
Increased Placental Cell Senescence and Oxidative Stress in Women with Pre-Eclampsia and Normotensive Post-Term Pregnancies

 and
and
Abstract
:1. Introduction
2. Results
2.1. Participants
2.2. Gene Expression
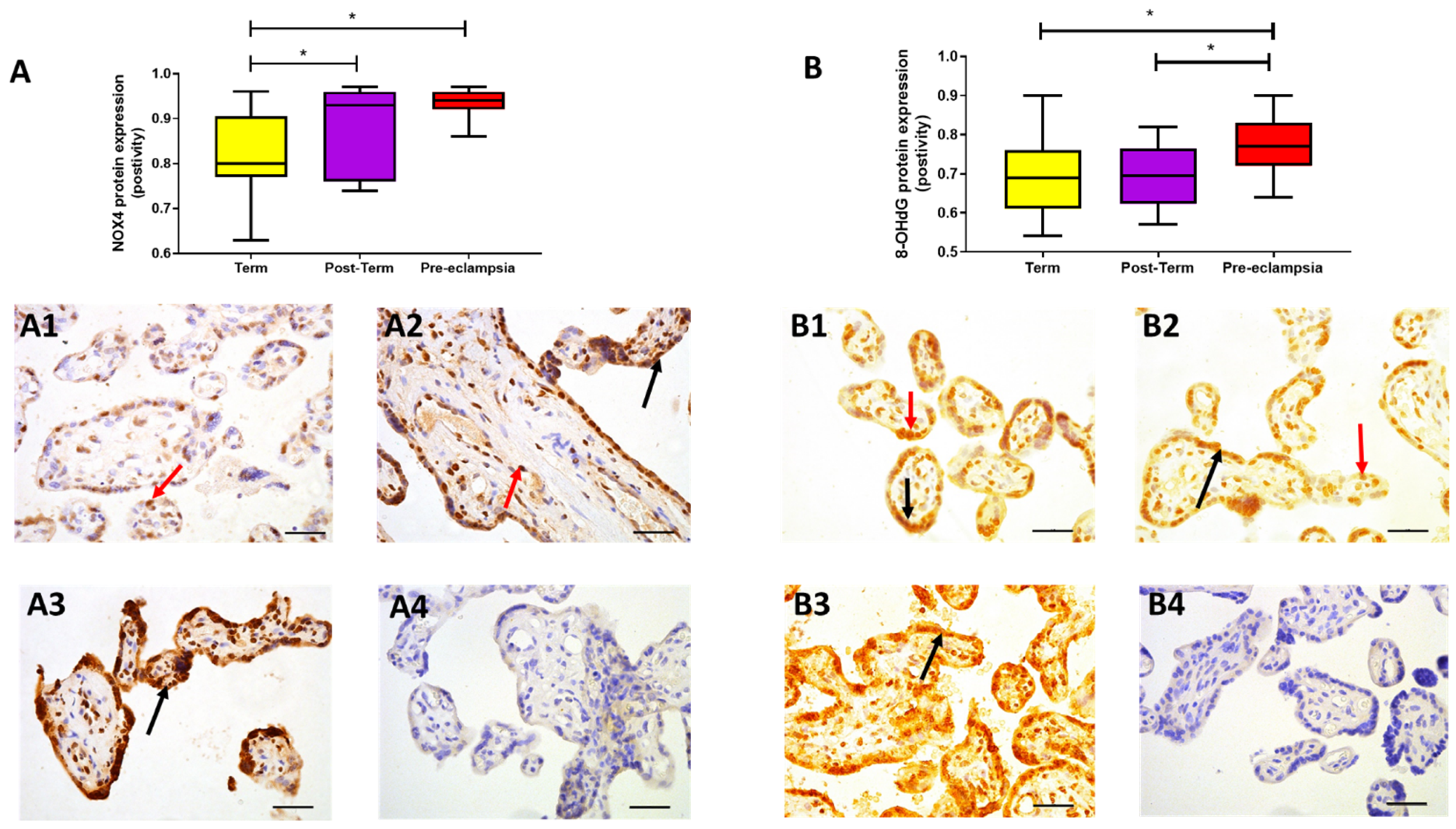
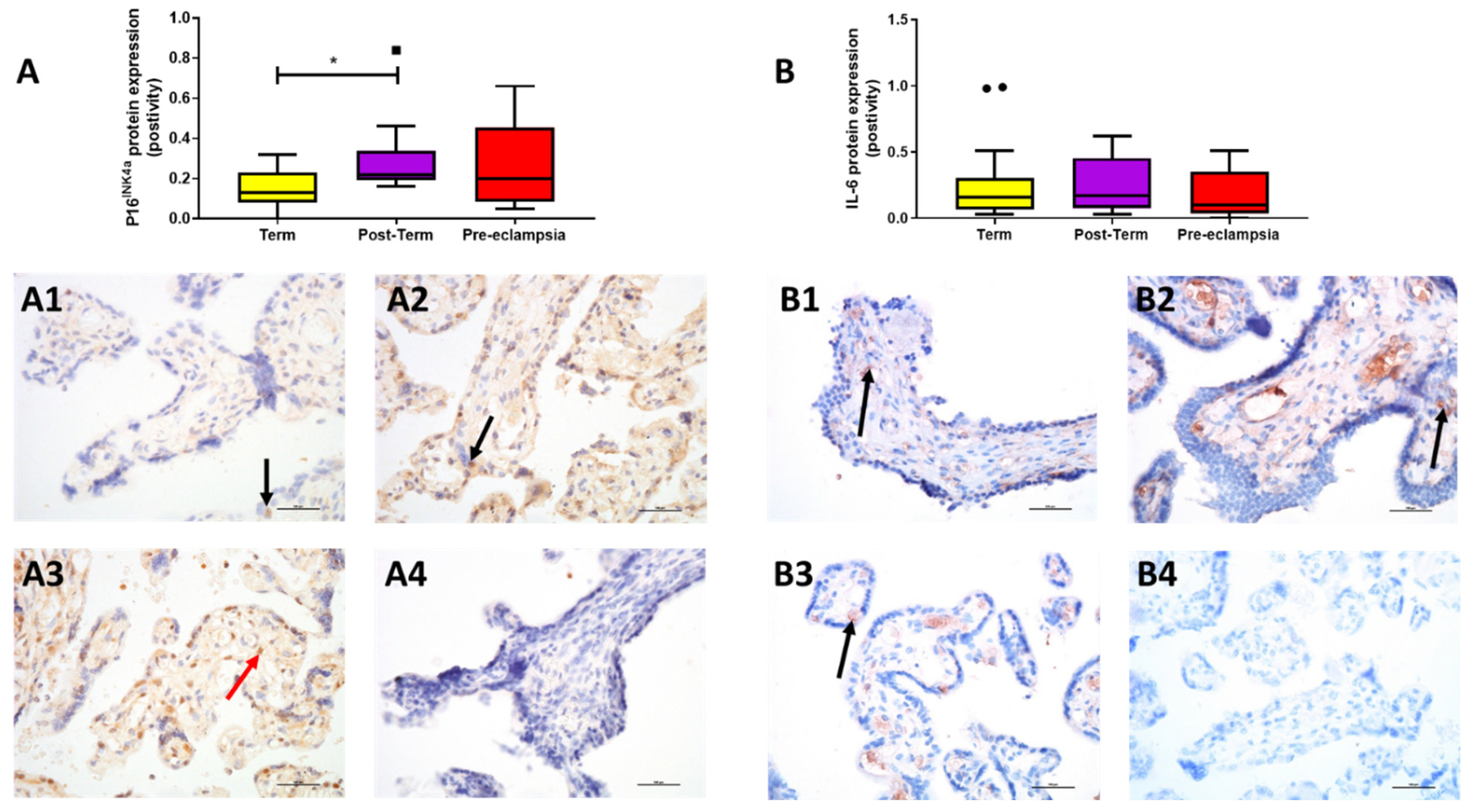
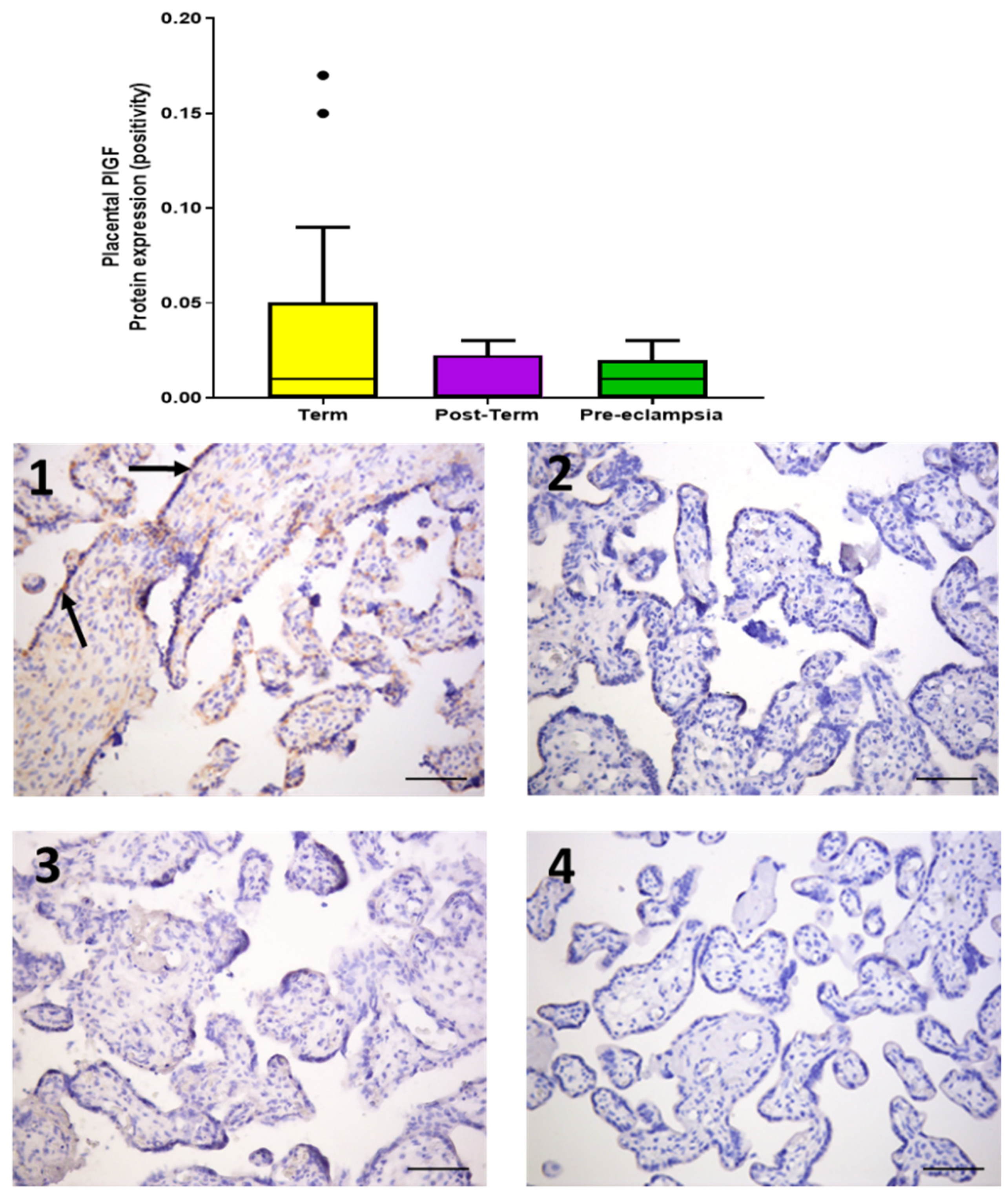
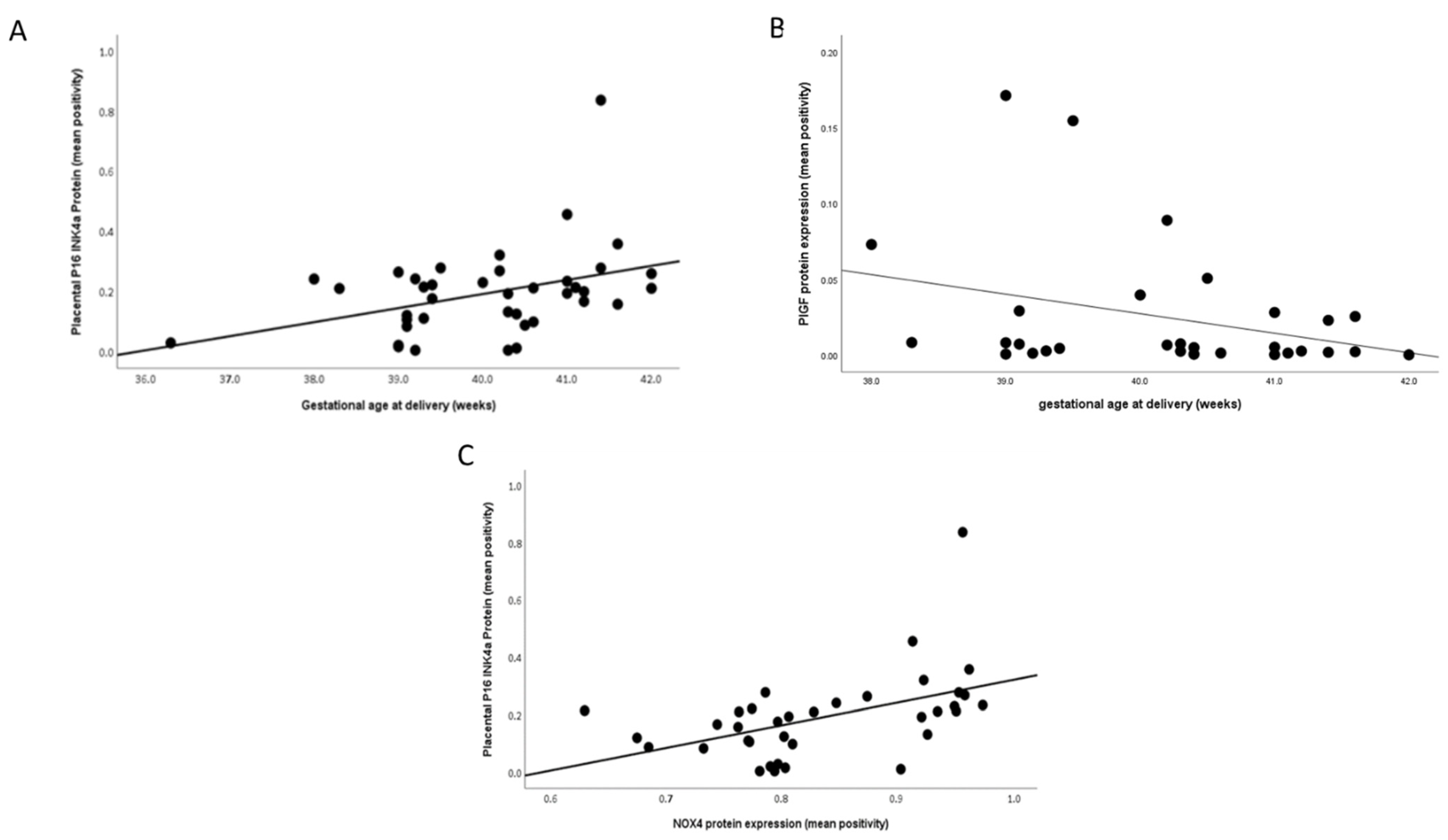
2.3. Protein Expression
3. Discussion
4. Materials and Methods
4.1. Cohort and Sample Collection
4.2. RNA Extraction, cDNA Synthesis, and Quantitative Reverse-Transcription Polymerase Chain Reaction (RT-qPCR)
4.3. Immunohistochemical Staining
4.4. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sultana, Z.; Maiti, K.; Aitken, J.; Morris, J.; Dedman, L.; Smith, R. Oxidative stress, placental ageing-related pathologies and adverse pregnancy outcomes. Am. J. Reprod. Immunol. 2017, 77, e12653. [Google Scholar] [CrossRef] [Green Version]
- van Deursen, J.M. The role of senescent cells in ageing. Nature 2014, 509, 439–446. [Google Scholar] [CrossRef] [Green Version]
- Kumari, R.; Jat, P. Mechanisms of Cellular Senescence: Cell Cycle Arrest and Senescence Associated Secretory Phenotype. Front. Cell Dev. Biol. 2021, 9, 645593. [Google Scholar] [CrossRef]
- WHO. Recommended definitions, terminology and format for statistical tables related to the perinatal period and use of a new certificate for cause of perinatal deaths. Modifications recommended by FIGO as amended October 14, 1976. Acta Obstet. Gynecol. Scand. 1977, 56, 247–253. [Google Scholar]
- Ayyavoo, A.; Derraik, J.G.; Hofman, P.L.; Cutfield, W.S. Postterm births: Are prolonged pregnancies too long? J. Pediatrics 2014, 164, 647–651. [Google Scholar] [CrossRef]
- Wennerholm, U.B.; Saltvedt, S.; Wessberg, A.; Alkmark, M.; Bergh, C.; Wendel, S.B.; Fadl, H.; Jonsson, M.; Ladfors, L.; Sengpiel, V.; et al. Induction of labour at 41 weeks versus expectant management and induction of labour at 42 weeks (SWEdish Post-term Induction Study, SWEPIS): Multicentre, open label, randomised, superiority trial. BMJ 2019, 367, l6131. [Google Scholar] [CrossRef] [Green Version]
- NICE. Inducing Labour; NICE Public Health Guidance: London, UK, 2008. [Google Scholar]
- Galal, M.; Symonds, I.; Murray, H.; Petraglia, F.; Smith, R. Postterm pregnancy. Facts Views Vis. Obgyn. 2012, 4, 175–187. [Google Scholar]
- Oberg, A.S.; Frisell, T.; Svensson, A.C.; Iliadou, A.N. Maternal and fetal genetic contributions to postterm birth: Familial clustering in a population-based sample of 475,429 Swedish births. Am. J. Epidemiol. 2013, 177, 531–537. [Google Scholar] [CrossRef] [Green Version]
- Harding, M. Post-term Prenancy. In Patient; NICE: London, UK, 2016. [Google Scholar]
- Jones, C.J.; Fox, H. Ultrastructure of the placenta in prolonged pregnancy. J. Pathol. 1978, 126, 173–179. [Google Scholar] [CrossRef]
- Maiti, K.; Sultana, Z.; Aitken, R.J.; Morris, J.; Park, F.; Andrew, B.; Riley, S.C.; Smith, R. Evidence that fetal death is associated with placental aging. Am. J. Obstet. Gynecol. 2017, 217, 441.e1–441.e14. [Google Scholar] [CrossRef] [Green Version]
- Cindrova-Davies, T.; Fogarty, N.M.E.; Jones, C.J.P.; Kingdom, J.; Burton, G.J. Evidence of oxidative stress-induced senescence in mature, post-mature and pathological human placentas. Placenta 2018, 68, 15–22. [Google Scholar] [CrossRef]
- Steegers, E.A.; von Dadelszen, P.; Duvekot, J.J.; Pijnenborg, R. Pre-eclampsia. Lancet 2010, 376, 631–644. [Google Scholar] [CrossRef]
- Brown, M.A.; Magee, L.A.; Kenny, L.C.; Karumanchi, S.A.; McCarthy, F.P.; Saito, S.; Hall, D.R.; Warren, C.E.; Adoyi, G.; Ishaku, S.; et al. The hypertensive disorders of pregnancy: ISSHP classification, diagnosis & management recommendations for international practice. Pregnancy Hypertens 2018, 13, 291–310. [Google Scholar] [PubMed]
- Ives, C.W.; Sinkey, R.; Rajapreyar, I.; Tita, A.T.N.; Oparil, S. Preeclampsia-Pathophysiology and Clinical Presentations: JACC State-of-the-Art Review. J. Am. Coll. Cardiol. 2020, 76, 1690–1702. [Google Scholar] [CrossRef] [PubMed]
- Carnero, A. Markers of Cellular Senescence. In Cell Senescence Methods and Protocols; Galluzzi, L., Vitala, I., Kepp, O., Kroemer, G., Eds.; Humana Press: Totowa, NJ, USA, 2013. [Google Scholar]
- Yudkin, P.L.; Wood, L.; Redman, C.W. Risk of unexplained stillbirth at different gestational ages. Lancet 1987, 1, 1192–1194. [Google Scholar] [CrossRef]
- Ferrari, F.; Facchinetti, F.; Saade, G.; Menon, R. Placental telomere shortening in stillbirth: A sign of premature senescence? J. Matern. Fetal Neonatal Med. 2016, 29, 1283–1288. [Google Scholar] [CrossRef]
- Smith, R.; Maiti, K.; Aitken, R.J. Unexplained antepartum stillbirth: A consequence of placental aging? Placenta 2013, 34, 310–313. [Google Scholar] [CrossRef]
- Williams, P.J.; Mistry, H.D.; Innes, B.A.; Bulmer, J.N.; Broughton Pipkin, F. Expression of AT1R, AT2R and AT4R and their roles in extravillous trophoblast invasion in the human. Placenta 2010, 31, 448–455. [Google Scholar] [CrossRef]
- Hirano, T. IL-6 in inflammation, autoimmunity and cancer. Int. Immunol. 2021, 33, 127–148. [Google Scholar] [CrossRef] [PubMed]
- Hannan, K.M.; Brandenburger, Y.; Jenkins, A.; Sharkey, K.; Cavanaugh, A.; Rothblum, L.; Moss, T.; Poortinga, G.; McArthur, G.A.; Pearson, R.B.; et al. mTOR-dependent regulation of ribosomal gene transcription requires S6K1 and is mediated by phosphorylation of the carboxy-terminal activation domain of the nucleolar transcription factor UBF. Mol. Cell Biol. 2003, 23, 8862–8877. [Google Scholar] [CrossRef] [Green Version]
- Hara, K.; Yonezawa, K.; Kozlowski, M.T.; Sugimoto, T.; Andrabi, K.; Weng, Q.P.; Kasuga, M.; Nishimoto, I.; Avruch, J. Regulation of eIF-4E BP1 phosphorylation by mTOR. J. Biol. Chem. 1997, 272, 26457–26463. [Google Scholar] [CrossRef] [Green Version]
- Chuprin, A.; Gal, H.; Biron-Shental, T.; Biran, A.; Amiel, A.; Rozenblatt, S.; Krizhanovsky, V. Cell fusion induced by ERVWE1 or measles virus causes cellular senescence. Genes Dev. 2013, 27, 2356–2366. [Google Scholar] [CrossRef] [Green Version]
- Cox, L.S.; Redman, C. The role of cellular senescence in ageing of the placenta. Placenta 2017, 52, 139–145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Londero, A.P.; Orsaria, M.; Marzinotto, S.; Grassi, T.; Fruscalzo, A.; Calcagno, A.; Bertozzi, S.; Nardini, N.; Stella, E.; Lelle, R.J.; et al. Placental aging and oxidation damage in a tissue micro-array model: An immunohistochemistry study. Histochem. Cell Biol. 2016, 146, 191–204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sukenik-Halevy, R.; Amiel, A.; Kidron, D.; Liberman, M.; Ganor-Paz, Y.; Biron-Shental, T. Telomere homeostasis in trophoblasts and in cord blood cells from pregnancies complicated with preeclampsia. Am. J. Obstet Gynecol. 2016, 214, 283.e1–283.e7. [Google Scholar] [CrossRef]
- Biron-Shental, T.; Kidron, D.; Sukenik-Halevy, R.; Goldberg-Bittman, L.; Sharony, R.; Fejgin, M.D.; Amiel, A. TERC telomerase subunit gene copy number in placentas from pregnancies complicated with intrauterine growth restriction. Early Hum. Dev. 2011, 87, 73–75. [Google Scholar] [CrossRef] [PubMed]
- Biron-Shental, T.; Sukenik-Halevy, R.; Sharon, Y.; Goldberg-Bittman, L.; Kidron, D.; Fejgin, M.D.; Amiel, A. Short telomeres may play a role in placental dysfunction in preeclampsia and intrauterine growth restriction. Am. J. Obstet. Gynecol. 2010, 202, 381.e1–381.e7. [Google Scholar] [CrossRef]
- Biron-Shental, T.; Sukenik-Halevy, R.; Sharon, Y.; Laish, I.; Fejgin, M.D.; Amiel, A. Telomere shortening in intra uterine growth restriction placentas. Early Hum. Dev. 2014, 90, 465–469. [Google Scholar] [CrossRef]
- Levine, R.J.; Maynard, S.E.; Qian, C.; Lim, K.H.; England, L.J.; Yu, K.F.; Schisterman, E.F.; Thadhani, R.; Sachs, B.P.; Epstein, F.H.; et al. Circulating angiogenic factors and the risk of preeclampsia. N. Engl. J. Med. 2004, 350, 672–683. [Google Scholar] [CrossRef] [Green Version]
- Redman, C.W.; Staff, A.C. Preeclampsia, biomarkers, syncytiotrophoblast stress, and placental capacity. Am. J. Obstet. Gynecol. 2015, 213 (Suppl. 4), S9–S11. [Google Scholar] [CrossRef]
- Redman, C.W.; Sargent, I.L.; Staff, A.C. IFPA Senior Award Lecture: Making sense of pre-eclampsia—two placental causes of preeclampsia? Placenta 2014, 35, S20–S25. [Google Scholar] [CrossRef] [PubMed]
- ACOG. Definition of Term Pregnancy, Committee Opinion No. 579; ACOG: New Orleans, LA, USA, 2013; pp. 1139–1140. [Google Scholar]
- Brown, M.A.; Lindheimer, M.D.; de Swiet, M.; Van Assche, A.; Moutquin, J.M. The classification and diagnosis of the hypertensive disorders of pregnancy: Statement from the International Society for the Study of Hypertension in Pregnancy (ISSHP). Hypertens Pregnancy 2001, 20, IX–XIV. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mistry, H.D.; Wilson, V.; Ramsay, M.M.; Symonds, M.E.; Broughton Pipkin, F. Reduced selenium concentrations and glutathione peroxidase activity in pre-eclamptic pregnancies. Hypertension 2008, 52, 881–888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mistry, H.D.; McCallum, L.A.; Kurlak, L.O.; Greenwood, I.A.; Broughton Pipkin, F.; Tribe, R.M. Novel expression and regulation of voltage-dependent potassium channels in placentas from women with preeclampsia. Hypertension 2011, 58, 497–504. [Google Scholar] [CrossRef] [Green Version]
- Murthi, P.; Fitzpatrick, E.; Borg, A.J.; Donath, S.; Brennecke, S.P.; Kalionis, B. GAPDH, 18S rRNA and YWHAZ are suitable endogenous reference genes for relative gene expression studies in placental tissues from human idiopathic fetal growth restriction. Placenta 2008, 29, 798–801. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Term 37–39+6 (n = 26) | Post-Term 41–42 (n = 11) | Pre-Eclampsia (n = 13) |
|---|---|---|---|
| Age at booking (year) | 31 ± 6.0 | 31 ± 6.4 | 34 ± 3.8 |
| BMI at booking (kg/m2) | 26 [23,32] | 27 [26,31] | 30 [26,31] |
| Nulliparous, n (%) | 10 (38) | 5 (45) | 8 (62) |
| Smoking, n (%) | 5 (19) | 3 (27) | 1 (8) |
| Systolic blood pressure (mmHg) | 110 ± 10.6 c | 118 ± 5.1 c | 158 ± 12.3 c |
| Diastolic blood pressure (mmHg) | 77 ± 7.8 c | 75 ± 6.0 c | 97 ± 6.8 c |
| Proteinuria | - | - | 1.0 [0.4, 1.4] |
| Gestational age at delivery (weeks) | 39.4 (36.3–40.6) a,c | 41.3 (41.0–42) a,b | 37.2 (31.1–41.1) b,c |
| Birth weight (g) | 3533 ± 378 a | 3873 ± 384 b | 2741 ± 1073.7 a,b |
| Birthweight centile | 74 [47,94] | 86 [65,94] | 48 [26,78] |
| Placental weight (g) | 624 ± 133.6 | 696 ± 92.6 b | 539 ± 182.9 b |
| Baby gender, female n (%) | 14 (54) | 5 (45) | 8 (62) |
| Caesarean Section n (%) | 19 (73) | 11 (73) | 8 (77) |
| Gene | Accession Number | Primers | Length (bp) |
|---|---|---|---|
| Nox4 | NM_016931.3 | 5′–TGAACTATGAGGTCAGCCTCTG–3′ 5′–TCTCACGAATCTCCTCATGGT–3′ | 107 |
| GPx1 | NM_201397 | 5′–CAGTCGGTGTATGCCTTCTCG–3′ 5′–GAGGGACGCCACATTCTCG–3′ | 105 |
| GPx3 | NM_002084 | 5′–GAGCTTGCACCATTCGGTCT–3′ 5′–GGGTAGGAAGGATCTCTGAGTTC–3′ | 94 |
| GPx4 | NM_001039847 | 5′–GAGGCAAGACCGAAGTAAACTAC–3′ 5′–CCGAACTGGTTACACGGGAA–3′ | 100 |
| P16INK4a | NM_000077.4 | 5′–CTTCGGCTGACTGGCTGG–3′ 5′–TCATCATGACCTGGATCGGC–3′ | 129 |
| P21 | NM_078467 | 5′–TGTCCGTCAGAACCCATGC–3′ 5′–AAAGTCGAAGTTCCATCGCTC–3′ | 139 |
| IL–6 | NM_000600 | 5′–ACTCACCTCTTCAGAACGAATTG–3′ 5′–CCATCTTTGGAAGGTTCAGGTTG–3′ | 149 |
| PlGF | NM_001207012 | 5′–GAACGGCTCGTCAGAGGTG–3′ 5′–ACAGTGCAGATTCTCATCGCC–3′ | 187 |
| sFlt–1 | NM_001159920 | 5′–TTTGCCTGAAATGGTGAGTAAGG–3′ 5′–TGGTTTGCTTGAGCTGTGTTC–3′ | 117 |
| B2M | NM_004048.2 | 5′–CTTATGCACGCTTAACTATCTTAACAA–3′ 5′–TAGGAGGGCTGGCAACTTAG–3′ | 127 |
| YWHAZ | NM_001135702.1 | 5′–ACTTTTGGTACATTGTGGCTTCAA–3′ 5′–CCGCCAGGACAAACCAGTAT–3′ | 94 |
| GAPDH | NM_002046.3 | 5′–GGAAGCTTGTCATCAATGGAA–3′ 5′–TGGACTCCACGACGTACTCA–3′ | 102 |
| Antigen | Suppler Information | Concentration (µg/mL) |
|---|---|---|
| NOX4 | Abcam, rabbit monoclonal: ab133303 | 2.18 |
| 8-0HdG | Abcam, mouse monoclonal: ab48508 | 12 |
| p16INK4α | Abcam, rabbit polyclonal: ab108349 | 0.7 |
| IL-6 | Abcam, mouse monoclonal: ab9324 | |
| PlGF | Abcam, rabbit polyclonal: ab196666 | 10 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Scaife, P.J.; Simpson, A.; Kurlak, L.O.; Briggs, L.V.; Gardner, D.S.; Broughton Pipkin, F.; Jones, C.J.P.; Mistry, H.D. Increased Placental Cell Senescence and Oxidative Stress in Women with Pre-Eclampsia and Normotensive Post-Term Pregnancies. Int. J. Mol. Sci. 2021, 22, 7295. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22147295
Scaife PJ, Simpson A, Kurlak LO, Briggs LV, Gardner DS, Broughton Pipkin F, Jones CJP, Mistry HD. Increased Placental Cell Senescence and Oxidative Stress in Women with Pre-Eclampsia and Normotensive Post-Term Pregnancies. International Journal of Molecular Sciences. 2021; 22(14):7295. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22147295
Chicago/Turabian StyleScaife, Paula J., Amy Simpson, Lesia O. Kurlak, Louise V. Briggs, David S. Gardner, Fiona Broughton Pipkin, Carolyn J. P. Jones, and Hiten D. Mistry. 2021. "Increased Placental Cell Senescence and Oxidative Stress in Women with Pre-Eclampsia and Normotensive Post-Term Pregnancies" International Journal of Molecular Sciences 22, no. 14: 7295. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22147295