
Evaluation of Anti-Melanogenesis Activity of Enriched Pueraria lobata Stem Extracts and Characterization of Its Phytochemical Components Using HPLC–PDA–ESI–MS/MS

 ,
,  , , and
, , and
Abstract
:1. Introduction
2. Results
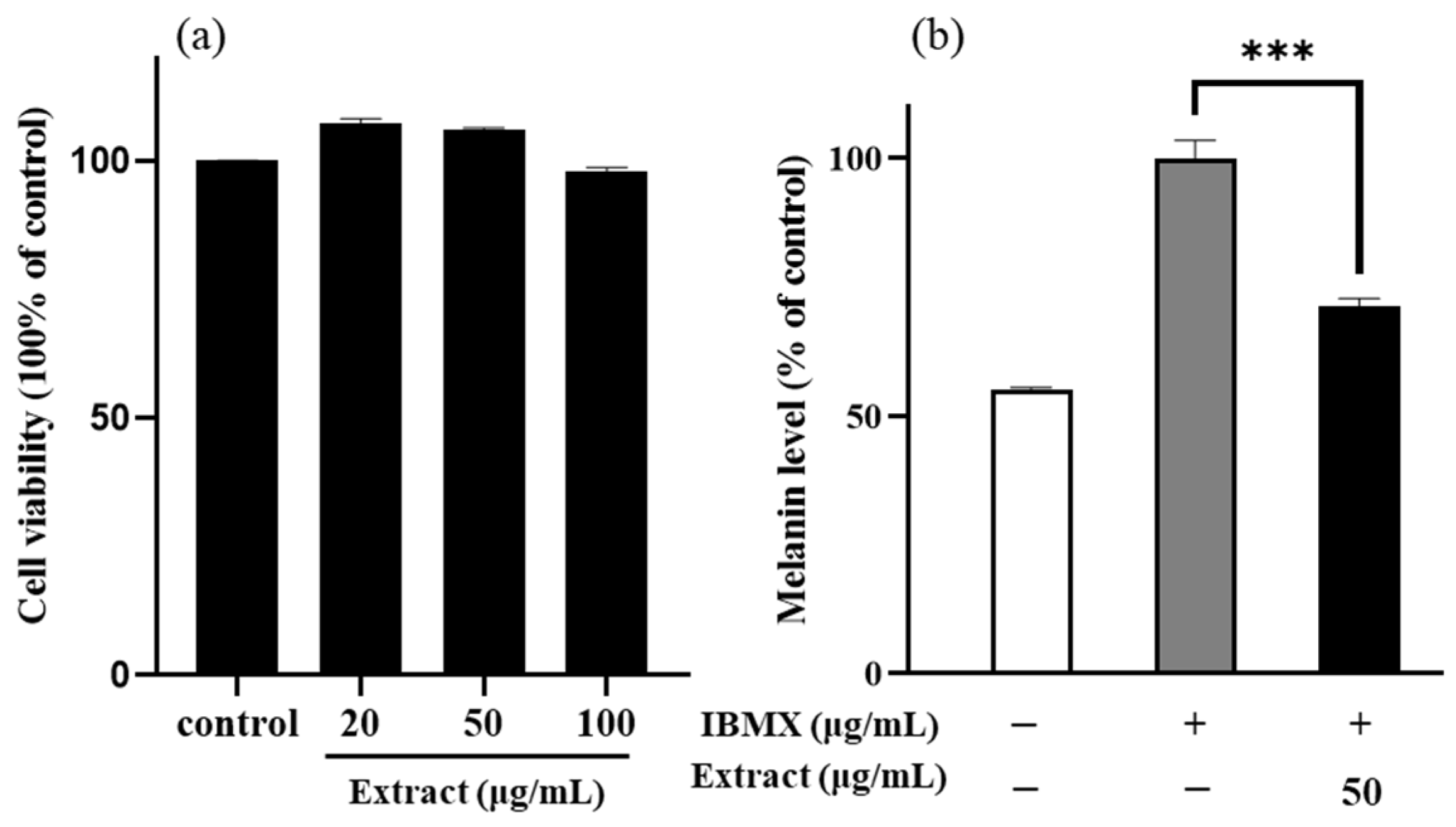
2.1. Effects of the Enriched PLS Extracts on the Viability and Melanin Content of the B16/F10 Melanoma Cells
2.2. Visual Observation of Melanin Pigmentations
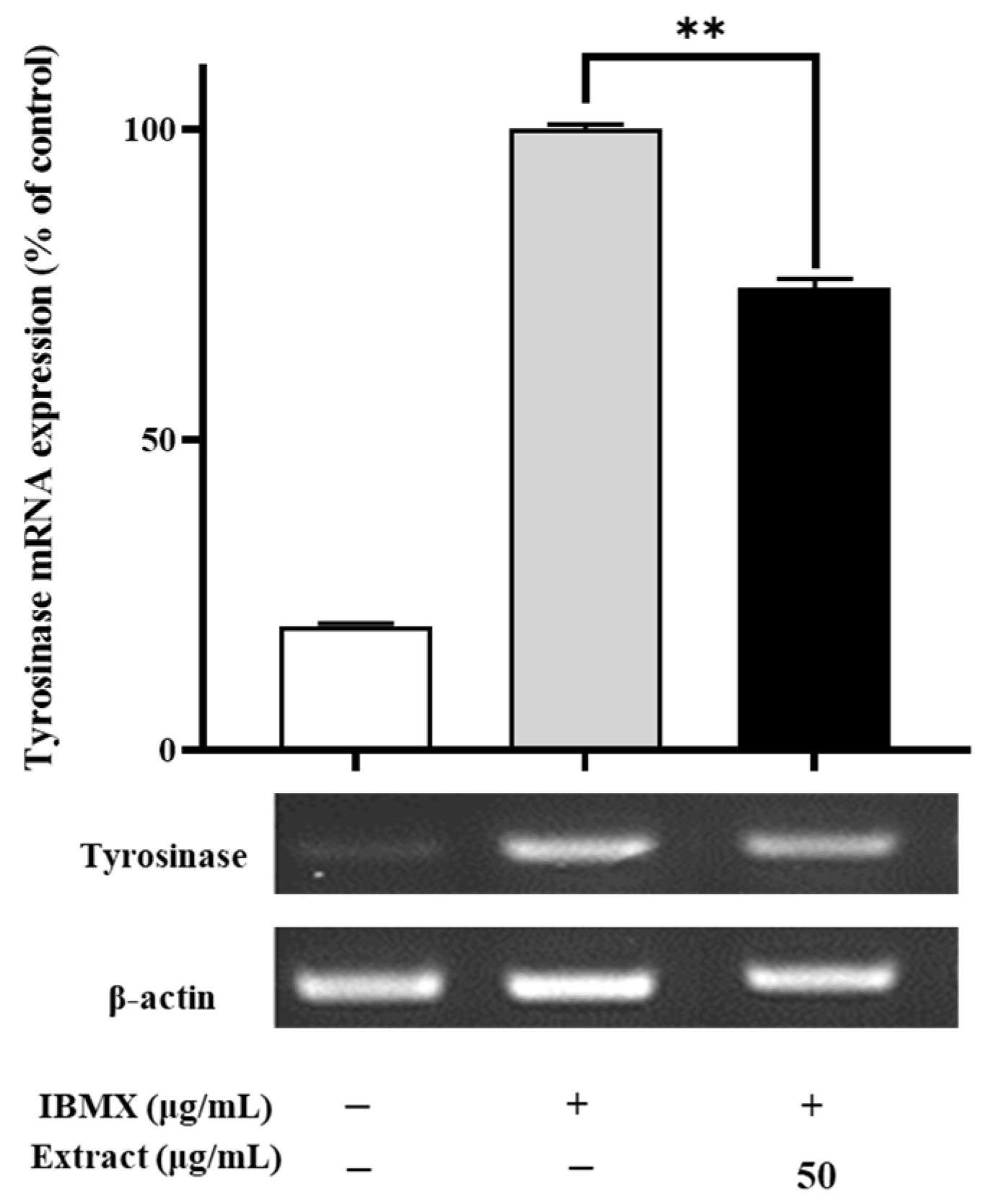
2.3. Effects of the Enriched PLS Extracts on the Tyrosinase Expression Levels
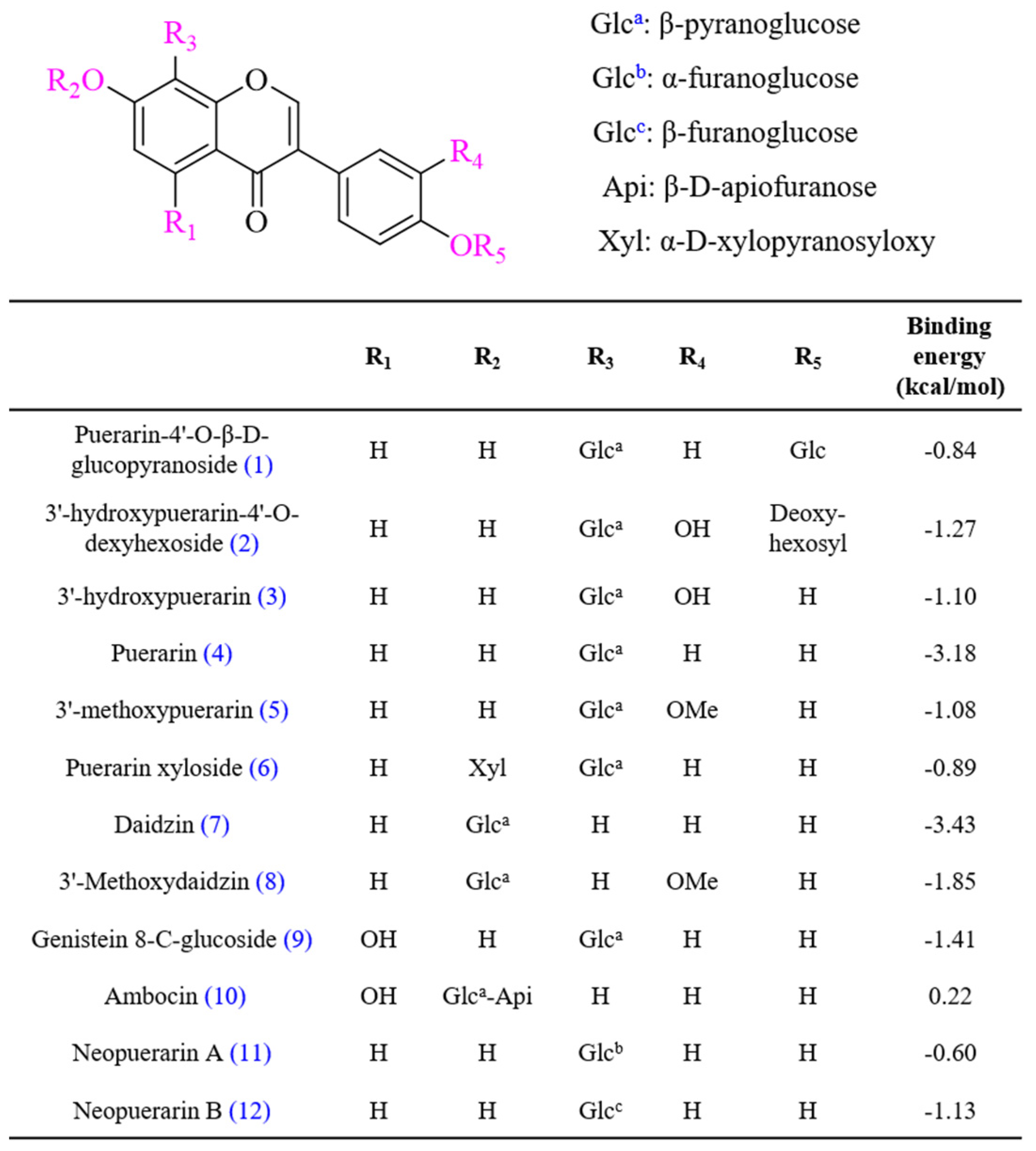
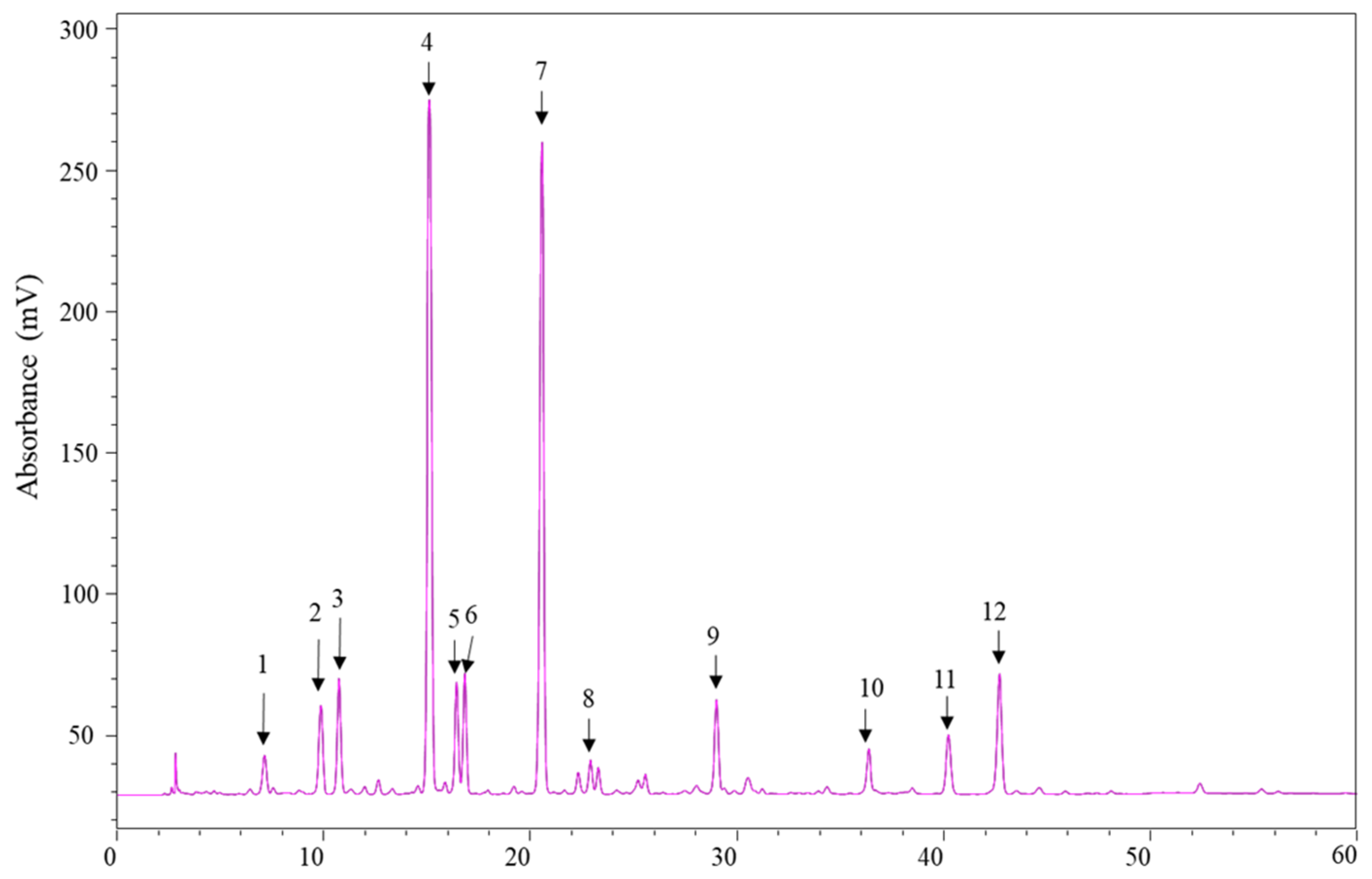
2.4. High-Performance Liquid Chromatography–Photodiode Array–Electrospray Ionization–Mass Spectrometry (HPLC–PDA–ESI–MS/MS) Analysis
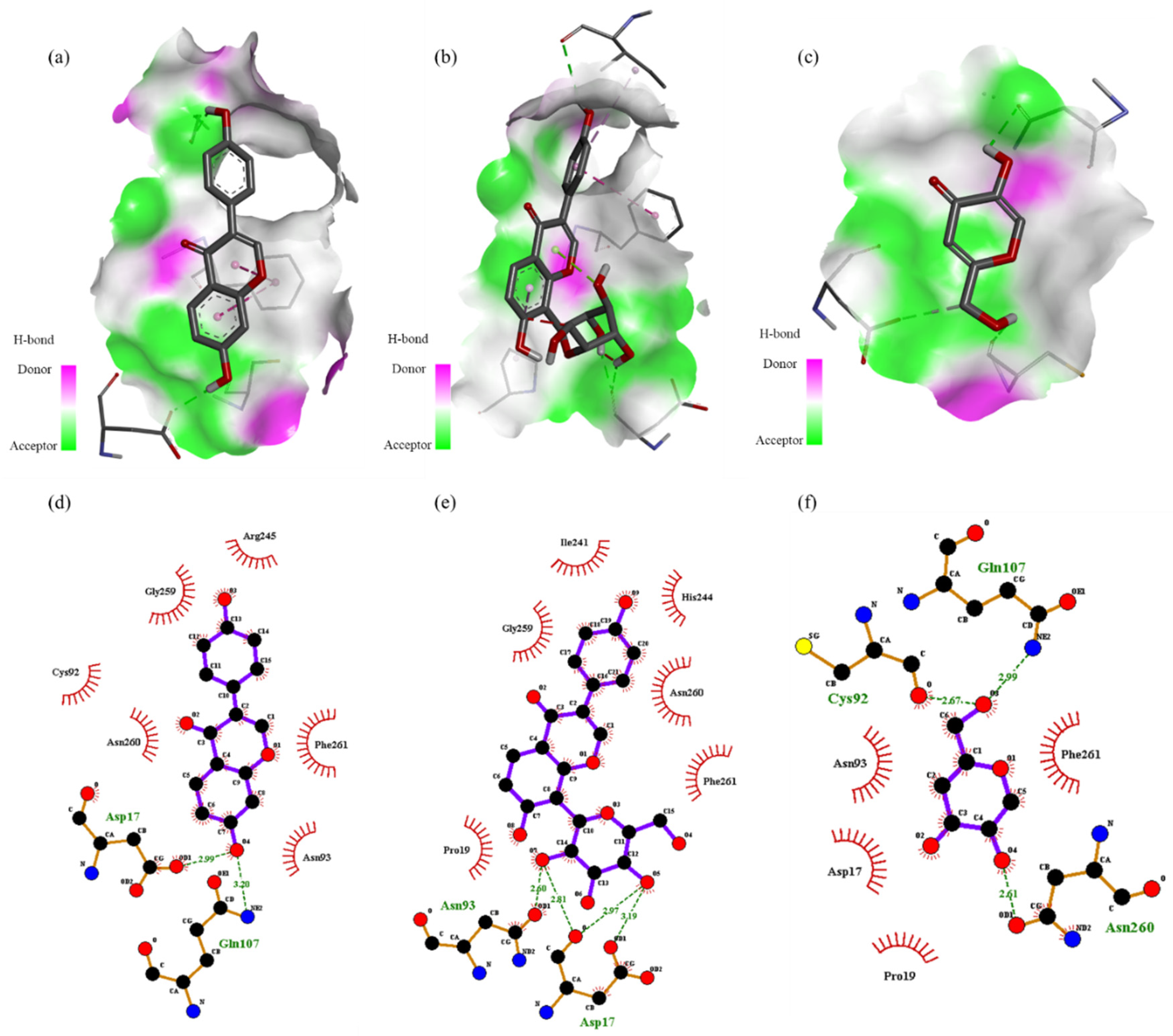
2.5. In Silico Docking
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Reagents
4.2. Preparation of Enriched PLS Extracts
4.3. Cell Culture
4.4. Cell Viability Assay
4.5. Determination of the Cellular Melanin Contents
4.6. Visual Evaluation of Melanin Pigmentation
4.7. RT-PCR Analysis
4.8. HPLC–PDA–ESI–MS/MS Analysis of PLS Extracts
4.9. Molecular Docking Simulation
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
Abbreviations
| PLS | Pueraria lobata stem |
| cAMP | cyclic adenosine monophosphate |
| IBMX | 3-isobutyl-1-methylxanthine |
| MITF | microphthalmia-associated transcription factor |
| RT-PCR | real-time quantitative polymerase chain reaction |
| HPLC–PDA–ESI–MS/MS | high-performance liquid chromatography–photodiode array–electrospray ionization–mass spectrometry |
| PKA | protein kinase A |
| DMEM | Dulbecco’s modified eagle’s medium |
| FBS | fetal bovine serum |
| ANOVA | analysis of variance |
| PBS | phosphate buffer saline |
| IC50 | Inhibitory concentration 50 |
References
- Hurbain, I.; Romao, M.; Sextius, P.; Bourreau, E.; Marchal, C.; Bernerd, F.; Duval, C.; Raposo, G. Melanosome distribution in keratinocytes in different skin types: Melanosome clusters are not degradative organelles. J. Investig. Dermatol. 2018, 138, 647–656. [Google Scholar] [CrossRef] [Green Version]
- Han, H.J.; Park, S.K.; Kang, J.Y.; Kim, J.M.; Yoo, S.K.; Heo, H.J. Anti-melanogenic effect of ethanolic extract of Sorghum bicolor on IBMX–induced melanogenesis in B16/F10 melanoma cells. Nutrients 2020, 12, 832. [Google Scholar] [CrossRef] [Green Version]
- Hwang, J.A.; Park, N.H.; Na, Y.J.; Lee, H.K.; Lee, J.H.; Kim, Y.J.; Lee, C.S. Coumestrol down-regulates melanin production in melan-a murine melanocytes through degradation of tyrosinase. Biol. Pharm. Bull. 2017, 40, 535–539. [Google Scholar] [CrossRef] [Green Version]
- Baek, S.-H.; Ahn, J.-W.; Nam, S.-H.; Yoon, C.-S.; Shin, J.-C.; Lee, S.-H. S-(−)-10, 11-dihydroxyfarnesoic acid methyl ester inhibits melanin synthesis in murine melanocyte cells. Int. J. Mol. Sci. 2014, 15, 12750–12763. [Google Scholar] [CrossRef]
- D’Mello, S.A.; Finlay, G.J.; Baguley, B.C.; Askarian-Amiri, M.E. Signaling pathways in melanogenesis. Int. J. Mol. Sci. 2016, 17, 1144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jang, E.J.; Shin, Y.; Park, H.J.; Kim, D.; Jung, C.; Hong, J.-Y.; Kim, S.; Lee, S.K. Anti-melanogenic activity of phytosphingosine via the modulation of the microphthalmia-associated transcription factor signaling pathway. J. Dermatol. Sci. 2017, 87, 19–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gillbro, J.; Olsson, M. The melanogenesis and mechanisms of skin-lightening agents-existing and new approaches. Int. J. Cosmet. Sci. 2011, 33, 210–221. [Google Scholar] [CrossRef]
- Kim, Y.-M.; Cho, S.-E.; Seo, Y.-K. The activation of melanogenesis by p-CREB and MITF signaling with extremely low-frequency electromagnetic fields on B16F10 melanoma. Life Sci. 2016, 162, 25–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sima, V.H.; Patris, S.; Aydogmus, Z.; Sarakbi, A.; Sandulescu, R.; Kauffmann, J.-M. Tyrosinase immobilized magnetic nanobeads for the amperometric assay of enzyme inhibitors: Application to the skin whitening agents. Talanta 2011, 83, 980–987. [Google Scholar] [CrossRef]
- Panichakul, T.; Rodboon, T.; Suwannalert, P.; Tripetch, C.; Rungruang, R.; Boohuad, N.; Youdee, P. Additive Effect of a Combination of Artocarpus lakoocha and Glycyrrhiza glabra Extracts on Tyrosinase Inhibition in Melanoma B16 Cells. Pharmaceuticals 2020, 13, 310. [Google Scholar] [CrossRef]
- Saeedi, M.; Eslamifar, M.; Khezri, K. Kojic acid applications in cosmetic and pharmaceutical preparations. Biomed. Pharmacother. 2019, 110, 582–593. [Google Scholar] [CrossRef] [PubMed]
- Nordlund, J.; Grimes, P.; Ortonne, J.P. The safety of hydroquinone. J. Eur. Acad. Dermatol. Venereol. 2006, 20, 781–787. [Google Scholar] [CrossRef]
- Burger, P.; Landreau, A.; Azoulay, S.; Michel, T.; Fernandez, X. Skin whitening cosmetics: Feedback and challenges in the development of natural skin lighteners. Cosmetics 2016, 3, 36. [Google Scholar] [CrossRef] [Green Version]
- Choi, Y.; Bose, S.; Shin, N.R.; Song, E.-J.; Nam, Y.-D.; Kim, H. Lactate-fortified Puerariae Radix fermented by Bifidobacterium breve improved diet-induced metabolic dysregulation via alteration of gut microbial communities. Nutrients 2020, 12, 276. [Google Scholar] [CrossRef] [Green Version]
- Jin, S.E.; Son, Y.K.; Min, B.-S.; Jung, H.A.; Choi, J.S. Anti-inflammatory and antioxidant activities of constituents isolated from Pueraria lobata roots. Arch. Pharm. Res. 2012, 35, 823–837. [Google Scholar] [CrossRef] [PubMed]
- Wagle, A.; Seong, S.H.; Jung, H.A.; Choi, J.S. Identifying an isoflavone from the root of Pueraria lobata as a potent tyrosinase inhibitor. Food Chem. 2019, 276, 383–389. [Google Scholar] [CrossRef]
- Mocan, A.; Carradori, S.; Locatelli, M.; Secci, D.; Cesa, S.; Mollica, A.; Riga, S.; Angeli, A.; Supuran, C.T.; Celia, C. Bioactive isoflavones from Pueraria lobata root and starch: Different extraction techniques and carbonic anhydrase inhibition. Food Chem. Toxicol. 2018, 112, 441–447. [Google Scholar] [CrossRef]
- Kang, Y.; Choi, J.-U.; Lee, E.-A.; Park, H.-R. Flaniostatin, a new isoflavonoid glycoside isolated from the leaves of Cudrania tricuspidata as a tyrosinase inhibitor. Food Sci. Biotechnol. 2013, 22, 1–4. [Google Scholar] [CrossRef]
- Kim, J.H.; Lee, E.S.; Lee, C.H. Melanin biosynthesis inhibitory effects of calycosin-7-O-β-d-glucoside isolated from astragalus (Astragalus membranaceus). Food Sci. Biotechnol. 2011, 20, 1481–1485. [Google Scholar] [CrossRef]
- Kim, D.E.; Hwang, Y.S.; Chang, B.Y.; Han, J.H.; Kim, D.S.; Kim, H.S.; Cho, H.K.; Kim, S.Y. Whitening effect and mechanism of aerial part of Pueraria lobata. Korean J. Pharmacogn. 2016, 47, 343–351. [Google Scholar]
- Mun, S.-C.; Mun, G.-S. Dynamics of phytoestrogen, isoflavonoids, and its isolation from stems of Pueraria lobata (Willd.) Ohwi growing in Democratic People’s Republic of Korea. J. Food Drug Anal. 2015, 23, 538–544. [Google Scholar] [CrossRef] [Green Version]
- Morgan, A.; Jeon, M.N.; Jeong, M.H.; Yang, S.Y.; Kim, Y.H. Chemical components from the stems of Pueraria lobata and their tyrosinase inhibitory activity. Nat. Prod. Sci. 2016, 22, 111–116. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.-J.; Yang, X.-P.; Wang, K.-W. Isolation of two new C-glucofuranosyl isoflavones from Pueraria lobata (Wild.) Ohwi with HPLC–MS guiding analysis. J. Asian Nat. Prod. Res. 2010, 12, 293–299. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Wang, X.; Fan, E. Optimisation of ultrasound-assisted extraction of puerarin and total isoflavones from Puerariae Lobatae Radix (Pueraria lobata (Wild.) Ohwi) with response surface methodology. Phytochem. Anal. 2012, 23, 513–519. [Google Scholar] [CrossRef]
- Zhao, C.; Chan, H.y.; Yuan, D.; Liang, Y.; Lau, T.y.; Chau, F.T. Rapid simultaneous determination of major isoflavones of Pueraria lobata and discriminative analysis of its geographical origins by principal component analysis. Phytochem. Anal. 2011, 22, 503–508. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Zhu, Y.; Wang, T.; Qi, J.; Liu, X. Enzyme-site blocking combined with optimization of molecular docking for efficient discovery of potential tyrosinase specific inhibitors from Puerariae lobatae radix. Molecules 2018, 23, 2612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Ying, J.; Jiang, D.; Chang, Z.; Li, H.; Zhang, G.; Gong, S.; Jiang, X.; Tao, J. Urotensin-II receptor stimulation of cardiac L-type Ca2+ channels requires the βγ subunits of Gi/o-protein and phosphatidylinositol 3-kinase-dependent protein kinase C β1 isoform. J. Biol. Chem. 2015, 290, 8644–8655. [Google Scholar] [CrossRef] [Green Version]
- Lv, J.; Jiang, S.; Yang, Y.; Zhang, X.; Gao, R.; Cao, Y.; Song, G. FGIN-1-27 Inhibits Melanogenesis by regulating protein kinase A/cAMP-responsive element-binding, protein kinase C-β, and mitogen-activated protein kinase pathways. Front. Pharmacol. 2020, 11, 2043. [Google Scholar] [CrossRef] [PubMed]
- Azam, M.S.; Kwon, M.; Choi, J.; Kim, H.-R. Sargaquinoic acid ameliorates hyperpigmentation through cAMP and ERK-mediated downregulation of MITF in α-MSH-stimulated B16F10 cells. Biomed. Pharmacother. 2018, 104, 582–589. [Google Scholar] [CrossRef] [PubMed]
- Bae-Harboe, Y.-S.C.; Park, H.-Y. Tyrosinase: A central regulatory protein for cutaneous pigmentation. J. Investig. Dermatol. 2012, 132, 2678–2680. [Google Scholar] [CrossRef] [Green Version]
- Lee, A.; Kim, J.Y.; Heo, J.; Cho, D.-H.; Kim, H.-S.; An, I.-S.; An, S.; Bae, S. The inhibition of melanogenesis via the PKA and ERK signaling pathways by Chlamydomonas reinhardtii extract in B16F10 melanoma cells and artificial human skin equivalents. J. Investig. Dermatol. 2018, 28, 2121–2132. [Google Scholar] [CrossRef]
- Mann, T.; Scherner, C.; Röhm, K.-H.; Kolbe, L. Structure-activity relationships of thiazolyl resorcinols, potent and selective inhibitors of human tyrosinase. Int. J. Mol. Sci. 2018, 19, 690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshimori, A.; Oyama, T.; Takahashi, S.; Abe, H.; Kamiya, T.; Abe, T.; Tanuma, S.-i. Structure–activity relationships of the thujaplicins for inhibition of human tyrosinase. Bioorg. Med. Chem. 2014, 22, 6193–6200. [Google Scholar] [CrossRef]
- Li, X.; Yang, H.-W.; Jiang, Y.; Oh, J.-Y.; Jeon, Y.-J.; Ryu, B. Ishophloroglucin A isolated from Ishige okamurae suppresses melanogenesis induced by α-MSH: In Vitro and In Vivo. Mar. Drugs 2020, 18, 470. [Google Scholar] [CrossRef] [PubMed]
- Wibowo, J.P.; Batista, F.A.; van Oosterwijk, N.; Groves, M.R.; Dekker, F.J.; Quax, W.J. A novel mechanism of inhibition by phenylthiourea on PvdP, a tyrosinase synthesizing pyoverdine of Pseudomonas aeruginosa. Int. J. Biol. Macromol. 2020, 146, 212–221. [Google Scholar] [CrossRef]
- Choi, J.; Park, S.-J.; Jee, J.-G. Analogues of ethionamide, a drug used for multidrug-resistant tuberculosis, exhibit potent inhibition of tyrosinase. Eur. J. Med. Chem. 2015, 106, 157–166. [Google Scholar] [CrossRef]
- Patil, S.; Sistla, S.; Jadhav, J. Interaction of small molecules with human tyrosinase: A surface plasmon resonance and molecular docking study. Int. J. Biol. Macromol. 2016, 92, 1123–1129. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak No. | RT (min) | Identification | Molecular Formular | λ Max (nm) | [M+H]+ | [M–H]– | Product ion (m/z) |
|---|---|---|---|---|---|---|---|
| 1 | 7.83 | Puerarin-4’-O-β-D-glucopyranoside | C27H30O14 | 248,303 | 579.20 | 577.15 | 297.32, 417.20 |
| 2 | 9.89 | 3’-hydroxypuerarin-4’-O-dexyhexoside | C27H30O14 | 249,297 | 579.20 | 623.15 [M+HCOOH–H]– | 433.14, 283.16, 313.39 |
| 3 | 10.88 | 3’-hydroxypuerarin | C21H20O10 | 250 | 433.10 | 431.10 | 313.40, 283.20 |
| 4 | 15.43 | Puerarin | C21H20O9 | 250,305 | 417.10 | 415.09 | 297.12 |
| 5 | 16.74 | 3’-methoxy puerarin | C22H22O10 | 250,306 | 447.15 | 445.10 | 327.08, 297.07 |
| 6 | 17.33 | Puerarin xyloside | C26H28O13 | 250 | 549.15 | 547.15 | 417.12, 297.06 |
| 7 | 20.72 | Daidzin | C21H20O9 | 250,306 | 417.15 | 461.10 [M+HCOOH–H]– | 255.07 |
| 8 | 23.08 | 3’-Methoxydaidzin | C22H22O10 | 248,300 | 447.20 | 491.15 [M+HCOOH–H]– | 285.08 |
| 9 | 29.03 | Genistein 8-C-glucoside | C21H20O10 | 260 | 433.10 | 431.10 | 311.34, 283.21 |
| 10 | 36.82 | Ambocin | C26H28O14 | 249,300 | 565.20 | 563.15 | 311.05, 283.06 |
| 11 | 40.28 | Neopuerarin A | C21H20O9 | 250,306 | 417.15 | 415.10 | 295.38, 267.26 |
| 12 | 42.46 | Neopuerarin B | C26H28O14 | 250,306 | 417.15 | 415.10 | 295.38, 267.26 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, D.; Kim, J.H.; Kim, C.T.; Jeong, W.S.; Kim, H.M.; Sim, J.; Kang, J.S. Evaluation of Anti-Melanogenesis Activity of Enriched Pueraria lobata Stem Extracts and Characterization of Its Phytochemical Components Using HPLC–PDA–ESI–MS/MS. Int. J. Mol. Sci. 2021, 22, 8105. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22158105
Gao D, Kim JH, Kim CT, Jeong WS, Kim HM, Sim J, Kang JS. Evaluation of Anti-Melanogenesis Activity of Enriched Pueraria lobata Stem Extracts and Characterization of Its Phytochemical Components Using HPLC–PDA–ESI–MS/MS. International Journal of Molecular Sciences. 2021; 22(15):8105. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22158105
Chicago/Turabian StyleGao, Dan, Jin Hyeok Kim, Cheong Taek Kim, Won Seok Jeong, Hyung Min Kim, Jaehoon Sim, and Jong Seong Kang. 2021. "Evaluation of Anti-Melanogenesis Activity of Enriched Pueraria lobata Stem Extracts and Characterization of Its Phytochemical Components Using HPLC–PDA–ESI–MS/MS" International Journal of Molecular Sciences 22, no. 15: 8105. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22158105