
Combined Exposure to Diazinon and Nicotine Exerts a Synergistic Adverse Effect In Vitro and Disrupts Brain Development and Behaviors In Vivo




Abstract
:1. Introduction
2. Results
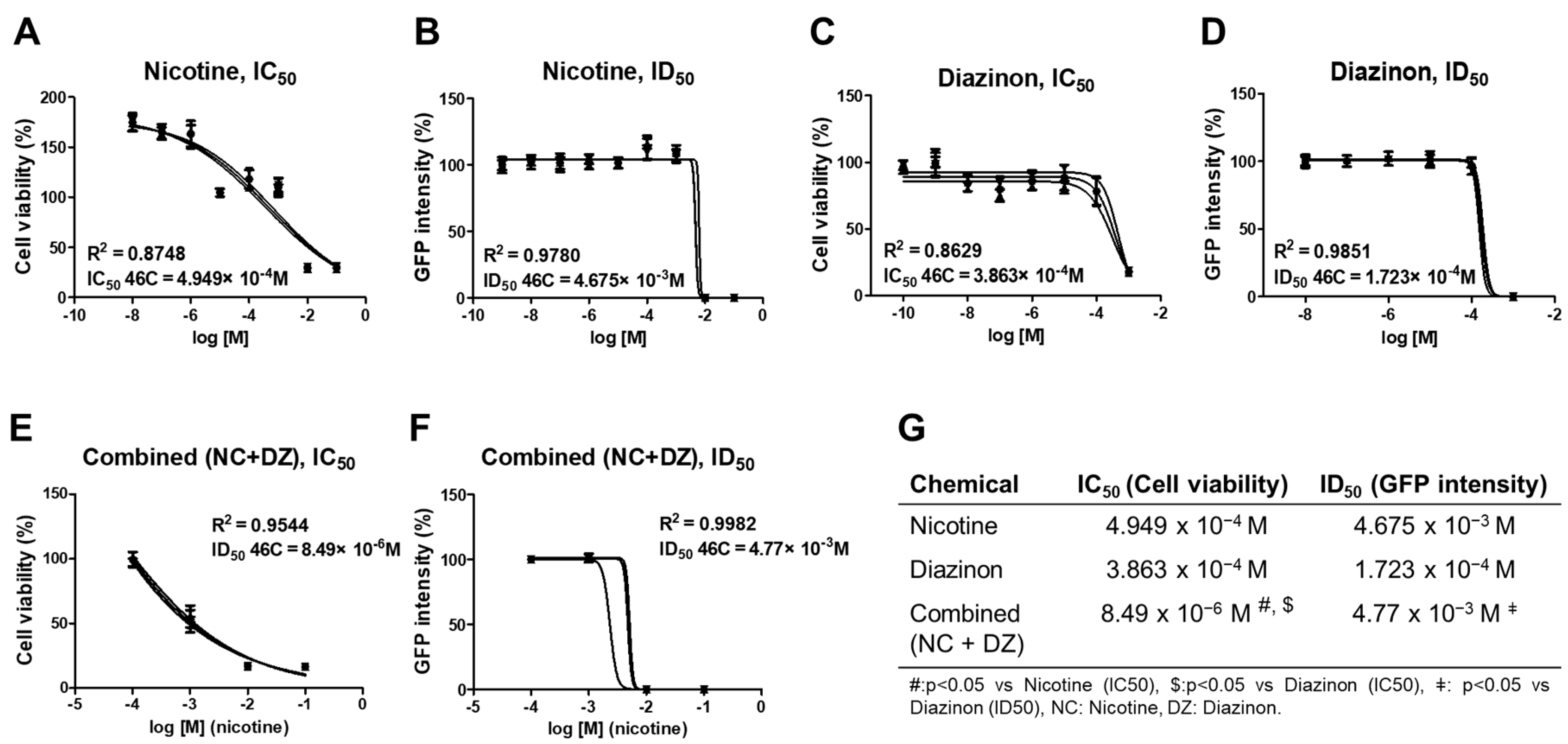
2.1. Combined Administration of Nicotine and Diazinon Exerted a Synergistic Adverse Effect on Cell Viability of Sox1-GFP Cells
2.2. Combined Administration of Nicotine and Diazinon Resulted in Abnormal Locomotor Function and Motor Learning Impairment
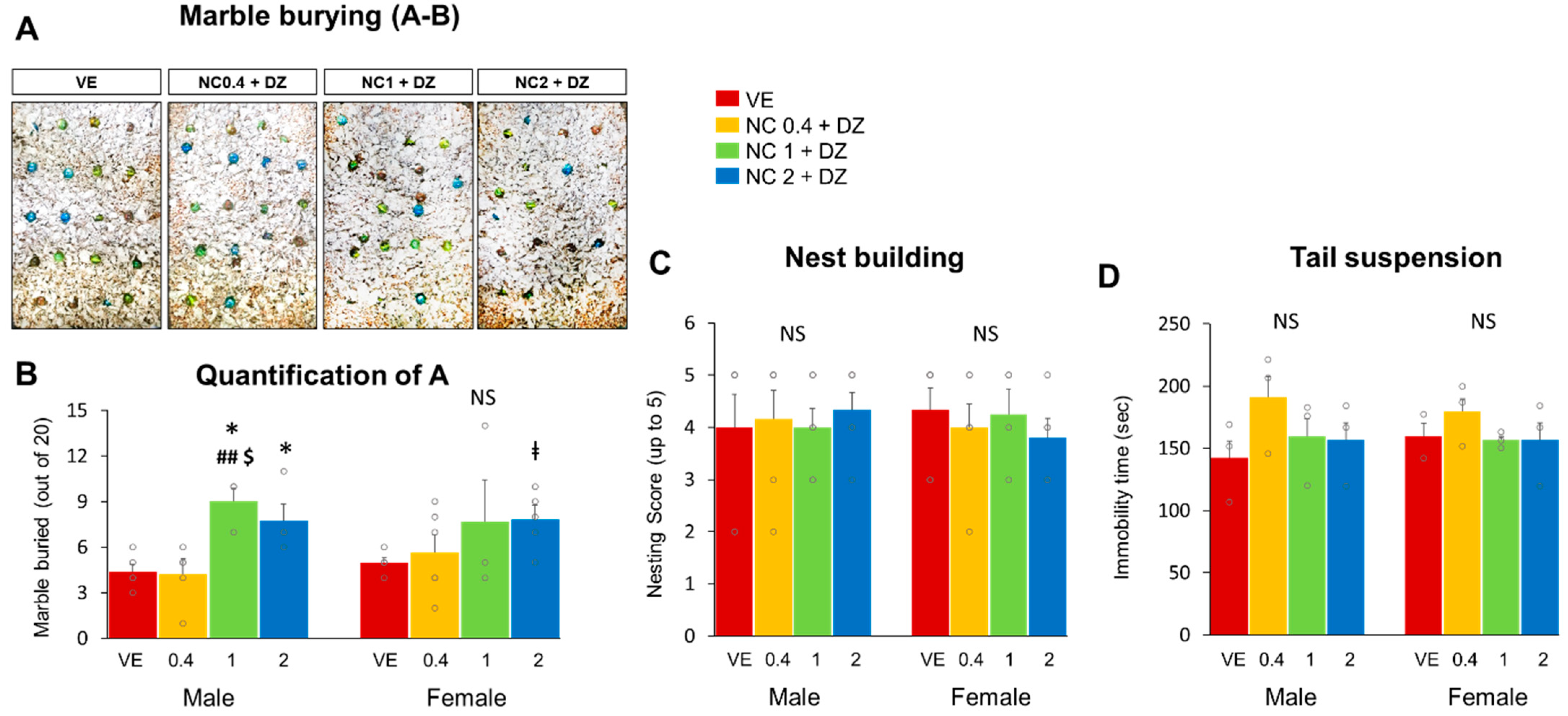
2.3. Combined Administration of Nicotine and Diazinon Partially Increased Compulsive-like Behavior in Mice, but Did Not Affect Depression-Related Behaviors
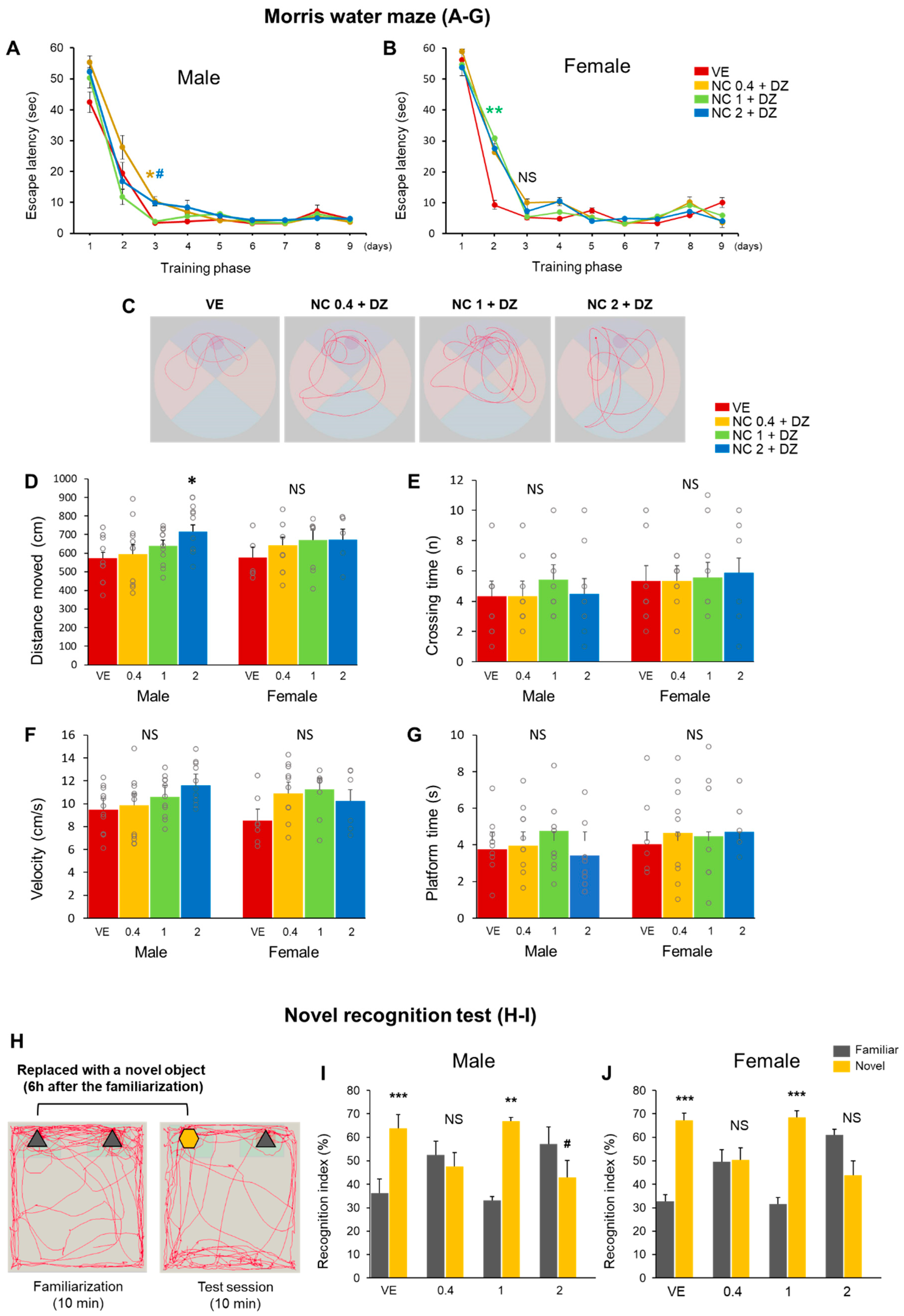
2.4. Combined Exposure of Nicotine and Diazinon Disrupted Spatial Learning and Cognition-Related Behaviors in Mouse Offfspring
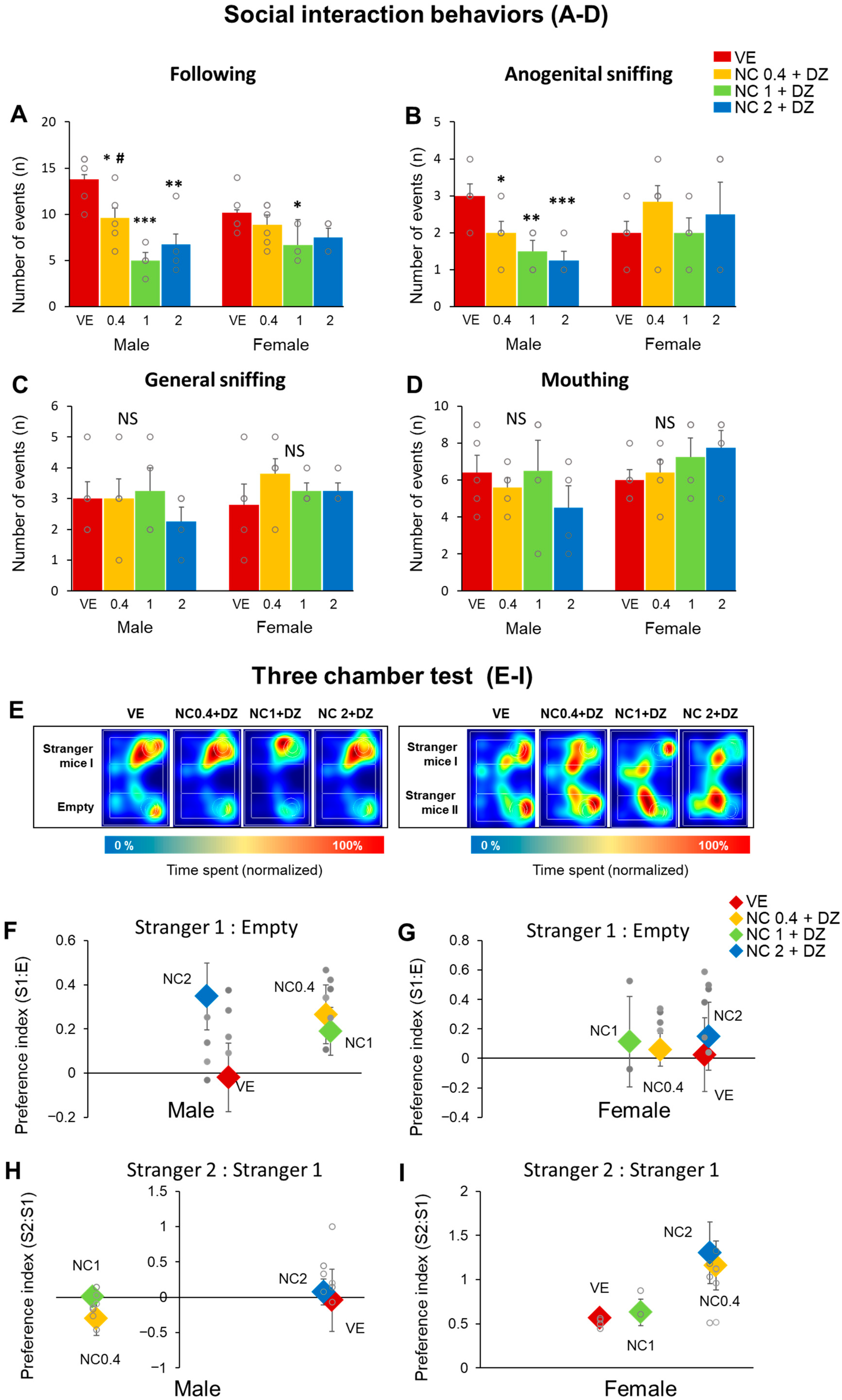
2.5. Simultaneous Exposure to Nicotine and Diazinon Disturbed Sociability-Related Behaviors, but Seldom Affected on the Animals’ Social Novelty Recognition
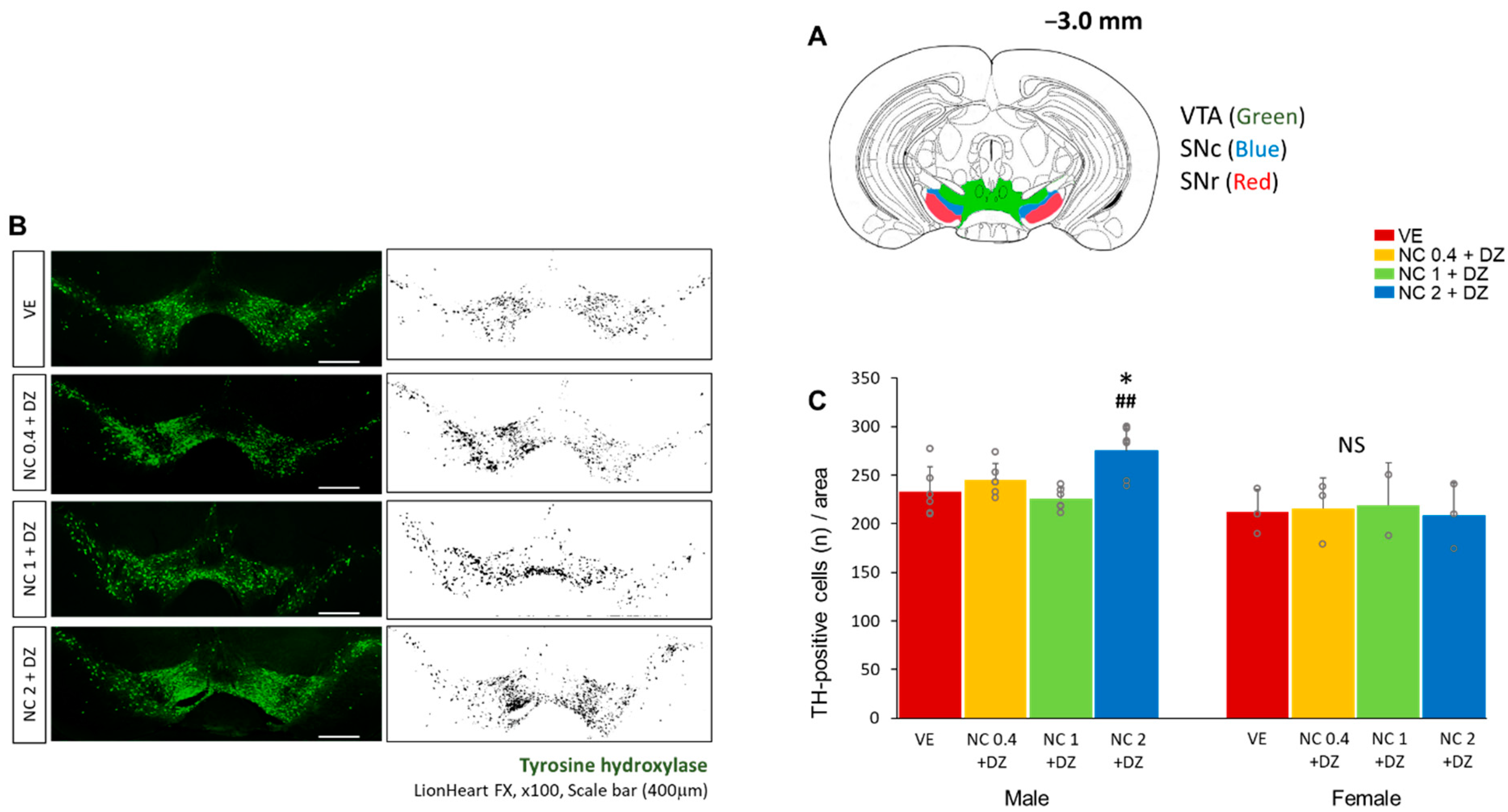
2.6. Combined Exposure of Nicotine and Diazinon Increased the Density of the Dopaminergic Neuron in Ventral Tegmental Area in Male Mice
3. Discussion
4. Materials and Methods
4.1. Mouse Embryonic Stem Cell Culture
4.2. Cell Viability Assay
4.3. Assessment for Neural Differentiation in Sox1-GFP Cells
4.4. Animals and Drug Treatments
4.5. Immunohistochemistry
4.6. Measurement of the TH-Positive Cells on the Ventral Tegmental Area
4.7. Experimental Behavior Analysis
4.8. Rotarod Test
4.9. Open Field Test
4.10. Marble Burying Test
4.11. Nest Building Test
4.12. Tail Suspension
4.13. Morris Water Maze
4.14. Nobel Object Recognition Test
4.15. Three-Chamber Sociability Test
4.16. Social Interaction
4.17. Data Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Marins, A.T.; Cerezer, C.; Leitemperger, J.W.; Severo, E.S.; Costa, M.D.; Fontoura, D.O.; Nunes, M.E.M.; Ribeiro, L.C.; Zanella, R.; Loro, V.L. A mixture of pesticides at environmental concentrations induces oxidative stress and cholinergic effects in the neotropical fish Rhamdia quelen. Ecotoxicology 2021, 30, 164–174. [Google Scholar] [CrossRef] [PubMed]
- Youbin, Q.; Chengzhi, C.; Yan, T.; Xuejun, J.; Chongying, Q.; Bin, P.; Baijie, T. The synergistic effect of benzo[a]pyrene and lead on learning and memory of mice. Toxicol. Ind. Health 2012, 29, 387–395. [Google Scholar] [CrossRef] [PubMed]
- Dwivedi, N.; Bhutia, Y.D.; Kumar, V.; Yadav, P.; Kushwaha, P.; Swarnkar, H.; Flora, S.J.S. Effects of combined exposure to dichlorvos and monocrotophos on blood and brain biochemical variables in rats. Hum. Exp. Toxicol. 2009, 29, 121–129. [Google Scholar] [CrossRef]
- Boussabbeh, M.; Ben Salem, I.; Hamdi, M.; Ben Fradj, S.; Abid-Essefi, S.; Bacha, H. Diazinon, an organophosphate pesticide, induces oxidative stress and genotoxicity in cells deriving from large intestine. Environ. Sci. Pollut. Res. 2016, 23, 2882–2889. [Google Scholar] [CrossRef]
- Dani, J.A. Overview of nicotinic receptors and their roles in the central nervous system. Biol. Psychiatry 2001, 49, 166–174. [Google Scholar] [CrossRef]
- Barraud, P.; Thompson, L.; Kirik, D.; Björklund, A.; Parmar, M. Isolation and characterization of neural precursor cells from theSox1-GFP reporter mouse. Eur. J. Neurosci. 2005, 22, 1555–1569. [Google Scholar] [CrossRef] [PubMed]
- Ernst, M.; Moolchan, E.T.; Robinson, M.L. Behavioral and Neural Consequences of Prenatal Exposure to Nicotine. J. Am. Acad. Child Adolesc. Psychiatry 2001, 40, 630–641. [Google Scholar] [CrossRef] [PubMed]
- Antunes, M.; Biala, G. The novel object recognition memory: Neurobiology, test procedure, and its modifications. Cogn. Process. 2011, 13, 93–110. [Google Scholar] [CrossRef] [Green Version]
- Kaidanovich-Beilin, O.; Lipina, T.; Vukobradovic, I.; Roder, J.; Woodgett, J.R. Assessment of Social Interaction Behaviors. J. Vis. Exp. 2011, e2473. [Google Scholar] [CrossRef] [PubMed]
- Morales, M.; Margolis, E.B. Ventral tegmental area: Cellular heterogeneity, connectivity and behaviour. Nat. Rev. Neurosci. 2017, 18, 73–85. [Google Scholar] [CrossRef] [PubMed]
- Gantz, S.; Ford, C.P.; Morikawa, H.; Williams, J.T. The Evolving Understanding of Dopamine Neurons in the Substantia Nigra and Ventral Tegmental Area. Annu. Rev. Physiol. 2018, 80, 219–241. [Google Scholar] [CrossRef]
- Mallozzi, M.; Bordi, G.; Garo, C.; Caserta, D. The effect of maternal exposure to endocrine disrupting chemicals on fetal and neonatal development: A review on the major concerns. Birth Defects Res. Part C 2016, 108, 224–242. [Google Scholar] [CrossRef]
- Aubert, J.; Stavridis, M.; Tweedie, S.; O’Reilly, M.; Vierlinger, K.; Li, M.; Ghazal, P.; Pratt, T.; Mason, J.; Roy, D.; et al. Screening for mammalian neural genes via fluorescence-activated cell sorter purification of neural precursors from Sox1-gfp knock-in mice. Proc. Natl. Acad. Sci. USA 2003, 100, 11836–11841. [Google Scholar] [CrossRef] [Green Version]
- Lee, A.; Picciotto, M.R. Perinatal nicotine exposure impairs learning of a skilled forelimb reaching task in male but not female adult mice. Behav. Brain Res. 2019, 367, 176–180. [Google Scholar] [CrossRef]
- Alkam, T.; Kim, H.-C.; Hiramatsu, M.; Mamiya, T.; Aoyama, Y.; Nitta, A.; Yamada, K.; Nabeshima, T. Evaluation of emotional behaviors in young offspring of C57BL/6J mice after gestational and/or perinatal exposure to nicotine in six different time-windows. Behav. Brain Res. 2013, 239, 80–89. [Google Scholar] [CrossRef]
- Li, J.; Bo, L.; Zhang, P.; Gao, Q.; Li, L.; Tang, J.; Wu, C.; Li, D.; Xiao, J.; Chen, J.; et al. Exposure to Nicotine During Pregnancy and Altered Learning and Memory in the Rat Offspring. Nicotine Tob. Res. 2014, 17, 661–666. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Tao, X.; Pang, G.; Mu, M.; Sun, Q.; Liu, F.; Hu, Y.; Tao, H.; Li, B.; Xu, K. Maternal Nicotine Exposure Alters Hippocampal Microglia Polarization and Promotes Anti-inflammatory Signaling in Juvenile Offspring in Mice. Front. Pharmacol. 2021, 12, 661304. [Google Scholar] [CrossRef] [PubMed]
- Cross, S.J.; Linker, K.E.; Leslie, F.M. Sex-dependent effects of nicotine on the developing brain. J. Neurosci. Res. 2017, 95, 422–436. [Google Scholar] [CrossRef] [Green Version]
- Rebuli, M.E.; Patisaul, H.B. Assessment of sex specific endocrine disrupting effects in the prenatal and pre-pubertal rodent brain. J. Steroid Biochem. Mol. Biol. 2016, 160, 148–159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vatanparast, J.; Naseh, M.; Baniasadi, M.; Haghdoost-Yazdi, H. Developmental exposure to chlorpyrifos and diazinon differentially affect passive avoidance performance and nitric oxide synthase-containing neurons in the basolateral complex of the amygdala. Brain Res. 2013, 1494, 17–27. [Google Scholar] [CrossRef] [PubMed]
- Alcaro, A.; Huber, R.; Panksepp, J. Behavioral functions of the mesolimbic dopaminergic system: An affective neuroethological perspective. Brain Res. Rev. 2007, 56, 283–321. [Google Scholar] [CrossRef] [Green Version]
- Keller, R.F.; Kanlikilicer, P.; Dragomir, A.; Fan, Y.; Akay, Y.M.; Akay, M. Investigating the Effect of Perinatal Nicotine Exposure on Dopaminergic Neurons in the VTA Using miRNA Expression Profiles. IEEE Trans. Nanobiosci. 2017, 16, 843–849. [Google Scholar] [CrossRef] [PubMed]
- Hegarty, S.V.; Sullivan, A.; O’Keeffe, G. Midbrain dopaminergic neurons: A review of the molecular circuitry that regulates their development. Dev. Biol. 2013, 379, 123–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lai, I.-C.; Dulcis, D. Nicotine-induced dopamine plasticity: A gateway to neurotransmitter replacement? Neural Regen. Res. 2020, 15, 73–74. [Google Scholar] [CrossRef] [PubMed]
- Nicholatos, J.W.; Francisco, A.; Bender, C.A.; Yeh, T.; Lugay, F.J.; Salazar, J.E.; Glorioso, C.; Libert, S. Nicotine promotes neuron survival and partially protects from Parkinson’s disease by suppressing SIRT6. Acta Neuropathol. Commun. 2018, 6, 120. [Google Scholar] [CrossRef]
- Chang, G.-Q.; Karatayev, O.; Leibowitz, S.F. Prenatal exposure to nicotine stimulates neurogenesis of orexigenic peptide-expressing neurons in hypothalamus and amygdala. J. Neurosci. 2013, 33, 13600–13611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brooks, A.; Henderson, B. Systematic Review of Nicotine Exposure’s Effects on Neural Stem and Progenitor Cells. Brain Sci. 2021, 11, 172. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.-J.A.; King, K.A.; Lee, R.E.; Sedtal, C.S.; Smith, A.M. Effects of nicotine exposure during prenatal or perinatal period on cell numbers in adult rat hippocampus and cerebellum: A stereology study. Life Sci. 2006, 79, 2221–2227. [Google Scholar] [CrossRef]
- Jung, Y.; Hsieh, L.S.; Lee, A.M.; Zhou, Z.; Coman, D.; Heath, C.J.; Hyder, F.; Mineur, Y.S.; Yuan, Q.; Goldman, D.; et al. An epigenetic mechanism mediates developmental nicotine effects on neuronal structure and behavior. Nat. Neurosci. 2016, 19, 905–914. [Google Scholar] [CrossRef]
- Dwyer, J.B.; Cardenas, A.; Franke, R.M.; Chen, Y.; Bai, Y.; Belluzzi, J.D.; Lotfipour, S.; Leslie, F.M. Prenatal nicotine sex-dependently alters adolescent dopamine system development. Transl. Psychiatry 2019, 9, 1–11. [Google Scholar] [CrossRef]
- Muneoka, K.; Nakatsu, T.; Fuji, J.-I.; Ogawa, T.; Takigawa, M. Prenatal Administration of Nicotine Results in Dopaminergic Alterations in the Neocortex. Neurotoxicol. Teratol. 1999, 21, 603–609. [Google Scholar] [CrossRef]
- Mangas, I.; Estevez, J.; Vilanova, E.; França, T.C.C. New insights on molecular interactions of organophosphorus pesticides with esterases. Toxicology 2017, 376, 30–43. [Google Scholar] [CrossRef]
- Hawkey, A.; Pippen, E.; White, H.; Kim, J.; Greengrove, E.; Kenou, B.; Holloway, Z.; Levin, E.D. Gestational and perinatal exposure to diazinon causes long-lasting neurobehavioral consequences in the rat. Toxicology 2020, 429, 152327. [Google Scholar] [CrossRef] [PubMed]
- Neishabouri, E.; Hassan, Z.; Azizi, E.; Ostad, S. Evaluation of immunotoxicity induced by diazinon in C57bl/6 mice. Toxicology 2004, 196, 173–179. [Google Scholar] [CrossRef] [PubMed]
- Jeon, B.-H.; Lee, B.; Jung, E.-M.; Jeung, E.-B. Regulatory effect of dexamethasone on tracheal calcium processing proteins and mucosal secretion. Endocr. Abstr. 2019, 70. [Google Scholar] [CrossRef]
- Benowitz, N.L.; Iii, P.J. Daily intake of nicotine during cigarette smoking. Clin. Pharmacol. Ther. 1984, 35, 499–504. [Google Scholar] [CrossRef]
- Tran, D.N.; Jung, E.-M.; Yoo, Y.-M.; Jeung, E.-B. 4-tert-Octylphenol Exposure Disrupts Brain Development and Subsequent Motor, Cognition, Social, and Behavioral Functions. Oxidative Med. Cell. Longev. 2020, 2020, 8875604. [Google Scholar] [CrossRef]
- Jung, E.-M.; Moffat, J.J.; Liu, J.; Dravid, S.M.; Gurumurthy, C.B.; Kim, W.-Y. Arid1b haploinsufficiency disrupts cortical interneuron development and mouse behavior. Nat. Neurosci. 2017, 20, 1694–1707. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Test | Measurement | Number of Animals per Group (Sex) | Age (Old) |
|---|---|---|---|
| Rotarod | Latency to fall (s) | n = 10 (5 male, 5 female) for VE, n = 10 (5 male, 5 female) for NC0.4 + DZ, n = 7 (4 male, 3 female) for NC1 + DZ, n = 8 (4 male, 3 female) for NC2 + DZ | 6~8 weeks |
| Open field | Time in center (s) | n = 10 (5 male, 5 female) for VE, n = 10 (5 male, 5 female) for NC0.4 + DZ, n = 7 (4 male, 3 female) for NC1 + DZ, n = 8 (4 male, 3 female) for NC2 + DZ | 4 weeks |
| Frequency (n) | |||
| Velocity (cm/s) | |||
| Distance traveled (cm) | |||
| Marble burying | Number of marbles buried (n) | n = 10 (5 male, 5 female) for VE, n = 10 (5 male, 5 female) for NC0.4 + DZ, n = 7 (4 male, 3 female) for NC1 + DZ, n = 8 (4 male, 3 female) for NC2 + DZ | 9 weeks |
| Nest building | Nesting score | n = 7 for each group (4 male, 3 female) | 4 weeks |
| Tail suspension | Immobility time (s) | n = 7 for each group (4 male, 3 female) | 9 weeks |
| Morris water maze | Escape latency (s) | n = 8 (4 male, 4 female) for VE, n = 8 (4 male, 4 female) for NC0.4 + DZ, n = 7 (4 male, 3 female) for NC1 + DZ, n = 8 (4 male, 3 female) for NC2 + DZ | 9~10 weeks |
| Noble object recognition | Recognition index (%) | n = 10 (5 male, 5 female) for VE, n = 10 (5 male, 5 female) for NC0.4 + DZ, n = 7 (4 male, 3 female) for NC1 + DZ, n = 8 (4 male, 3 female) for NC2 + DZ | 5 weeks |
| Three-chamber sociability | Time spent in chamber (s) | n = 10 (5 male, 5 female) for VE, n = 10 (5 male, 5 female) for NC0.4 + DZ, n = 7 (4 male, 3 female) for NC1 + DZ, n = 8 (4 male, 3 female) for NC2 + DZ | 5 weeks |
| Social interaction | Following (n) Genital sniffing(n) Anogenital sniffing (n) Mouthing | n = 10 (5 male, 5 female) for VE, n = 10 (5 male, 5 female) for NC0.4 + DZ, n = 7 (4 male, 3 female) for NC1 + DZ, n = 8 (4 male, 3 female) for NC2 + DZ | 9 weeks |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, B.; Park, S.M.; Jeong, S.; Kim, K.; Jeung, E.-B. Combined Exposure to Diazinon and Nicotine Exerts a Synergistic Adverse Effect In Vitro and Disrupts Brain Development and Behaviors In Vivo. Int. J. Mol. Sci. 2021, 22, 7742. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22147742
Lee B, Park SM, Jeong S, Kim K, Jeung E-B. Combined Exposure to Diazinon and Nicotine Exerts a Synergistic Adverse Effect In Vitro and Disrupts Brain Development and Behaviors In Vivo. International Journal of Molecular Sciences. 2021; 22(14):7742. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22147742
Chicago/Turabian StyleLee, Bonn, Seon Mi Park, SunHwa Jeong, KangMin Kim, and Eui-Bae Jeung. 2021. "Combined Exposure to Diazinon and Nicotine Exerts a Synergistic Adverse Effect In Vitro and Disrupts Brain Development and Behaviors In Vivo" International Journal of Molecular Sciences 22, no. 14: 7742. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22147742