 2017, 9, 40–49. [Google Scholar] [CrossRef] [Green Version]
- Abdellatif, Y.; Ibrahim, M. Non-Enzymatic Anti-Oxidants Potential in Enhancing Hibiscus Sabdariffa L. Tolerance to Oxidative Stress. Int. J. Bot. 2018, 14, 43–58. [Google Scholar]
- Sun, Y.-L.; Hong, S.-K. Effects of Citric Acid as an Important Component of the Responses to Saline and Alkaline Stress in the Halophyte Leymus Chinensis (Trin.). Plant Growth Regul. 2011, 64, 129–139. [Google Scholar] [CrossRef]
- Ahmed, S.; Abdel-Razek, M.; Hafez, W.; Aziz, A.E. Environmental Impacts of Some Organic Extracts on Sugar Beet Yield under Saline-Sodic Soil Conditions. J. Soil Sci. Agric. Eng. 2017, 8, 821–827. [Google Scholar] [CrossRef]
- Levi, A.; Paterson, A.H.; Cakmak, I.; Saranga, Y. Metabolite and Mineral Analyses of Cotton Near-isogenic Lines Introgressed with QTLs for Productivity and Drought-related Traits. Physiol. Plant. 2011, 141, 265–275. [Google Scholar] [CrossRef] [PubMed]
- Miyazawa, K. Drought Stress Alleviation of Cabbage Seedlings by Citric Acid Application. In Proceedings of the XXIX International Horticultural Congress on Horticulture: Sustaining Lives, Livelihoods and Landscapes (IHC2014), Brisbane, QLD, Australia, 17 August 2014. [Google Scholar]
- El-Desouky, S.; Ismaeil, F.; Wanas, A.; Fathy, E.; AbdEl-All, M.; Abd, M. Effect of Yeast Extract, Amino Acids and Citric Acid on Physioanatomical Aspects and Productivity of Tomato Plants Grown in Late Summer Season. Minufiya J. Agric. Res 2011, 36, 859–884. [Google Scholar]
- Zhang, L.; Livingstone, J.; Tarui, Y.; Hirasawa, E. Effects of Citric Acid, Sucrose, and Proton Concentration in Suppressing Defoliation in Hibiscus Plants Grown under Low-Illumination Conditions. HortTechnology 2009, 19, 305–308. [Google Scholar] [CrossRef]
- Gong, F.; Fan, W. Effects of Exogenous Citric Acids on Nutrient Activation of Calcareous Yellow Soil and Promotion Effects of Nutrient Absorption and Growth of Rosa Roxburghii Seedlings. Sci. Agric. Sin. 2018, 51, 2164–2177. [Google Scholar]
- Al Mahmud, J.; Hasanuzzaman, M.; Nahar, K.; Bhuyan, M.B.; Fujita, M. Insights into Citric Acid-Induced Cadmium Tolerance and Phytoremediation in Brassica Juncea L.: Coordinated Functions of Metal Chelation, Antioxidant Defense and Glyoxalase Systems. Ecotoxicol. Environ. Saf. 2018, 147, 990–1001. [Google Scholar] [CrossRef]
- Song, J.; Markewitz, D.; Wu, S.; Sang, Y.; Duan, C.; Cui, X. Exogenous Oxalic Acid and Citric Acid Improve Lead (Pb) Tolerance of Larix Olgensis A. Henry Seedlings. Forests 2018, 9, 510. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.-C.; Zhang, S.-L.; Wu, K.-J.; Li, R.; He, X.-R.; He, D.-N.; Huang, C.; Wei, H. The Effects of Exogenous Organic Acids on the Growth, Photosynthesis and Cellular Ultrastructure of Salix Variegata Franch. Under Cd Stress. Ecotoxicol. Environ. Saf. 2020, 187, 109790. [Google Scholar] [CrossRef]
- Zaheer, I.E.; Ali, S.; Rizwan, M.; Farid, M.; Shakoor, M.B.; Gill, R.A.; Najeeb, U.; Iqbal, N.; Ahmad, R. Citric Acid Assisted Phytoremediation of Copper by Brassica Napus L. Ecotoxicol. Environ. Saf. 2015, 120, 310–317. [Google Scholar] [CrossRef]
- Shakoor, M.B.; Ali, S.; Hameed, A.; Farid, M.; Hussain, S.; Yasmeen, T.; Najeeb, U.; Bharwana, S.A.; Abbasi, G.H. Citric Acid Improves Lead (Pb) Phytoextraction in Brassica Napus L. by Mitigating Pb-Induced Morphological and Biochemical Damages. Ecotoxicol. Environ. Saf. 2014, 109, 38–47. [Google Scholar] [CrossRef]
- Afshan, S.; Ali, S.; Bharwana, S.A.; Rizwan, M.; Farid, M.; Abbas, F.; Ibrahim, M.; Mehmood, M.A.; Abbasi, G.H. Citric Acid Enhances the Phytoextraction of Chromium, Plant Growth, and Photosynthesis by Alleviating the Oxidative Damages in Brassica Napus L. Environ. Sci. Pollut. Res. 2015, 22, 11679–11689. [Google Scholar] [CrossRef]
- Farid, M.; Ali, S.; Rizwan, M.; Ali, Q.; Abbas, F.; Bukhari, S.A.H.; Saeed, R.; Wu, L. Citric Acid Assisted Phytoextraction of Chromium by Sunflower; Morpho-Physiological and Biochemical Alterations in Plants. Ecotoxicol. Environ. Saf. 2017, 145, 90–102. [Google Scholar] [CrossRef]
- Sebastian, A.; Prasad, M. Exogenous Citrate and Malate Alleviate Cadmium Stress in Oryza Sativa L.: Probing Role of Cadmium Localization and Iron Nutrition. Ecotoxicol. Environ. Saf. 2018, 166, 215–222. [Google Scholar] [CrossRef]
- Faraz, A.; Faizan, M.; Sami, F.; Siddiqui, H.; Hayat, S. Supplementation of Salicylic Acid and Citric Acid for Alleviation of Cadmium Toxicity to Brassica Juncea. J. Plant Growth Regul. 2020, 39, 641–655. [Google Scholar] [CrossRef]
- Hassan, M.; Dagari, M.; Muazu, A.; Sanusi, K. Effect of Citric Acid on Cadmium Ion Uptake and Morphological Parameters of Hydroponically Grown Jute Mallow (Corchorus Olitorius). Int. J. Chem. Mater. Environ. Res. 2016, 3, 14–19. [Google Scholar]
- Anwer, S.; Ashraf, M.Y.; Hussain, M.; Ashraf, M.; Jamil, A. Citric Acid Mediated Phytoextraction of Cadmium by Maize (Zea Mays L.). Pak. J. Bot. 2012, 44, 1831–1836. [Google Scholar]
- Sun, Q.; Wang, X.; Ding, S.; Yuan, X. Effects of Exogenous Organic Chelators on Phytochelatins Production and Its Relationship with Cadmium Toxicity in Wheat (Triticum Aestivum L.) under Cadmium Stress. Chemosphere 2005, 60, 22–31. [Google Scholar] [CrossRef] [PubMed]
- Qiu, R.; Liu, W.; Zeng, X.; Tang, Y.; Brewer, E.; Fang, X. Effects of Exogenous Citric Acid and Malic Acid Addition on Nickel Uptake and Translocation in Leaf Mustard (Brassica Juncea Var. Foliosa Bailey) and Alyssum Corsicum. Int. J. Environ. Pollut. 2009, 38, 15–25. [Google Scholar] [CrossRef]
- Irtelli, B.; Navari-Izzo, F. Influence of Sodium Nitrilotriacetate (NTA) and Citric Acid on Phenolic and Organic Acids in Brassica Juncea Grown in Excess of Cadmium. Chemosphere 2006, 65, 1348–1354. [Google Scholar] [CrossRef]
- Ben Massoud, M.; Karmous, I.; El Ferjani, E.; Chaoui, A. Alleviation of Copper Toxicity in Germinating Pea Seeds by IAA, GA3, Ca and Citric Acid. J. Plant Interact. 2018, 13, 21–29. [Google Scholar] [CrossRef] [Green Version]
- Kaur, R.; Yadav, P.; Sharma, A.; Thukral, A.K.; Kumar, V.; Kohli, S.K.; Bhardwaj, R. Castasterone and Citric Acid Treatment Restores Photosynthetic Attributes in Brassica Juncea L. under Cd (II) Toxicity. Ecotoxicol. Environ. Saf. 2017, 145, 466–475. [Google Scholar] [CrossRef]
- Kumar, A.; Pal, L.; Agrawal, V. Glutathione and Citric Acid Modulates Lead-and Arsenic-Induced Phytotoxicity and Genotoxicity Responses in Two Cultivars of Solanum Lycopersicum L. Acta Physiol. Plant. 2017, 39, 1–12. [Google Scholar] [CrossRef]
- Gao, Y.; Miao, C.; Mao, L.; Zhou, P.; Jin, Z.; Shi, W. Improvement of Phytoextraction and Antioxidative Defense in Solanum Nigrum L. under Cadmium Stress by Application of Cadmium-Resistant Strain and Citric Acid. J. Hazard. Mater. 2010, 181, 771–777. [Google Scholar] [CrossRef]
- Najeeb, U.; Xu, L.; Ali, S.; Jilani, G.; Gong, H.; Shen, W.; Zhou, W. Citric Acid Enhances the Phytoextraction of Manganese and Plant Growth by Alleviating the Ultrastructural Damages in Juncus Effusus L. J. Hazard. Mater. 2009, 170, 1156–1163. [Google Scholar] [CrossRef]
- Amir, W.; Farid, M.; Ishaq, H.K.; Farid, S.; Zubair, M.; Alharby, H.F.; Bamagoos, A.A.; Rizwan, M.; Raza, N.; Hakeem, K.R. Accumulation Potential and Tolerance Response of Typha Latifolia L. under Citric Acid Assisted Phytoextraction of Lead and Mercury. Chemosphere 2020, 257, 127247. [Google Scholar] [CrossRef]
- Shahid, M.; Dumat, C.; Pourrut, B.; Silvestre, J.; Laplanche, C.; Pinelli, E. Influence of EDTA and Citric Acid on Lead-Induced Oxidative Stress to Vicia Faba Roots. J. Soils Sediments 2014, 14, 835–843. [Google Scholar] [CrossRef]
- Lu, L.; Tian, S.; Yang, X.; Peng, H.; Li, T. Improved Cadmium Uptake and Accumulation in the Hyperaccumulator Sedum Alfredii: The Impact of Citric Acid and Tartaric Acid. J. Zhejiang Univ. Sci. B 2013, 14, 106–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- An, Y.; Zhou, P.; Xiao, Q.; Shi, D. Effects of Foliar Application of Organic Acids on Alleviation of Aluminum Toxicity in Alfalfa. J. Plant Nutr. Soil Sci. 2014, 177, 421–430. [Google Scholar] [CrossRef]
- Hall, J.L. Cellular Mechanisms for Heavy Metal Detoxification and Tolerance. J. Exp. Bot. 2002, 53, 1–11. [Google Scholar] [CrossRef]
- Ghori, N.H.; Ghori, T.; Hayat, M.Q.; Imadi, S.R.; Gul, A.; Altay, V.; Ozturk, M. Heavy Metal Stress and Responses in Plants. Int. J. Environ. Sci. Technol. 2019, 16, 1807–1828. [Google Scholar] [CrossRef]
- Dresler, S.; Hanaka, A.; Bednarek, W.; Maksymiec, W. Accumulation of Low-Molecular-Weight Organic Acids in Roots and Leaf Segments of Zea Mays Plants Treated with Cadmium and Copper. Acta Physiol. Plant. 2014, 36, 1565–1575. [Google Scholar] [CrossRef] [Green Version]
- Sazanova, K.; Osmolovskaya, N.; Schiparev, S.; Yakkonen, K.; Kuchaeva, L.; Vlasov, D. Organic Acids Induce Tolerance to Zinc-and Copper-Exposed Fungi under Various Growth Conditions. Curr. Microbiol. 2015, 70, 520–527. [Google Scholar] [CrossRef] [PubMed]
- Yu, G.; Ma, J.; Jiang, P.; Li, J.; Gao, J.; Qiao, S.; Zhao, Z. The Mechanism of Plant Resistance to Heavy Metal. IOP Conf. Ser. Earth Environ. Sci. 2019, 310, 052004. [Google Scholar] [CrossRef]
- DalCorso, G.; Farinati, S.; Furini, A. Regulatory Networks of Cadmium Stress in Plants. Plant Signal. Behav. 2010, 5, 663–667. [Google Scholar] [CrossRef]
- Martinoia, E. Vacuolar Transporters–Companions on a Longtime Journey. Plant Physiol. 2018, 176, 1384–1407. [Google Scholar] [CrossRef]
- Vatansever, R.; Ozyigit, I.I.; Filiz, E. Essential and Beneficial Trace Elements in Plants, and Their Transport in Roots: A Review. Appl. Biochem. Biotechnol. 2017, 181, 464–482. [Google Scholar] [CrossRef]
- Salt, D.; Kato, N.; Kramer, U.; Smith, R.; Raskin, I. The Role of Root Exudates in Nickel Hyperaccumulation and Tolerance in Accumulator and Non Accumulator Species of Thlaspi. In Phytoremediation of Contaminated Soil and Water; Terry, N., Banuelos, G., Eds.; CRC Press: Boca Raton, FL, USA, 2000; pp. 189–200. [Google Scholar]
- Ma, J.F.; Hiradate, S. Form of Aluminium for Uptake and Translocation in Buckwheat (Fagopyrum Esculentum Moench). Planta 2000, 211, 355–360. [Google Scholar] [CrossRef]
- Yang, Z.M.; Sivaguru, M.; Horst, W.J.; Matsumoto, H. Aluminium Tolerance Is Achieved by Exudation of Citric Acid from Roots of Soybean (Glycine Max). Physiol. Plant. 2000, 110, 72–77. [Google Scholar] [CrossRef]
- De Noronha, A.L.O.; Guimaraes, L.; Duarte, H.A. Structural and Thermodynamic Analysis of the First Mononuclear Aqueous Aluminum Citrate Complex Using DFT Calculations. J. Chem. Theory Comput. 2007, 3, 930–937. [Google Scholar] [CrossRef]
- Javed, M.T.; Stoltz, E.; Lindberg, S.; Greger, M. Changes in PH and Organic Acids in Mucilage of Eriophorum Angustifolium Roots after Exposure to Elevated Concentrations of Toxic Elements. Environ. Sci. Pollut. Res. 2013, 20, 1876–1880. [Google Scholar] [CrossRef] [Green Version]
- Seshadri, B.; Bolan, N.; Naidu, R. Rhizosphere-Induced Heavy Metal (Loid) Transformation in Relation to Bioavailability and Remediation. J. Soil Sci. Plant Nutr. 2015, 15, 524–548. [Google Scholar] [CrossRef] [Green Version]
- Mariano, E.D.; Jorge, R.A.; Keltjens, W.G.; Menossi, M. Metabolism and Root Exudation of Organic Acid Anions under Aluminium Stress. Braz. J. Plant Physiol. 2005, 17, 157–172. [Google Scholar] [CrossRef]
- Sharma, S.S.; Dietz, K.; Mimura, T. Vacuolar Compartmentalization as Indispensable Component of Heavy Metal Detoxification in Plants. Plant Cell Environ. 2016, 39, 1112–1126. [Google Scholar] [CrossRef]
- Manara, A. Plant Responses to Heavy Metal Toxicity. In Plants and Heavy Metals; Springer: Dordrecht, The Netherlands, 2012; pp. 27–53. [Google Scholar]
- Yeh, T.; Lin, C.; Chuang, C.; Pan, C. The Effect of Varying Soil Organic Levels on Phytoextraction of Cu and Zn Uptake, Enhanced by Chelator EDTA, DTPA, EDDS and Citric Acid. J. Environ. Anal. Toxicol. 2012, 2, 2. [Google Scholar] [CrossRef]
- Dumont, S.; Rivoal, J. Consequences of Oxidative Stress on Plant Glycolytic and Respiratory Metabolism. Front. Plant Sci. 2019, 10, 166. [Google Scholar] [CrossRef] [PubMed]
- Jagtap, V.; Bhargava, S. Variation in the Antioxidant Metabolism of Drought Tolerantand Drought Susceptible Varieties of Sorghum Bicolor (L.) Moench. Exposed to High Light, Low Water and High Temperature Stress. J. Plant Physiol. 1995, 145, 195–197. [Google Scholar] [CrossRef]
- Gong, M.; Chen, S.-N.; Song, Y.-Q.; Li, Z.-G. Effect of Calcium and Calmodulin on Intrinsic Heat Tolerance in Relation to Antioxidant Systems in Maize Seedlings. Funct. Plant Biol. 1997, 24, 371–379. [Google Scholar] [CrossRef]
- Kordrostami, M.; Rabiei, B.; Ebadi, A. Oxidative Stress in Plants: Production, Metabolism, and Biological Roles of Reactive Oxygen Species. In Handbook of Plant and Crop Stress; CRC Press: Boca Raton, FL, USA, 2019; pp. 85–92. [Google Scholar]
- Mittler, R. Oxidative Stress, Antioxidants and Stress Tolerance. Trends Plant Sci. 2002, 7, 405–410. [Google Scholar] [CrossRef]
- Gupta, K.J.; Shah, J.K.; Brotman, Y.; Jahnke, K.; Willmitzer, L.; Kaiser, W.M.; Bauwe, H.; Igamberdiev, A.U. Inhibition of Aconitase by Nitric Oxide Leads to Induction of the Alternative Oxidase and to a Shift of Metabolism towards Biosynthesis of Amino Acids. J. Exp. Bot. 2012, 63, 1773–1784. [Google Scholar] [CrossRef] [Green Version]
- Demidchik, V. Mechanisms of Oxidative Stress in Plants: From Classical Chemistry to Cell Biology. Environ. Exp. Bot. 2015, 109, 212–228. [Google Scholar] [CrossRef]
- Preciado-Rangel, P.; Gaucín-Delgado, J.M.; Salas-Pérez, L.; Chavez, E.S.; Mendoza-Vllarreal, R.; Ortiz, J.C.R. The Effect of Citric Acid on the Phenolic Compounds, Flavonoids and Antioxidant Capacity of Wheat Sprouts. Rev. Fac. Cienc. Agrar. UNCuyo 2018, 50, 119–127. [Google Scholar]
- Salas-Pérez, L.; Gaucín Delgado, J.; Preciado-Rangel, P.; Gonzales Fuentes, J.; Ayala Garay, A.; Segura Castruita, M. The Application of Citric Acid Increases the Quality and Antioxidant Capacity of Lentil Sprouts. Rev. Mex. Cienc. Agrícolas 2018, 9, 4301–4309. [Google Scholar]
- Mallhi, Z.I.; Rizwan, M.; Mansha, A.; Ali, Q.; Asim, S.; Ali, S.; Hussain, A.; Alrokayan, S.H.; Khan, H.A.; Alam, P. Citric Acid Enhances Plant Growth, Photosynthesis, and Phytoextraction of Lead by Alleviating the Oxidative Stress in Castor Beans. Plants 2019, 8, 525. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Balibrea, S.; Moreno, D.A.; García-Viguera, C. Influence of Light on Health-promoting Phytochemicals of Broccoli Sprouts. J. Sci. Food Agric. 2008, 88, 904–910. [Google Scholar] [CrossRef]
- Zhao, Z.; Hu, L.; Hu, T.; Fu, J. Differential Metabolic Responses of Two Tall Fescue Genotypes to Heat Stress. Acta Prataculturae Sin. 2015, 24, 58–69. [Google Scholar]
- Maxwell, D.P.; Wang, Y.; McIntosh, L. The Alternative Oxidase Lowers Mitochondrial Reactive Oxygen Production in Plant Cells. Proc. Natl. Acad. Sci. USA 1999, 96, 8271–8276. [Google Scholar] [CrossRef] [Green Version]
- Kumari, A.; Pathak, P.K.; Bulle, M.; Igamberdiev, A.U.; Gupta, K.J. Alternative Oxidase Is an Important Player in the Regulation of Nitric Oxide Levels under Normoxic and Hypoxic Conditions in Plants. J. Exp. Bot. 2019, 70, 4345–4354. [Google Scholar] [CrossRef] [PubMed]
- Naikoo, M.I.; Dar, M.I.; Raghib, F.; Jaleel, H.; Ahmad, B.; Raina, A.; Khan, F.A.; Naushin, F. Role and Regulation of Plants Phenolics in Abiotic Stress Tolerance: An Overview. Plant Signal. Mol. 2019, 157–168. [Google Scholar]
- Sharma, A.; Shahzad, B.; Kumar, V.; Kohli, S.K.; Sidhu, G.P.S.; Bali, A.S.; Handa, N.; Kapoor, D.; Bhardwaj, R.; Zheng, B. Phytohormones Regulate Accumulation of Osmolytes under Abiotic Stress. Biomolecules 2019, 9, 285. [Google Scholar] [CrossRef] [Green Version]
- Chalker-Scott, L. Do Anthocyanins Function as Osmoregulators in Leaf Tissues? Adv. Bot. Res. 2002, 37, 103–106. [Google Scholar]
- Wahid, A.; Ghazanfar, A. Possible Involvement of Some Secondary Metabolites in Salt Tolerance of Sugarcane. J. Plant Physiol. 2006, 163, 723–730. [Google Scholar] [CrossRef]
- Reynoso-Camacho, R.; Ramos-Gomez, M.; Loarca-Pina, G. Advances in Agricultural and Food Biotechnology. Bioactive Components in Common Beans (Phaseolus Vulgaris L.); Research Signpost: Trivandrum, India, 2006; pp. 217–236. [Google Scholar]
- Li, W.; Zhang, J.; Tan, S.; Zheng, Q.; Zhao, X.; Gao, X.; Lu, Y. Citric Acid-enhanced Dissolution of Polyphenols during Soaking of Different Teas. J. Food Biochem. 2019, 43, e13046. [Google Scholar] [CrossRef]
- Szabados, L.; Savouré, A. Proline: A Multifunctional Amino Acid. Trends Plant Sci. 2010, 15, 89–97. [Google Scholar] [CrossRef]
- Kavi Kishor, P.B.; Hima Kumari, P.; Sunita, M.; Sreenivasulu, N. Role of Proline in Cell Wall Synthesis and Plant Development and Its Implications in Plant Ontogeny. Front. Plant Sci. 2015, 6, 544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yıldırım, E.; Dursun, A. Effect of Foliar Salicylic Acid Applications on Plant Growth and Yield of Tomato under Greenhouse Conditions. In Proceedings of the International Symposium on Strategies Towards Sustainability of Protected Cultivation in Mild Winter Climate, Antalya, Turkey, 6–11 April 2008; pp. 395–400. [Google Scholar]
- Ulloa, J.; Aguilar-Pusian, J.; Rosas-Ulloa, P.; Galavíz-Ortíz, K.M.d.C.; Ulloa-Rangel, B. Effect of Soaking Conditions with Citric Acid, Ascorbic Acid and Potassium Sorbate on the Physicochemical and Microbiological Quality of Minimally Processed Jackfruit. CyTA-J. Food 2010, 8, 193–199. [Google Scholar] [CrossRef]
- Limón, R.I.; Peñas, E.; Torino, M.I.; Martínez-Villaluenga, C.; Dueñas, M.; Frias, J. Fermentation Enhances the Content of Bioactive Compounds in Kidney Bean Extracts. Food Chem. 2015, 172, 343–352. [Google Scholar] [CrossRef] [Green Version]
- Markakis, P.; Jurd, L. Anthocyanins and Their Stability in Foods. Crit. Rev. Food Sci. Nutr. 1974, 4, 437–456. [Google Scholar] [CrossRef]
- De la Fuente, J.M.; Ramírez-Rodríguez, V.; Cabrera-Ponce, J.L.; Herrera-Estrella, L. Aluminum Tolerance in Transgenic Plants by Alteration of Citrate Synthesis. Science 1997, 276, 1566–1568. [Google Scholar] [CrossRef] [PubMed]
- Koyama, H.; Kawamura, A.; Kihara, T.; Hara, T.; Takita, E.; Shibata, D. Overexpression of Mitochondrial Citrate Synthase in Arabidopsis Thaliana Improved Growth on a Phosphorus-Limited Soil. Plant Cell Physiol. 2000, 41, 1030–1037. [Google Scholar] [CrossRef] [PubMed]
- Guerinot, M.L. Improving Rice Yields—Ironing out the Details. Nat. Biotechnol. 2001, 19, 417–418. [Google Scholar] [CrossRef]
- Han, D.; Wang, L.; Wang, Y.; Yang, G.; Gao, C.; Yu, Z.; Li, T.; Zhang, X.; Ma, L.; Xu, X. Overexpression of Malus Xiaojinensis CS1 Gene in Tobacco Affects Plant Development and Increases Iron Stress Tolerance. Sci. Hortic. 2013, 150, 65–72. [Google Scholar] [CrossRef]
- Delhaize, E.; Hebb, D.M.; Ryan, P.R. Expression of a Pseudomonas Aeruginosa Citrate Synthase Gene in Tobacco Is Not Associated with Either Enhanced Citrate Accumulation or Efflux. Plant Physiol. 2001, 125, 2059–2067. [Google Scholar] [CrossRef] [Green Version]
- Anoop, V.M.; Basu, U.; McCammon, M.T.; McAlister-Henn, L.; Taylor, G.J. Modulation of Citrate Metabolism Alters Aluminum Tolerance in Yeast and Transgenic Canola Overexpressing a Mitochondrial Citrate Synthase. Plant Physiol. 2003, 132, 2205–2217. [Google Scholar] [CrossRef] [Green Version]
- Kruse, A.; Fieuw, S.; Heineke, D.; Müller-Röber, B. Antisense Inhibition of Cytosolic NADP-Dependent Isocitrate Dehydrogenase in Transgenic Potato Plants. Planta 1998, 205, 82–91. [Google Scholar] [CrossRef]
- Diatloff, E.; Roberts, M.; Sanders, D.; Roberts, S.K. Characterization of Anion Channels in the Plasma Membrane of Arabidopsis Epidermal Root Cells and the Identification of a Citrate-Permeable Channel Induced by Phosphate Starvation. Plant Physiol. 2004, 136, 4136–4149. [Google Scholar] [CrossRef] [Green Version]
- Yamaguchi, M.; Sasaki, T.; Sivaguru, M.; Yamamoto, Y.; Osawa, H.; Ahn, S.J.; Matsumoto, H. Evidence for the Plasma Membrane Localization of Al-Activated Malate Transporter (ALMT1). Plant Cell Physiol. 2005, 46, 812–816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, X.; Li, R.; Shi, J.; Wang, J.; Sun, Q.; Zhang, H.; Xing, Y.; Qi, Y.; Zhang, N.; Guo, Y.-D. Brassica Oleracea MATE Encodes a Citrate Transporter and Enhances Aluminum Tolerance in Arabidopsis Thaliana. Plant Cell Physiol. 2014, 55, 1426–1436. [Google Scholar] [CrossRef] [Green Version]
- Lei, G.J.; Yokosho, K.; Yamaji, N.; Ma, J.F. Two MATE Transporters with Different Subcellular Localization Are Involved in Al Tolerance in Buckwheat. Plant Cell Physiol. 2017, 58, 2179–2189. [Google Scholar] [CrossRef]
- Liu, M.Y.; Lou, H.Q.; Chen, W.W.; Piñeros, M.A.; Xu, J.M.; Fan, W.; Kochian, L.V.; Zheng, S.J.; Yang, J.L. Two Citrate Transporters Coordinately Regulate Citrate Secretion from Rice Bean Root Tip under Aluminum Stress. Plant Cell Environ. 2018, 41, 809–822. [Google Scholar] [CrossRef]
- Maron, L.G.; Piñeros, M.A.; Guimarães, C.T.; Magalhaes, J.V.; Pleiman, J.K.; Mao, C.; Shaff, J.; Belicuas, S.N.; Kochian, L.V. Two Functionally Distinct Members of the MATE (Multi-drug and Toxic Compound Extrusion) Family of Transporters Potentially Underlie Two Major Aluminum Tolerance QTLs in Maize. Plant J. 2010, 61, 728–740. [Google Scholar] [CrossRef]
- Yokosho, K.; Yamaji, N.; Ma, J.F. Isolation and Characterisation of Two MATE Genes in Rye. Funct. Plant Biol. 2010, 37, 296–303. [Google Scholar] [CrossRef]
- Yokosho, K.; Yamaji, N.; Ma, J.F. An Al-inducible MATE Gene Is Involved in External Detoxification of Al in Rice. Plant J. 2011, 68, 1061–1069. [Google Scholar] [CrossRef]
- Fan, W.; Xu, J.-M.; Lou, H.-Q.; Xiao, C.; Chen, W.-W.; Yang, J.-L. Physiological and Molecular Analysis of Aluminium-Induced Organic Acid Anion Secretion from Grain Amaranth (Amaranthus Hypochondriacus L.) Roots. Int. J. Mol. Sci. 2016, 17, 608. [Google Scholar] [CrossRef] [Green Version]
- Furukawa, J.; Yamaji, N.; Wang, H.; Mitani, N.; Murata, Y.; Sato, K.; Katsuhara, M.; Takeda, K.; Ma, J.F. An Aluminum-Activated Citrate Transporter in Barley. Plant Cell Physiol. 2007, 48, 1081–1091. [Google Scholar] [CrossRef] [Green Version]
- Magalhaes, J.V.; Liu, J.; Guimaraes, C.T.; Lana, U.G.; Alves, V.M.; Wang, Y.-H.; Schaffert, R.E.; Hoekenga, O.A.; Pineros, M.A.; Shaff, J.E. A Gene in the Multidrug and Toxic Compound Extrusion (MATE) Family Confers Aluminum Tolerance in Sorghum. Nat. Genet. 2007, 39, 1156–1161. [Google Scholar] [CrossRef]
- Ribeiro, A.P.; de Souza, W.R.; Martins, P.K.; Vinecky, F.; Duarte, K.E.; Basso, M.F.; da Cunha, B.A.; Campanha, R.B.; de Oliveira, P.A.; Centeno, D.C. Overexpression of BdMATE Gene Improves Aluminum Tolerance in Setaria Viridis. Front. Plant Sci. 2017, 8, 865. [Google Scholar] [CrossRef] [Green Version]
- Zhu, D.; Li, R.; Liu, X.; Sun, M.; Wu, J.; Zhang, N.; Zhu, Y. The Positive Regulatory Roles of the TIFY10 Proteins in Plant Responses to Alkaline Stress. PLoS ONE 2014, 9, e111984. [Google Scholar] [CrossRef]
- Sun, M.; Sun, X.; Zhao, Y.; Zhao, C.; DuanMu, H.; Yu, Y.; Ji, W.; Zhu, Y. Ectopic Expression of GsPPCK3 and SCMRP in Medicago Sativa Enhances Plant Alkaline Stress Tolerance and Methionine Content. PLoS ONE 2014, 9, e89578. [Google Scholar] [CrossRef]
- Barone, P.; Rosellini, D.; LaFayette, P.; Bouton, J.; Veronesi, F.; Parrott, W. Bacterial Citrate Synthase Expression and Soil Aluminum Tolerance in Transgenic Alfalfa. Plant Cell Rep. 2008, 27, 893–901. [Google Scholar] [CrossRef]
- Deng, W.; Luo, K.; Li, Z.; Yang, Y.; Hu, N.; Wu, Y. Overexpression of Citrus Junos Mitochondrial Citrate Synthase Gene in Nicotiana Benthamiana Confers Aluminum Tolerance. Planta 2009, 230, 355–365. [Google Scholar] [CrossRef]
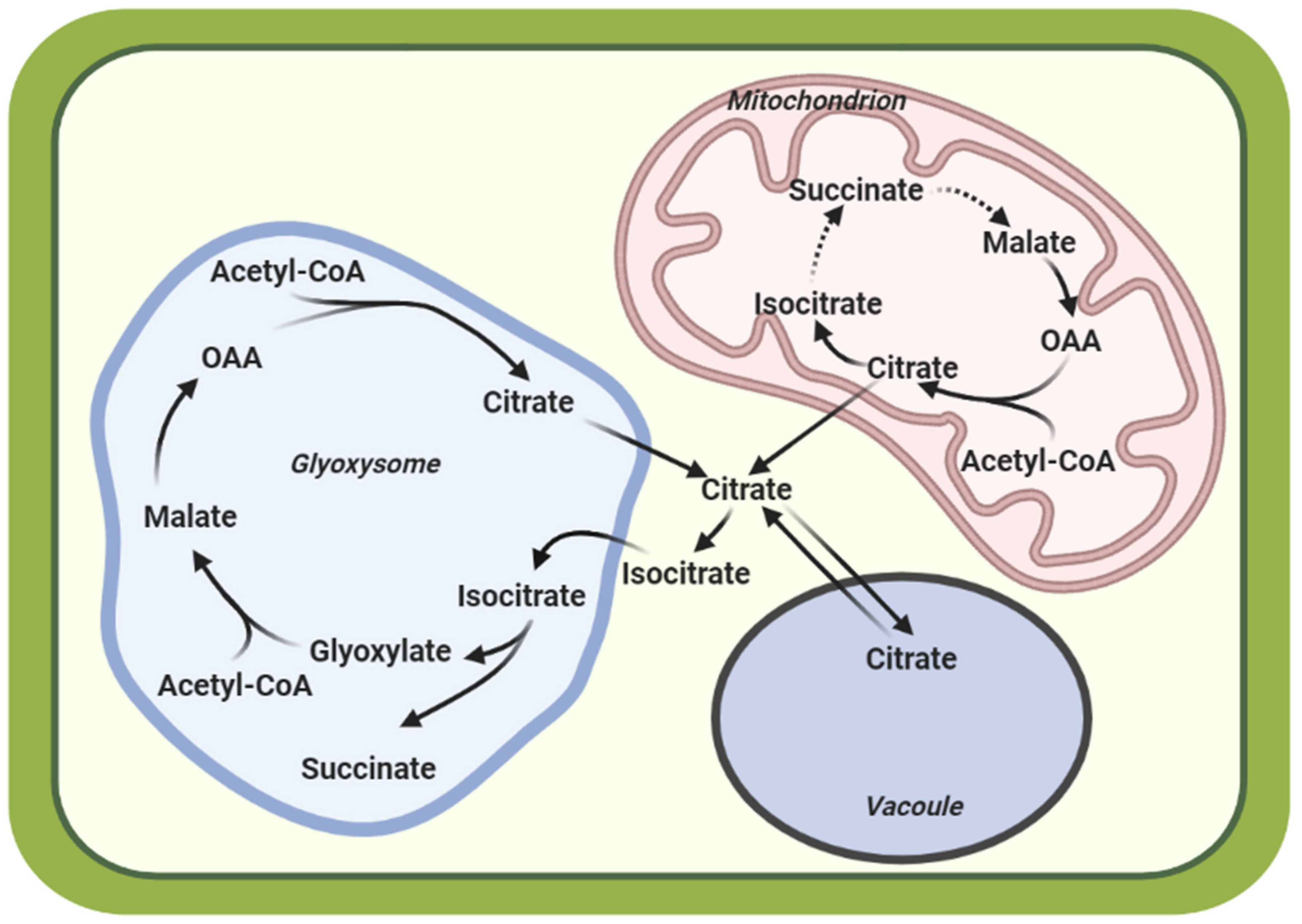
- Sweetlove, L.J.; Beard, K.F.; Nunes-Nesi, A.; Fernie, A.R.; Ratcliffe, R.G. Not Just a Circle: Flux Modes in the Plant TCA Cycle. Trends Plant Sci. 2010, 15, 462–470. [Google Scholar] [CrossRef]
- Ganjewala, D.; Kaur, G.; Srivastava, N. Metabolic Engineering of Stress Protectant Secondary Metabolites to Confer Abiotic Stress Tolerance in Plants. In Molecular Approaches in Plant Biology and Environmental Challenges; Springer: Singapore, 2019; pp. 207–227. [Google Scholar]
- Araújo, W.L.; Martins, A.O.; Fernie, A.R.; Tohge, T. 2-Oxoglutarate: Linking TCA Cycle Function with Amino Acid, Glucosinolate, Flavonoid, Alkaloid, and Gibberellin Biosynthesis. Front. Plant Sci. 2014, 5, 552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katz, E.; Fon, M.; Lee, Y.; Phinney, B.; Sadka, A.; Blumwald, E. The Citrus Fruit Proteome: Insights into Citrus Fruit Metabolism. Planta 2007, 226, 989–1005. [Google Scholar] [CrossRef]
- Walker, R.P.; Battistelli, A.; Moscatello, S.; Chen, Z.-H.; Leegood, R.C.; Famiani, F. Phosphoenolpyruvate Carboxykinase in Cherry (Prunus Avium L.) Fruit during Development. J. Exp. Bot. 2011, 62, 5357–5365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cercós, M.; Soler, G.; Iglesias, D.J.; Gadea, J.; Forment, J.; Talón, M. Global Analysis of Gene Expression during Development and Ripening of Citrus Fruit Flesh. A Proposed Mechanism for Citric Acid Utilization. Plant Mol. Biol. 2006, 62, 513–527. [Google Scholar] [CrossRef] [PubMed]
- Fatland, B.L.; Nikolau, B.J.; Wurtele, E.S. Reverse Genetic Characterization of Cytosolic Acetyl-CoA Generation by ATP-Citrate Lyase in Arabidopsis. Plant Cell 2005, 17, 182–203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Degu, A.; Hatew, B.; Nunes-Nesi, A.; Shlizerman, L.; Zur, N.; Katz, E.; Fernie, A.R.; Blumwald, E.; Sadka, A. Inhibition of Aconitase in Citrus Fruit Callus Results in a Metabolic Shift towards Amino Acid Biosynthesis. Planta 2011, 234, 501–513. [Google Scholar] [CrossRef]
- Li, S.; Wang, W.; Ma, Y.; Liu, S.; Grierson, D.; Yin, X.; Chen, K. Citrus CitERF6 Contributes to Citric Acid Degradation via Upregulation of CitAclα1, Encoding ATP-Citrate Lyase Subunit α. J. Agric. Food Chem. 2020, 68, 10081–10087. [Google Scholar] [CrossRef]
- Signorelli, S.; Arellano, J.B.; Melø, T.B.; Borsani, O.; Monza, J. Proline Does Not Quench Singlet Oxygen: Evidence to Reconsider Its Protective Role in Plants. Plant Physiol. Biochem. 2013, 64, 80–83. [Google Scholar] [CrossRef] [Green Version]
- Shimajiri, Y.; Oonishi, T.; Ozaki, K.; Kainou, K.; Akama, K. Genetic Manipulation of the Γ-aminobutyric Acid (GABA) Shunt in Rice: Overexpression of Truncated Glutamate Decarboxylase (GAD 2) and Knockdown of Γ-aminobutyric Acid Transaminase (GABA-T) Lead to Sustained and High Levels of GABA Accumulation in Rice Kernels. Plant Biotechnol. J. 2013, 11, 594–604. [Google Scholar]
- Ali, Q.; Habib-ur-Rehman Athar, M.Z.; Haider, S.S.; Aslam, N.; Shehzad, F.; Naseem, J.; Ashraf, R.; Ali, A.; Hussain, S.M. Role of Amino Acids in Improving Abiotic Stress Tolerance to Plants. In Plant Tolerance to Environmental Stress: Role of Phytoprotectants; Hasanuzzaman, M., Fujita, M., Oku, H., Islam, M.T., Eds.; CRC Press: Boca Raton, FL, USA, 2019; pp. 175–204. [Google Scholar]
- Igamberdiev, A.U.; Eprintsev, A.T. Organic Acids: The Pools of Fixed Carbon Involved in Redox Regulation and Energy Balance in Higher Plants. Front. Plant Sci. 2016, 7, 1042. [Google Scholar] [CrossRef] [Green Version]
- Ludwig, M. The Roles of Organic Acids in C4 Photosynthesis. Front. Plant Sci. 2016, 7, 647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, Y.-C.; Kuang, J.-F.; Chen, J.-Y.; Liu, X.-C.; Xiao, Y.-Y.; Fu, C.-C.; Wang, J.-N.; Wu, K.-Q.; Lu, W.-J. Banana Transcription Factor MaERF11 Recruits Histone Deacetylase MaHDA1 and Represses the Expression of MaACO1 and Expansins during Fruit Ripening. Plant Physiol. 2016, 171, 1070–1084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crifò, T.; Puglisi, I.; Petrone, G.; Recupero, G.R.; Piero, A.R.L. Expression Analysis in Response to Low Temperature Stress in Blood Oranges: Implication of the Flavonoid Biosynthetic Pathway. Gene 2011, 476, 1–9. [Google Scholar] [CrossRef]
- Xing, S.; van Deenen, N.; Magliano, P.; Frahm, L.; Forestier, E.; Nawrath, C.; Schaller, H.; Gronover, C.S.; Prüfer, D.; Poirier, Y. ATP Citrate Lyase Activity Is Post-translationally Regulated by Sink Strength and Impacts the Wax, Cutin and Rubber Biosynthetic Pathways. Plant J. 2014, 79, 270–284. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Du, X.; Liang, X.; Han, R.; Liang, Z.; Liu, Y.; Liu, F.; Zhao, J. Different Roles of the Mevalonate and Methylerythritol Phosphate Pathways in Cell Growth and Tanshinone Production of Salvia Miltiorrhiza Hairy Roots. PLoS ONE 2012, 7, e46797. [Google Scholar] [CrossRef] [PubMed]
- Liao, P.; Hemmerlin, A.; Bach, T.J.; Chye, M.-L. The Potential of the Mevalonate Pathway for Enhanced Isoprenoid Production. Biotechnol. Adv. 2016, 34, 697–713. [Google Scholar] [CrossRef]
- Lipko, A.; Swiezewska, E. Isoprenoid Generating Systems in Plants—A Handy Toolbox How to Assess Contribution of the Mevalonate and Methylerythritol Phosphate Pathways to the Biosynthetic Process. Prog. Lipid Res. 2016, 63, 70–92. [Google Scholar] [CrossRef]
- Li, C.; Schilmiller, A.L.; Liu, G.; Lee, G.I.; Jayanty, S.; Sageman, C.; Vrebalov, J.; Giovannoni, J.J.; Yagi, K.; Kobayashi, Y. Role of β-Oxidation in Jasmonate Biosynthesis and Systemic Wound Signaling in Tomato. Plant Cell 2005, 17, 971–986. [Google Scholar] [CrossRef] [Green Version]
- Piero, A.R.L.; Cicero, L.L.; Puglisi, I. The Metabolic Fate of Citric Acid as Affected by Cold Storage in Blood Oranges. J. Plant Biochem. Biotechnol. 2014, 23, 161–166. [Google Scholar] [CrossRef]
- Chen, M.; Jiang, Q.; Yin, X.-R.; Lin, Q.; Chen, J.-Y.; Allan, A.C.; Xu, C.-J.; Chen, K.-S. Effect of Hot Air Treatment on Organic Acid-and Sugar-Metabolism in Ponkan (Citrus Reticulata) Fruit. Sci. Hortic. 2012, 147, 118–125. [Google Scholar] [CrossRef]
- Phan, T.-T.; Li, J.; Sun, B.; Liu, J.-Y.; Zhao, W.-H.; Huang, C.; Yang, L.-T.; Li, Y.-R. ATP-Citrate Lyase Gene (SoACLA-1), a Novel ACLA Gene in Sugarcane, and Its Overexpression Enhance Drought Tolerance of Transgenic Tobacco. Sugar. Tech. 2017, 19, 258–269. [Google Scholar] [CrossRef]
- Ye, N.; Zhu, G.; Liu, Y.; Zhang, A.; Li, Y.; Liu, R.; Shi, L.; Jia, L.; Zhang, J. Ascorbic Acid and Reactive Oxygen Species Are Involved in the Inhibition of Seed Germination by Abscisic Acid in Rice Seeds. J. Exp. Bot. 2012, 63, 1809–1822. [Google Scholar] [CrossRef] [Green Version]
- Sadak, M.S.; Elhamid, E.; Mostafa, H.M. Alleviation of Adverse Effects of Salt Stress in Wheat Cultivars by Foliar Treatment with Antioxidants I. Changes in Growth, Some Biochemical Aspects and Yield Quantity and Quality. Am. Eur. J. Agric. Environ. Sci. 2013, 13, 1476–1487. [Google Scholar]
- Hedrich, R.; Marten, I. Malate-induced Feedback Regulation of Plasma Membrane Anion Channels Could Provide a CO2 Sensor to Guard Cells. EMBO J. 1993, 12, 897–901. [Google Scholar] [CrossRef]
- Van Oosten, M.J.; Pepe, O.; De Pascale, S.; Silletti, S.; Maggio, A. The Role of Biostimulants and Bioeffectors as Alleviators of Abiotic Stress in Crop Plants. Chem. Biol. Technol. Agric. 2017, 4, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Chavoushi, M.; Najafi, F.; Salimi, A.; Angaji, S. Improvement in Drought Stress Tolerance of Safflower during Vegetative Growth by Exogenous Application of Salicylic Acid and Sodium Nitroprusside. Ind. Crops Prod. 2019, 134, 168–176. [Google Scholar] [CrossRef]
- Roy, P.R.; Tahjib-Ul-Arif, M.; Polash, M.A.S.; Hossen, M.Z.; Hossain, M.A. Physiological Mechanisms of Exogenous Calcium on Alleviating Salinity-Induced Stress in Rice (Oryza Sativa L.). Physiol. Mol. Biol. Plants 2019, 25, 611–624. [Google Scholar] [CrossRef]
- Tahjib-Ul-Arif, M.; Afrin, S.; Polash, M.A.S.; Akter, T.; Ray, S.R.; Hossain, M.T.; Hossain, M.A. Role of Exogenous Signaling Molecules in Alleviating Salt-Induced Oxidative Stress in Rice (Oryza Sativa L.): A Comparative Study. Acta Physiol. Plant. 2019, 41, 1–14. [Google Scholar] [CrossRef]
- Sohag, A.A.M.; Tahjib-Ul-Arif, M.; Afrin, S.; Khan, M.K.; Hannan, M.A.; Skalicky, M.; Mortuza, M.G.; Brestic, M.; Hossain, M.A.; Murata, Y. Insights into Nitric Oxide-Mediated Water Balance, Antioxidant Defence and Mineral Homeostasis in Rice (Oryza Sativa L.) under Chilling Stress. Nitric Oxide 2020, 100, 7–16. [Google Scholar] [CrossRef]
- Sohag, A.A.M.; Tahjib-Ul-Arif, M.; Polash, M.A.S.; Chowdhury, M.B.; Afrin, S.; Burritt, D.J.; Murata, Y.; Hossain, M.A.; Hossain, M.A. Exogenous Glutathione-Mediated Drought Stress Tolerance in Rice (Oryza Sativa L.) Is Associated with Lower Oxidative Damage and Favorable Ionic Homeostasis. Iran. J. Sci. Technol. Trans. Sci. 2020, 44, 955–971. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stress | Treatment | Plant Species | Organ/Tissue | Duration | Endogenous CA Level | Reference |
|---|---|---|---|---|---|---|
| Salinity | 50 to 250 mM NaCl | Helianthus annuus | Shoot | 7 days | ↑ | [19] |
| 20 and 120 mM NaCl | Solanum lycopersicum | Shoot | 10 days | ↑ | [21] | |
| 100 and 200 mM NaCl | Root exudates | 4 weeks | ↑ | [20] | ||
| 100 and 200 mM NaCl | Acacia nilotica | Root exudates | 4 weeks | ↑ | [20] | |
| 25 to 200 mM NaCl | Trigonella foenum-graecum | Seedling | 5 days | ↑ | [22] | |
| Drought | 40, 70, and 100% FC | Solanum lycopersicum | Fruit | 120 days | ↑ | [27] |
| Irrigated and dryland | Gossypium hirsutum | Leaf | 108 days | ↑ | [23] | |
| Withholding water | Clusia sp. | Leaf | 16 days | ↑ | [24] | |
| Withholding water | Aptenia cordifolia | Leaf | 10 days | ↑ | [25] | |
| −20, −20 to −40, and −40 to −60 kPa | Solanum tuberosum | Tuber | 42 days | = | [26] | |
| Heat | 25/20 °C and 35/30 °C (D/N) | Festuca arundinacea | Leaf | 28 days | ↑ | [29] |
| 22°C and 30 °C (daytime) | Solanum tuberosum | Tuber | 42 days | = | [26] | |
| 20/15 °C and 35/30 °C | Poa pratensis | Leaf | 18 days | = | [28] | |
| 30/25 °C and 45/40 °C | Hybrid bermudagrass | Leaf | 18 days | ↑ | [28] | |
| 25/20 °C and 35/30 °C (D/N) | Lolium arundinaceum | Leaf | 15 days | ↑ | [17] | |
| HMs | 50 µM CdCl2 | Solanum nigrum | Root | 24 h | ↑ | [30] |
| 0.6 mM CdCl2 | Brassica juncea | Shoot | 7 days | ↑ | [33] | |
| 150 μM NiCl2.6H20 | Amaranthus paniculatus | Leaf and root | 1 week | ↑ | [32] | |
| 50 µM K2Cr2O7 | Oryza sativa | Root exudates | 8 days | ↑ | [34] | |
| 100 µM K2Cr2O7 | Oryza sativa | Root exudates | 8 days | ↓ | [34] | |
| 100 µM K2Cr2O7 | Oryza sativa | Root exudates | 16 days | ↑ | [34] | |
| 50 μM AlCl3 | Secale cereale and Triticum aestivum | Root exudates | 12 h | ↑ | [37] | |
| 15 μM AlCl3.6H2O | Glycine max | Root exudates | 24 h | ↑ | [39] | |
| 50 μM AlCl3 | Brachiaria brizantha | Root exudates | 12 h | ↓ | [36] | |
| 30 μM AlCl3 | Cassia tora | Root exudates | 9 h | ↑ | [35] |
| Plant Species | Stress | CA Treatments and Method of Application | Effects | Outcomes | References |
|---|---|---|---|---|---|
| Gossypium barbadense (Cotton) | Salt (205, 135, and 35 mM NaCl) | Foliar spray of 2.5 g L−1 potassium citrate | Increased growth, yield, and photosynthetic pigments. Increased TSS, TSP, TPC, FAA, and proline. Enhanced CAT, POX, and SOD activities. | Improved growth and yield but no effects on fiber properties. Increased salt tolerance. | [18] |
| Carica papaya (Papaya) | Salt (NaCl) | Seed soaking with CA (500 mg L−1) as sildenafil citrate | Increased germination rate. | Improved the tolerance and development of papaya plants in saline environments. | [42] |
| Phaseolus vulgaris (Common Bean) | Drought | Spraying of CA (0.5, 1.0, 1.5, and 2 g L−1) | Increased relative water content (RWC) and Chl. Increased plant growth and productivity. | Application of CA at 1.5 g L−1 was most effective for drought alleviation. | [14] |
| Zea mays (Maize) | Salt (NaCl) (4.2–4.6 dSm−1) | Foliar spray of CA with ascorbic acid and salicylic acid (100 or 200 ppm) | Increased leaf area index, net assimilation rate, growth rate, and photosynthetic pigments. Enhanced CAT, POX, PPO, and PAL activities. Decreased proline and Na+. Increased K+. | Improved tolerance to salinity. Enhanced growth and yield. | [43] |
| Leymus chinensis (Chinese ryegrass) | Salt (200 mM NaCl) and alkaline stress (100 mM Na2CO3) | Irrigation with CA (50 mg L−1) | Increased growth and CA exudation. Increased RWC and CO2 assimilation rate. Enhanced MDA content, CAT, APX, and SOD activities. | Improved tolerance to saline and alkaline stress. | [46] |
| Gossypium barbadense (Cotton) | Drought | Foliar spray of CA (500 ppm) | Increased growth, number of fruiting branches, number of open bolls per plant, seed index, boll weight, lint percentage, and seed cotton yield. Increased Chl a, Chl b, Chl a+b, carotenoid, and proline contents in leaves. Enhanced CAT and POX activities. | Reduced drought sensitivity but no significant effects on fiber properties. | [15] |
| Hibiscus sabdariffa (Roselle) | Salt (75 mM NaCl) | Foliar spray of CA (10 mM) | Increased TPC and proline accumulation. Reduced GSH content. Enhanced SOD activity but decreased CAT, POX, and PAL activities | Improved flower production under salinity condition. | [45] |
| Brassica oleracea (Cabbage) | Drought | Spraying of CA (5 mM) | Increased P uptake. Decreased hydrogen peroxide production. | Alleviated drought-induced oxidative stress. | [49] |
| Melissa officinalis (Lemon balm) | Salt (0.0, 1.6, 3.1, and 6.3 dSm−1) | Foliar spray of CA (0.3 g L−1) | Increased levels of α-pinene, β-bisabolene, monoterpene hydrocarbons (MCH) and oxygenated sesquiterpenes (SCHO). | Improved growth. | [44] |
| Beta vulgaris (Sugar beet) | Salt (12.50 dSm−1) | Soil application of CA (300 mg L−1) | Increased K, N, and P when added in combination with tomato peel extract. Increased CAT and POX activity when added in combination with banana peel extract. | Banana extract and CA reduced soil salinity. Increased root and sugar yield. | [47] |
| Plant Species | Stress | CA dose | Effects | Outcomes | Reference |
|---|---|---|---|---|---|
| Lolium arundinaceum | Heat stress: (25/20 °C and 35/30 °C, day/night) in growth chambers | Foliar spraying of CA (0, 0.2, 2, and 20 mM) | Increased growth. Increased Chl content, photochemical efficiency (Fv/Fm) and SOD, POX, and CAT activities. Decreased EL and MDA content. Increased expression of heat shock protein genes. | Alleviated growth and physiological damage caused by high temperature | [17] |
| Lycopersicon esculentum | Heat stress as manipulated by late summer sowing (air temp up to 35 °C) | Spraying of CA (2.5 and 5 g L−1) | Increased yield and fertility of pollen grains. Increased vitamin C content, TSS, minerals. Increased stem thickness, epidermis, phloem and xylem tissues. Enhanced POX, SOD, and CAT activities. | Increased yield during late summer | [50] |
| Hibiscus rosa-sinensis | Cold stress (˂10 °C) | CA (5 mM) in nutrient solution | Increased the number of leaves remaining on plants grown under low-illumination. | Suppressed defoliation | [51] |
| Leymus chinensis | Alkaline stress (100 mM Na2CO3) | Spraying of CA (50 mg L−1) | Increased growth, relative growth rate, and photosynthesis. Enhanced CAT, SOD, and APX activities. | Increased stress tolerance | [46] |
| Rosa roxburghii | Calcareous yellow soil (pH higher than 8) | CA (40, 80 and 120 mg kg−1 soil) | Increased growth, total biomass, root development, root-shoot ratio, and total root surface area. Increased nutrient contents. | Increased seedling growth | [52] |
| Plant Species | HM Stress | Treatments | Effects | Outcomes | References |
|---|---|---|---|---|---|
| Brassica napus | Cu (50 and 100 µM as CuSO4) | CA (2.5 mM) in nutrient solution | Increased plant growth, biomass, Chl content, stomatal conductance, and water use efficiency. Enhanced POX, SOD, CAT, and APX activities. Reduced H2O2, MDA, and EL. | Minimized Cu toxicity and enhanced biomass production. | [56] |
| Brassica napus | Cd (10 and 50 µM as CdCl2) | CA (2.5 mM) in solution medium | Enhanced plant growth and biomass, gas exchange activities, and antioxidant enzymes activity. Reduced oxidative stress by reducing H2O2 and MDA production and decreasing EL. | Mitigated Cd stress. | [16] |
| Solanum nigram | Cd (50 mg Cd2+ kg−1 dry soil) | CA (20 mmol kg−1 soil) applied in soil | Promoted plant growth, biomass, and antioxidative defense e.g., SOD and POX activity at initial stage. | Slightly reduced Cd stress. | [70] |
| Brassica juncea | Cd (0.6 mmol kg−1 soil as CdCl2) | CA (0.6 mmol kg−1 soil) applied in soil | Increased plant height, Chl a+b, carotenoid, anthocyanins, and flavonoids in leaves. Non-significant increment of the activities of SOD, POX, CAT, and GPX. Reduced MDA levels. | Alleviated Cd-induced toxicity. | [68] |
| Brassica juncea | Cd (0.6 mM) as CdCl2 | Soil treatment with CA (0 and 0.6 mM) | Significantly increased Chl a+b, carotenoid, and polyphenols. Non-significant increase in flavonoids, anthocyanins and total carbohydrate content. Induced stomatal opening. Reduced ROS production. | Alleviated Cd stress. | [33] |
| Brassica napus | Cr (100 and 500 μM) | Irrigated with CA (2.5 and 5.0 mM) | Increased plant growth, biomass, Chl a, Chl b, Chl a+b, carotenoid, and soluble protein concentrations. Enhanced activities SOD, POX, CAT, and APX. Reduced MDA and EL. | Improved Cr stress tolerance. | [58] |
| Brassica juncea | Cd (0.5 mM Cd and 1.0 mM CdCl2) | CA (0.5 and 1.0 mM) in nutrient solution | Increased plant growth, leaf RWC, and Chl content. Enhanced activities of APX, MDHAR, DHAR, GR, GPX, SOD, and CAT. Reduced oxidative damage. | Enhanced Cd stress tolerance by regulating antioxidant defense. | [53] |
| Helianthus annuus (Sunflower) | Cr (5, 10 and 20 mg kg−1 dry weight) | CA treatment (2.5 and 5.0 mM) | Increased plant growth and biomass, Chl, carotenoid, photosynthesis, gas exchange, and soluble proteins. Enhanced activities of antioxidant enzymes. Reduced production of ROS and MDA. | Improved Cr stress tolerance. | [59] |
| Juncus effusus | Mn (50, 100 and 500 μM as MnSO4) | CA (5 mM) in the nutrient solution | Increased shoot length and root number. | Alleviated Mn toxicity and enhanced growth. | [71] |
| Germinating pea seeds | Cu (as 200 µM CuCl2) | Irrigated with CA (as 100 µM Na-citrate) | Reduced oxidative stress. Decreased H2O2, MDA, carbonyl groups, lipid peroxidation, and protein oxidation. | Enhanced growth and reduced stress. | [67] |
| Zea mays (Maize) | Cd as CdCl2 (300 mg kg−1) | Irrigation with CA (0.25, 0.5, 1.0 and 2 g kg−1 soil) | Increased root and shoot length, biomass. Reduced bioaccumulation coefficient and translocation factor. Reduced Cd uptake. | CA proved inefficient for Cd phytoextraction, however, ameliorated the toxicity of Cd | [63] |
| Brassica juncea | Cd (150 mg Cd2+ kg−1 soil) | CA (10 and 20 mmol kg−1 soil) | Increased shoot phenolic acids. Reduced ROS production. | Improved Cd stress tolerance. | [66] |
| Brassica napus | Pb as Pb(NO3)2 (50 and 100 μM) | CA (2.5 mM) in solution media | Increased plant height, root length, leaf growth, fresh and dry weight, Chl content, SPAD values, Pn, E, Gs, and Pn/E. Enhanced SOD, POX, CAT, and APX activities. Prevented lipid membrane damage. Reduced MDA and H2O2 production. | Increased Pb stress tolerance. | [57] |
| Solanum lycopersicum | Pb (10 μM as Pb(NO3)2) and As (10 μM as Na2HAsO4) | CA (250 μM) in nutrient solution | Increased Chl a and Chl b content. Decreased Pb accumulation, α-tocopherol content, and MDA levels. | Increased Pb and As tolerance. | [69] |
| Roots of Vicia faba | Pb (5 μM) as Pb(NO3)2 | CA (550 μM and 1000 μM) in nutrient culture | Non-significant effect on antioxidant enzyme activities (i.e., SOD, GPX, APX, and GR). | CA did not mitigate Pb toxicity | [73] |
| Sedum alfredii | Cd (100 µmol L−1 CdCl2) | CA (0, 10, 50, 100, 500 µmol L−1) in solution culture | Increased plant growth and biomass. | Improved Cd stress tolerance | [74] |
| Corchorus olitorius | Cd (20 mg L−1) as Cd(NO3)2. 4H2O | 5 mM CA in nutrient culture | Enhanced antioxidant enzyme activity. Decreased Cd2+ uptake and accumulation. | Improved Cd stress tolerance | [62] |
| Salix variegate | Cd (50 μmol L−1) as CdCl2·2. 5H2O | CA (100 μmol L−1) in nutrient solution | Increased biomass, carotenoid, Chl a, Chl b and Chl a+b content. Increased net photosynthesis rate, stomatal conductance, chloroplast size and width. | Reduced stress and enhanced growth, biomass, and photosynthesis. | [55] |
| Brassica juncea | Ni as NiSO4 (0.003 mmol L−1) | CA (0.5, 1.0, and 5.0 mmol L−1) in nutrient solution | Reduced Ni uptake but had no effect on Ni translocation. | Reduce stress by reducing Ni uptake. | [65] |
| Brassica juncea | Cd (0.6 mM) | Foliar spray of CA (0.6 mM) | Increased plant growth. Increased antioxidant activity. Reduced ROS. | Enhanced growth and efficacy of photosynthetic machinery | [61] |
| Helianthus annuus (Sunflower) | Cr (5, 10, and 20 mg kg−1) | Irrigation with CA (2.5 and 5 mM) | Increased plant growth, Chl, carotenoid, Pn, E, Gs, and water use efficiency. | Increased tolerance to Cr stress. | [40] |
| Larix olgensis | 100 mg kg−1 Pb from Pb(NO3)2 | Root irrigation and foliar spraying of CA (0.2, 1.0, 5.0, and 10.0 mmol L−1) | Increased plant growth and biomass, proline, total Chl, and carotenoid content. Enhanced SOD and POX activities. Reduced Pb content and MDA levels. | Improved tolerance to Pb stress | [54] |
| Oryza sativa (Rice) | Cd as CdCl2 (25.0 µM) | CA (50.0 µM) in nutrient solution | Increased GSH, Chl, carotenoid, and anthocyanin contents. Decreased Cd content in leaves. | Enhanced Cd tolerance and promoted higher biomass production | [60] |
| Triticum aestivum (Wheat) | 20 µM Cd (added as CdCl2) | Irrigation with CA (10, 50, 100, and 500 µM) | Increased index of tolerance, root and shoot biomass. Decreased Cd uptake, MDA levels, and PCs-SH production in roots. | Reduced bioavailability of Cd. | [64] |
| Medicago sativa (Alfalfa) | 100 µM Al in nutrient solution | Foliar spraying with 100 µM of CA | Increased growth. Reduced lipid peroxidation. | Alleviated Al toxicity through roots Al detoxification | [75] |
| Typha latifolia | Pb and Hg (1, 2.5 and 5 mM) | CA (5 mM) in nutrient medium | Increased fresh and dry biomass of root, stem, and leaf. Increased Chl a, Chl b, Chl a+b, carotenoid, soluble protein contents, and SPAD values. Decreased ROS, MDA, and EL. Enhanced the activities of SOD, POX, APX, and CAT. | Improved stress tolerance with increased physiological parameters. | [72] |
| Gene(s) | Origins | Transgenic Plants | Phenotype | References |
|---|---|---|---|---|
| Citrate Synthase (CS) | Pseudomonas aeruginosa | Nicotiana tabacum | Al stress tolerance | [120] |
| CS | Pseudomonas aeruginosa | Papaya sp. | Al stress tolerance | [120] |
| CS | Pseudomonas aeruginosa | Tobacco plant | Al stress intolerance | [124] |
| AACT1 | Hordeum vulgare | Tobacco cells | Al stress tolerance | [136] |
| MATE | Sorghum bicolor | Arabidopsis thaliana | Al stress tolerance | [137] |
| CS | Pseudomonas aeruginosa | Medicago sativa | Al stress tolerance | [141] |
| CS | Citrus junos | Nicotiana benthamiana | Al stress tolerance | [142] |
| MATE1 | Zea mays | Arabidopsis thaliana | Al stress tolerance | [132] |
| MATE | Vigna umbellate | Solanum lycopersicum | Al stress tolerance | [134] |
| MATE | Brachypodium distachyon | Setaria viridis | Al stress tolerance | [138] |
| Mitochondrial Citrate Synthase (mtCS) | Arabidopsis thaliana | Daucus carota | Al stress tolerance | [121] |
| mtCS | Arabidopsis thaliana | Brassica napus | Al stress tolerance | [125] |
| TIFY10a | Glycine soja | Medicago sativa | Alkaline stress tolerance | [139] |
| PPCK3 | Glycine soja | Medicago sativa | Alkaline stress tolerance | [140] |
| CS1 | Malus xiaojinensis | Nicotiana tabacum | Fe stress tolerance | [123] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tahjib-Ul-Arif, M.; Zahan, M.I.; Karim, M.M.; Imran, S.; Hunter, C.T.; Islam, M.S.; Mia, M.A.; Hannan, M.A.; Rhaman, M.S.; Hossain, M.A.; et al. Citric Acid-Mediated Abiotic Stress Tolerance in Plants. Int. J. Mol. Sci. 2021, 22, 7235. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22137235
Tahjib-Ul-Arif M, Zahan MI, Karim MM, Imran S, Hunter CT, Islam MS, Mia MA, Hannan MA, Rhaman MS, Hossain MA, et al. Citric Acid-Mediated Abiotic Stress Tolerance in Plants. International Journal of Molecular Sciences. 2021; 22(13):7235. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22137235
Chicago/Turabian StyleTahjib-Ul-Arif, Md., Mst. Ishrat Zahan, Md. Masudul Karim, Shahin Imran, Charles T. Hunter, Md. Saiful Islam, Md. Ashik Mia, Md. Abdul Hannan, Mohammad Saidur Rhaman, Md. Afzal Hossain, and et al. 2021. "Citric Acid-Mediated Abiotic Stress Tolerance in Plants" International Journal of Molecular Sciences 22, no. 13: 7235. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22137235