
Expression Silencing of Glutathione Peroxidase 4 in Mouse Erythroleukemia Cells Delays In Vitro Erythropoiesis


Abstract
:1. Introduction
2. Results
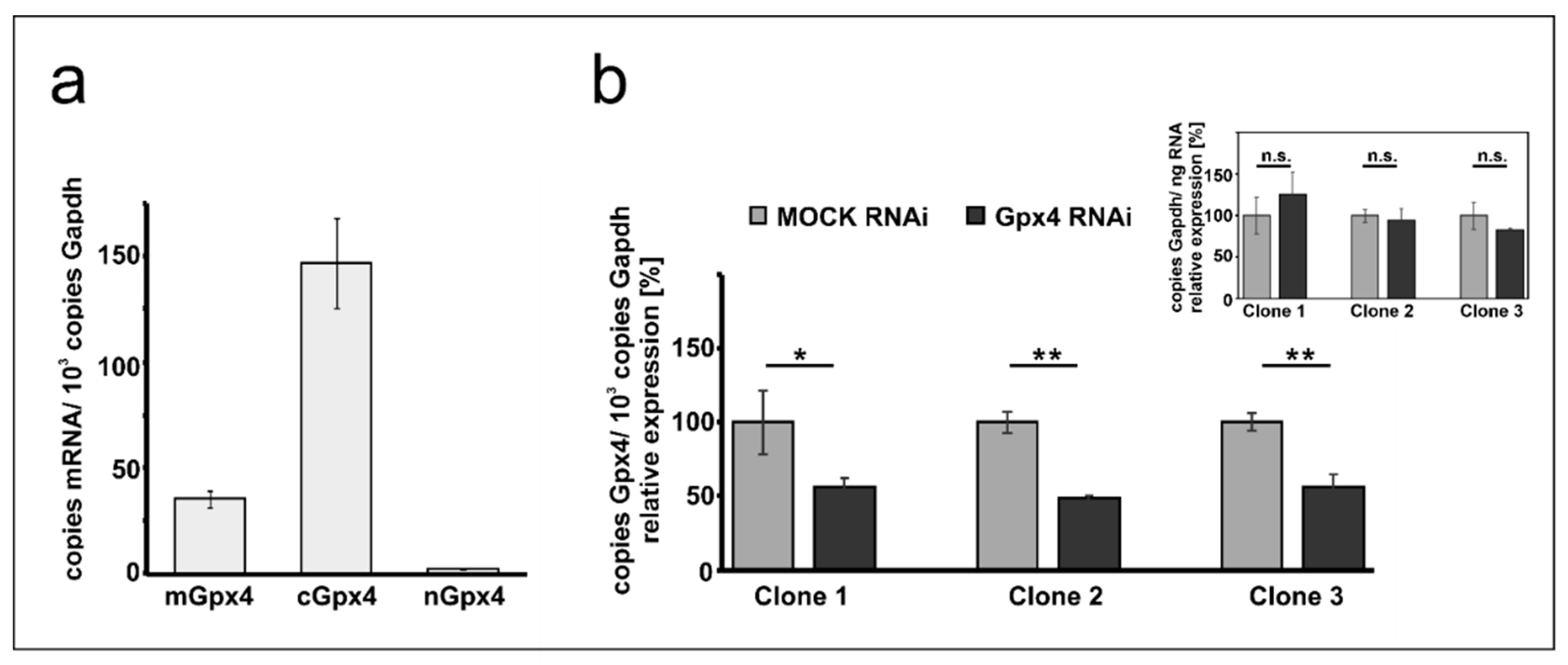
2.1. Expression of Gpx4 Isoforms in Mouse Erythroleukemia Cells
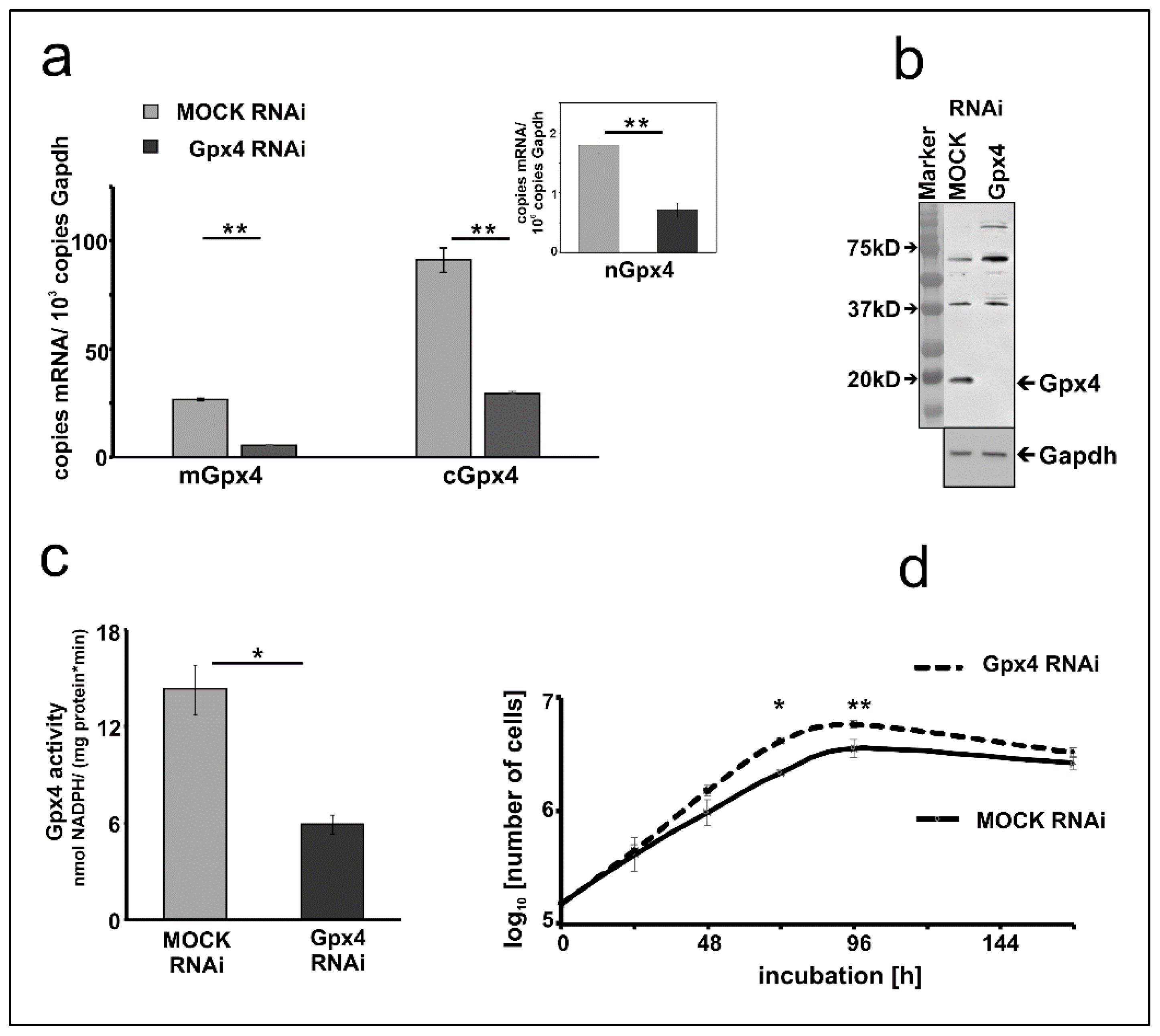
2.2. Stable Transfection of MEL Cells with Anti-Gpx4 Antisense Construct Reduced Gpx4 Expression
2.3. Basic Characterization of Gpx4 Compromised MEL Cells
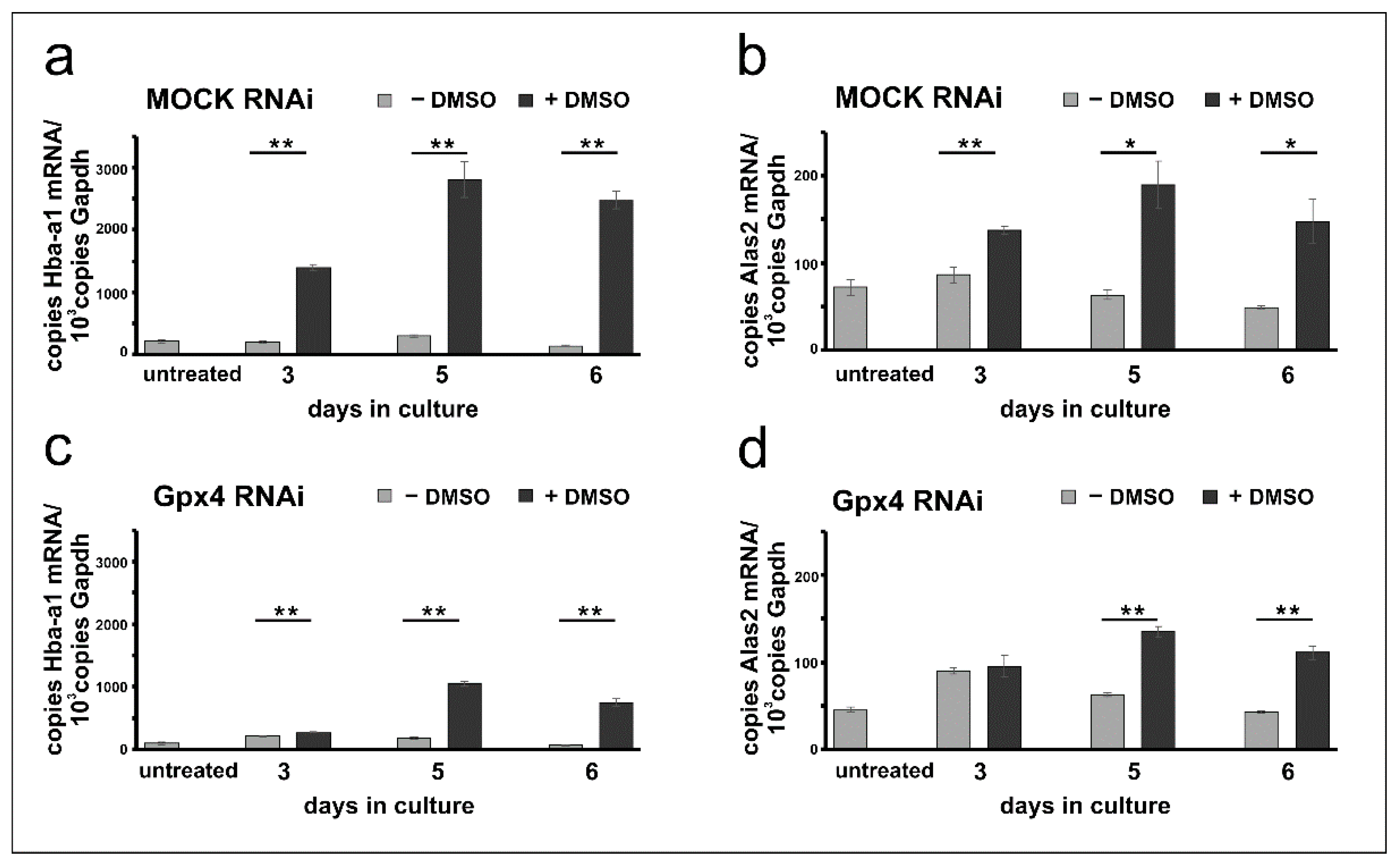
2.4. Erythropoiesis Can Be Induced in Mock-Transfected and Gpx4 siRNA Transfected MEL Cells
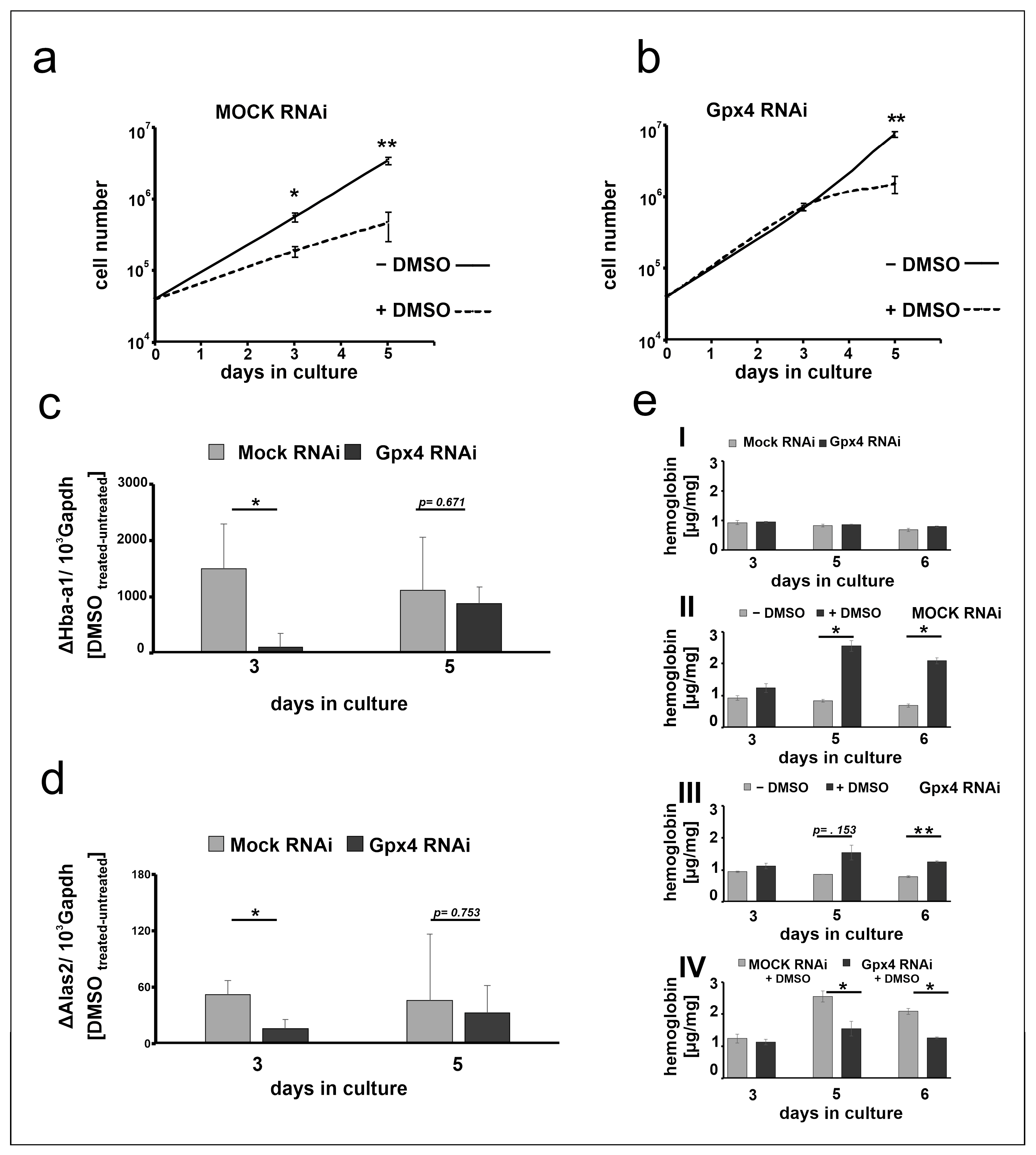
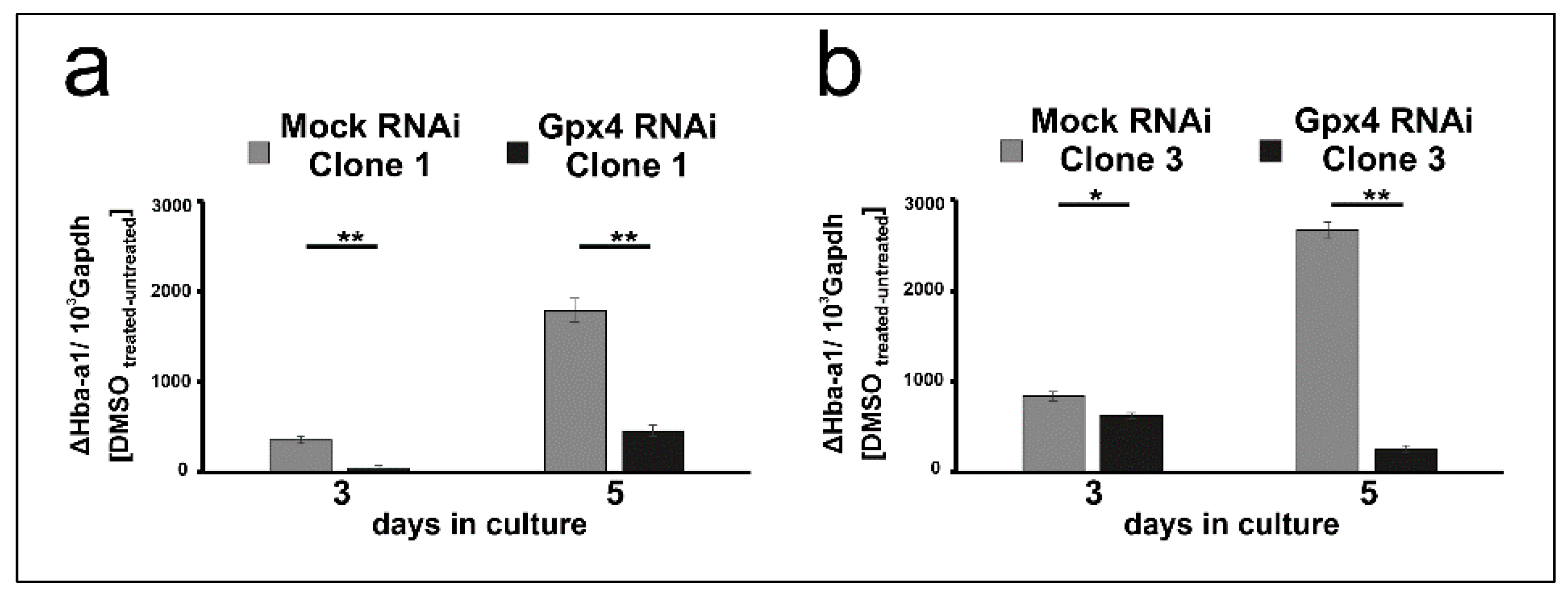
2.5. Stable Knockdown of Gpx4 Expression Delays DMSO-Induced Erythroid Differentiation
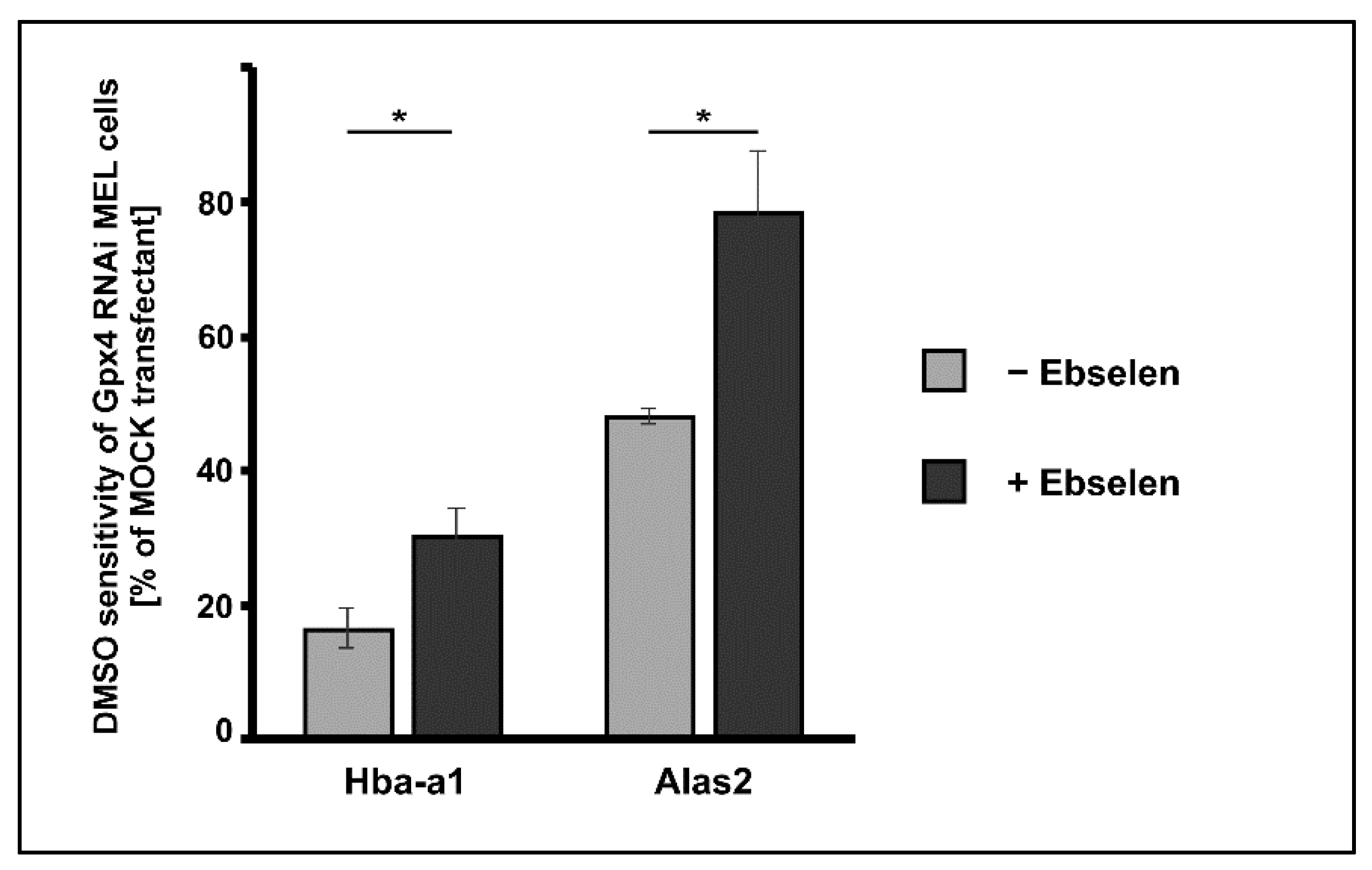
2.6. Delay of DMSO Induced In Vitro Erythropoiesis in MEL Cells Caused by Gpx4 Deficiency Was Partially Rescued by Ebselen
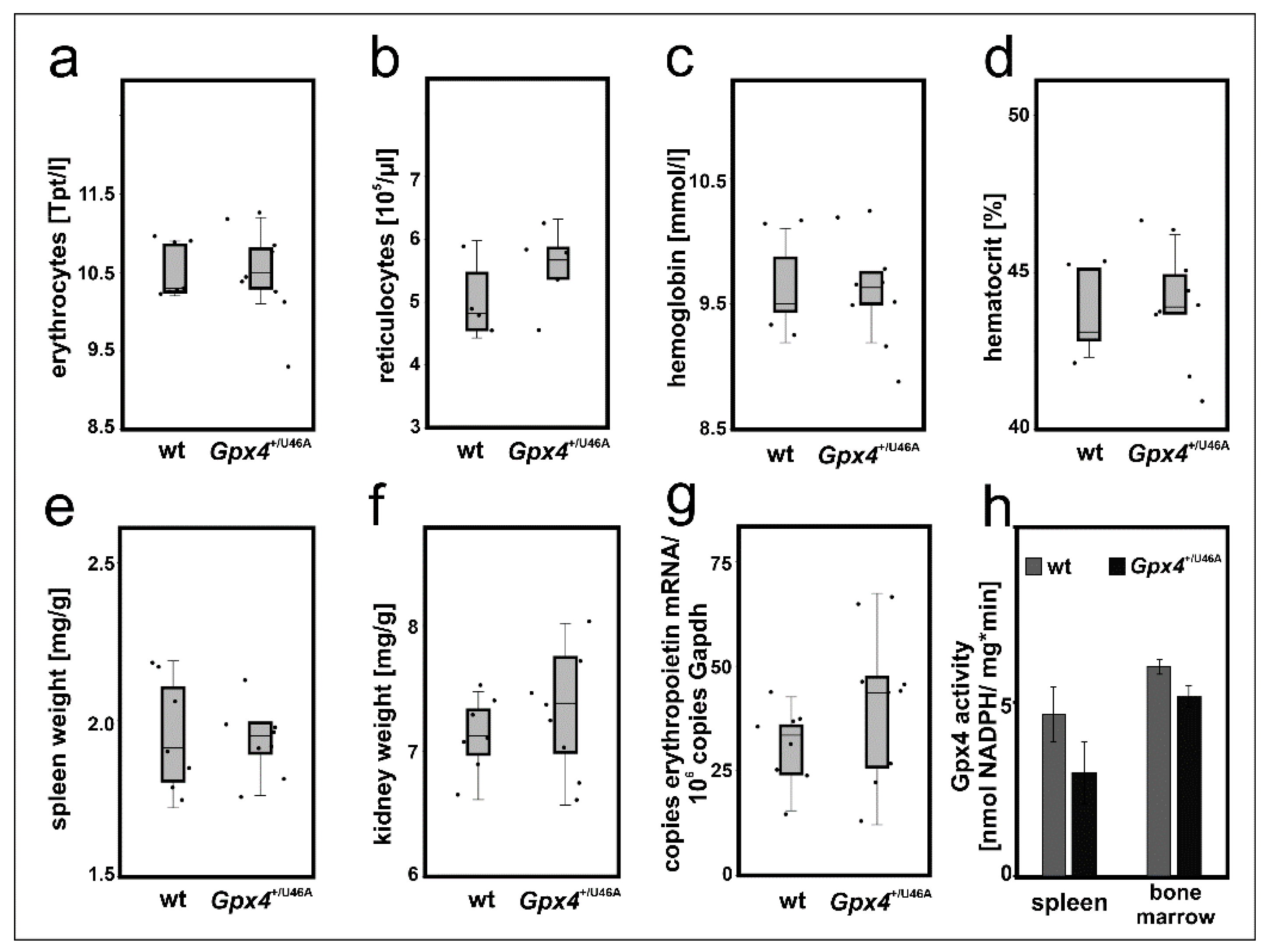
2.7. Heterozygous Expression of Catalytically Inactive Gpx4 in Mice Does Not Induce a Defective Erythropoietic Phenotype
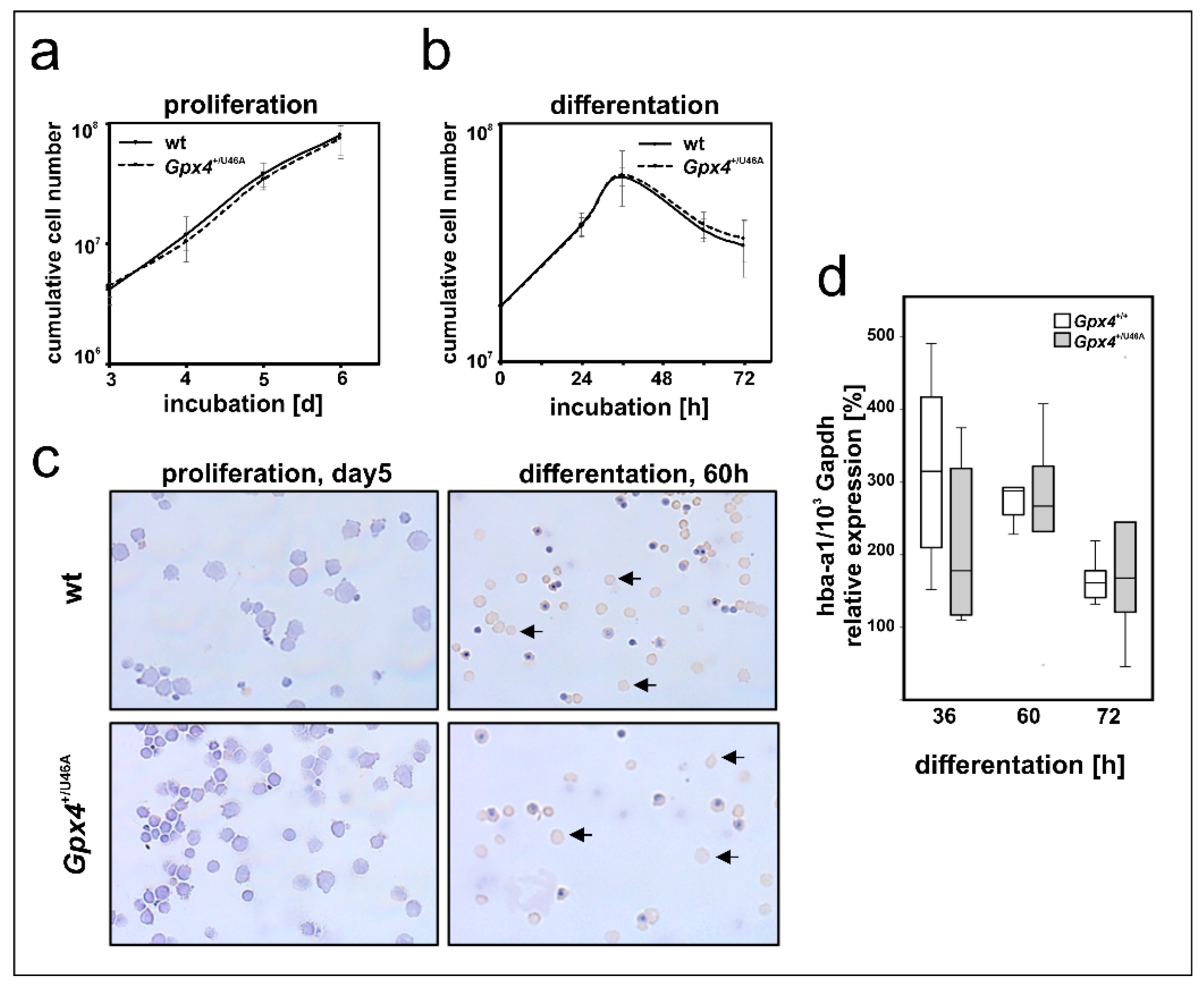
2.8. Heterozygous Expression of Catalytically Inactive Gpx4 in Mice Does Not Impact Ex Vivo Erythropoiesis of Embryonic Liver Derived Erythroblasts
3. Discussion
3.1. Gpx4 Isoforms Are Expressed in MEL Cells
3.2. Gpx4 and Erythropoiesis
3.3. Heterozygous Expression of a Catalytically Inactive Gpx4 Mutant Does Not Compromise In Vivo Erythropoiesis in Mice
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Animals, Breeding, and Isolation of Spleen and Bone Marrow
4.3. PCR Genotyping
4.4. MEL Cell Culture
4.5. Antisense Transfection and Selection Procedure
4.6. DMSO Induced Erythroid Differentiation in Stable MEL Cell Transfectants
4.7. RNA Isolation and Quantitative Real-Time PCR
4.8. Photometric Hemoglobin Determination
4.9. Immunoblotting
4.10. Gpx4 Activity Assay
4.11. Ex Vivo Erythropoiesis
4.11.1. Cultivation of Murine Erythroid Progenitors
4.11.2. Induction and Analysis of Terminal Differentiation
4.12. Blood Analysis and Organ Collection
4.13. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dzierzak, E.; Philipsen, S. Erythropoiesis: Development and differentiation. Cold Spring Harb. Perspect. Med. 2013, 3, a011601. [Google Scholar] [CrossRef]
- Nandakumar, S.K.; Ulirsch, J.C.; Sankaran, V.G. Advances in understanding erythropoiesis: Evolving perspectives. Br. J. Haematol. 2016, 173, 206–218. [Google Scholar] [CrossRef] [Green Version]
- Franco, R.S. Measurement of red cell lifespan and aging. Transfus. Med. Hemother. 2012, 39, 302–307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.H.; Crocker, P.R.; Westaby, S.; Key, N.; Mason, D.Y.; Gordon, S.; Weatherall, D.J. Isolation and immunocytochemical characterization of human bone marrow stromal macrophages in hemopoietic clusters. J. Exp. Med. 1988, 168, 1193–1198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moras, M.; Lefevre, S.D.; Ostuni, M.A. From Erythroblasts to Mature Red Blood Cells: Organelle Clearance in Mammals. Front. Physiol. 2017, 8, 1076. [Google Scholar] [CrossRef]
- Jacobsen, R.N.; Perkins, A.C.; Levesque, J.P. Macrophages and regulation of erythropoiesis. Curr. Opin. Hematol. 2015, 22, 212–219. [Google Scholar] [CrossRef] [PubMed]
- Sukhbaatar, N.; Weichhart, T. Iron Regulation: Macrophages in Control. Pharmaceuticals 2018, 11, 137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toppo, S.; Flohe, L.; Ursini, F.; Vanin, S.; Maiorino, M. Catalytic mechanisms and specificities of glutathione peroxidases: Variations of a basic scheme. Biochim. Biophys. Acta 2009, 1790, 1486–1500. [Google Scholar] [CrossRef] [PubMed]
- Margis, R.; Dunand, C.; Teixeira, F.K.; Margis-Pinheiro, M. Glutathione peroxidase family—An evolutionary overview. FEBS J. 2008, 275, 3959–3970. [Google Scholar] [CrossRef] [PubMed]
- Toppo, S.; Vanin, S.; Bosello, V.; Tosatto, S.C. Evolutionary and structural insights into the multifaceted glutathione peroxidase (Gpx) superfamily. Antioxid. Redox Signal. 2008, 10, 1501–1514. [Google Scholar] [CrossRef]
- Liang, H.; Yoo, S.E.; Na, R.; Walter, C.A.; Richardson, A.; Ran, Q. Short form glutathione peroxidase 4 is the essential isoform required for survival and somatic mitochondrial functions. J. Biol. Chem. 2009, 284, 30836–30844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Savaskan, N.E.; Ufer, C.; Kuhn, H.; Borchert, A. Molecular biology of glutathione peroxidase 4: From genomic structure to developmental expression and neural function. Biol. Chem. 2007, 388, 1007–1017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puglisi, R.; Maccari, I.; Pipolo, S.; Conrad, M.; Mangia, F.; Boitani, C. The nuclear form of glutathione peroxidase 4 is associated with sperm nuclear matrix and is required for proper paternal chromatin decondensation at fertilization. J. Cell. Physiol. 2012, 227, 1420–1427. [Google Scholar] [CrossRef] [PubMed]
- Imai, H.; Hirao, F.; Sakamoto, T.; Sekine, K.; Mizukura, Y.; Saito, M.; Kitamoto, T.; Hayasaka, M.; Hanaoka, K.; Nakagawa, Y. Early embryonic lethality caused by targeted disruption of the mouse PHGPx gene. Biochem. Biophys. Res. Commun. 2003, 305, 278–286. [Google Scholar] [CrossRef]
- Yant, L.J.; Ran, Q.; Rao, L.; Van Remmen, H.; Shibatani, T.; Belter, J.G.; Motta, L.; Richardson, A.; Prolla, T.A. The selenoprotein GPX4 is essential for mouse development and protects from radiation and oxidative damage insults. Free Radic. Biol. Med. 2003, 34, 496–502. [Google Scholar] [CrossRef]
- Brutsch, S.H.; Wang, C.C.; Li, L.; Stender, H.; Neziroglu, N.; Richter, C.; Kuhn, H.; Borchert, A. Expression of inactive glutathione peroxidase 4 leads to embryonic lethality, and inactivation of the Alox15 gene does not rescue such knock-in mice. Antioxid. Redox Signal. 2015, 22, 281–293. [Google Scholar] [CrossRef]
- Ingold, I.; Aichler, M.; Yefremova, E.; Roveri, A.; Buday, K.; Doll, S.; Tasdemir, A.; Hoffard, N.; Wurst, W.; Walch, A.; et al. Expression of a Catalytically Inactive Mutant Form of Glutathione Peroxidase 4 (Gpx4) Confers a Dominant-negative Effect in Male Fertility. J. Biol. Chem. 2015, 290, 14668–14678. [Google Scholar] [CrossRef] [Green Version]
- Brutsch, S.H.; Rademacher, M.; Roth, S.R.; Muller, K.; Eder, S.; Viertel, D.; Franz, C.; Kuhn, H.; Borchert, A. Male Subfertility Induced by Heterozygous Expression of Catalytically Inactive Glutathione Peroxidase 4 Is Rescued in Vivo by Systemic Inactivation of the Alox15 Gene. J. Biol. Chem. 2016, 291, 23578–23588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borchert, A.; Wang, C.C.; Ufer, C.; Schiebel, H.; Savaskan, N.E.; Kuhn, H. The role of phospholipid hydroperoxide glutathione peroxidase isoforms in murine embryogenesis. J. Biol. Chem. 2006, 281, 19655–19664. [Google Scholar] [CrossRef] [Green Version]
- Yoo, S.E.; Chen, L.; Na, R.; Liu, Y.; Rios, C.; Van Remmen, H.; Richardson, A.; Ran, Q. Gpx4 ablation in adult mice results in a lethal phenotype accompanied by neuronal loss in brain. Free Radic. Biol. Med. 2012, 52, 1820–1827. [Google Scholar] [CrossRef] [Green Version]
- Canli, O.; Alankus, Y.B.; Grootjans, S.; Vegi, N.; Hultner, L.; Hoppe, P.S.; Schroeder, T.; Vandenabeele, P.; Bornkamm, G.W.; Greten, F.R. Glutathione peroxidase 4 prevents necroptosis in mouse erythroid precursors. Blood 2016, 127, 139–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujita, H.; Yamamoto, M.; Yamagami, T.; Hayashi, N.; Sassa, S. Erythroleukemia differentiation. Distinctive responses of the erythroid-specific and the nonspecific delta-aminolevulinate synthase mRNA. J. Biol. Chem. 1991, 266, 17494–17502. [Google Scholar] [CrossRef]
- Singer, D.; Cooper, M.; Maniatis, G.M.; Marks, P.A.; Rifkind, R.A. Erythropoietic differentiation in colonies of cells transformed by Friend virus. Proc. Natl. Acad. Sci. USA 1974, 71, 2668–2670. [Google Scholar] [CrossRef] [Green Version]
- Hafid-Medheb, K.; Augery-Bourget, Y.; Minatchy, M.N.; Hanania, N.; Robert-Lezenes, J. Bcl-XL is required for heme synthesis during the chemical induction of erythroid differentiation of murine erythroleukemia cells independently of its antiapoptotic function. Blood 2003, 101, 2575–2583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seguin, A.; Takahashi-Makise, N.; Yien, Y.Y.; Huston, N.C.; Whitman, J.C.; Musso, G.; Wallace, J.A.; Bradley, T.; Bergonia, H.A.; Kafina, M.D.; et al. Reductions in the mitochondrial ABC transporter Abcb10 affect the transcriptional profile of heme biosynthesis genes. J. Biol. Chem. 2017, 292, 16284–16299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsiftsoglou, A.S.; Wong, W. Molecular and cellular mechanisms of leukemic hemopoietic cell differentiation: An analysis of the Friend system. Anticancer Res. 1985, 5, 81–99. [Google Scholar]
- Friend, C.; Scher, W.; Holland, J.G.; Sato, T. Hemoglobin synthesis in murine virus-induced leukemic cells in vitro: Stimulation of erythroid differentiation by dimethyl sulfoxide. Proc. Natl. Acad. Sci. USA 1971, 68, 378–382. [Google Scholar] [CrossRef] [Green Version]
- Tsiftsoglou, A.S.; Vizirianakis, I.S.; Strouboulis, J. Erythropoiesis: Model systems, molecular regulators, and developmental programs. IUBMB Life 2009, 61, 800–830. [Google Scholar] [CrossRef]
- Altamura, S.; Vegi, N.M.; Hoppe, P.S.; Schroeder, T.; Aichler, M.; Walch, A.; Okreglicka, K.; Hultner, L.; Schneider, M.; Ladinig, C.; et al. Glutathione peroxidase 4 and vitamin E control reticulocyte maturation, stress erythropoiesis and iron homeostasis. Haematologica 2020, 105, 937–950. [Google Scholar] [CrossRef] [Green Version]
- Marks, P.A.; Sheffery, M.; Rifkind, R.A. Induction of transformed cells to terminal differentiation and the modulation of gene expression. Cancer Res. 1987, 47, 659–666. [Google Scholar]
- Borchert, A.; Kuttner, G.; Giessmann, E.; Wang, C.C.; Wessner, H.; Volkmer, R.; Hohne, W.; Kuhn, H. Defining the immunoreactive epitope for the monoclonal anti-human glutathione peroxidase-4 antibody anti-hGPx4 Mab63-1. Immunol. Lett. 2010, 133, 85–93. [Google Scholar] [CrossRef]
- Chan, R.Y.; Seiser, C.; Schulman, H.M.; Kuhn, L.C.; Ponka, P. Regulation of transferrin receptor mRNA expression. Distinct regulatory features in erythroid cells. Eur. J. Biochem. 1994, 220, 683–692. [Google Scholar] [CrossRef]
- Ouled-Haddou, H.; Messaoudi, K.; Demont, Y.; Lopes Dos Santos, R.; Carola, C.; Caulier, A.; Vong, P.; Jankovsky, N.; Lebon, D.; Willaume, A.; et al. A new role of glutathione peroxidase 4 during human erythroblast enucleation. Blood Adv. 2020, 4, 5666–5680. [Google Scholar] [CrossRef]
- Azad, G.K.; Tomar, R.S. Ebselen, a promising antioxidant drug: Mechanisms of action and targets of biological pathways. Mol. Biol. Rep. 2014, 41, 4865–4879. [Google Scholar] [CrossRef] [PubMed]
- Paulson, R.F.; Shi, L.; Wu, D.C. Stress erythropoiesis: New signals and new stress progenitor cells. Curr. Opin. Hematol. 2011, 18, 139–145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamane, T. Cellular Basis of Embryonic Hematopoiesis and Its Implications in Prenatal Erythropoiesis. Int. J. Mol. Sci. 2020, 21, 9346. [Google Scholar] [CrossRef] [PubMed]
- Morrison, S.J.; Hemmati, H.D.; Wandycz, A.M.; Weissman, I.L. The purification and characterization of fetal liver hematopoietic stem cells. Proc. Natl. Acad. Sci. USA 1995, 92, 10302–10306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- von Lindern, M.; Deiner, E.M.; Dolznig, H.; Parren-van Amelsvoort, M.; Hayman, M.J.; Mullner, E.W.; Beug, H. Leukemic transformation of normal murine erythroid progenitors: V- and c-ErbB act through signaling pathways activated by the EpoR and c-Kit in stress erythropoiesis. Oncogene 2001, 20, 3651–3664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dolznig, H.; Kolbus, A.; Leberbauer, C.; Schmidt, U.; Deiner, E.M.; Mullner, E.W.; Beug, H. Expansion and differentiation of immature mouse and human hematopoietic progenitors. Methods Mol. Med. 2005, 105, 323–344. [Google Scholar] [CrossRef]
- Matte, A.; de Franceschi, L. Oxidation and erythropoiesis. Curr. Opin. Hematol. 2019, 26, 145–151. [Google Scholar] [CrossRef]
- Petrak, J.; Myslivcova, D.; Man, P.; Cmejlova, J.; Cmejla, R.; Vyoral, D. Proteomic analysis of erythroid differentiation induced by hexamethylene bisacetamide in murine erythroleukemia cells. Exp. Hematol. 2007, 35, 193–202. [Google Scholar] [CrossRef] [PubMed]
- Savaskan, N.E.; Borchert, A.; Brauer, A.U.; Kuhn, H. Role for glutathione peroxidase-4 in brain development and neuronal apoptosis: Specific induction of enzyme expression in reactive astrocytes following brain injury. Free Radic. Biol. Med. 2007, 43, 191–201. [Google Scholar] [CrossRef]
- Bozo, I.Y.; Drobyshev, A.Y.; Redko, N.A.; Komlev, V.S.; Isaev, A.A.; Deev, R.V. Bringing a Gene-Activated Bone Substitute into Clinical Practice: From Bench to Bedside. Front. Bioeng. Biotechnol. 2021, 9, 599300. [Google Scholar] [CrossRef] [PubMed]
- Levinson, S.S.; Goldman, J. Measuring hemoglobin in plasma by reaction with tetramethylbenzidine. Clin. Chem. 1982, 28, 471–474. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Beug, H.; Palmieri, S.; Freudenstein, C.; Zentgraf, H.; Graf, T. Hormone-dependent terminal differentiation in vitro of chicken erythroleukemia cells transformed by ts mutants of avian erythroblastosis virus. Cell 1982, 28, 907–919. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Amplicon | Forward/Reverse | Gene ID |
|---|---|---|
| Gapdh | 5′-CCATCACCATCTTCCAGGAGCGA-3′ 5′-GGATGACCTTGCCCACAGCCTTG-3′ | 14433 |
| mGpx4 | 5′-GAGATGAGCTGGGGCCGTCTGA-3′ 5′-ACGCAGCCGTTCTTATCAATGAGAA-3′ | 625249 |
| m + cGpx4 | 5′-CGCCTGGTCTGGCAGGCACCA-3′ 5′-ACGCAGCCGTTCTTATCAATGAGAA-3′ | 625249 |
| nGpx4 | 5′-AGTTCCTGGGCTTGTGTGCATCC-3′ 5′-ACGCAGCCGTTCTTATCAATGAGAA-3′ | 625249 |
| Epo | 5′-CACCCTGCTGCTTTTACTCTCCTT-3′ 5′-CTTCTGCACAACCCATCGTGACAT-3′ | 13856 |
| Hba-a1 | 5′-ATGGTGCTCTCTGGGGAAGACAAA-3′ 5′-TCATCGAGGTGGCCTGCAGCATT-3′ | 15122 |
| Alas2 | 5′-ATGGTGGCAGCAGCTATGTTGCTA-3′ 5′-CTTGAACTTCTGGAGCTGCCCTC-3′ | 11656 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rademacher, M.; Kuhn, H.; Borchert, A. Expression Silencing of Glutathione Peroxidase 4 in Mouse Erythroleukemia Cells Delays In Vitro Erythropoiesis. Int. J. Mol. Sci. 2021, 22, 7795. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22157795
Rademacher M, Kuhn H, Borchert A. Expression Silencing of Glutathione Peroxidase 4 in Mouse Erythroleukemia Cells Delays In Vitro Erythropoiesis. International Journal of Molecular Sciences. 2021; 22(15):7795. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22157795
Chicago/Turabian StyleRademacher, Marlena, Hartmut Kuhn, and Astrid Borchert. 2021. "Expression Silencing of Glutathione Peroxidase 4 in Mouse Erythroleukemia Cells Delays In Vitro Erythropoiesis" International Journal of Molecular Sciences 22, no. 15: 7795. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22157795