
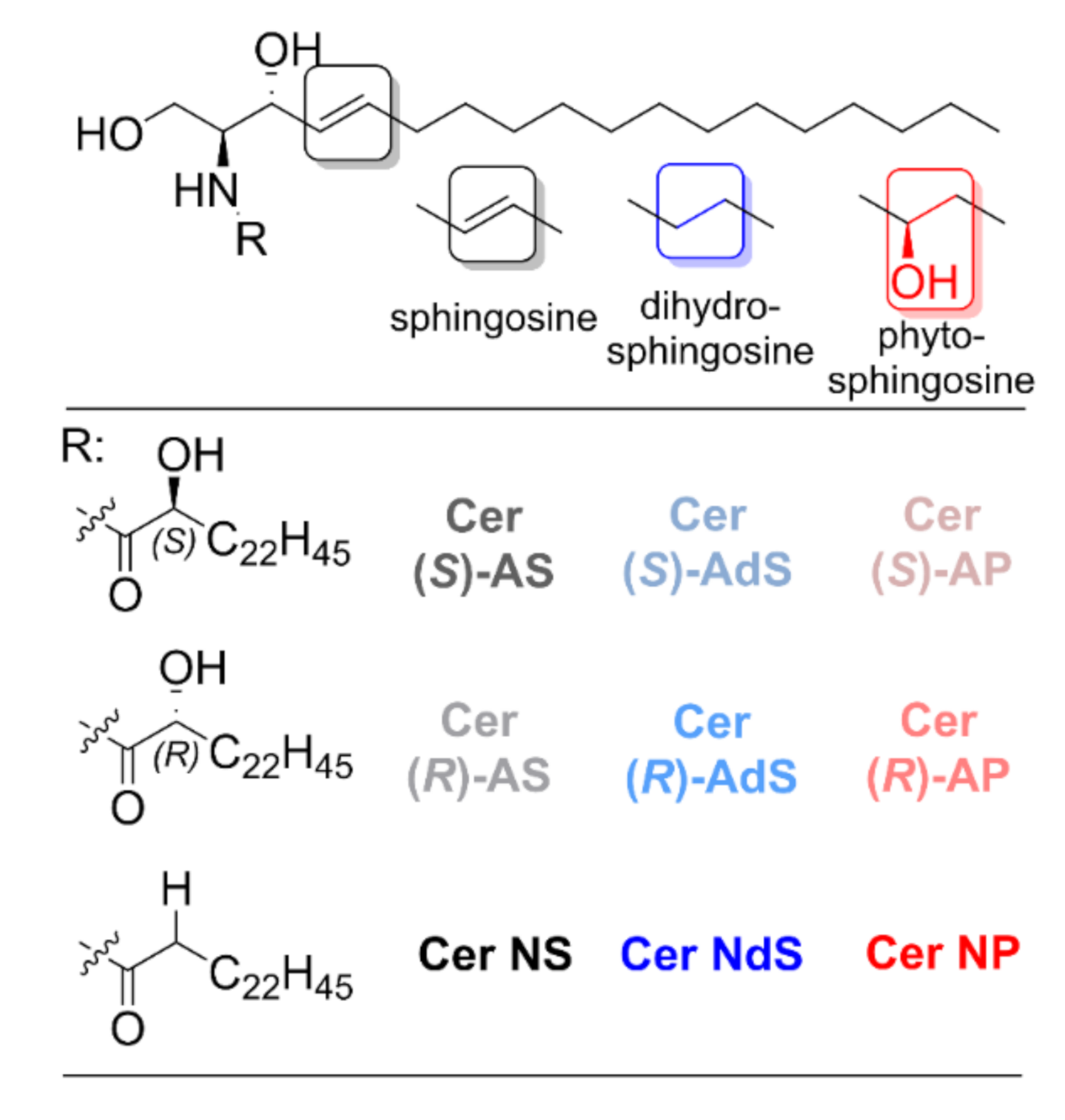
Effects of (R)- and (S)-α-Hydroxylation of Acyl Chains in Sphingosine, Dihydrosphingosine, and Phytosphingosine Ceramides on Phase Behavior and Permeability of Skin Lipid Models

 ,
,
Abstract
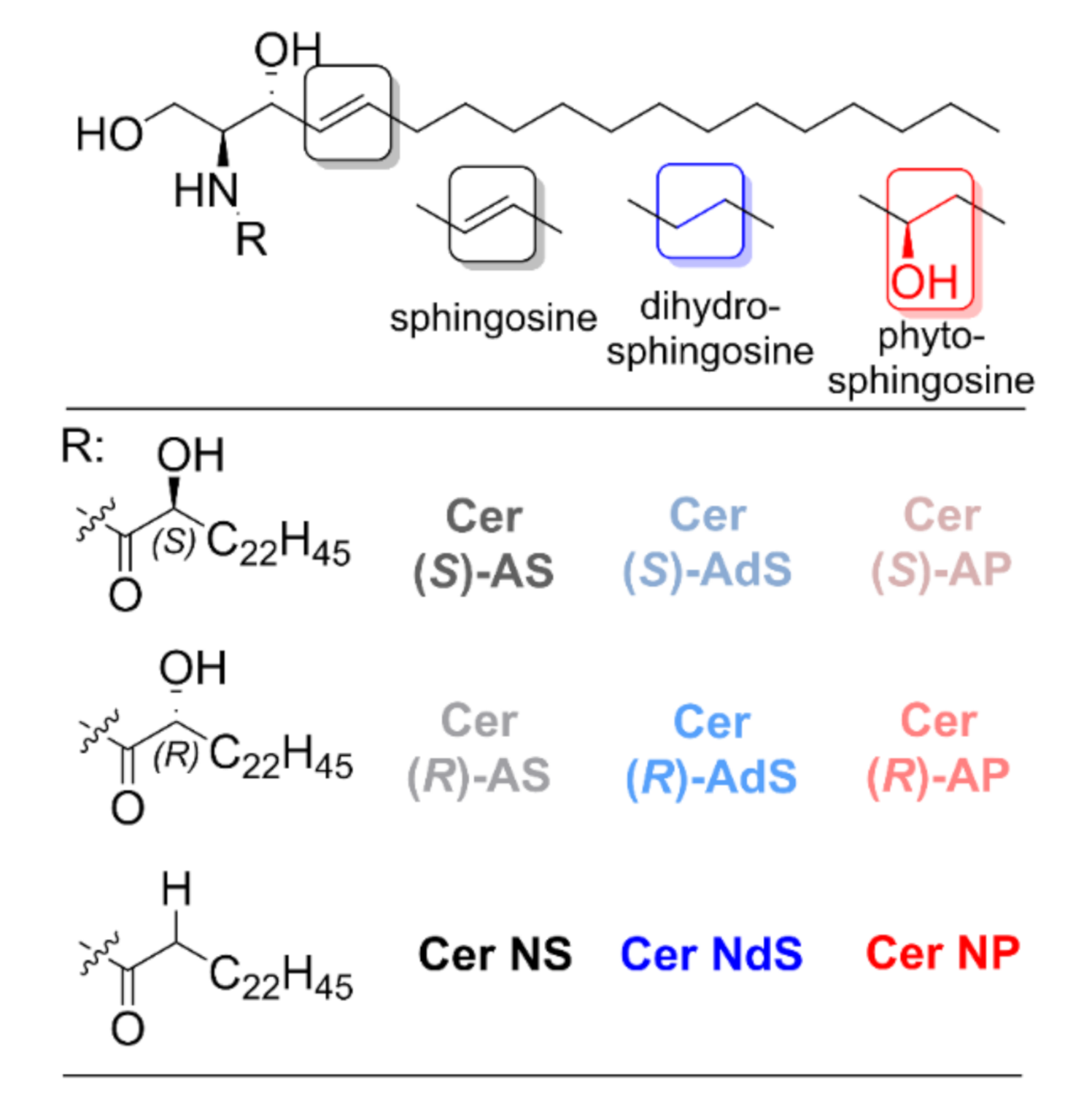
:1. Introduction
2. Results
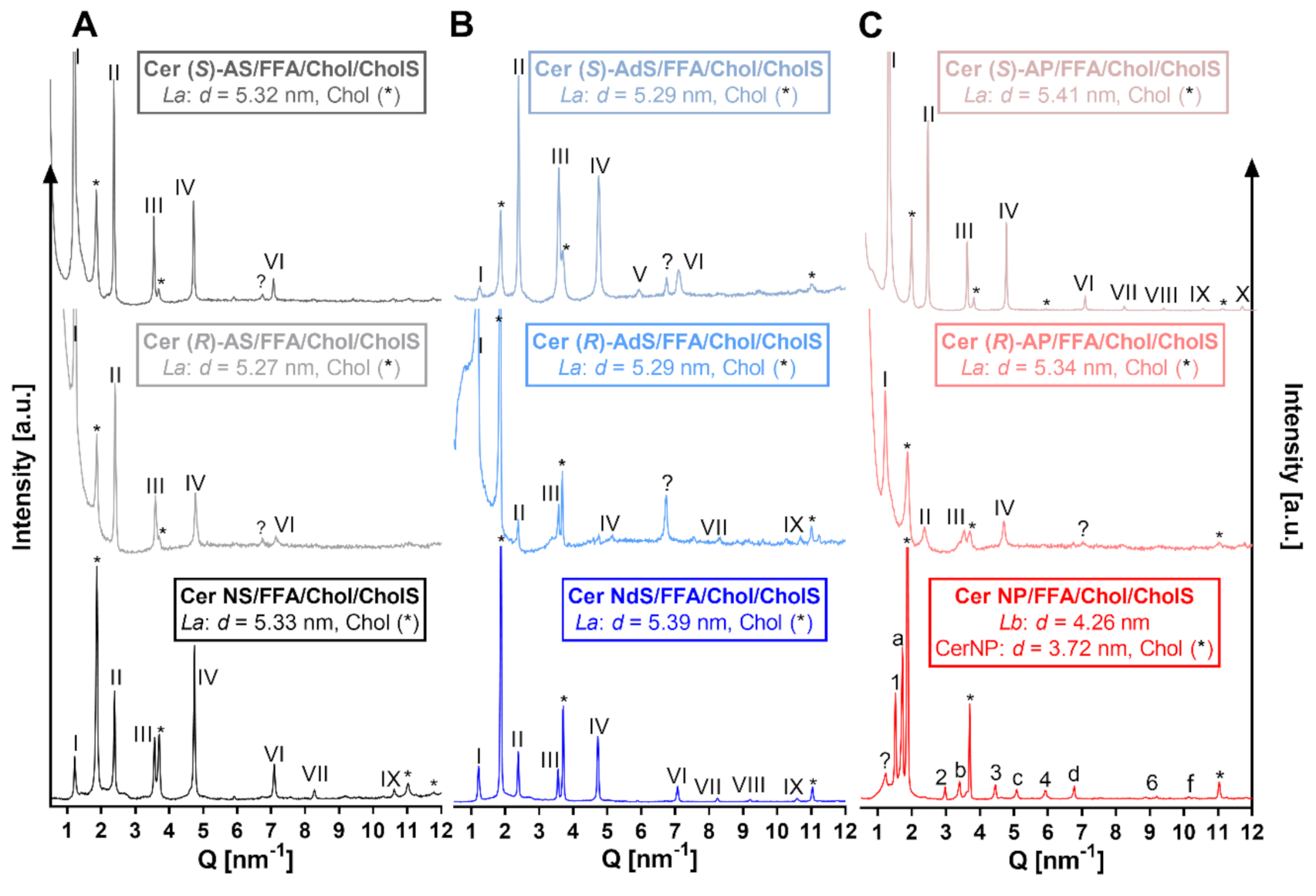
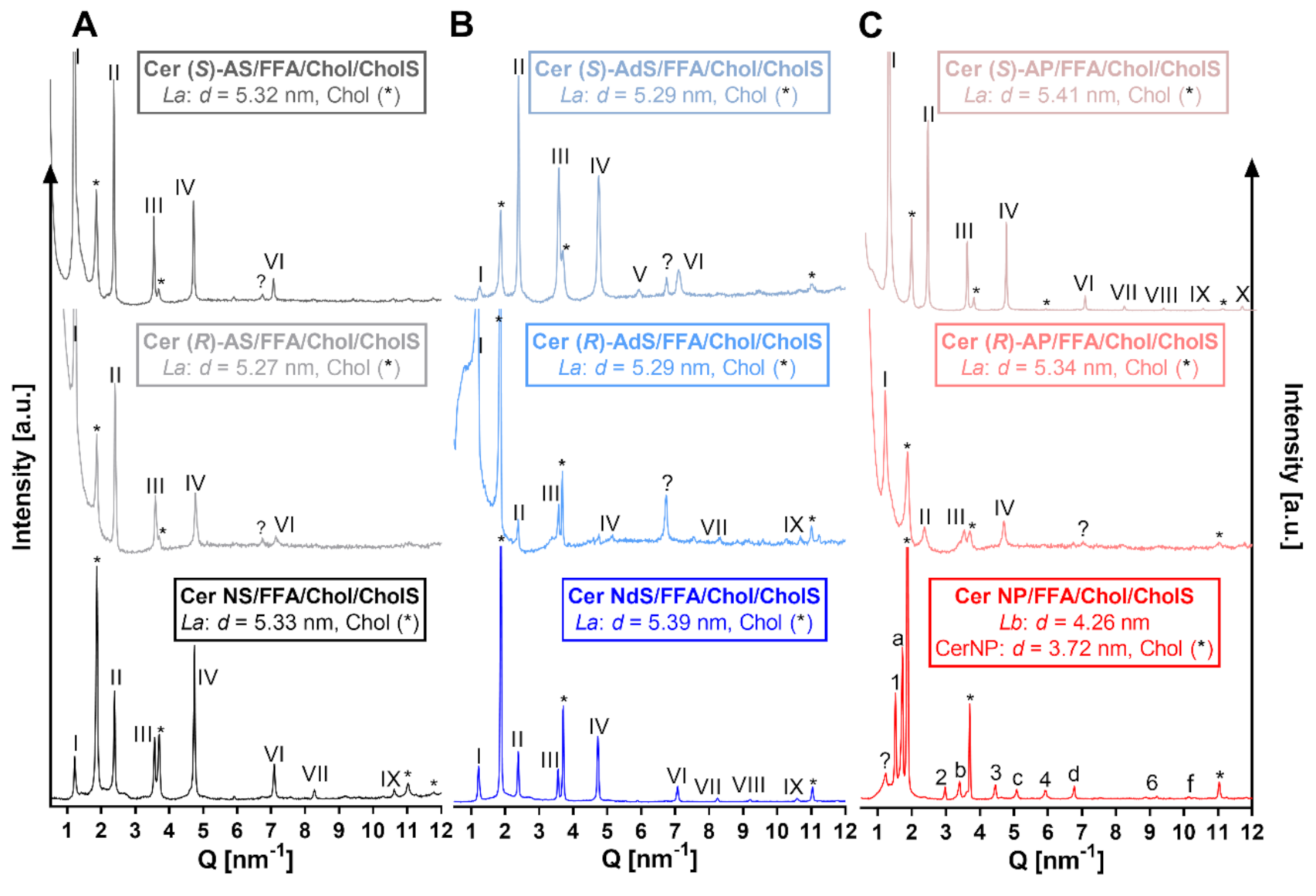
2.1. Lamellar Lipid Arrangement
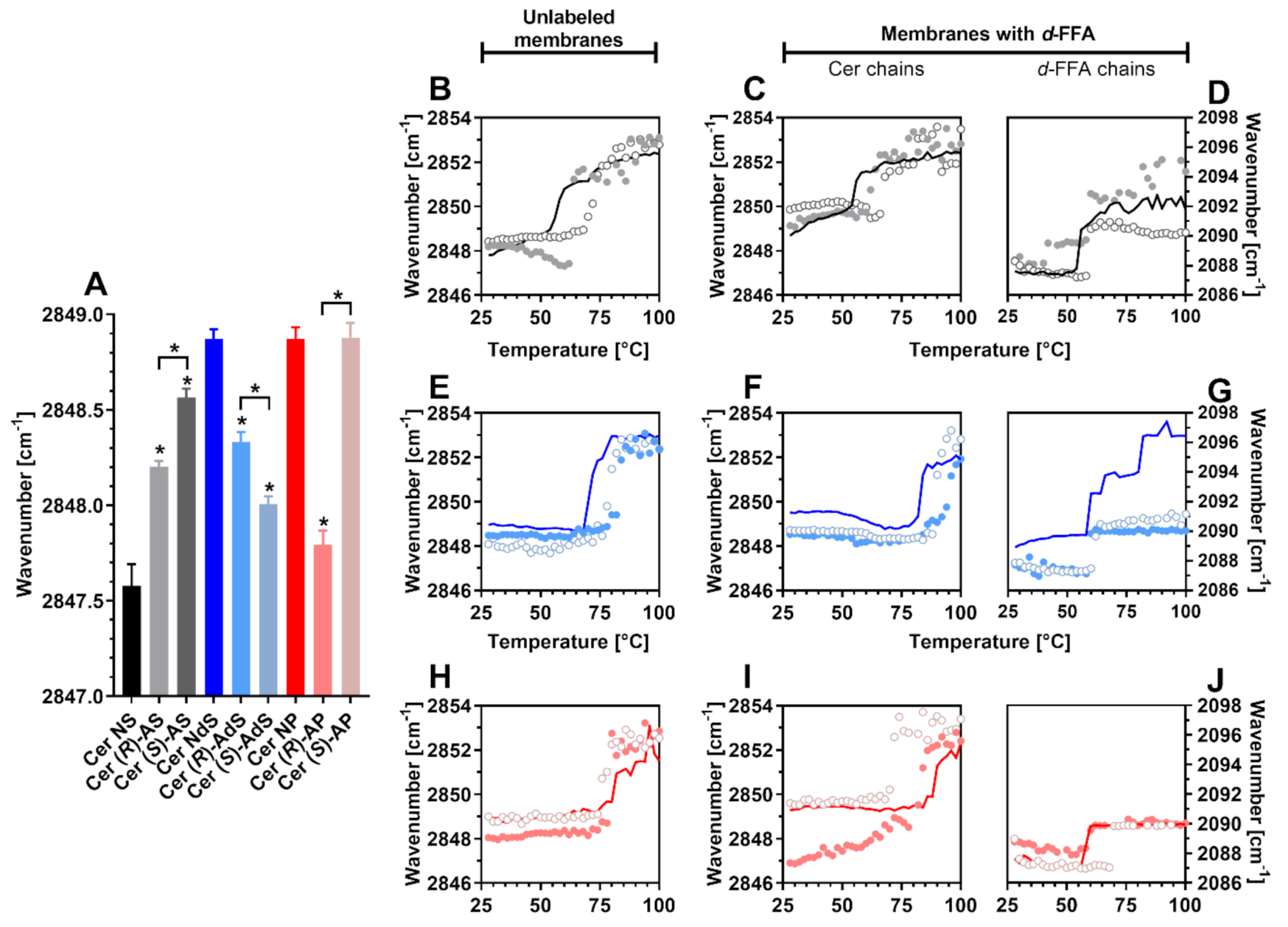
2.2. Lipid Chain Order
2.3. Phase Transitions
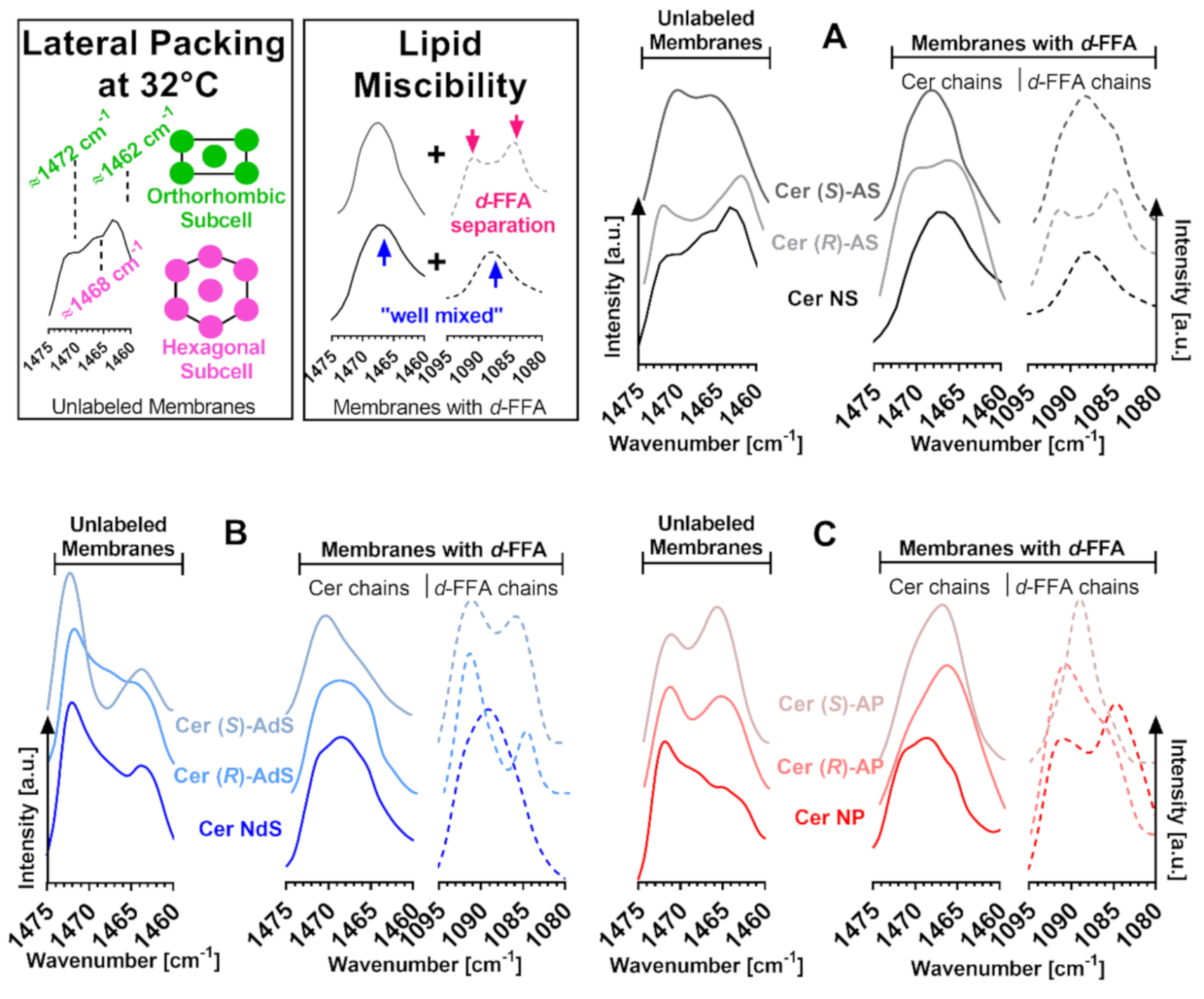
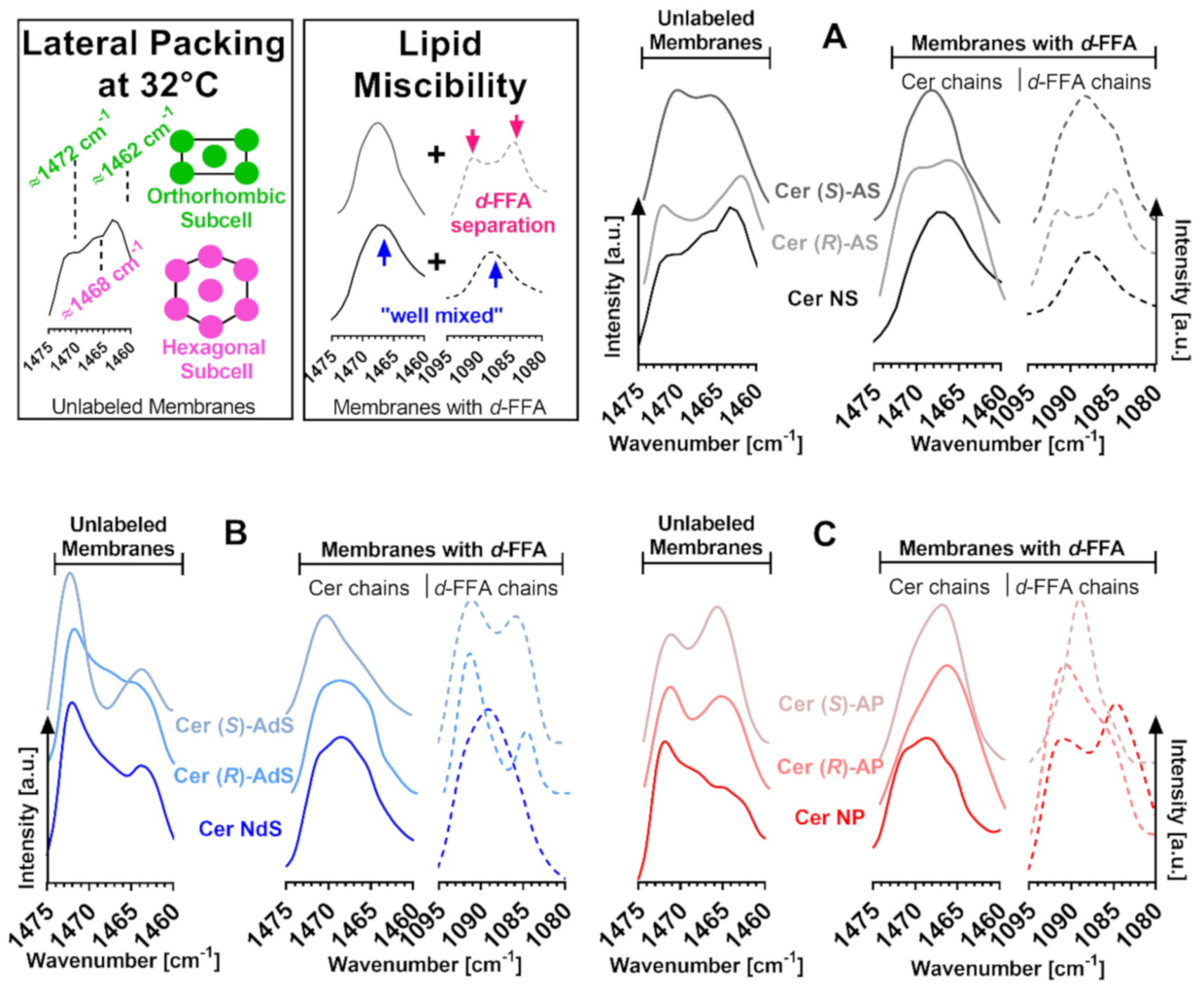
2.4. Lateral Chain Packing and Lipid Mixing
2.5. Model Membrane Permeability
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Preparation of the Lipid Membranes
4.3. X-ray Diffraction (XRD)
4.4. Attenuated Total Reflectance Fourier Transform Infrared Spectroscopy (FTIR)
4.5. Water Loss through the Lipid Membranes
4.6. Permeation Experiments
4.7. Data Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Elias, P.M. Epidermal Barrier Function-Intercellular Lamellar Lipid Structures, Origin, Composition and Metabolism. J. Control. Release 1991, 15, 199–208. [Google Scholar] [CrossRef]
- Wertz, P.W.; van den Bergh, B. The physical, chemical and functional properties of lipids in the skin and other biological barriers. Chem. Phys. Lipids 1998, 91, 85–96. [Google Scholar] [CrossRef]
- Bouwstra, J.A.; Dubbelaar, F.E.; Gooris, G.S.; Ponec, M. The lipid organisation in the skin barrier. Acta Derm. Venereol. Suppl. 2000, 208, 23–30. [Google Scholar]
- Bouwstra, J.; Pilgram, G.; Gooris, G.; Koerten, H.; Ponec, M. New aspects of the skin barrier organization. Skin Pharmacol. Physiol. 2001, 14, 52–62. [Google Scholar] [CrossRef] [PubMed]
- Feingold, K.; Elias, P. The important role of lipids in the epidermis and their role in the formation and maintenance of the cutaneous barrier. Biochim. Biophys. Acta 2014, 1841, 279. [Google Scholar] [CrossRef] [PubMed]
- Motta, S.; Monti, M.; Sesana, S.; Caputo, R.; Carelli, S.; Ghidoni, R. Ceramide composition of the psoriatic scale. Biochim. Biophys. Acta 1993, 1182, 147–151. [Google Scholar] [CrossRef]
- Breiden, B.; Sandhoff, K. The role of sphingolipid metabolism in cutaneous permeabilitybarrier formation. Biochim. Biophys. Acta 2014, 1841, 441–452. [Google Scholar] [CrossRef] [PubMed]
- Skolova, B.; Jandovska, K.; Pullmannova, P.; Tesar, O.; Roh, J.; Hrabalek, A.; Vavrova, K. The role of the trans double bond in skin barrier sphingolipids: Permeability and infrared spectroscopic study of model ceramide and dihydroceramide membranes. Langmuir ACS J. Surf. Colloids 2014, 30, 5527–5535. [Google Scholar] [CrossRef]
- Skolova, B.; Janusova, B.; Zbytovska, J.; Gooris, G.; Bouwstra, J.; Slepicka, P.; Berka, P.; Roh, J.; Palat, K.; Hrabalek, A.; et al. Ceramides in the skin lipid membranes: Length matters. Langmuir ACS J. Surf. Colloids 2013, 29, 15624–15633. [Google Scholar] [CrossRef]
- Skolova, B.; Janusova, B.; Vavrova, K. Ceramides with a pentadecasphingosine chain and short acyls have strong permeabilization effects on skin and model lipid membranes. BBA Biomembr. 2016, 1858, 220–232. [Google Scholar] [CrossRef] [PubMed]
- Skolova, B.; Hudska, K.; Pullmannova, P.; Kovacik, A.; Palat, K.; Roh, J.; Fleddermann, J.; Estrela-Lopis, I.; Vavrova, K. Different phase behavior and packing of ceramides with long (C16) and very long (C24) acyls in model membranes: Infrared spectroscopy using deuterated lipids. J. Phys. Chem. B 2014, 118, 10460–10470. [Google Scholar] [CrossRef] [PubMed]
- Pullmannová, P.; Pavlíková, L.; Kováčik, A.; Sochorová, M.; Školová, B.; Slepička, P.; Maixner, J.; Zbytovská, J.; Vávrová, K. Permeability and microstructure of model stratum corneum lipid membranes containing ceramides with long (C16) and very long (C24) acyl chains. Biophys. Chem. 2017, 224, 20–31. [Google Scholar] [CrossRef] [PubMed]
- Kovacik, A.; Roh, J.; Vavrova, K. The chemistry and biology of 6-hydroxyceramide, the youngest member of the human sphingolipid family. ChemBioChem 2014, 15, 1555–1562. [Google Scholar] [CrossRef] [PubMed]
- Rerek, M.E.; Van Wyck, D.; Mendelsohn, R.; Moore, D.J. FTIR spectroscopic studies of lipid dynamics in phytosphingosine ceramide models of the stratum corneum lipid matrix. Chem. Phys. Lipids 2005, 134, 51–58. [Google Scholar] [CrossRef]
- Rerek, M.E.; Chen, H.-C.; Markovic, B.; Van Wyck, D.; Garidel, P.; Mendelsohn, A.R.; Moore, D. Phytosphingosine and Sphingosine Ceramide Headgroup Hydrogen Bonding: Structural Insights through Thermotropic Hydrogen/Deuterium Exchange. J. Phys. Chem. B 2001, 105, 9355–9362. [Google Scholar] [CrossRef]
- Skolova, B.; Kovacik, A.; Tesar, O.; Opalka, L.; Vavrova, K. Phytosphingosine, sphingosine and dihydrosphingosine ceramides in model skin lipid membranes: Permeability and biophysics. BBA Biomembr. 2017, 1859, 824–834. [Google Scholar] [CrossRef]
- Kováčik, A.; Šilarová, M.; Pullmannová, P.; Maixner, J.; Vávrová, K. Effects of 6-Hydroxyceramides on the Thermotropic Phase Behavior and Permeability of Model Skin Lipid Membranes. Langmuir ACS J. Surf. Colloids 2017, 33, 2890–2899. [Google Scholar] [CrossRef] [PubMed]
- t’Kindt, R.; Jorge, L.; Dumont, E.; Couturon, P.; David, F.; Sandra, P.; Sandra, K. Profiling and characterizing skin ceramides using reversed-phase liquid chromatography-quadrupole time-of-flight mass spectrometry. Anal. Chem. 2012, 84, 403–411. [Google Scholar] [CrossRef]
- Uchida, Y.; Hama, H.; Alderson, N.L.; Douangpanya, S.; Wang, Y.; Crumrine, D.A.; Elias, P.M.; Holleran, W.M. Fatty acid 2-hydroxylase, encoded by FA2H, accounts for differentiation-associated increase in 2-OH ceramides during keratinocyte differentiation. J. Biol. Chem. 2007, 282, 13211–13219. [Google Scholar] [CrossRef] [Green Version]
- Hama, H. Fatty acid 2-Hydroxylation in mammalian sphingolipid biology. Biochim. Biophys. Acta 2010, 1801, 405–414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rabionet, M.; Gorgas, K.; Sandhoff, R. Ceramide synthesis in the epidermis. Biochim. Biophys. Acta 2014, 1841, 422–434. [Google Scholar] [CrossRef]
- Mislow, K.; Bleicher, S. The Configuration of Cerebronic Acid. J. Am. Chem. Soc. 1954, 76, 2825–2826. [Google Scholar] [CrossRef]
- Raudenkolb, S.; Wartewig, S.; Neubert, R.H.H. Polymorphism of ceramide 6: A vibrational spectroscopic and X-ray powder diffraction investigation of the diastereomers of N-(α-hydroxyoctadecanoyl)-phytosphingosine. Chem. Phys. Lipids 2005, 133, 89–102. [Google Scholar] [CrossRef] [PubMed]
- Garidel, P. The thermotropic phase behaviour of phyto-ceramide 1 as investigated by ATR-FTIR and DSC. Phys. Chem. Chem. Phys. 2002, 4, 2714–2720. [Google Scholar] [CrossRef]
- Garidel, P.; Folting, B.; Schaller, I.; Kerth, A. The microstructure of the stratum corneum lipid barrier: Mid-infrared spectroscopic studies of hydrated ceramide:palmitic acid:cholesterol model systems. Biophys. Chem. 2010, 150, 144–156. [Google Scholar] [CrossRef] [PubMed]
- Ruettinger, A.; Kiselev, M.A.; Hauss, T.; Dante, S.; Balagurov, A.; Neubert, R. Fatty acid interdigitation in stratum corneum model membranes: A neutron diffraction study. Eur. Biophys. J. 2008, 37, 759–771. [Google Scholar] [CrossRef] [PubMed]
- Schröter, A.; Kessner, D.; Kiselev, M.A.; Hauß, T.; Dante, S.; Neubert, R.H. Basic nanostructure of stratum corneum lipid matrices based on ceramides [EOS] and [AP]: A neutron diffraction study. Biophys. J. 2009, 97, 1104–1114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kessner, D.; Ruettinger, A.; Kiselev, M.; Wartewig, S.; Neubert, R. Properties of ceramides and their impact on the stratum corneum structure. Skin Pharmacol. Physiol. 2008, 21, 58–74. [Google Scholar] [CrossRef]
- Schmitt, T.; Lange, S.; Sonnenberger, S.; Dobner, B.; Deme, B.; Neubert, R.H.H.; Gooris, G.; Bouwstra, J.A. Determination of the influence of C24 D/(2R)- and L/(2S)-isomers of the CER[AP] on the lamellar structure of stratum corneum model systems using neutron diffraction. Chem. Phys. Lipids 2017, 209, 29–36. [Google Scholar] [CrossRef]
- Mojumdar, E.H.; Kariman, Z.; van Kerckhove, L.; Gooris, G.S.; Bouwstra, J.A. The role of ceramide chain length distribution on the barrier properties of the skin lipid membranes. Biochim. Biophys. Acta 2014, 1838, 2473–2483. [Google Scholar] [CrossRef] [Green Version]
- Bouwstra, J.A.; Gooris, G.S.; van der Spek, J.A.; Bras, W. Structural investigations of human stratum corneum by small-angle X-ray scattering. J. Investig. Dermatol. 1991, 97, 1005–1012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouwstra, J.; Gooris, G.; Salomons-de Vries, M.; Van der Spek, J.; Bras, W. Structure of human stratum corneum as a function of temperature and hydration: A wide-angle X-ray diffraction study. Int. J. Pharm. 1992, 84, 205–216. [Google Scholar] [CrossRef]
- de Jager, M.W.; Gooris, G.S.; Dolbnya, I.P.; Bras, W.; Ponec, M.; Bouwstra, J.A. The phase behaviour of skin lipid mixtures based on synthetic ceramides. Chem. Phys. Lipids 2003, 124, 123–134. [Google Scholar] [CrossRef]
- Dahlén, B.; Pascher, I. Molecular arrangements in sphingolipids. Thermotropic phase behaviour of tetracosanoylphytosphingosine. Chem. Phys. Lipids 1979, 24, 119–133. [Google Scholar] [CrossRef]
- Dahlen, B.; Pascher, I. Molecular arrangements in sphingolipids. Crystal structure of N-tetracosanoylphytosphingosine. Acta Crystallogr. Sect. B 1972, 28, 2396–2404. [Google Scholar] [CrossRef]
- Mendelsohn, R.; Moore, D.J. Vibrational spectroscopic studies of lipid domains in biomembranes and model systems. Chem. Phys. Lipids 1998, 96, 141–157. [Google Scholar] [CrossRef]
- Snyder, R.G.; Schachtschneider, J.H. Vibrational analysis of the n-paraffins—I: Assignments of infrared bands in the spectra of C3H8 through n-C19H40. Spectrochim. Acta 1963, 19, 85–116. [Google Scholar] [CrossRef]
- Kováčik, A.; Pullmannová, P.; Maixner, J.; Vávrová, K. Effects of Ceramide and Dihydroceramide Stereochemistry at C-3 on the Phase Behavior and Permeability of Skin Lipid Membranes. Langmuir ACS J. Surf. Colloids 2018, 34, 521–529. [Google Scholar] [CrossRef] [PubMed]
- Pascher, I. Molecular arrangements in sphingolipids Conformation and hydrogen bonding of ceramide and their implication on membrane stability and permeability. BBA Biomembr. 1976, 455, 433–451. [Google Scholar] [CrossRef]
- van Smeden, J.; Boiten, W.A.; Hankemeier, T.; Rissmann, R.; Bouwstra, J.A.; Vreeken, R.J. Combined LC/MS-platform for analysis of all major stratum corneum lipids, and the profiling of skin substitutes. Biochim. Biophys. Acta 2014, 1841, 70–79. [Google Scholar] [CrossRef]
- Opálka, L.; Kováčik, A.; Maixner, J.; Vávrová, K. Omega-O-Acylceramides in Skin Lipid Membranes: Effects of Concentration, Sphingoid Base, and Model Complexity on Microstructure and Permeability. Langmuir ACS J. Surf. Colloids 2016, 32, 12894–12904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schroeter, A.; Stahlberg, S.; Skolova, B.; Sonnenberger, S.; Eichner, A.; Huster, D.; Vavrova, K.; Hauss, T.; Dobner, B.; Neubert, R.H.; et al. Phase separation in ceramide[NP] containing lipid model membranes: Neutron diffraction and solid-state NMR. Soft Matter 2017, 13, 2107–2119. [Google Scholar] [CrossRef]
- Engelbrecht, T.N.; Schroeter, A.; Hauß, T.; Demé, B.; Scheidt, H.A.; Huster, D.; Neubert, R.H. The impact of ceramides NP and AP on the nanostructure of stratum corneum lipid bilayer. Part I: Neutron diffraction and 2H NMR studies on multilamellar models based on ceramides with symmetric alkyl chain length distribution. Soft Matter 2012, 8, 6599–6607. [Google Scholar] [CrossRef]
- Uche, L.E.; Gooris, G.S.; Beddoes, C.M.; Bouwstra, J.A. New insight into phase behavior and permeability of skin lipid models based on sphingosine and phytosphingosine ceramides. BBA Biomembr. 2019, 1861, 1317–1328. [Google Scholar] [CrossRef] [PubMed]
- Kováčik, A.; Vogel, A.; Adler, J.; Pullmannová, P.; Vávrová, K.; Huster, D. Probing the role of ceramide hydroxylation in skin barrier lipid models by 2H solid-state NMR spectroscopy and X-ray powder diffraction. BBA Biomembr. 2018, 1860, 1162–1170. [Google Scholar] [CrossRef] [PubMed]
- Pullmannova, P.; Ermakova, E.; Kovacik, A.; Opalka, L.; Maixner, J.; Zbytovska, J.; Kucerka, N.; Vavrova, K. Long And Very Long Lamellar Phases In Model Stratum Corneum Lipid Membranes. J. Lipid Res. 2019, 60, 963–971. [Google Scholar] [CrossRef]
- Caussin, J.; Gooris, G.S.; Janssens, M.; Bouwstra, J.A. Lipid organization in human and porcine stratum corneum differs widely, while lipid mixtures with porcine ceramides model human stratum corneum lipid organization very closely. BBA Biomembr. 2008, 1778, 1472–1482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wertz, P.W.; Downing, D.T. Ceramides of pig epidermis: Structure determination. J. Lipid Res. 1983, 24, 759–765. [Google Scholar] [CrossRef]
- Wertz, P.W.; Downing, D.T. Acylglucosylceramides of pig epidermis: Structure determination. J. Lipid Res. 1983, 24, 753–758. [Google Scholar] [CrossRef]
- Ponec, M.; Weerheim, A.; Lankhorst, P.; Wertz, P. New Acylceramide in Native and Reconstructed Epidermis. J. Investig. Dermatol. 2003, 120, 581–588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Groen, D.; Gooris, G.S.; Bouwstra, J.A. Model membranes prepared with ceramide EOS, cholesterol and free fatty acids form a unique lamellar phase. Langmuir ACS J. Surf. Colloids 2010, 26, 4168–4175. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Membrane Name | Model Membrane Composition |
|---|---|
| Cer NS | Cer NS/FFAs/Chol (1:1:1 mol) + 5 wt% of CholS |
| Cer (R)-AS | Cer (R)-AS/FFAs/Chol (1:1:1 mol) + 5 wt% of CholS |
| Cer (S)-AS | Cer (S)-AS/FFAs/Chol (1:1:1 mol) + 5 wt% of CholS |
| Cer NdS | Cer NdS/FFAs/Chol (1:1:1 mol) + 5 wt% of CholS |
| Cer (R)-AdS | Cer (R)-AdS/FFAs/Chol (1:1:1 mol) + 5 wt% of CholS |
| Cer (S)-AdS | Cer (S)-AdS/FFAs/Chol (1:1:1 mol) + 5 wt% of CholS |
| Cer NP | Cer NP/FFAs/Chol (1:1:1 mol) + 5 wt% of CholS |
| Cer (R)-AP | Cer (R)-AP/FFAs/Chol (1:1:1 mol) + 5 wt% of CholS |
| Cer (S)-AP | Cer (S)-AP/FFAs/Chol (1:1:1 mol) + 5 wt% of CholS |
| Unlabeled Membranes | Membranes with d-FFA [°C] | ||
|---|---|---|---|
| [°C] | Cer Chains | d-FFA Chains | |
| Cer NS | 57 (37) | 55 | 55 |
| Cer (R)-AS | 63 (63) | 61 | 59 |
| Cer (S)-AS | 71 (69) | 67 | 59 |
| Cer NdS | 73 (65) | 83 | 59 |
| Cer (R)-AdS | 83 (65) | 95 | 59 |
| Cer (S)-AdS | 79 (77) | 89 | 63 |
| Cer NP | 81 (60) | 89 | 58 |
| Cer (R)-AP | 79 (60) | 79 | 59 |
| Cer (S)-AP | 75 (74) | 71 | 69 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kováčik, A.; Pullmannová, P.; Opálka, L.; Šilarová, M.; Maixner, J.; Vávrová, K. Effects of (R)- and (S)-α-Hydroxylation of Acyl Chains in Sphingosine, Dihydrosphingosine, and Phytosphingosine Ceramides on Phase Behavior and Permeability of Skin Lipid Models. Int. J. Mol. Sci. 2021, 22, 7468. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22147468
Kováčik A, Pullmannová P, Opálka L, Šilarová M, Maixner J, Vávrová K. Effects of (R)- and (S)-α-Hydroxylation of Acyl Chains in Sphingosine, Dihydrosphingosine, and Phytosphingosine Ceramides on Phase Behavior and Permeability of Skin Lipid Models. International Journal of Molecular Sciences. 2021; 22(14):7468. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22147468
Chicago/Turabian StyleKováčik, Andrej, Petra Pullmannová, Lukáš Opálka, Michaela Šilarová, Jaroslav Maixner, and Kateřina Vávrová. 2021. "Effects of (R)- and (S)-α-Hydroxylation of Acyl Chains in Sphingosine, Dihydrosphingosine, and Phytosphingosine Ceramides on Phase Behavior and Permeability of Skin Lipid Models" International Journal of Molecular Sciences 22, no. 14: 7468. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22147468