
Chromatin Dynamics and Gene Expression Response to Heat Exposure in Field-Conditioned versus Laboratory-Cultured Nematostella vectensis

 ,
,
Abstract
:1. Introduction
2. Results
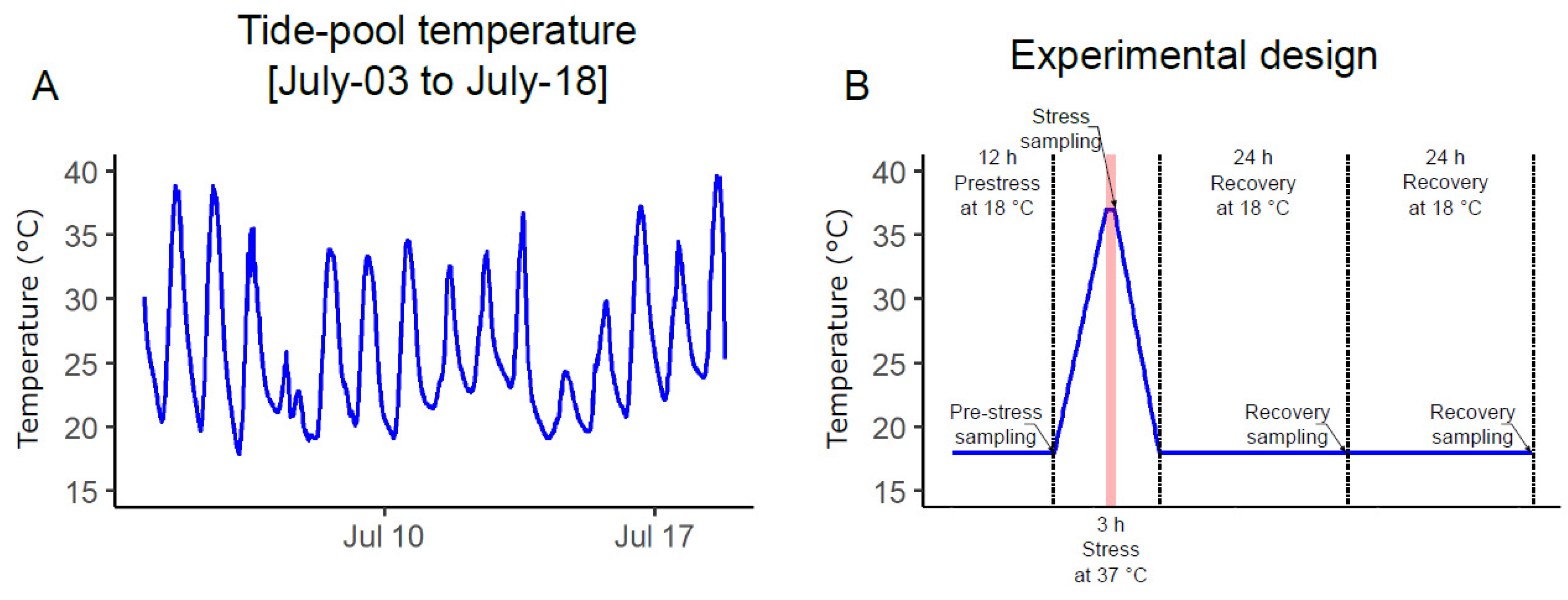
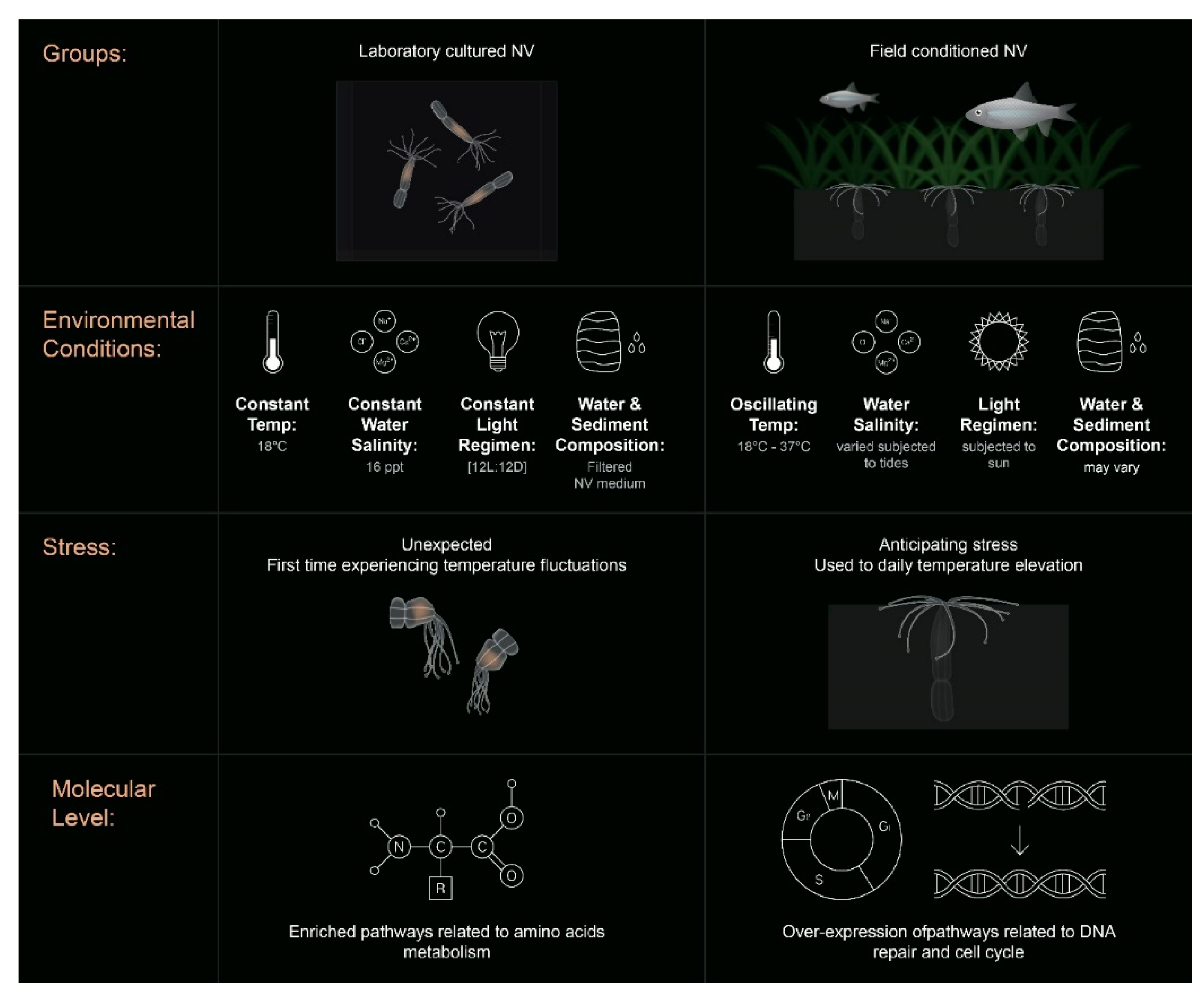
2.1. Field Conditioned N. vectensis
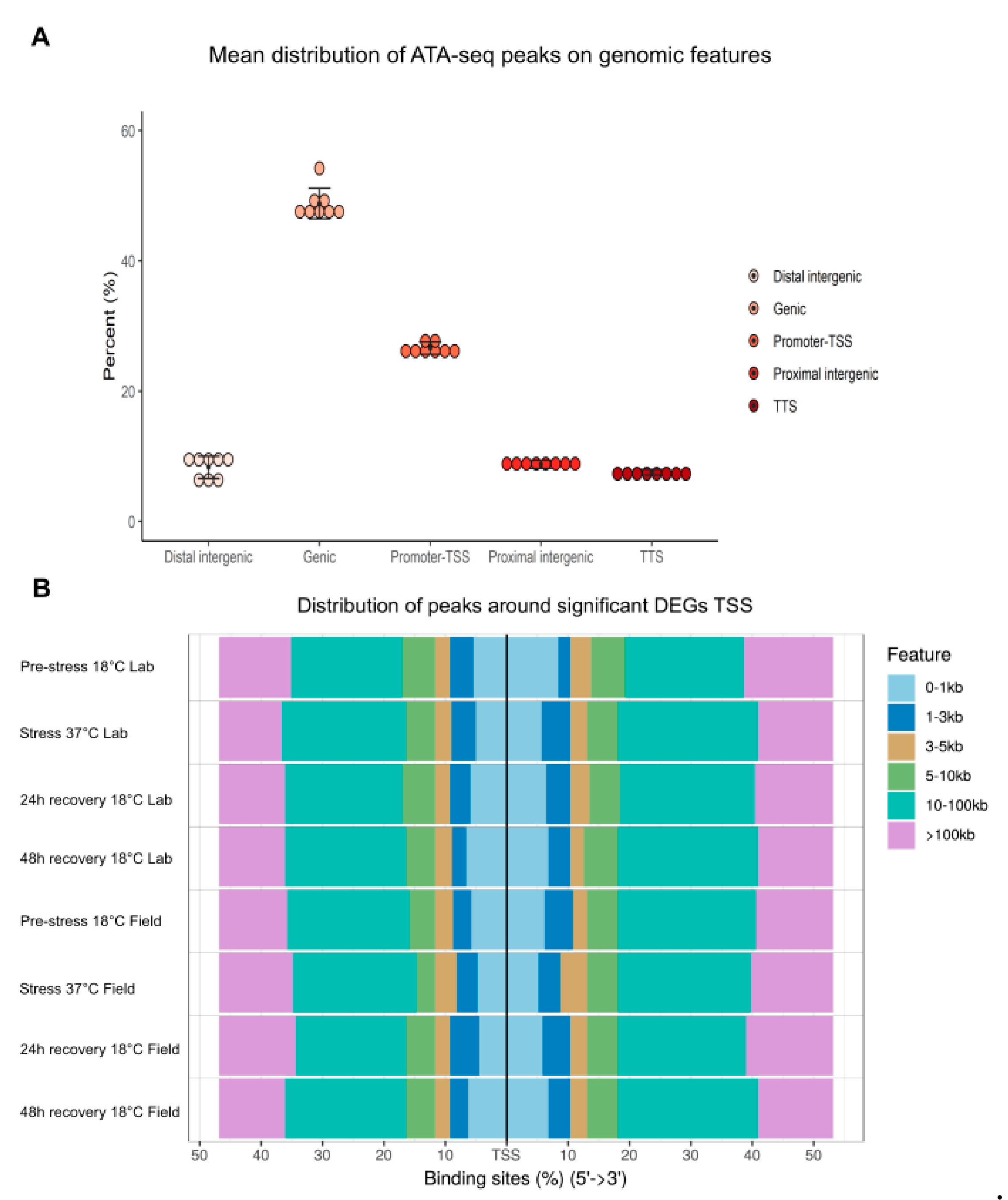
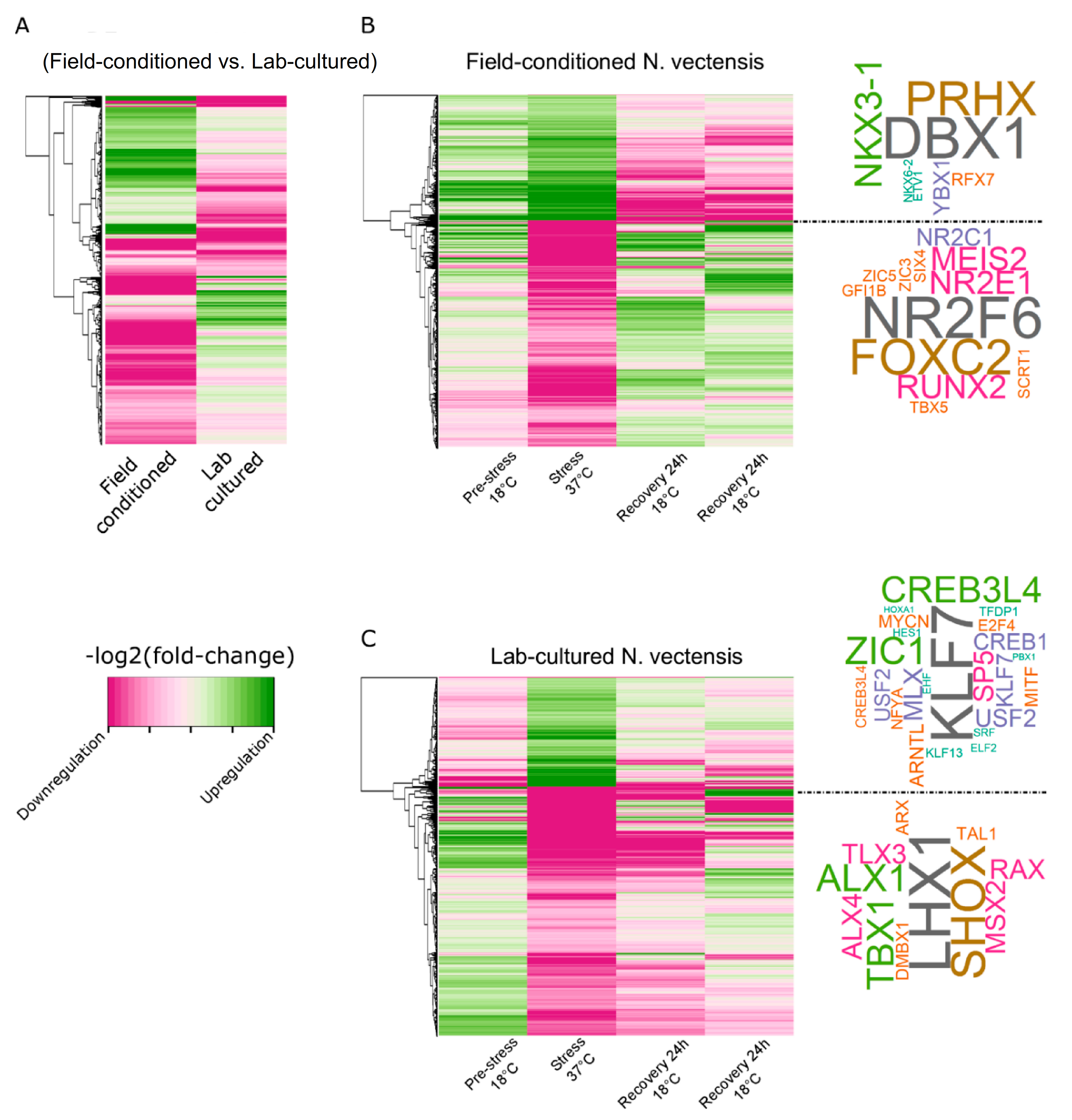
2.2. Chromatin Accessibility Profiles and Differential Expression Analysis of Field and Lab N. vectensis
2.3. Rearing Conditions Affect TF Enrichment and Transcriptional Response during Heat Exposure
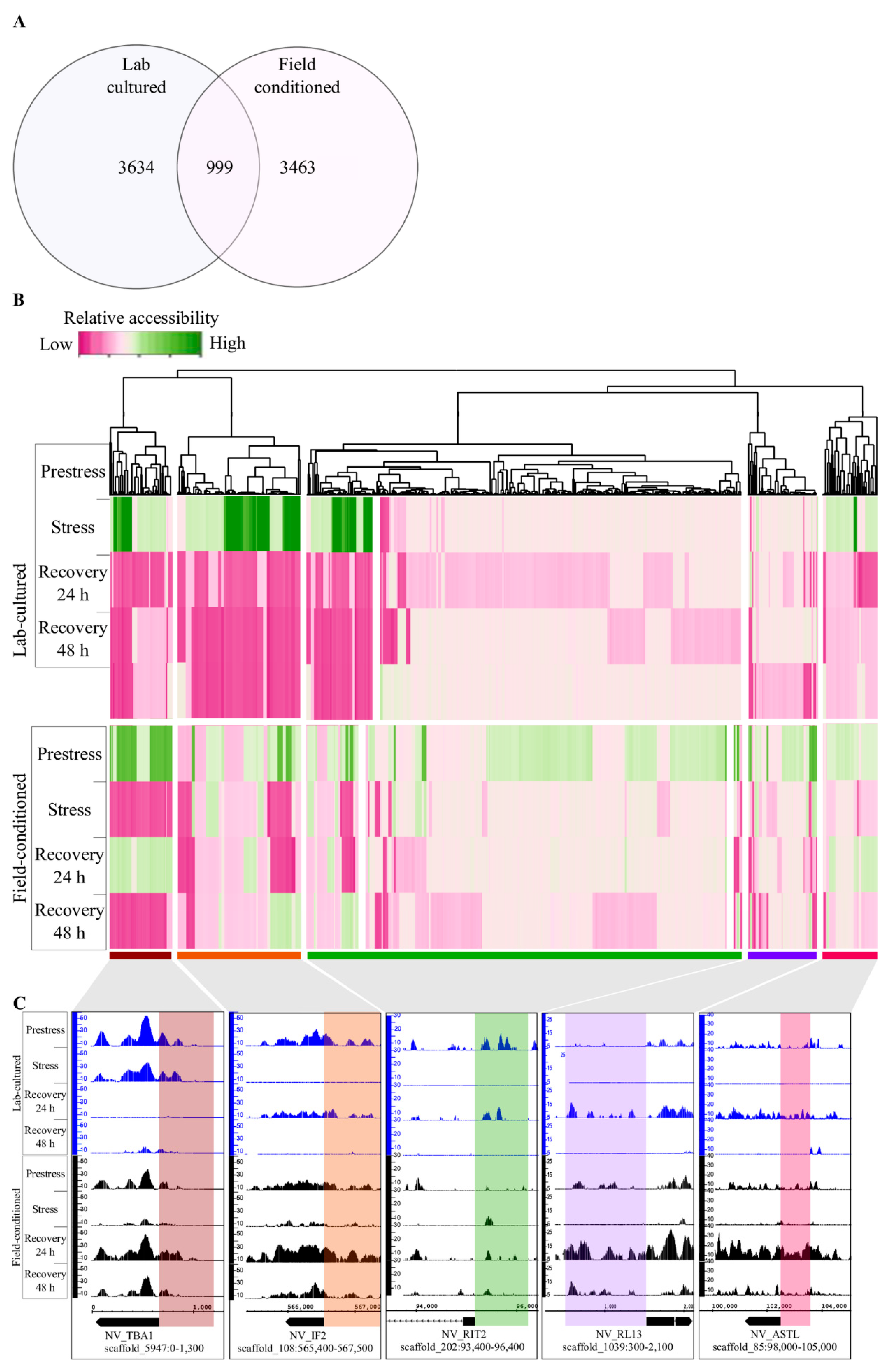
2.4. Changes in DNA Regulatory Landscape in Response to Heat Exposure
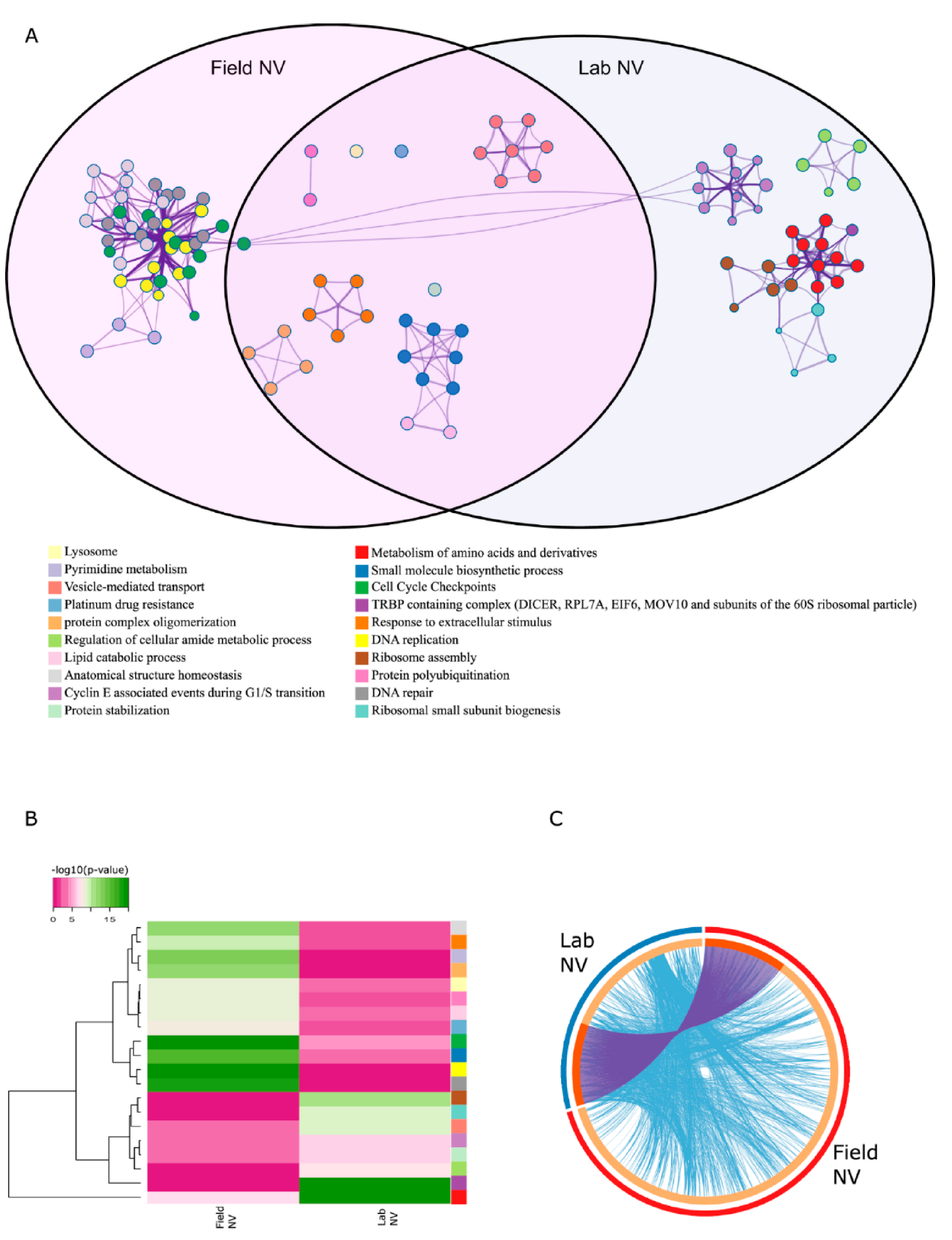
2.5. Enrichment Analysis Revealed Different Response Pathways
3. Discussion
4. Limitations of the Study
5. Materials and Methods
5.1. Animal Culture
5.2. Experimental Design
5.3. ATAC-Seq Nuclear Isolation and Library Preparation
5.4. RNA-Seq Library Preparation
5.5. Data Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Calisi, R.M.; Bentley, G.E. Lab and Field Experiments: Are They the Same Animal? Horm. Behav. 2009, 56, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Hand, C.; Uhlinger, K.R. The Unique, Widely Distributed, Estuarine Sea Anemone, Nematostella vectensis Stephenson: A Review, New Facts, and Questions. Estuaries 1994, 17, 501. [Google Scholar] [CrossRef]
- Finnerty, J.R.; Pang, K.; Burton, P.; Paulson, D.; Martindale, M.Q. Origins of Bilateral Symmetry: Hox and Dpp Expression in a Sea Anemone. Science 2004, 304, 1335–1337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwaiger, M.; Schönauer, A.; Rendeiro, A.F.; Pribitzer, C.; Schauer, A.; Gilles, A.F.; Schinko, J.B.; Renfer, E.; Fredman, D.; Technau, U. Evolutionary Conservation of the Eumetazoan Gene Regulatory Landscape. Genome Res. 2014, 24, 639–650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sebé-Pedrós, A.; Saudemont, B.; Chomsky, E.; Plessier, F.; Mailhé, M.-P.; Renno, J.; Loe-Mie, Y.; Lifshitz, A.; Mukamel, Z.; Schmutz, S.; et al. Cnidarian Cell Type Diversity and Regulation Revealed by Whole-Organism Single-Cell RNA-Seq. Cell 2018, 173, 1520–1534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sullivan, J.C.; Reitzel, A.M.; Finnerty, J.R. A High Percentage of Introns in Human Genes Were Present Early in Animal Evolution. Genome Inform. 2006, 17, 219–229. [Google Scholar] [PubMed]
- Reitzel, A.M.; Tarrant, A.M.; Levy, O. Circadian Clocks in the Cnidaria: Environmental Entrainment, Molecular Regulation, and Organismal Outputs. Integr. Comp. Biol. 2013, 53, 118–130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reitzel, A.M.; Sullivan, J.C.; Traylor-Knowles, N.; Finnerty, J.R. Genomic Survey of Candidate Stress-Response Genes in the Estuarine Anemone Nematostella Vectensis. Biol. Bull. 2008, 214, 233–254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reitzel, A.M.; Chu, T.; Edquist, S.; Genovese, C.; Church, C.; Tarrant, A.M.; Finnerty, J.R. Physiological and Developmental Responses to Temperature by the Sea Anemone Nematostella Vectensis. Mar. Ecol. Prog. Ser. 2013, 484, 115–130. [Google Scholar] [CrossRef] [Green Version]
- Palumbi, S.R.; Barshis, D.J.; Traylor-Knowles, N.; Bay, R.A. Mechanisms of Reef Coral Resistance to Future Climate Change. Science 2014, 344, 895–898. [Google Scholar] [CrossRef]
- Knighton, L.E.; Wolfgeher, D.; Reitzel, A.M.; Truman, A.W. Dataset of Nematostella Vectensis Hsp70 Isoform Interactomes upon Heat Shock. Data Brief 2019, 27, 104580. [Google Scholar] [CrossRef]
- Friedman, L.E.; Gilmore, T.D.; Finnerty, J.R. Intraspecific Variation in Oxidative Stress Tolerance in a Model Cnidarian: Differences in Peroxide Sensitivity between and within Populations of Nematostella Vectensis. PLoS ONE 2018, 13, e0188265. [Google Scholar] [CrossRef] [Green Version]
- Helm, R.R.; Martín-Díaz, M.L.; Tarrant, A.M. Phylogenetic Analysis of Cnidarian Peroxiredoxins and Stress-Responsive Expression in the Estuarine Sea Anemone Nematostella Vectensis. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2018, 221, 32–43. [Google Scholar] [CrossRef]
- Elran, R.; Raam, M.; Kraus, R.; Brekhman, V.; Sher, N.; Plaschkes, I.; Chalifa-Caspi, V.; Lotan, T. Early and Late Response of Nematostella Vectensis Transcriptome to Heavy Metals. Mol. Ecol. 2014, 23, 4722–4736. [Google Scholar] [CrossRef]
- Suzuki, M.M.; Bird, A. DNA Methylation Landscapes: Provocative Insights from Epigenomics. Nat. Rev. Genet. 2008, 9, 465–476. [Google Scholar] [CrossRef]
- Eirin-Lopez, J.M.; Putnam, H.M. Marine Environmental Epigenetics. Ann. Rev. Mar. Sci. 2019, 11, 335–368. [Google Scholar] [CrossRef] [Green Version]
- Tsompana, M.; Buck, M.J. Chromatin Accessibility: A Window into the Genome. Epigenetics Chromatin 2014, 7, 33. [Google Scholar] [CrossRef] [Green Version]
- Cyr, N.E.; Romero, L.M. Fecal Glucocorticoid Metabolites of Experimentally Stressed Captive and Free-Living Starlings: Implications for Conservation Research. Gen. Comp. Endocrinol. 2008, 158, 20–28. [Google Scholar] [CrossRef]
- Fuchikawa, T.; Eban-Rothschild, A.; Nagari, M.; Shemesh, Y.; Bloch, G. Potent Social Synchronization Can Override Photic Entrainment of Circadian Rhythms. Nat. Commun. 2016, 7, 11662. [Google Scholar] [CrossRef] [Green Version]
- Tomotani, B.M.; Flores, D.E.F.L.; Tachinardi, P.; Paliza, J.D.; Oda, G.A.; Valentinuzzi, V.S. Field and Laboratory Studies Provide Insights into the Meaning of Day-Time Activity in a Subterranean Rodent (Ctenomys Aff. Knighti), the Tuco-Tuco. PLoS ONE 2012, 7, e37918. [Google Scholar] [CrossRef] [Green Version]
- Vanin, S.; Bhutani, S.; Montelli, S.; Menegazzi, P.; Green, E.W.; Pegoraro, M.; Sandrelli, F.; Costa, R.; Kyriacou, C.P. Unexpected Features of Drosophila Circadian Behavioural Rhythms under Natural Conditions. Nature 2012, 484, 371–375. [Google Scholar] [CrossRef] [PubMed]
- Weizman, E.N.; Tannenbaum, M.; Tarrant, A.M.; Hakim, O.; Levy, O. Chromatin Dynamics Enable Transcriptional Rhythms in the Cnidarian Nematostella Vectensis. PLoS Genet. 2019, 15, e1008397. [Google Scholar] [CrossRef] [PubMed]
- Neumann, D. Timing in Tidal, Semilunar, and Lunar Rhythms. In Annual, Lunar, and Tidal Clocks; Numata, H., Helm, B., Eds.; Springer Japan: Tokyo, Japan, 2014; pp. 3–24. [Google Scholar]
- Naylor, E. Chronobiology of Marine Organisms; Cambridge University Press: Cambridge, UK, 2010. [Google Scholar]
- Hillyer, K.E.; Tumanov, S.; Villas-Bôas, S.; Davy, S.K. Metabolite Profiling of Symbiont and Host during Thermal Stress and Bleaching in a Model Cnidarian-Dinoflagellate Symbiosis. J. Exp. Biol. 2016, 219, 516–527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petrou, K.; Nielsen, D.A.; Heraud, P. Single-Cell Biomolecular Analysis of Coral Algal Symbionts Reveals Opposing Metabolic Responses to Heat Stress and Expulsion. Front. Mar. Sci. 2018, 5, 110. [Google Scholar] [CrossRef] [Green Version]
- Dixon, G.; Abbott, E.; Matz, M. Meta-Analysis of the Coral Environmental Stress Response: Acropora Corals Show Opposing Responses Depending on Stress Intensity. Mol. Ecol. 2020, 29, 2855–2870. [Google Scholar] [CrossRef]
- Tarrant, A.M.; Helm, R.R.; Levy, O.; Rivera, H.E. Environmental Entrainment Demonstrates Natural Circadian Rhythmicity in the Cnidarian Nematostella Vectensis. J. Exp. Biol. 2019, 222, jeb205393. [Google Scholar] [CrossRef]
- Rabinowitz, C.; Moiseeva, E.; Rinkevich, B. In Vitro Cultures of Ectodermal Monolayers from the Model Sea Anemone Nematostella Vectensis. Cell Tissue Res. 2016, 366, 693–705. [Google Scholar] [CrossRef]
- Langmead, B.; Salzberg, S.L. Fast Gapped-Read Alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Liu, T.; Meyer, C.A.; Eeckhoute, J.; Johnson, D.S.; Bernstein, B.E.; Nusbaum, C.; Myers, R.M.; Brown, M.; Li, W.; et al. Model-Based Analysis of ChIP-Seq (MACS). Genome Biol. 2008, 9, R137. [Google Scholar] [CrossRef] [Green Version]
- Buske, F.A.; Bodén, M.; Bauer, D.C.; Bailey, T.L. Assigning Roles to DNA Regulatory Motifs Using Comparative Genomics. Bioinformatics 2010, 26, 860–866. [Google Scholar] [CrossRef] [Green Version]
- Quinlan, A.R.; Hall, I.M. BEDTools: A Flexible Suite of Utilities for Comparing Genomic Features. Bioinformatics 2010, 26, 841–842. [Google Scholar] [CrossRef] [Green Version]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast Universal RNA-Seq Aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhou, B.; Pache, L.; Chang, M.; Khodabakhshi, A.H.; Tanaseichuk, O.; Benner, C.; Chanda, S.K. Metascape Provides a Biologist-Oriented Resource for the Analysis of Systems-Level Datasets. Nat. Commun. 2019, 10, 1523. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Name | % Alignment to Genome | % Alignment to Mitochondrial DNA |
|---|---|---|
| Pre-stress 18 °C Lab (rep-1) | 87.44% | 0.68% |
| Pre-stress 18 °C Lab (rep-2) | 85.15% | 0.71% |
| Pre-stress 18 °C Lab (rep-3) | 90.48% | 0.13% |
| Pre-stress 18 °C Field (rep-1) | 84.93% | 5.72% |
| Pre-stress 18 °C Field (rep-2) | 88.32% | 7.96% |
| Pre-stress 18 °C Field (rep-3) | 89.85% | 11.01% |
| Stress 37 °C Lab (rep-1) | 85.89% | 2.11% |
| Stress 37 °C Lab (rep-2) | 86.67% | 1.07% |
| Stress 37 °C Lab (rep-3) | 89.71% | 1.33% |
| Stress 37 °C Field (rep-1) | 82.25% | 2.11% |
| Stress 37 °C Field (rep-2) | 89.23% | 11.34% |
| Stress 37 °C Field (rep-3) | 86.49% | 13.93% |
| 24 h recovery 18 °C Lab (rep-1) | 91.37% | 0.93% |
| 24 h recovery 18 °C Lab (rep-2) | 91.85% | 3.45% |
| 24 h recovery 18 °C Lab (rep-3) | 82.26% | 1.26% |
| 24 h recovery 18 °C Field (rep-1) | 89.70% | 4.72% |
| 24 h recovery 18 °C Field (rep-2) | 84.97% | 6.66% |
| 24 h recovery 18 °C Field (rep-3) | 87.60% | 3.47% |
| 48 h recovery 18 °C Lab (rep-1) | 86.23% | 1.35% |
| 48 h recovery 18 °C Lab (rep-2) | 92.05% | 1.03% |
| 48 h recovery 18 °C Lab (rep-3) | 89.55% | 1.06% |
| 48 h recovery 18 °C Field (rep-1) | 83.37% | 5.13% |
| 48 h recovery 18 °C Field (rep-2) | 89.31% | 5.10% |
| 48 h recovery 18 °C Field (rep-3) | 82.38% | 0.89% |
| Sample Name | Rep 1 vs. Rep 2 | Rep 2 vs. Rep 3 | Rep 1 vs. Rep 3 |
|---|---|---|---|
| Pre-stress 18 °C Lab | 0.868 | 0.826 | 0.952 |
| Pre-stress 18 °C Field | 0.983 | 0.989 | 0.999 |
| Stress 37 °C Lab | 0.845 | 0.847 | 0.865 |
| Stress 37 °C Field | 0.815 | 0.746 | 0.83 |
| 24 h recovery 18 °C Lab | 0.805 | 0.816 | 0.789 |
| 24 h recovery 18 °C Field | 0.852 | 0.821 | 0.879 |
| 48 h recovery 18 °C Lab | 0.702 | 0.702 | 0.902 |
| 48 h recovery 18 °C Field | 0.959 | 0.191 | 0.17 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Weizman, E.; Rinsky, M.; Simon-Blecher, N.; Lampert-Karako, S.; Yaron, O.; Tarrant, A.M.; Levy, O. Chromatin Dynamics and Gene Expression Response to Heat Exposure in Field-Conditioned versus Laboratory-Cultured Nematostella vectensis. Int. J. Mol. Sci. 2021, 22, 7454. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22147454
Weizman E, Rinsky M, Simon-Blecher N, Lampert-Karako S, Yaron O, Tarrant AM, Levy O. Chromatin Dynamics and Gene Expression Response to Heat Exposure in Field-Conditioned versus Laboratory-Cultured Nematostella vectensis. International Journal of Molecular Sciences. 2021; 22(14):7454. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22147454
Chicago/Turabian StyleWeizman, Eviatar, Mieka Rinsky, Noa Simon-Blecher, Sarit Lampert-Karako, Orly Yaron, Ann M. Tarrant, and Oren Levy. 2021. "Chromatin Dynamics and Gene Expression Response to Heat Exposure in Field-Conditioned versus Laboratory-Cultured Nematostella vectensis" International Journal of Molecular Sciences 22, no. 14: 7454. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22147454