
Bacterial Infection and Non-Hodgkin B-Cell Lymphoma: Interactions between Pathogen, Host and the Tumor Environment


Abstract
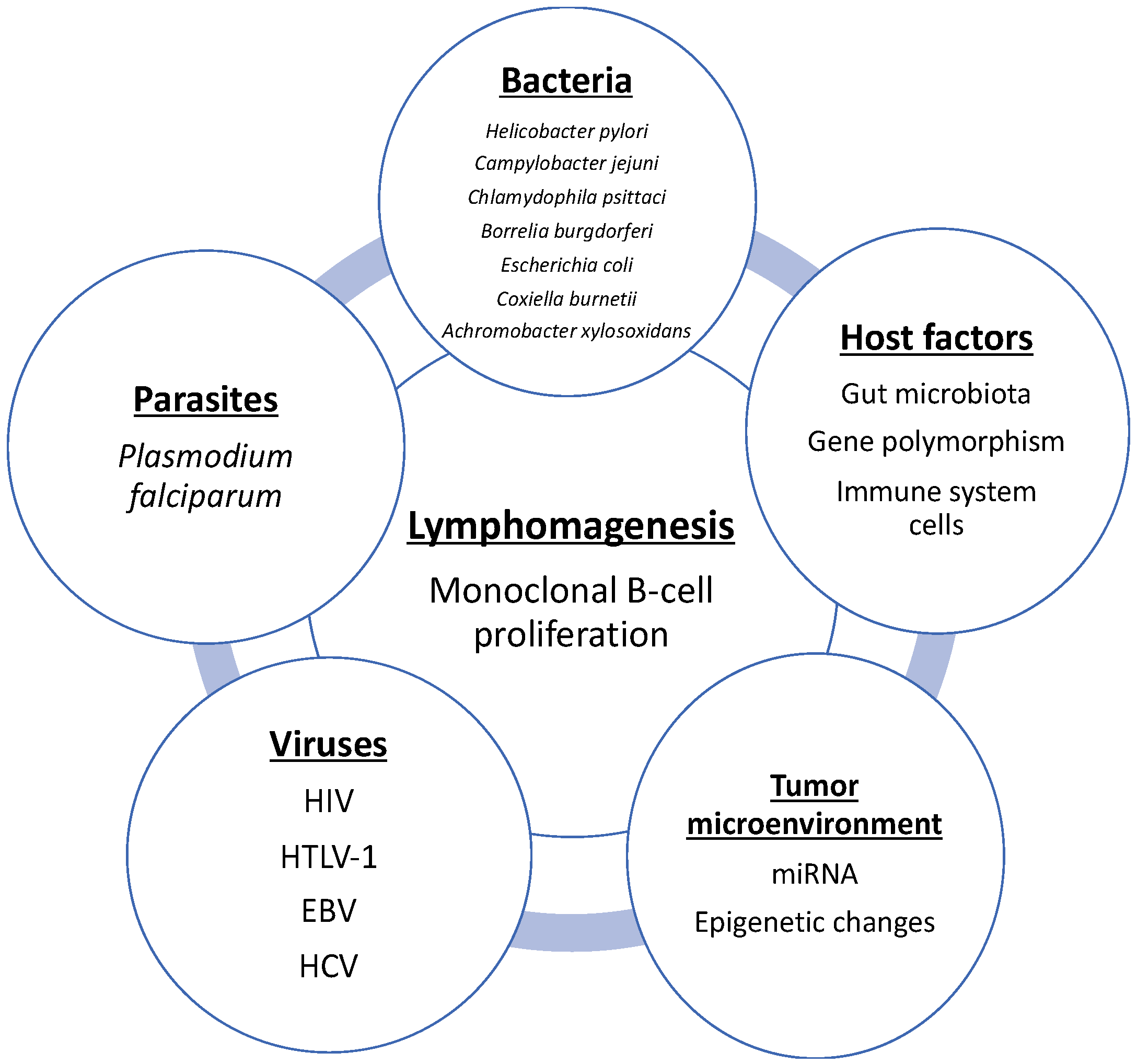
:1. Introduction
2. Role of Bacteria in Gastrointestinal Lymphoma
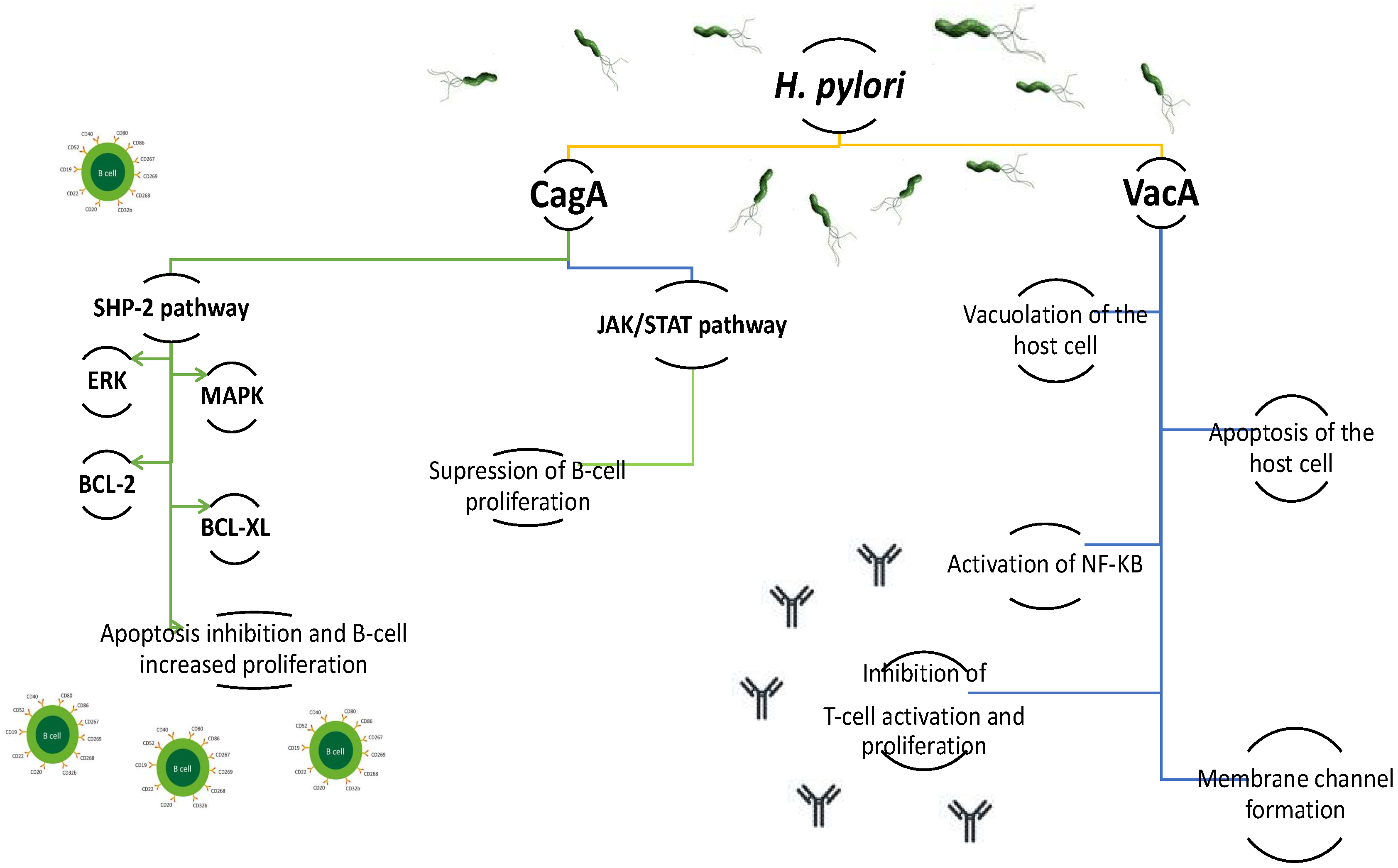
2.1. H. pylori and MALT, DLBCL, and Burkitt lymphoma
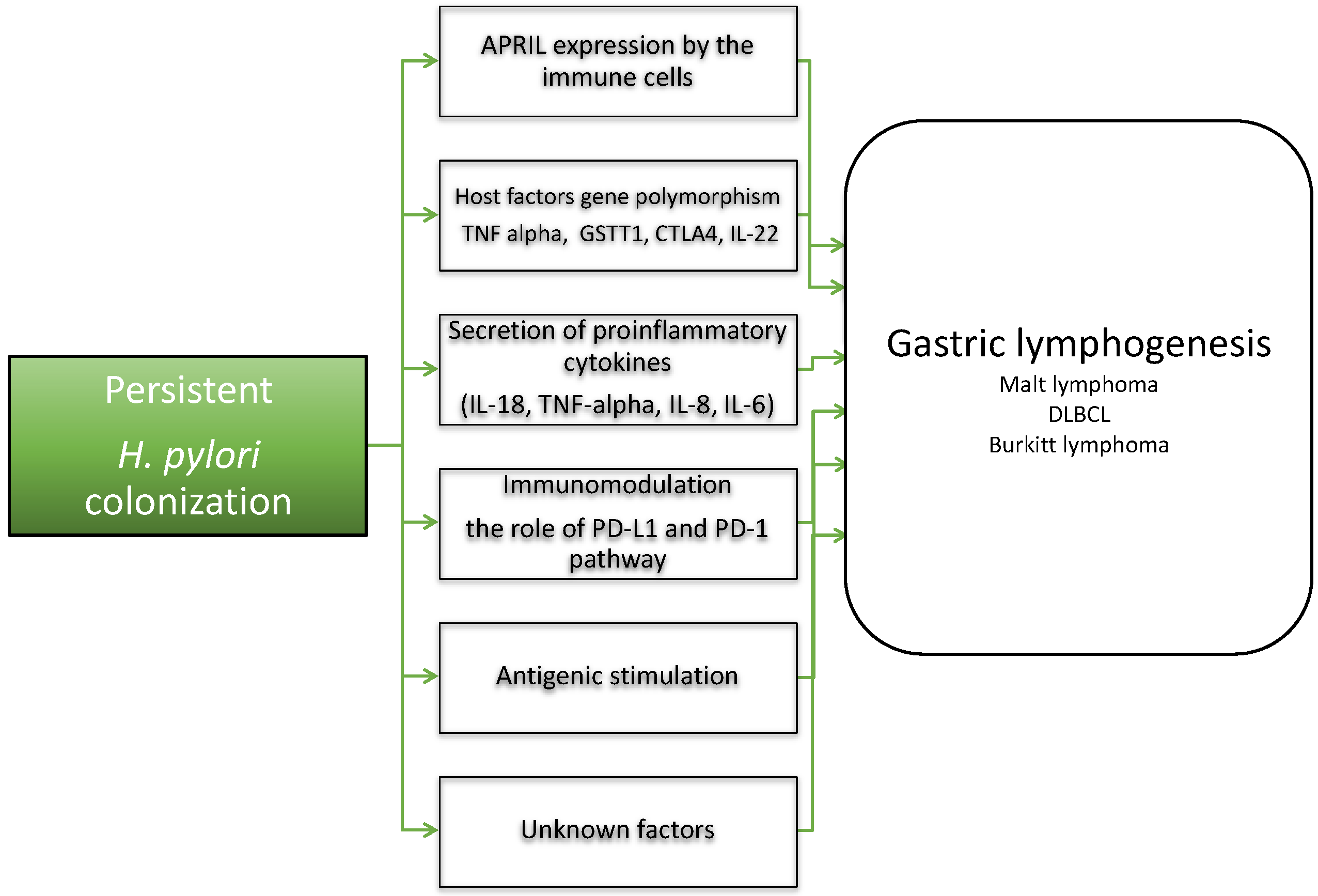
2.2. H. pylori and Host Factors in Gastric Lymphogenesis
2.3. The Role of Epigenetic Factors and Molecular Factors in Gastric Lymphogenesis
2.4. H. pylori Eradication in B-Cell Lymphomas
2.5. H. pylori and Gastric DLBCL
2.6. H. pylori and Burkitt Lymphoma
2.7. Campylobacter jejuni and IPSID
3. Role of Bacteria in Skin and Ocular NHL Lymphomas
3.1. Borrelia burgdorferi and Cutaneous NHL
3.2. Chlamydophila psittaci (Ch. psittaci) and Ocular MALT
4. Role of Bacteria in Pulmonary Lymphogenesis
Achromobacter xylosoxidans and BALT
5. Escherichia coli and Primary Bladder MALT
6. Coxiella burnetii (C. burnetii) and Various NHL
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pratap, S.; Scordino, T.S. Molecular and cellular genetics of non-Hodgkin lymphoma: Diagnostic and prognostic implications. Exp. Mol. Pathol. 2019, 106, 44–51. [Google Scholar] [CrossRef] [PubMed]
- Sindel, A.; Al-Juhaishi, T.; Yazbeck, V. Marginal Zone Lymphoma: State-of-the-Art Treatment. Curr. Treat. Options Oncol. 2019, 5, 90. [Google Scholar] [CrossRef]
- Swerdlow, S.H.; Campo, E.; Pileri, S.A.; Harris, N.L.; Stein, H.; Siebert, R.; Advani, R.; Ghielmini, M.; Salles, G.A.; Zelenetz, A.D.; et al. The 2016 revision of the World Health Organization classification of lymphoid neoplasms. Blood 2016, 127, 2375–2390. [Google Scholar] [CrossRef] [Green Version]
- Zucca, E.; Bertoni, F. The spectrum of MALT lymphoma at different sites: Biological and therapeutic relevance. Blood 2016, 127, 2082–2092. [Google Scholar] [CrossRef] [Green Version]
- Raderer, M.; Kiesewetter, B.; Ferreri, A.J. Clinicopathologic characteristics and treatment of marginal zone lymphoma of mucosa- associated lymphoid tissue (MALT lymphoma). CA Cancer J. Clin. 2016, 66, 153–171. [Google Scholar] [CrossRef] [PubMed]
- Laliani, G.; Ghasemian Sorboni, S.; Lari, R.; Yaghoubi, A.; Soleimanpour, S.; Khazaei, M.; Hasanian, S.M.; Avan, A. Bacteria and cancer: Different sides of the same coin. Life Sci. 2020, 1, 117398. [Google Scholar] [CrossRef] [PubMed]
- Rybojad, P.; Los, R.; Sawicki, M.; Tabarkiewicz, J.; Malm, A. Anaerobic bacteria colonizing the lower airways Anaerobic bacteria colonizing the lower airways in lung cancer patients in lung cancer patients. Folia Histochem Cytobiol. 2011, 49, 263–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karpiński, T.M. Role of Oral Microbiota in Cancer Development. Microorganisms 2019, 7, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levine, A.M. Lymphomas and leukemias due to infectious organisms. Hematology 2012, 17 (Suppl. 1), S87–S89. [Google Scholar] [CrossRef]
- Chiu, B.C.; Hou, N. Epidemiology and etiology of Non-Hodgkin lymphoma. Cancer Treat. Res. 2015, 165, 1–25. [Google Scholar]
- Suarez, F.; Lecuit, M. Infection-associated non-Hodgkin lymphomas. Clin. Microbiol. Infect. 2015, 21, 991–997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, S.; Vuai, M.S.; Zhong, M. The role of bacteria in cancer therapy—Enemies in the past, but allies at present. Infect. Agent Cancer 2018, 13, 9. [Google Scholar] [CrossRef]
- Stevens, W.B.; Netea, M.G.; Kater, A.P.; van der Velden, W.J. ‘Trained immunity’: Consequences for lymphoid malignancies. Haematologica 2016, 101, 1460–1468. [Google Scholar] [CrossRef] [Green Version]
- Melenotte, C.; Mezouar, S.; Mège, J.L.; Gorvel, J.P.; Kroemer, G.; Raoult, D. Bacterial infection and non-Hodgkin’s lymphoma. Crit. Rev. Microbiol. 2020, 46, 270–287. [Google Scholar] [CrossRef] [PubMed]
- Ferreri, A.J.; Ernberg, I.; Copie-Bergman, C. Infectious agents and lymphoma development: Molecular and clinical aspects. J. Intern. Med. 2009, 265, 421–438. [Google Scholar] [CrossRef] [PubMed]
- Andersen, L.P. Colonization and infection by Helicobacter pylori in humans. Helicobacter 2007, 12, 12–16. [Google Scholar] [CrossRef]
- Saleem, N.; Howden, C.W. Update on the management of Helicobacter pylori infection. Curr. Treat. Options Gastroenterol. 2020, 17, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Fischbach., W.; Malfertheiner, P. Helicobacter pylori infection. Dtsch. Arztebl. Int. 2018, 22, 429–436. [Google Scholar]
- Matysiak-Budnik, T.; Fabiani, B.; Hennequin, C.; Thieblemont, C.; Malamut, G.; Cadiot, G.; Bouché, O.; Ruskoné-Fourmestraux, A. Gastrointestinal lymphomas: French Intergroup clinical practice recommendations for diagnosis, treatment and follow-up (SNFGE, FFCD, GERCOR, UNICANCER, SFCD, SFED, SFRO, SFH). Dig. Liver Dis. 2018, 50, 124–131. [Google Scholar] [CrossRef] [PubMed]
- Floch, P.; Magraud, F.; Lehours, P. Helicobacter pylori strains and gastric MALT Lymphoma. Toxins 2017, 9, 132. [Google Scholar] [CrossRef] [PubMed]
- Pellicano, R.; Ianiro, G.; Fagoonee, S.; Settanni, C.R.; Gasbarrini, A. Extragastric diseases and Helicobacter pylori. Helicobacter 2020, 25 (Suppl. 1), e12741. [Google Scholar] [CrossRef]
- Nakamura, S.; Matsumoto, T. Helicobacter pylori and gastric mucosa-associated lymphoid tissue lymphoma: Recent progress in pathogenesis and management. World J. Gastroenterol. 2013, 7, 8181–8187. [Google Scholar] [CrossRef]
- Denic, M.; Touati, E.; De Reuse, H. Pathogenesis of Helicobacter pylori infection. Helicobacter 2020, 25 (Suppl. 1), e12736. [Google Scholar] [CrossRef] [PubMed]
- Kuo, S.H.; Chen, L.T.; Lin, C.W.; Wu, M.S.; Hsu, P.N.; Tsai, H.J.; Chu, C.Y.; Tzeng, Y.S.; Wang, H.P.; Yeh, K.H.; et al. Detection of the Helicobacter pylori CagA protein in gastric mucosa-associated lymphoid tissue lymphoma cells: Clinical and biological significance. Blood Cancer J. 2013, 3, e125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Backert, S.; Tegtmeyer, N. Type IV secretion and signal transduction of Helicobacter pylori CagA through interactions with host cell receptors. Toxins 2017, 9, 115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuo, S.H.; Wu, M.S.; Yeh, K.H.; Lin, C.W.; Hsu, P.N.; Chen, L.T.; Cheng, A.L. Novel insights of lymphomagenesis of Helicobacter pylori-dependent gastric mucosa-associated lymphoid tissue lymphoma. Cancers 2019, 11, 547. [Google Scholar] [CrossRef] [Green Version]
- Kuo, S.H.; Yeh, K.H.; Chen, L.T.; Lin, C.W.; Hsu, P.N.; Wu, M.S.; Liou, J.M.; Tsai, H.J.; Tzeng, Y.S.; Cheng, A.L. Helicobacter pylori CagA translocation is closely associated with the expression of CagA-signaling molecules in low-grade gastric mucosa-associated lymphoid tissue lymphoma. Am. J. Surg Pathol. 2015, 39, 761–766. [Google Scholar] [CrossRef]
- Delchier, J.C.; Lamarque, D.; Levy, M.; Tkoub, E.M.; Copie Bergman, C.; Deforges, L.; Chaumette, M.T.; Haioun, C. Helicobacter pylori and gastric lymphoma: High seroprevalence of CagA in diffuse large B-cell lymphoma but not in low-grade lymphoma of mucosa-associated lymphoid tissue type. Am. J. Gastroenterol. 2001, 96, 2324–2328. [Google Scholar] [CrossRef] [PubMed]
- Kuo, S.H.; Chen, L.T.; Lin, C.W.; Yeh, K.H.; Shun, C.T.; Tzeng, Y.S.; Liou, J.M.; Wu, M.S.; Hsu, P.N.; Cheng, A.L. Expressions of the CagA protein and CagA-signaling molecules predict Helicobacter pylori dependence of early-stage gastric DLBCL. Blood 2017, 129, 188–198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atherton, J.C.; Peek, R.M., Jr.; Tham, K.T.; Cover, T.L.; Blaser, M.J. Clinical and pathological importance of heterogeneity in vacA, the vacuolating cytotoxine gene of Helicobacter pylori. Gastroenterology 1997, 112, 92–99. [Google Scholar] [CrossRef]
- Papagiannakis, P.; Michalopoulos, C.; Papalexi, F.; Dalampoura, D.; Diamantidis, M.D. The role of Helicobacter pylori infection in hematological disorders. Eur. J. Intern. Med. 2013, 24, 685–690. [Google Scholar] [CrossRef]
- Luo, J.J.; Li, C.Y.; Liu, S.; Yu, W.; Tang, A.Y.; Cai, H.L.; Zhang, Y. Overexpression of Helicobacetr pylori VacA N-terminal fragment induces proinflammatory cytokine expression and apoptosis in human monocytic cell line through activation of NF-kB. Can. J. Microbiol. 2013, 59, 523–533. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, C.A.; Figueiredo, C.; Lic, C.B.; Ferreira, R.M.; Pardo, M.L.; Ruiz Liso, J.M.; Alonso, P.; Sala, N.; Sanz-Anquela, J.M. Helicobacter pylori cagA and vacA genotypes as predictors of progression of gastric preneoplastic lesions: A long-term follow-up in a high- rise area in Spain. Am. J. Gastroenterol. 2011, 106, 867–874. [Google Scholar] [CrossRef]
- Hasanzadeh, L.; Ghaznavi-Rad, E.; Soufian, S.; Farjadi, V.; Abtahi, H. Expression and antigenic evaluation of vacA antigenic fragment of Helicobacter pylori. Iran. J. Basic Med. Sci. 2013, 16, 835–840. [Google Scholar]
- de Brito, B.B.; da Silva, F.A.F.; Soares, A.S.; Pereira, V.A.; Santos, M.L.C.; Sampaio, M.M.; Neves, P.H.M.; de Melo, F.F. Pathogenesis and clinical management of Helicobacter pylori gastric infection. World J. Gastroenterol. 2019, 7, 5578–5589. [Google Scholar] [CrossRef] [PubMed]
- Burkitt, M.D.; Duckworth, C.A.; Williams, J.M.; Pritchard, D.M. Helicobacter pylori-induced gastric pathology: Insights from in vivo and ex vivo models. Dis. Model. Mech. 2017, 10, 89–104. [Google Scholar] [CrossRef] [Green Version]
- Abadi, A.T.B. Strategies used by Helicobacter pylori to establish persistent infection. World J. Gastroenterol. 2017, 23, 2870–2882. [Google Scholar] [CrossRef]
- Minaga, K.; Watanabe, T.; Kamata, K.; Asano, N.; Kudo, M. Nucleotide-binding oligomerization domain 1 and Helicobacter pylori infection: A review. World J. Gastroenterol. 2018, 24, 1725–1733. [Google Scholar] [CrossRef]
- Blosse, A.; Peru, S.; Levy, M.; Marteyn, B.; Floch, P.; Sifré, E.; Giese, A.; Prochazkova-Carlotti, M.; Azzi Martin, L.; Dubus, P.; et al. APRIL-producing eosinophils are involved in gastric MALT lymphomagenesis induced by Helicobacter sp infection. Sci. Rep. 2020, 9, 14858. [Google Scholar] [CrossRef]
- Barth, T.F.; Bentz, M.; Döhner, H.; Möller, P. Molecular aspects of B-cell lymphomas of the gastrointestinal tract. Clin. Lymphoma. 2001, 2, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Craig, V.J.; Cogliatti, S.B.; Arnold, I.; Gerke, C.; Balandat, J.E.; Wündisch, T.; Müller, A. B-cell receptor signaling and CD40 ligand-independent T cell help cooperate in Helicobacter-induced MALT lymphomagenesis. Leukemia 2010, 24, 1186–1196. [Google Scholar] [CrossRef] [Green Version]
- Munari, F.; Lonardi, S.M.; Cassatella, M.A.; Doglioni, C.; Cangi, M.G.; Amedei, A.; Facchetti, F.; Eishi, Y.; Rugge, M.; Fassan, M.; et al. Tumor-associated macrophages as major source of APRIL in gastric MALT lymphoma. Blood 2011, 117, 6612–6616. [Google Scholar] [CrossRef] [Green Version]
- Jubel, J.M.; Barbati, Z.R.; Burger, C.; Wirtz, D.C.; Schildberg, F.A. The role of PD-1 in acute and chronic infection. Front. Immunol. 2020, 11, 487. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.Y.; Chen, J.H.; Kao, J.T.; Liu, K.C.; Lai, C.H.H.; Wang, Y.M.; Hsieh, C.T.; Tzen, J.T.C.; Hsu, P.N. Expression of CD25(high) regulatory T cells and PD-1 in gastric infiltrating CD4(+) T lymphocytes in patients with Helicobacter pylori infection. Clin. Vaccine Immunol. 2011, 18, 1198–1201. [Google Scholar] [CrossRef] [Green Version]
- Shen, B.; Qian, A.; Lao, W.; Li, W.; Chen, X.; Zhang, B.; Wang, H.; Yuan, F.; Sun, Y. Relationship between Helicobacter pylori and expression of programmed death-1 and its ligand in gastric intraepithelial neoplasia and early-stage gastric cancer. Cancer Manag. Res. 2019, 11, 3909–3919. [Google Scholar] [CrossRef] [Green Version]
- Holokai, L.; Chakrabarti, J.; Broda, T.; Chang, J.; Hawkins, J.A.; Sundaram, N.; Wroblewski, L.E.; Peek, R.M., Jr.; Wang, J.; Helmrath, M.; et al. Increased programmed death-ligand 1 is an early epithelial cell response to Helicobacter pylori infection. PLoS Pathog. 2019, 15, e1007468. [Google Scholar] [CrossRef] [Green Version]
- Liao, F.; Hsu, Y.C.; Kuo, S.H.; Yang, Y.C.; Chen, J.P.; Hsu, P.N.; Lin, C.W.; Chen, L.T.; Cheng, A.L.; Fann, C.S.; et al. Genetic polymorphisms and tissue expression of interleukin-22 associated with risk and therapeutic response of gastric mucosa-associated lymphoid tissue lymphoma. Blood Cancer J. 2014, 4, eXX. [Google Scholar]
- Pereira, M.I.; Medeiros, J.A. Role of Helicobacter pylori in gastric mucosa associated lymphoid tissue lymphomas. World J. Gastroenterol. 2014, 20, 684–698. [Google Scholar] [CrossRef]
- Deutsch, A.J.; Steinbauer, E.; Hofmann, N.A.; Strunk, D.; Gerlza, T.; Beham-Schmid, C.; Schaider, H.; Neumeister, P. Chemokine receptors in gastric MALT lymphoma: Loss of CXCR4 and upregulation of CXCR7 is associated with progression to diffuse large B-cell lymphoma. Mod. Pathol. 2013, 26, 182–194. [Google Scholar] [CrossRef]
- Blosse, A.; Levy, M.; Robe, C.; Staedel, C.; Copie-Bergman, C.; Lehours, P. Deregulation of miRNA in Helicobacter pylori-induced gastric MALT lymphoma: From Mice to Human. J. Clin. Med. 2019, 8, 845. [Google Scholar] [CrossRef] [Green Version]
- Craig, V.J.; Cogliatti, S.B.; Rehrauer, H.; Wündisch, T.; Müller, A. Epigenetic silencing of microRNA-203 dysregulates ABL1 expression and drives Helicobacter-associated gastric lymphomagenesis. Cancer Res. 2011, 71, 3616–3624. [Google Scholar] [CrossRef] [Green Version]
- Saito, Y.; Suzuki, H.; Tsugawa, H.; Imaeda, H.; Matsuzaki, J.; Hirata, K.; Hosoe, N.; Nakamura, M.; Mukai, M.; Saito, H.; et al. Overexpression of miR-142-5p and miR-155 in gastric mucosa-associated lymphoid tissue (MALT) lymphoma resistant to Helicobacter pylori eradication. PLoS ONE 2012, 7, e47396. [Google Scholar] [CrossRef] [PubMed]
- Ferreri, A.J.; Govi, S.; Ponzoni, M. Marginal zone lymphomas and infectious agents. Semin. Cancer Biol. 2013, 23, 431–440. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Kim, K.M.; Kim, J.J.; Lee, J.H.; Rhee, J.C.; Ko, Y.H. Methylation of p16INK4A and mitotic arrest defective protein 2 (MAD2) genes in gastric marginal-zone B-cell lymphomas. Acta Haematol. 2008, 120, 217–224. [Google Scholar] [CrossRef]
- Yamamoto, M.L.; Schiestl, R.H. Intestinal microbiome and lymphoma development. Cancer J. 2014, 20, 190–194. [Google Scholar] [CrossRef] [Green Version]
- Saxena, A.; Mukhopadhyay, A.K.; Nandi, S.P. Helicobacter pylori: Perturbation and restoration of gut microbiome. J. Biosci. 2020, 45, 110. [Google Scholar] [CrossRef]
- Elsalem, L.; Jum’ah, A.A.; Alfaqih, M.A.; Aloudat, O. The bacterial microbiota of gastrointestinal cancers: Role in cancer pathogenesis and therapeutic perspectives. Clin. Exp. Gastroenterol. 2020, 13, 151–185. [Google Scholar] [CrossRef]
- Dutta, D.; Lim, S.H. Bidirectional interaction between intestinal microbiome and cancer: Opportunities for therapeutic interventions. Biomark Res. 2020, 8, 31. [Google Scholar] [CrossRef]
- Zucca, E.; Arcaini, L.; Buske, C.; Johnson, P.W.; Ponzoni, M.; Raderer, M.; Ricardi, U.; Salar, A.; Stamatopoulos, K.; Thieblemont, C.; et al. ESMO Guidelines Committee. Marginal zone lymphomas: ESMO Clinical Practice Guidelines for diagnosis, treatment and follow-up. Ann. Oncol. 2020, 31, 17–29. [Google Scholar] [CrossRef] [Green Version]
- Malfertheiner, P.; Megraud, F.; O’Morain, C.A.; Gisbert, J.P.; Kuipers, E.J.; Axon, A.T.; Bazzoli, F.; Gasbarrini, A.; Atherton, J.; Graham, D.Y.; et al. European Helicobacter and Microbiota Study Group and Consensus panel. Management of Helicobacter pylori infection-the Maastricht V/Florence Consensus Report. Gut 2017, 66, 6–30. [Google Scholar] [CrossRef] [Green Version]
- Wotherspoon, A.C.; Doglioni, C.; Diss, T.C.; Pan, L.; Moschini, A.; de Boni, M.; Isaacson, P.G. Regression of primary low-grade B-cell gastric lymphoma of mucosa-associated lymphoid tissue type after eradication of Helicobacter pylori. Lancet 1993, 342, 575–577. [Google Scholar] [CrossRef]
- Raderer, M.; Wohrer, S.; Kiesewetter, B.; Dolak, W.; Lagler, H.; Wotherspoon, A.; Muellauer, L.; Chott, A. Antibiotic treatment as sole management of Helicobacter pylori-negative gastric MALT lymphoma: A single center experience with prolonged follow-up. Ann. Hematol. 2015, 94, 969–973. [Google Scholar] [CrossRef]
- Paydas, S. Helicobacter pylori eradication in gastric diffuse large B cell lymphoma. World J. Gastroenterol. 2015, 21, 3773–3776. [Google Scholar] [CrossRef]
- Moleiro, J.; Ferreira, S.; Lage, P.; Dias Pereira, A. Gastric malt lymphoma: Analysis of a series of consecutive patients over 20 years. UE Gastroenterol. J. 2016, 4, 395–402. [Google Scholar] [CrossRef] [Green Version]
- Asano, N.; Iijima, K.; Koike, T.; Imatani, A.; Shimosegawa, T. Helicobacter pylori-negative gastric mucosa-associated lymphoid tissue lymphomas: A review. World J. Gastroenterol. 2015, 21, 8014–8020. [Google Scholar] [CrossRef]
- Megraud, F.; Coenen, S.; Versporten, A.; Kist, M.; Lopez-Brea, M.; Hirschl, A.M.; Andersen, L.P.; Goossens, H.; Glupczynski, Y.; Study Group participants. Helicobacter pylori resistance to antibiotics in Europe and its relationship to antibiotic consumption. Gut 2013, 62, 34–42. [Google Scholar] [CrossRef] [PubMed]
- Krzyżek, P.; Pawełka, D.; Iwańczak, B.; Kempiński, R.; Leśniakowski, K.; Mégraud, F.; Łaczmański, Ł.; Biernat, M.; Gościniak, G. High primary antibiotic resistance of Helicobacter pylori strains isolated from pediatric and adult patients in Poland during 2016–2018. Antibiotics 2020, 9, 228. [Google Scholar] [CrossRef] [PubMed]
- Younes, B.K.; Doghri, R.; Mrad, K.; Bedhiafi, W.; Benammar-Elgaaied, A.; Sola, B.; Aissa-Fennira, F.B. PTEN loss and cyclin A2 upregulation define a PI3K/AKT pathway activation in Helicobacter pylori-induced MALT and DLBCL gastric lymphoma with features of MALT. Appl. Immunohistochem. Mol. Morphol. 2021, 29, 56–61. [Google Scholar] [CrossRef]
- Ferreri, A.J.; Govi, S.; Raderer, M.; Mulè, A.; Andriani, A.; Caracciolo, D.; Devizzi, L.; Ilariucci, F.; Luminari, S.; Viale, E.; et al. Helicobacter pylori eradication as exclusive treatment for limited-stage gastric diffuse large B-cell lymphoma: Results of multicenter phase 2 trial. Blood 2012, 120, 3858–3860. [Google Scholar] [CrossRef] [Green Version]
- Tsai, H.J.; Tai, J.J.; Chen, L.T.; Wu, M.S.; Yeh, K.H.; Lin, C.W.; Wang, T.E.; Wang, H.P.; Yu, F.J.; Liou, J.M.; et al. Taiwan Cooperative Oncology Group A multicenter prospective study of first-line antibiotic therapy for early-stage gastric mucosa-associated lymphoid tissue lymphoma and diffuse large B-cell lymphoma with histological evidence of mucosa-associated lymphoid tissue. Haematologica 2020, 105, e349–e354. [Google Scholar]
- Kuo, S.H.; Yeh, K.H.; Chen, L.T.; Lin, C.W.; Hsu, P.N.; Hsu, C.; Wu, M.S.; Tzeng, Y.S.; Tsai, H.J.; Wang, H.P.; et al. Helicobacter pylori-related diffuse large B-cell lymphoma of the stomach: A distinct entity with lower aggressiveness and higher chemosensitivity. Blood Cancer J. 2014, 4, e220. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Y.; Xiao, Y.; Zhou, R.; Liao, Y.; Zhou, J.; Ma, X. Prognostic significance of Helicobacter pylori-infection in gastric diffuse large B-cell lymphoma. BMC Cancer 2019, 19, 842. [Google Scholar] [CrossRef] [PubMed]
- Kurucu, N.; Akyüz, C.; Yalçın, B.; Bajin, I.Y.; Varan, A.; Orhan, D.; Karnak, I.; Aydın, B.; Kutluk, T. Primary gastric lymphoma: A report of 16 pediatric cases treated at a single institute and review of the literature. Pediatr. Hematol. Oncol. 2020, 37, 656–664. [Google Scholar] [CrossRef]
- Baumgaertner, I.; Copie-Bergman, C.; Levy, M.; Haioun, C.; Charachon, A.; Baia, M.; Sobhani, I.; Delchier, J.C. Complete remission of gastric Burkitt’s lymphoma after eradication of Helicobacter pylori. World J. Gastroenterol. 2009, 15, 5746–5750. [Google Scholar] [CrossRef]
- Moschovi, M.; Menegas, D.; Stefanaki, K.; Constantinidou, C.V.; Tzortzatou-Stathopoulou, F. Med Primary gastric Burkitt lymphoma in childhood: Associated with Helicobacter pylori? Pediatr. Oncol. 2003, 41, 444–447. [Google Scholar] [CrossRef]
- Ruan, W.; Febo-Rodriguez, L.; Daignault, C.; Gulati, N.; Narine, K.; Sher, A.C.; Elghetany, M.T.; Cortes-Santiago, N.; Fishman, D.S.; Olive, A.P. Endoscopic ultrasound-guided diagnosis of Helicobacter pylori-associated gastric Burkitt’s lymphoma in an adolescent patient: A rare case. Clin. J. Gastroenterol. 2021, 14, 88–91. [Google Scholar] [CrossRef]
- Grewal, S.S.; Hunt, J.P.; O’Connor, S.C.; Gianturco, L.E.; Richardson, M.W.; Lehmann, L.E. Helicobacter pylori associated gastric Burkitt lymphoma. Pediatr. Blood Cancer. 2008, 50, 888–890. [Google Scholar] [CrossRef]
- Campuzano-Maya, G. Hematologic manifestations of Helicobacter pylori infection. World J. Gastroenterol. 2014, 20, 12818–12838. [Google Scholar] [CrossRef]
- Al-Saleem, T. Immunoproliferative small intestinal disease (IPSID): A model for mature B-cell neoplasms. Blood 2005, 105, 2274–2280. [Google Scholar] [CrossRef]
- Ewers, E.C.; Sheffler, R.L.; Wang, J.; Ngauy, V. Immunoproliferative small intestinal disease associated with overwhelming polymicrobial gastrointestinal infection with transformation to diffuse large B-cell lymphoma. Am. J. Trop Med. Hyg. 2016, 94, 1177–1181. [Google Scholar] [CrossRef] [Green Version]
- Lecuit, M.; Abachin, E.; Martin, A.; Poyart, C.; Pochart, P.; Suarez, F.; Bengoufa, D.; Feuillard, J.; Lavergne, A.; Gordon, J.I.; et al. Immunoproliferative small intestinal disease associated with Campylobacter jejuni. N. Engl. J. Med. 2004, 350, 239–248. [Google Scholar] [CrossRef] [PubMed]
- Mesnard, B.; De Vroey, B.; Maunoury, V.; Lecuit, M. Immunoproliferative small intestinal disease associated with Campylobacter jejuni. Dig. Liver Dis. 2012, 44, 799–800. [Google Scholar] [CrossRef]
- Roncati, L.; Maiorana, A. IgA plasmablastic large B-cell lymphoma. Diagnosis 2017, 4, 105–107. [Google Scholar] [CrossRef]
- Kreling, V.; Falcone, F.H.; Kehrenberg, C.; Hensel, A. Campylobacter sp.: Pathogenicity factors and prevention methods-new molecular targets for innovative antivirulence drugs? Appl. Microbiol. Biotechnol. 2020, 104, 10409–10436. [Google Scholar] [CrossRef] [PubMed]
- He, Z.; Gharaibeh, R.Z.; Newsome, R.C.; Pope, J.L.; Dougherty, M.W.; Tomkovich, S.; Pons, B.; Mirey, G.; Vignard, J.; Hendrixson, D.R.; et al. Campylobacter jejuni promotes colorectal tumorigenesis through the action of cytolethal distending toxin. Gut 2019, 68, 289–300. [Google Scholar] [CrossRef] [Green Version]
- Travaglino, A. Borrelia burgdorferi in primary cutaneous lymphomas: A systematic review and meta-analysis. J. Dtsch. Dermatol. Ges. 2020, 18, 1379–1384. [Google Scholar] [CrossRef] [PubMed]
- Sriskandarajah, P.; Dearden, C.E. Epidemiology and environmental aspects of marginal zone lymphomas. Best Pract. Res. Clin. Haematol. 2017, 30, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Tadiotto Cicogna, G.; Ferranti, M.; Alaibac, M. Diagnostic Workup of Primary Cutaneous B Cell Lymphomas: A Clinician’s Approach. Front. Oncol. 2020, 10, 988. [Google Scholar] [CrossRef]
- Leinweber, B.; Colli, C.; Chott, A.; Kerl, H.; Cerroni, L. Differential diagnosis of cutaneous infiltrates of B lymphocytes with follicular growth pattern. Am. J. Dermatopathol. 2004, 26, 4–13. [Google Scholar] [CrossRef]
- Colli, C.; Leinweber, B.; Mullegger, R.; Chott, A.; Kerl, H.; Cerroni, L. Borrelia burgdorferi-associated lymphocytoma cutis: Clinicopathologic, immunophenotypic, and molecular study of 106 cases. J. Cutan. Pathol. 2004, 31, 232–240. [Google Scholar] [CrossRef] [PubMed]
- Aberer, E.; Fingerle, V.; Wutte, N.; Fink-Puches, R.; Cerroni, L. Within European margins. Lancet 2011, 377, 178. [Google Scholar] [CrossRef]
- Travaglino, A.; Mirella Pace, M.; Varricchio, S.; Della Pepa, R.; Iuliano, A.; Picardi, M.; Pane, F.; Staibano, S.; Mascolo, M. Prevalence of Chlamydia psittaci, Chlamydia pneumoniae, and Chlamydia trachomatis determined by molecular testing in ocular adnexa lymphoma specimens. Am. J. Clin. Pathol. 2020, 153, 427–434. [Google Scholar] [CrossRef] [PubMed]
- Chanudet, E.; Adam, P.; Nicholson, A.G.; Wotherspoon, A.C.; Ranaldi, R.; Goteri, G.; Pileri, S.A.; Ye, H.; Müller-Hermelink, H.K.; Du, M.Q. Chlamydiae and Mycoplasma infections in pulmonary MALT lymphoma. Br. J. Cancer 2007, 97, 949–951. [Google Scholar] [CrossRef] [Green Version]
- Ferreri, A.J.M.; Guidoboni, M.; Ponzoni, M.; De Conciliis, C.; Dell’Oro, S.; Fleischhauer, K.; Caggiari, L.; Lettini, A.A.; Dal Cin, E.; Ieri, R.; et al. Evidence for an association between Chlamydia psittaci and ocular adnexal lymphomas. J. Natl. Cancer Inst. 2004, 96, 586–594. [Google Scholar] [CrossRef]
- Zhang, G.S.; Winter, J.N.; Variakojis, D.; Reich, S.; Lissner, G.S.; Bryar, P.; Regner, M.; Mangold, K.; Kaul, K. Lack of an association between Chlamydia psittaci and ocular adnexal lymphoma. Leuk. Lymphoma 2007, 48, 577–583. [Google Scholar] [CrossRef] [PubMed]
- Mutoh, T.; Takatsuki, H.; Mannoji, K.; Kawamura, K.; Okura, N.; Ohshima, K. Regression of uterine cervical diffuse large B-cell lymphoma transformed from mucosa-associated lymphoid tissue lymphoma subsequent to Chlamydia trachomatis eradication. Rinsho Ketsueki 2017, 58, 912–916. [Google Scholar]
- Sassone, M.; Ponzoni, M.; Ferreri, A.J. Ocular adnexal marginal zone lymphoma: Clinical presentation, pathogenesis, diagnosis, prognosis, and treatment. Best Pract. Res. Clin. Haematol. 2017, 30, 118–130. [Google Scholar] [CrossRef]
- Collina, F.; De Chiara, A.; De Renzo, A.; De Rosa, G.; Botti, G.; Franco, R. Chlamydia psittaci in ocular adnexa MALT lymphoma: A possible role in lymphomagenesis and a different geographical distribution. Infect. Agent Cancer 2012, 7, 8. [Google Scholar] [CrossRef] [Green Version]
- Kalogeropoulos, D.; Papoudou-Bai, A.; Kanavaros, P.; Kalogeropoulos, C. Ocular adnexal marginal zone lymphoma of mucosa-associated lymphoid tissue. Clin. Exp. Med. 2018, 18, 151–163. [Google Scholar] [CrossRef] [PubMed]
- Ferreri, A.J.; Govi, S.; Pasini, E.; Mappa, S.; Bertoni, F.; Zaja, F.; Montalbán, C.; Stelitano, C.; Cabrera, M.E.; GiordanonResti, A.; et al. Chlamydophila psittaci eradication with doxycycline as first-line targeted therapy for ocular adnexae lymphoma: Final results of an international phase II trial. J. Clin. Oncol. 2012, 30, 2988–2994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanguedolce, F.; Zanelli, M.; Zizzo, M.; Bisagni, A.; Soriano, A.; Cocco, G.; Palicelli, A.; Santandrea, G.; Caprera, C.; Corsi, M.; et al. Primary pulmonary B-cell lymphoma: A review and update. Cancers 2021, 13, 415. [Google Scholar] [CrossRef] [PubMed]
- Adam, P.; Czapiewski, P.; Colak, S.; Kosmidis, P.; Tousseyn, T.; Sagaert, X.; Boudova, L.; Okoń, K.; Morresi-Hauf, A.; Agostinelli, M.C.; et al. Prevalence of Achromobacter xylosoxidans in pulmonary mucosa-associated lymphoid tissue lymphoma in different regions of Europe. Br. J. Haematol. 2014, 164, 804–810. [Google Scholar] [CrossRef] [PubMed]
- Aoyama, S.; Masaki, A.; Sakamoto, Y.; Takino, H.; Murase, T.; Ohshima, K.; Yoshino, T.; Kato, S.; Inagaki, H. Achromobacter infection is rare in Japanese patients with pulmonary B-cell lymphoma. Intern. Med. 2018, 57, 789–794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cools, P.; Ho, E.; Vranckx, K.; Schelstraete, P.; Wurth, B.; Franckx, H.; Ieven, G.; Van Simaey, L.; Van Daele, S.; Verhulst, S.; et al. Epidemic Achromobacter xylosoxidans strain among Belgian cystic fibrosis patients and review of literature. BMC Microbiol. 2016, 16, 122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borie, R.; Caro, V.; Nunes, H.; Kambouchner, M.; Cazes, A.; Antoine, M.; Crestani, B.; Leroy, K.; Copie-Bergman, C.; Kwasiborski, A.; et al. No evidence for a pathogen associated with pulmonary MALT lymphoma: A metagenomics investigation. Infect. Agent Cancer. 2021, 16, 10. [Google Scholar] [CrossRef]
- Klein, T.O.; Soll, B.A.; Issel, B.F.; Fraser, C. Bronchus-associated lymphoid tissue lymphoma and Mycobacterium tuberculosis infection: An unusual case and a review of the literature. Respir. Care 2007, 52, 755–758. [Google Scholar]
- Gaur, S.; Trayner, E.; Aish, L.; Weinstein, R. Bronchus-associated lymphoid tissue lymphoma arising in a patient with bronchiectasis and chronic Mycobacterium avium infection. Am. J. Hematol. 2004, 77, 22–25. [Google Scholar] [CrossRef]
- Ishimatsu, Y.; Mukae, H.; Matsumoto, K.; Harada, T.; Hara, A.; Hara, S.; Amenomori, M.; Fujita, H.; Sakamoto, N.; Hayashi, T.; et al. Two cases with pulmonary mucosa-associated lymphoid tissue lymphoma successfully treated with clarithromycin. Chest 2010, 138, 730–733. [Google Scholar] [CrossRef]
- Sammassimo, S.; Pruneri, G.; Andreola, G.; Montoro, J.; Steffanoni, S.; Nowakowski, G.S.; Gandini, S.; Negri, M.; Habermann, T.M.; Raderer, M.; et al. A retrospective international study on primary extranodal marginal zone lymphoma of the lung (BALT lymphoma) on behalf of International Extranodal Lymphoma Study Group (IELSG). Hematol. Oncol. 2016, 34, 177–183. [Google Scholar] [CrossRef]
- Perrone, S.; D’Elia, G.M.; Annechini, G.; Pulsoni, A. Infectious aetiology of Marginal Zone Lymphoma and role of anti-infective therapy. Mediterr. J. Hematol. Infect. Dis. 2016, 8, e2016006. [Google Scholar] [CrossRef]
- Lyapichev, K.A.; Ivashkevich, Y.; Chernov, Y.; Chinenov, D.; Shpot, E.; Bessonov, A.A.; Dabaja, B.S.; Konoplev, S. MALT lymphoma of the urinary bladder shows a dramatic female predominance, uneven geographic distribution, and possible infectious etiology. Res. Rep. Urol. 2021, 13, 49–62. [Google Scholar]
- Kröber, S.M.; Aepinus, C.; Ruck, P.; Müller-Hermelink, H.K.; Horny, H.P.; Kaiserling, E. Extranodal marginal zone B cell lymphoma of MALT type involving the mucosa of both the urinary bladder and stomach. J. Clin. Pathol. 2002, 55, 554–557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simpson, W.G.; Lopez, A.; Babbar, P.; Payne, L.F. Primary bladder lymphoma, diffuse large B-cell type: Case report and literature review of 26 cases. Urol. Ann. 2015, 7, 268–272. [Google Scholar] [CrossRef]
- Lucioni, M.; Nicola, M.; Croci, G.A.; Rattotti, S.; Gotti, M.; Arcaini, L.; Paulli, M.; Cristina, S.; Valentini, S.; Martinengo, C. Antibiotic therapy-induced remission of bladder mucosa-associated lymphoid tissue (MALT) lymphoma carrying t(11;18)(q21;q21) apoptosis inhibitor 2-MALT1. J. Clin. Oncol. 2013, 31, e304–e306. [Google Scholar] [CrossRef]
- Fujimura, M.; Chin, K.; Sekita, N.; Kajimoto, S.; Kamijima, S.; Suzuki, H.; Ichikawa, T.; Mikami, K. Regression of mucosa-associated lymphoid tissue lymphoma of the bladder after antibiotic therapy: A case report. Hinyokika Kiyo. 2008, 54, 783–786. [Google Scholar]
- Terlizzi, M.E.; Gribaudo, G.; Maffei, M.E. UroPathogenic Escherichia coli (UPEC) infections: Virulence factors, bladder responses, antibiotic, and non-antibiotic antimicrobial strategies. Front. Microbiol. 2017, 8, 1566. [Google Scholar] [CrossRef]
- Hannan, T.J.; Totsika, M.; Mansfield, K.J.; Moore, K.H.; Schembri, M.A.; Hultgren, S.J. Host-pathogen checkpoints and population bottlenecks in persistent and intracellular uropathogenic Escherichia coli bladder infection. FEMS Microbiol Rev. 2012, 36, 616–648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Roeden, S.E.; van Houwelingen, F.; Donkers, C.M.J.; Hogewoning, M.M.A.; van der Hoek, W.; Kampschreur, L.M.; Bonten, M.J.M.; Hoepelman, A.I.M.; Bleeker-Rovers, C.P.; Wever, P.C.; et al. Exposure to Coxiella burnetii and risk of non-Hodgkin lymphoma: A retrospective population-based analysis in the Netherlands. Lancet Haematol. 2018, 5, e211–e219. [Google Scholar] [CrossRef]
- Melenotte, C.; Million, M.; Audoly, G.; Gorse, A.; Dutronc, H.; Roland, G.; Dekel, M.; Moreno, A.; Cammilleri, S.; Carrieri, M.P.; et al. B-cell non-Hodgkin lymphoma linked to Coxiella burnetii. Blood 2016, 127, 113–121. [Google Scholar] [CrossRef] [Green Version]
- Mezouar, S.; Omar Osman, I.; Melenotte, C.; Slimani, C.; Chartier, C.; Raoult, D.; Mege, J.L.; Devaux, C.A. High concentrations of serum soluble E-Cadherin in patients with Q Fever. Front. Cell Infect. Microbiol. 2019, 21, 219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Melenotte, C.; Mezouar, S.; Ben Amara, A.; Benatti, S.; Chiaroni, J.; Devaux, C.; Costello, R.; Kroemer, G.; Mege, J.L.; Raoult, D. A transcriptional signature associated with non-Hodgkin lymphoma in the blood of patients with Q fever. PLoS ONE 2019, 14, e0217542. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Pathogen | Type of Lymphoma | Localization |
|---|---|---|
| Helicobacter pylori | Gastric MALT lymphoma, EMZL, non-gastric MALT DLBCL lymphoma Burkitt lymphoma IPSID | Stomach, intestine |
| Campylobacter jejuni | IPSID | Intestine |
| Borrelia burgdorferi, Borrelia afzelii | MALT lymphoma | Skin |
| Chlamydophila psittaci | Ocular adnexal MALT lymphoma | Eye |
| Achromobacter xylosoxidans | BALT lymphoma | Lungs |
| Chlamydophila pneumoniae, Chlamydia trachomatis, Mycoplasma pneumoniae, | BALT lymphoma | Lungs |
| Mycobacterium tuberculosis, Mycobacterium avium | MALT lymphoma | Lungs |
| Eschericha coli | MALT lymphoma | Bladder |
| Coxiella burnetii | MALT lymphoma, DLBCL, FL, B-CLL | Various localizations |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Biernat, M.M.; Wróbel, T. Bacterial Infection and Non-Hodgkin B-Cell Lymphoma: Interactions between Pathogen, Host and the Tumor Environment. Int. J. Mol. Sci. 2021, 22, 7372. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22147372
Biernat MM, Wróbel T. Bacterial Infection and Non-Hodgkin B-Cell Lymphoma: Interactions between Pathogen, Host and the Tumor Environment. International Journal of Molecular Sciences. 2021; 22(14):7372. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22147372
Chicago/Turabian StyleBiernat, Monika Maria, and Tomasz Wróbel. 2021. "Bacterial Infection and Non-Hodgkin B-Cell Lymphoma: Interactions between Pathogen, Host and the Tumor Environment" International Journal of Molecular Sciences 22, no. 14: 7372. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22147372