
Uptake of Nanotitania by Gingival Epithelial Cells Promotes Inflammatory Response and Is Accelerated by Porphyromonas gingivalis Lipopolysaccharide: An In Vitro Study

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
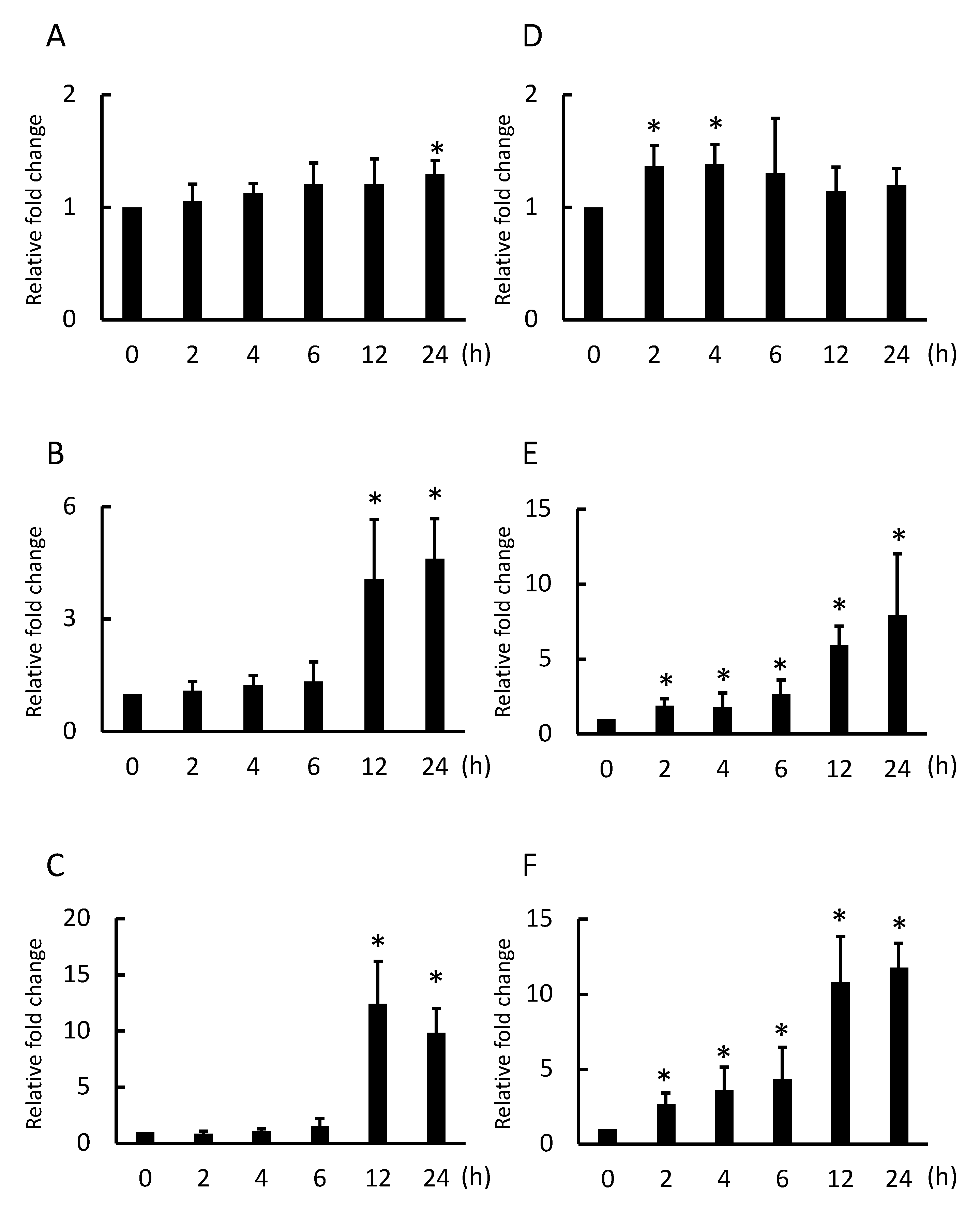
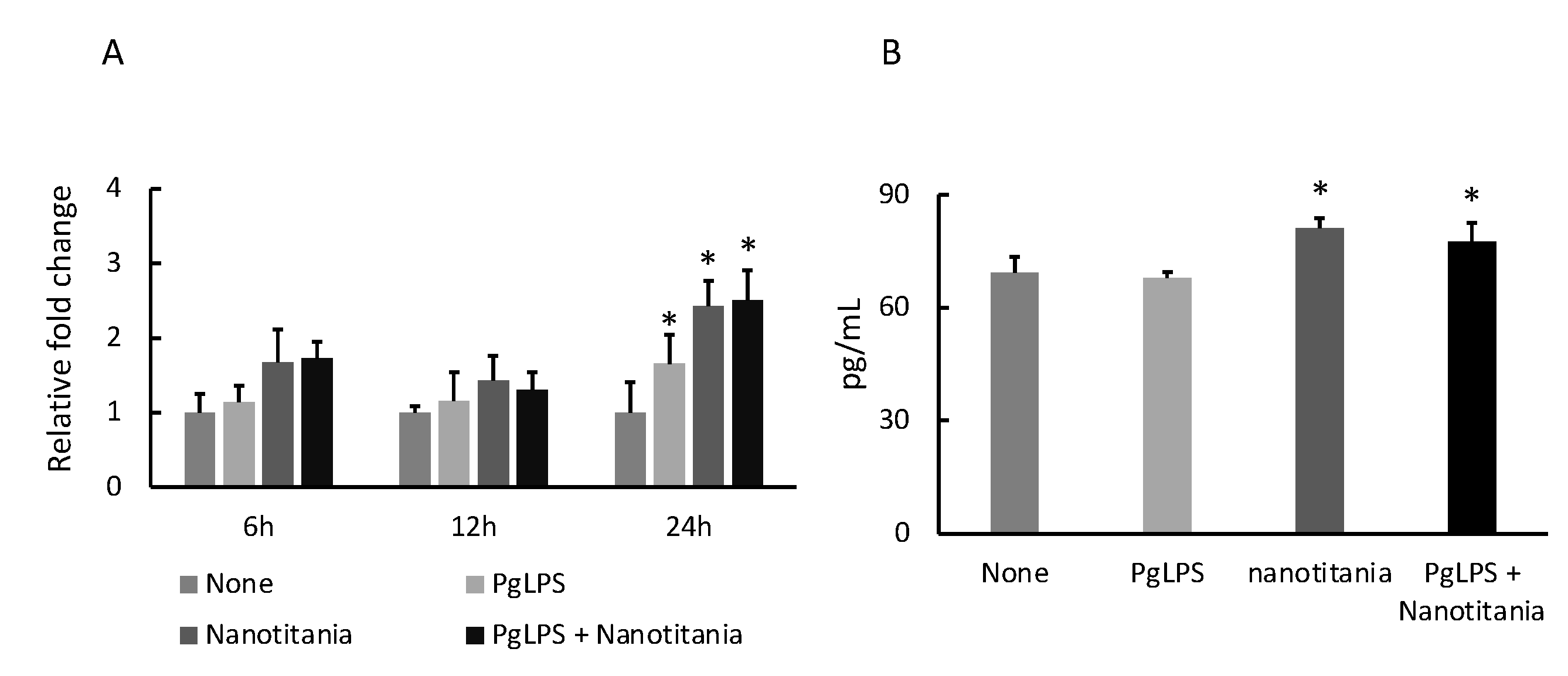
2.1. Nanotitania Upregulates the Expression of Inflammatory Cytokines
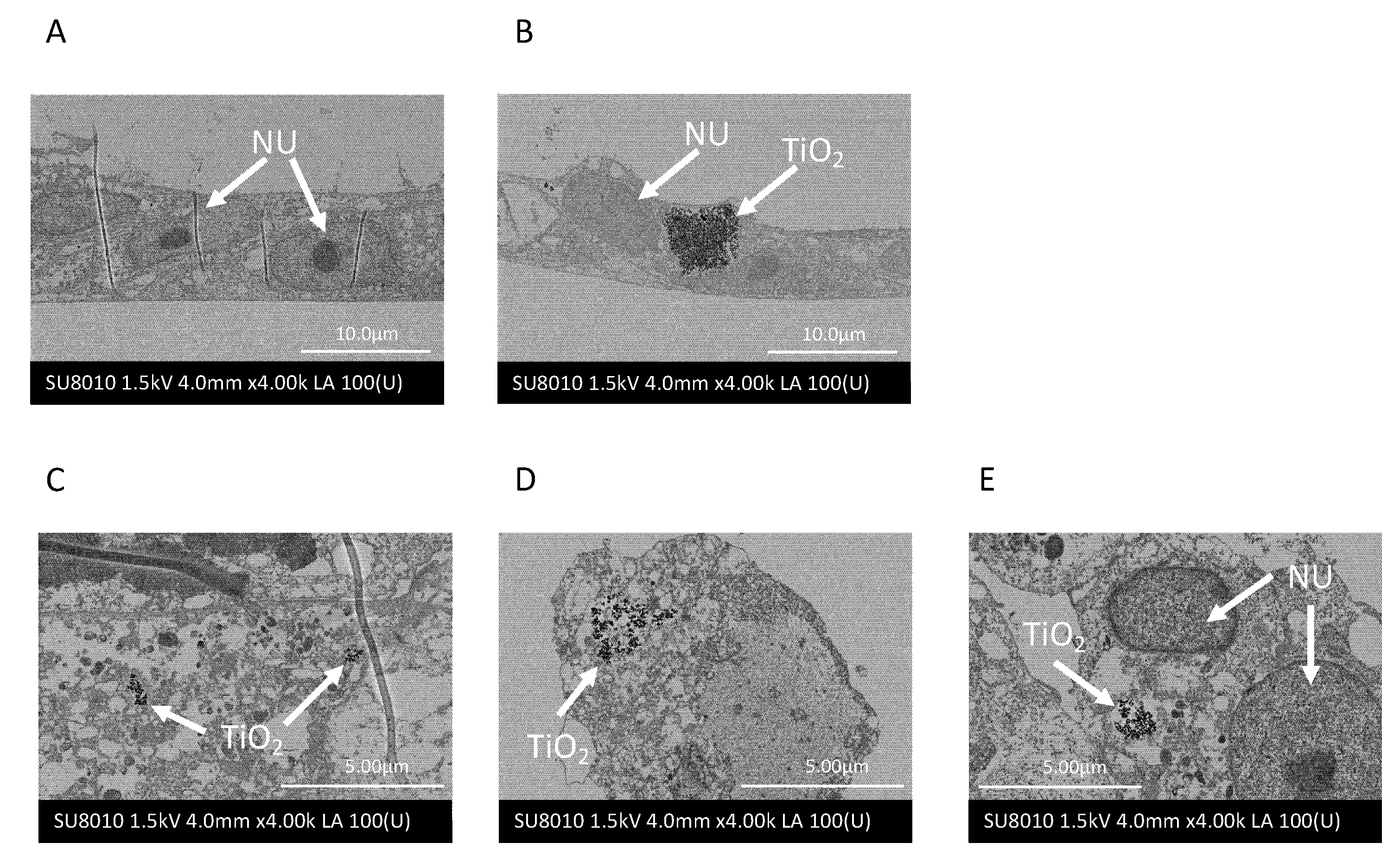
2.2. Nanotitania Uptake by Gingival Epithelial Cells
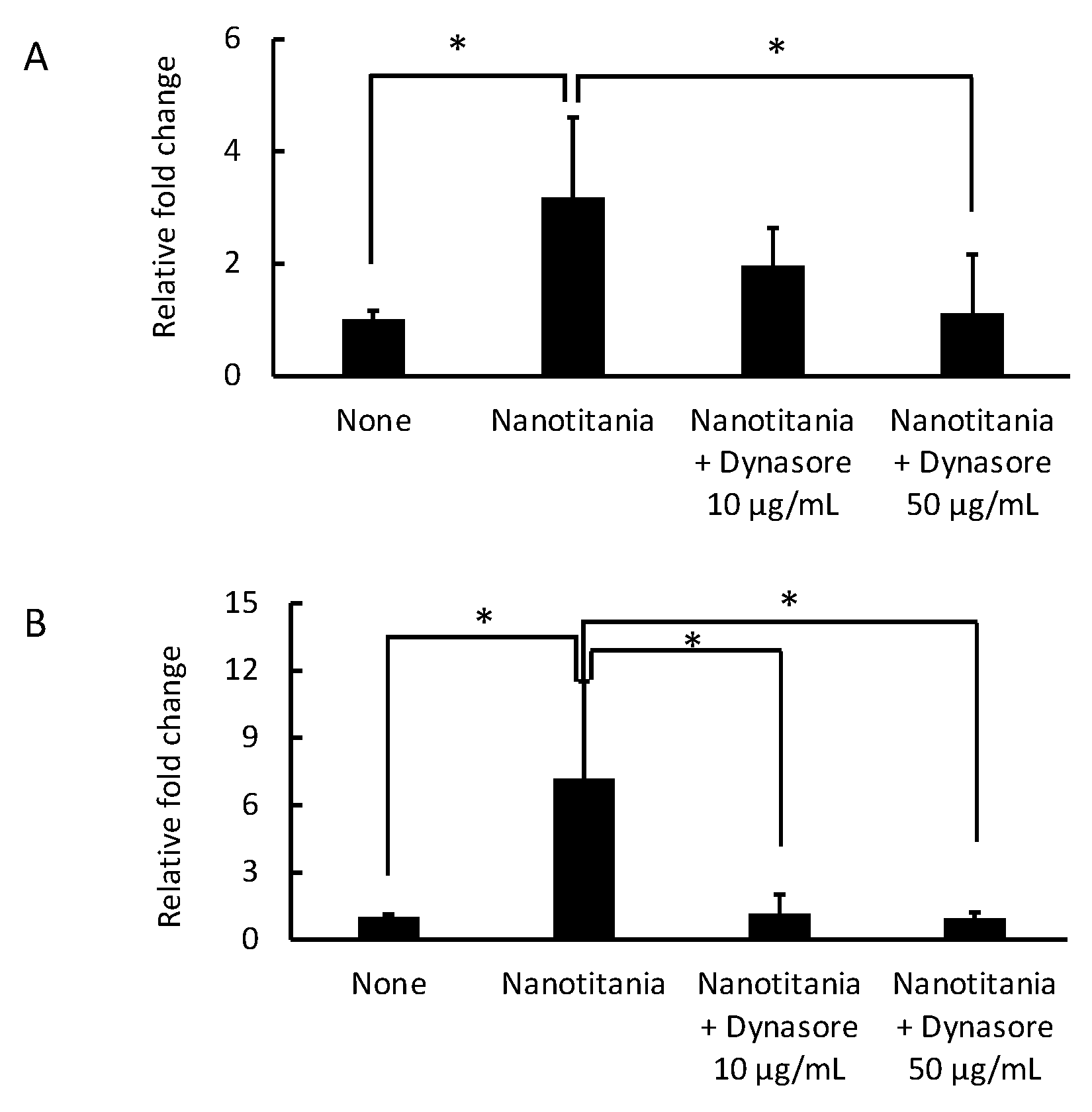
2.3. Endocytosis Inhibitor Blocks the Induction of Cytokine Production by the Uptake of Nanotitania in the Gingival Epithelium
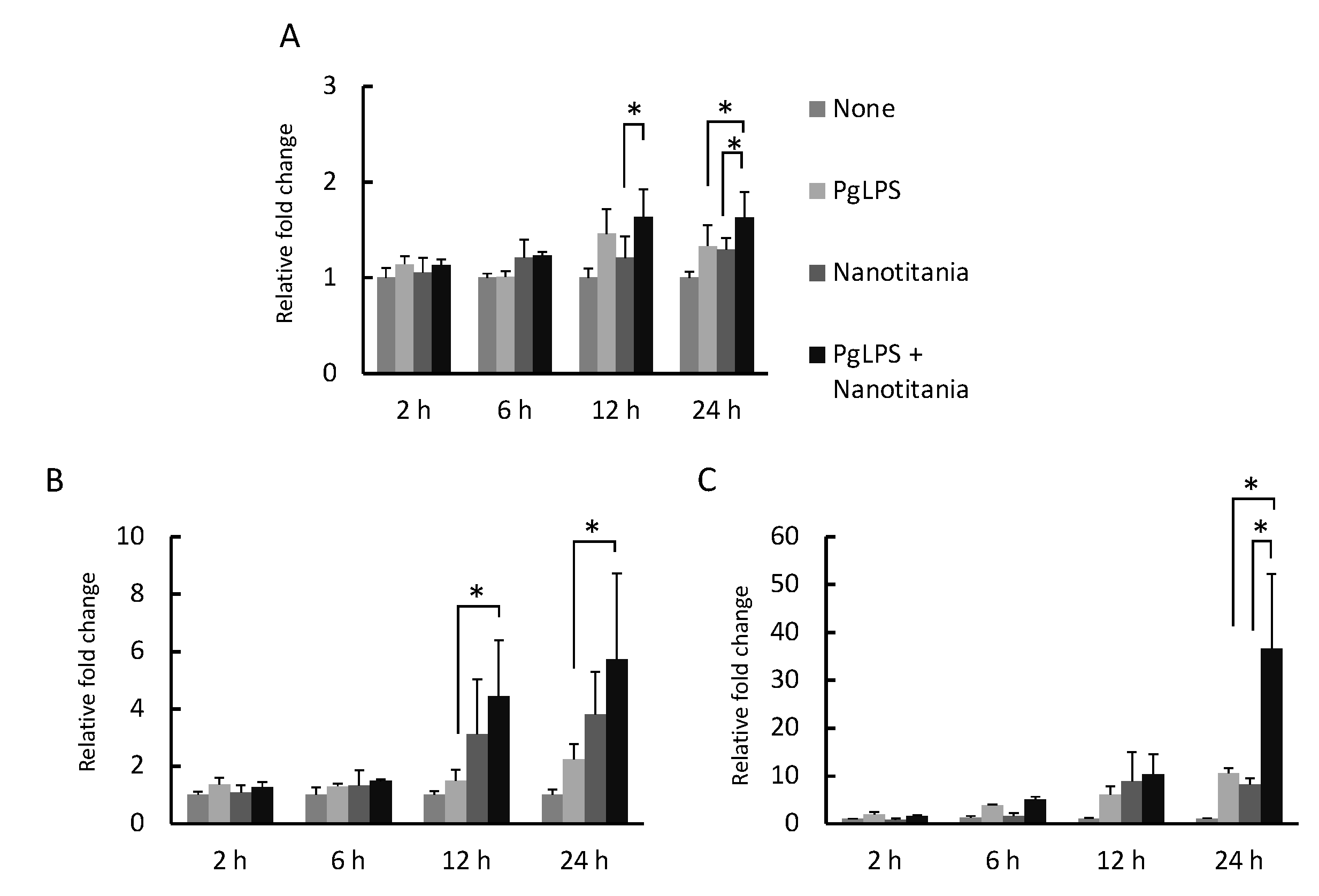
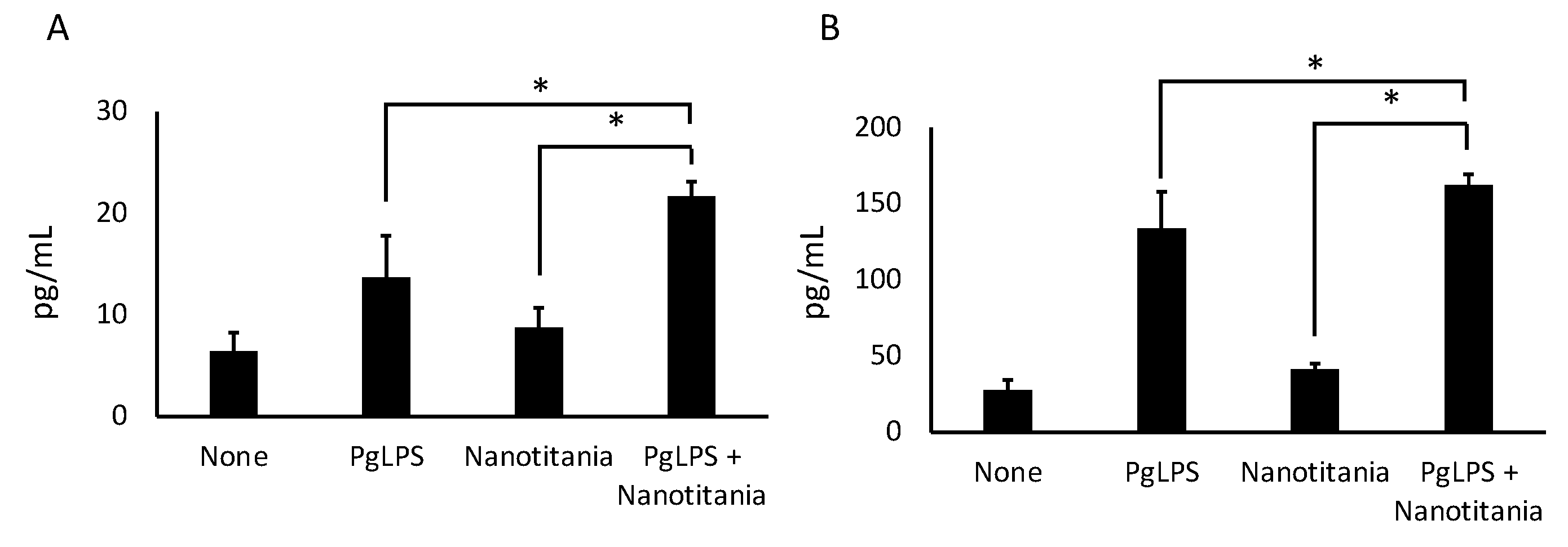
2.4. Co-Culture with Nanotitania and PgLPS Upregulates the Expression of Inflammatory Cytokines
2.5. Nanotitania Upregulates the Expression of IL-11
3. Discussion
4. Materials and Methods
4.1. Cell Culture and Reagents
4.2. Sample Preparation for Reverse Transcription-Quantitative Polymerase Chain Reaction
4.3. Reverse Transcription-Quantitative Polymerase Chain Reaction
4.4. Scanning Electron Microscope Observation
4.5. Enzyme-Linked Immunosorbent Assay
4.6. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Bornstein, M.M.; Horner, K.; Jacobs, R. Use of cone beam computed tomography in implant dentistry: Current concepts, indications and limitations for clinical practice and research. Periodontol. 2000 2017, 73, 51–72. [Google Scholar] [CrossRef]
- Romanos, G.E.; Delgado-Ruiz, R.; Sculean, A. Concepts for prevention of complications in implant therapy. Periodontol. 2000 2019, 81, 7–17. [Google Scholar] [CrossRef]
- Psillas, G.; Papaioannou, D.; Petsali, S.; Dimas, G.G.; Constantinidis, J. Odontogenic maxillary sinusitis: A comprehensive review. J. Dent. Sci. 2021, 16, 474–481. [Google Scholar] [CrossRef] [PubMed]
- Lindhe, J.; Meyle, J. Group D of European Workshop on Periodontology. Peri-implant diseases: Consensus Report of the Sixth European Workshop on Periodontology. J. Clin. Periodontol. 2008, 35, 282–285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tallarico, M.; Canullo, L.; Wang, H.-L.; Cochran, D.; Meloni, S. Classification System for Peri-implantitis: A Narrative Review with a Proposal of a New Evidence-Based Etiology Codification. Int J. Oral Maxillofac. Implants 2018, 33, 871–879. [Google Scholar] [CrossRef] [PubMed]
- Araujo, M.G.; Lindhe, J. Peri-implant health. J. Periodontol. 2018, 89, S249–S256. [Google Scholar] [CrossRef] [Green Version]
- Fransson, C.; Tomasi, C.; Pikner, S.S.; Gröndahl, K.; Wennström, J.L.; Leyland, A.H.; Berglundh, T. Severity and pattern of peri-implantitis-associated bone loss. J. Clin. Periodontol. 2010, 37, 442–448. [Google Scholar] [CrossRef]
- Fu, J.H.; Wang, H.L. Breaking the wave of peri-implantitis. Periodontol. 2000 2020, 84, 145–160. [Google Scholar] [CrossRef]
- López, C.Y.T.; Bueno, J.D.; Torres, I.Z.; López, M.L.; Macuas, A.H.; Álvarez, J.E. TiO2 Nanotubes Transformation into 4 nm Anatase Nanoparticles: Anodizing Industrial Recycled Titanium for Photocatalytic Water Remediation. Int. J. App. Nanotechnol. Res. 2019, 4, 26–44. [Google Scholar] [CrossRef]
- Lan, W.-C.; Huang, T.-S.; Cho, Y.-C.; Huang, Y.-T.; Walinski, C.J.; Chiang, P.-C.; Rusilin, M.; Pai, F.-T.; Huang, C.-C.; Huang, M.-S. The Potential of a Nanostructured Titanium Oxide Layer with Self-Assembled Monolayers for Biomedical Applications: Surface Properties and Biomechanical Behaviors. Appl. Sci. 2020, 10, 590. [Google Scholar] [CrossRef] [Green Version]
- White, P.C.; Chicca, I.J.; Cooper, P.R.; Milward, M.R.; Chapple, I.L. Neutrophil Extracellular Traps in Periodontitis: A Web of Intrigue. J. Dent. Res. 2016, 95, 26–34. [Google Scholar] [CrossRef] [PubMed]
- Wheelis, S.E.; Wilson, T.G., Jr.; Valderrama, P.; Rodrigues, D.C. Surface characterization of titanium implant healing abutments before and after placement. Clin. Implant. Dent. Relat. Res. 2018, 20, 180–190. [Google Scholar] [CrossRef] [PubMed]
- Noumbissi, S.; Scarano, A.; Gupta, S. A Literature Review Study on Atomic Ions Dissolution of Titanium and Its Alloys in Implant Dentistry. Materials 2019, 12, 368. [Google Scholar] [CrossRef] [Green Version]
- Noronha Oliveira, M.; Schunemann, W.V.H.; Mathew, M.T.; Henriques, B.; Magini, R.S.; Teughels, W.; Souza, J.C.M. Can degradation products released from dental implants affect peri-implant tissues? J. Periodontal. Res. 2018, 53, 1–11. [Google Scholar] [CrossRef]
- Bressan, E.; Ferroni, L.; Gardin, C.; Bellin, G.; Sbricoli, L.; Sivolella, S.; Brunello, G.; Schwartz-Arad, D.; Mijiritsky, E.; Penarrocha, M.; et al. Metal Nanoparticles Released from Dental Implant Surfaces: Potential Contribution to Chronic Inflammation and Peri-Implant Bone Loss. Materials 2019, 12, 2036. [Google Scholar] [CrossRef] [Green Version]
- Senna, P.; Antoninha Del Bel Cury, A.; Kates, S.; Meirelles, L. Surface Damage on Dental Implants with Release of Loose Particles after Insertion into Bone. Clin. Implant. Dent. Relat. Res. 2015, 17, 681–692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delgado-Ruiz, R.; Romanos, G. Potential Causes of Titanium Particle and Ion Release in Implant Dentistry: A Systematic Review. Int. J. Mol. Sci. 2018, 19, 3585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dodo, C.G.; Meirelles, L.; Aviles-Reyes, A.; Ruiz, K.G.S.; Abranches, J.; Cury, A.A.D.B. Pro-inflammatory analysis of macrophages in contact with titanium particles and Porphyromonas gingivalis. Braz. Dent. J. 2017, 28, 428–434. [Google Scholar] [CrossRef] [Green Version]
- Ishikawa, T.; Sugawara, S.; Kihara, H.; Hanasaka, T.; Hatakeyama, W.; Sasaki, M.; Kondo, H. Titanium nanoparticles potentially affect gingival tissue through IL-13α2 receptor expression. J. Oral Sci. 2021, 63, 263–266. [Google Scholar] [CrossRef]
- Darveau, R.P.; Pham, T.T.; Lemley, K.; Reife, R.A.; Bainbridge, B.W.; Coats, S.R.; Howald, W.N.; Way, S.S.; Hajjar, A.M. Porphyromonas gingivalis lipopolysaccharide contains multiple lipid A species that functionally interact with both toll-like receptors 2 and 4. Infect. Immun. 2004, 72, 5041–5051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kent, L.W.; Rahemtulla, F.; Michalek, S.M. Interleukin (IL)-1 and Porphyromonas gingivalis lipopolysaccharide stimulation of IL-6 production by fibroblasts derived from healthy or periodontally diseased human gingival tissue. J. Periodontol. 1999, 70, 274–282. [Google Scholar] [CrossRef] [PubMed]
- Giacaman, R.A.; Asrani, A.C.; Ross, K.F.; Herzberg, M.C. Cleavage of protease-activated receptors on an immortalized oral epithelial cell line by Porphyromonas gingivalis gingipains. Microbiology 2009, 155 Pt 10, 3238–3246. [Google Scholar] [CrossRef] [Green Version]
- Matsuda, T.; Hirano, T.; Nagasawa, S.; Kishimoto, T. Identification of alpha 2-macroglobulin as a carrier protein for IL-6. J. Immunol. 1989, 142, 148–152. [Google Scholar] [PubMed]
- Sauder, D.N.; Kilian, P.L.; McLane, J.A.; Quick, T.W.; Jakubovic, H.; Davis, S.C.; Eaglstein, W.H.; Mertz, P.M. Interleukin-1 enhances epidermal wound healing. Lymphokine Res. 1990, 9, 465–473. [Google Scholar]
- Kupper, T.S. The activated keratinocyte: A model for inducible cytokine production by non-bone marrow-derived cells in cutaneous inflammatory and immune responses. J. Investig. Dermatol. 1990, 94, 146S–150S. [Google Scholar] [CrossRef] [Green Version]
- Koch, A.E.; Polverini, P.J.; Kunkel, S.L.; Harlow, L.A.; DiPietro, L.A.; Elner, V.M.; Elner, S.G.; Strieter, R.M. Interleukin-8 as a macrophage-derived mediator of angiogenesis. Science 1992, 258, 1798–1801. [Google Scholar] [CrossRef]
- Maekawa, T.; Krauss, J.L.; Abe, T.; Jotwani, R.; Triantafilou, M.; Triantafilou, K.; Hashim, A.; Hoch, S.; Curtis, M.A.; Nussbaum, G.; et al. Porphyromonas gingivalis manipulates complement and TLR signaling to uncouple bacterial clearance from inflammation and promote dysbiosis. Cell Host Microbe 2014, 15, 768–778. [Google Scholar] [CrossRef] [Green Version]
- Leira, Y.; Iglesias-Rey, R.; Gómez-Lado, N.; Aguiar, P.; Campos, F.; D’Aiuto, F.; Castillo, J.; Blanco, J.; Sobrino, T. Porphyromonas gingivalis lipopolysaccharide-induced periodontitis and serum amyloid-beta peptides. Arch. Oral Biol. 2019, 99, 120–125. [Google Scholar] [CrossRef]
- Kirchhausen, T.; Macia, E.; Pelish, H.E. Use of dynasore, the small molecule inhibitor of dynamin, in the regulation of endocytosis. Methods Enzymol. 2008, 438, 77–93. [Google Scholar] [PubMed] [Green Version]
- Doherty, G.J.; McMahon, H.T. Mechanisms of endocytosis. Annu Rev. Biochem. 2009, 78, 857–902. [Google Scholar] [CrossRef] [Green Version]
- Michael Danielsen, E.; Hansen, G.H. Small molecule pinocytosis and clathrin-dependent endocytosis at the intestinal brush border: Two separate pathways into the enterocyte. Biochim. Biophys. Acta 2016, 1858, 233–243. [Google Scholar] [CrossRef]
- Garbers, C.; Scheller, J. Interleukin-6 and interleukin-11: Same same but different. Biol. Chem. 2013, 394, 1145–1161. [Google Scholar] [CrossRef] [PubMed]
- Lokau, J.; Nitz, R.; Agthe, M.; Monhasery, N.; Aparicio-Siegmund, S.; Schumacher, N.; Wolf, J.; Möller-Hackbarth, K.; Waetzig, G.H.; Grötzinger, J.; et al. Proteolytic Cleavage Governs Interleukin-11 Trans-signaling. Cell Rep. 2016, 14, 1761–1773. [Google Scholar] [CrossRef] [Green Version]
- Paul, S.R.; Bennett, F.; Calvetti, J.A.; Kelleher, K.; Wood, C.R.; O’Hara, R.M., Jr.; Leary, A.C.; Sibley, B.; Clark, S.C.; Williams, D.A. Molecular cloning of a cDNA encoding interleukin 11, a stromal cell-derived lymphopoietic and hematopoietic cytokine. Proc. Natl. Acad. Sci. USA 1990, 87, 7512–7516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andus, T.; Holstege, A. Cytokines and the liver in health and disease. Effects on liver metabolism and fibrogenesis. Acta Gastroenterol. Belg. 1994, 57, 236–244. [Google Scholar] [PubMed]
- Maheshwari, A.; Janssens, K.; Bogie, J.; Van Den Haute, C.; Struys, T.; Lambrichts, I.; Baekelandt, V.; Stinissen, P.; Hendriks, J.J.; Slaets, H.; et al. Local overexpression of interleukin-11 in the central nervous system limits demyelination and enhances remyelination. Mediators Inflamm. 2013, 685317. [Google Scholar] [CrossRef]
- Du, X.X.; Neben, T.; Goldman, S.; Williams, D.A. Effects of recombinant human interleukin-11 on hematopoietic reconstitution in transplant mice: Acceleration of recovery of peripheral blood neutrophils and platelets. Blood 1993, 81, 27–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amarasekara, D.S.; Yun, H.; Kim, S.; Lee, N.; Kim, H.; Rho, J. Regulation of Osteoclast Differentiation by Cytokine Networks. Immune Netw. 2018, 18, e8. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Fujita, N.; Oh-hara, T.; Morinaga, Y.; Nakagawa, T.; Yamada, M.; Tsuruo, T. Production of interleukin-11 in bone-derived endothelial cells and its role in the formation of osteolytic bone metastasis. Oncogene 1998, 16, 693–703. [Google Scholar] [CrossRef] [Green Version]
- McCoy, E.M.; Hong, H.; Pruitt, H.C.; Feng, X. IL-11 produced by breast cancer cells augments osteoclastogenesis by sustaining the pool of osteoclast progenitor cells. BMC Cancer 2013, 13, 16. [Google Scholar] [CrossRef] [Green Version]
- Liang, M.; Ma, Q.; Ding, N.; Luo, F.; Bai, Y.; Kang, F.; Gong, X.; Dong, R.; Dai, J.; Dai, Q.; et al. IL-11 is essential in promoting osteolysis in breast cancer bone metastasis via RANKL-independent activation of osteoclastogenesis. Cell Death Dis. 2019, 10, 353. [Google Scholar] [CrossRef] [Green Version]
- Putoczki, T.L.; Thiem, S.; Loving, A.; Busuttil, R.A.; Wilson, N.J.; Ziegler, P.K.; Nguyen, P.M.; Preaudet, A.; Farid, R.; Edwards, K.M.; et al. Interleukin-11 is the dominant IL-6 family cytokine during gastrointestinal tumorigenesis and can be targeted therapeutically. Cancer Cell 2013, 24, 257–271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ernst, M.; Putoczki, T.L. Molecular pathways: IL11 as a tumor-promoting cytokine-translational implications for cancers. Clin Cancer Res. 2014, 20, 5579–5588. [Google Scholar] [CrossRef] [Green Version]
- Kohal, R.J.; Dennison, D.K. Clinical Longevity of Zirconia Implants with the Focus on Biomechanical and Biological Outcome. Curr. Oral Health Rep. 2020, 7, 344–351. [Google Scholar] [CrossRef]
- Neale, S.D.; Haynes, D.R.; Howie, D.W.; Murray, D.W.; Athanasou, N.A. The effect of particle phagocytosis and metallic wear particles on osteoclast formation and bone resorption in vitro. J. Arthroplasty 2000, 15, 654–662. [Google Scholar] [CrossRef] [PubMed]
- Angireddy, R.; Kazmi, H.R.; Srinivasan, S.; Sun, L.; Iqbal, J.; Fuchs, S.Y.; Guha, M.; Kijima, T.; Yuen, T.; Zaidi, M.; et al. Cytochrome c oxidase dysfunction enhances phagocytic function and osteoclast formation in macrophages. FASEB J. 2019, 33, 9167–9181. [Google Scholar] [CrossRef] [PubMed]
- Takito, J.; Nakamura, M. Heterogeneity and Actin Cytoskeleton in Osteoclast and Macrophage Multinucleation. Int. J. Mol. Sci. 2020, 21, 6629. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, T.; Terashima, J.; Shimoyama, Y.; Ohashi, Y.; Mikami, T.; Takeda, Y.; Sasaki, M. Effects of butyric acid, a bacterial metabolite, on the migration of ameloblastoma mediated by laminin 332. J. Oral Sci. 2020, 62, 435–438. [Google Scholar] [CrossRef]
- Shin, M.; Nozaki, T.; Idrizi, F.; Isogai, S.; Ogasawara, K.; Ishida, K.; Yuge, S.; Roscoe, B.; Wolfe, S.A.; Fukuhara, S.; et al. Valves Are a Conserved Feature of the Zebrafish Lymphatic System. Dev. Cell 2019, 51, 374–386.e5. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sugawara, S.; Ishikawa, T.; Sato, S.; Kihara, H.; Taira, M.; Sasaki, M.; Kondo, H. Uptake of Nanotitania by Gingival Epithelial Cells Promotes Inflammatory Response and Is Accelerated by Porphyromonas gingivalis Lipopolysaccharide: An In Vitro Study. Int. J. Mol. Sci. 2021, 22, 8084. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22158084
Sugawara S, Ishikawa T, Sato S, Kihara H, Taira M, Sasaki M, Kondo H. Uptake of Nanotitania by Gingival Epithelial Cells Promotes Inflammatory Response and Is Accelerated by Porphyromonas gingivalis Lipopolysaccharide: An In Vitro Study. International Journal of Molecular Sciences. 2021; 22(15):8084. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22158084
Chicago/Turabian StyleSugawara, Shiho, Taichi Ishikawa, Shu Sato, Hidemichi Kihara, Masayuki Taira, Minoru Sasaki, and Hisatomo Kondo. 2021. "Uptake of Nanotitania by Gingival Epithelial Cells Promotes Inflammatory Response and Is Accelerated by Porphyromonas gingivalis Lipopolysaccharide: An In Vitro Study" International Journal of Molecular Sciences 22, no. 15: 8084. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22158084