
Modulation of Nrf2 and NF-κB Signaling Pathways by Naturally Occurring Compounds in Relation to Cancer Prevention and Therapy. Are Combinations Better Than Single Compounds?


Abstract
:1. Introduction
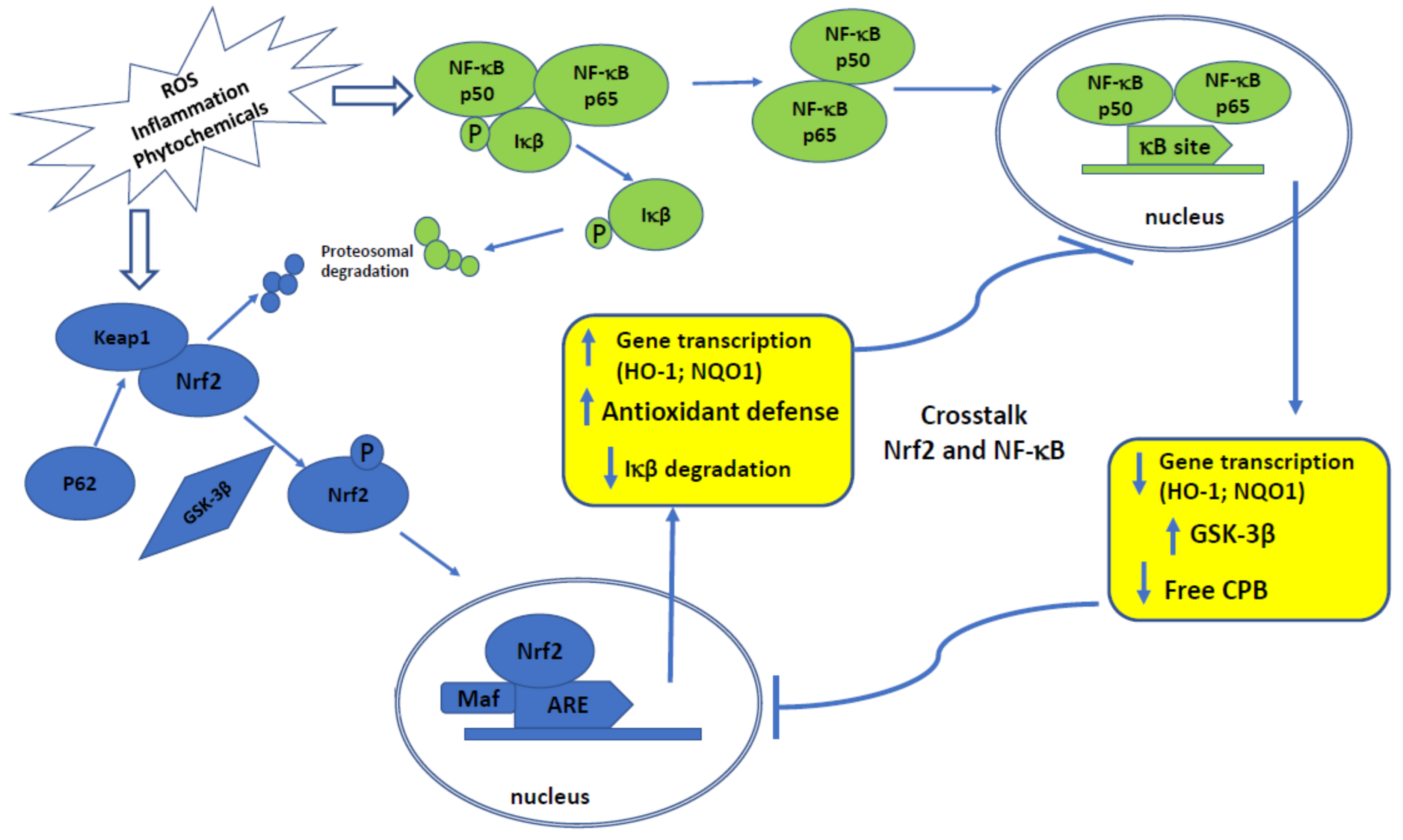
2. Overview of Nrf2 and NF-κB Signaling Pathways and Their Interconnections
3. Phytochemicals as Modulators of Nrf2 and NF-kB Signaling Pathway
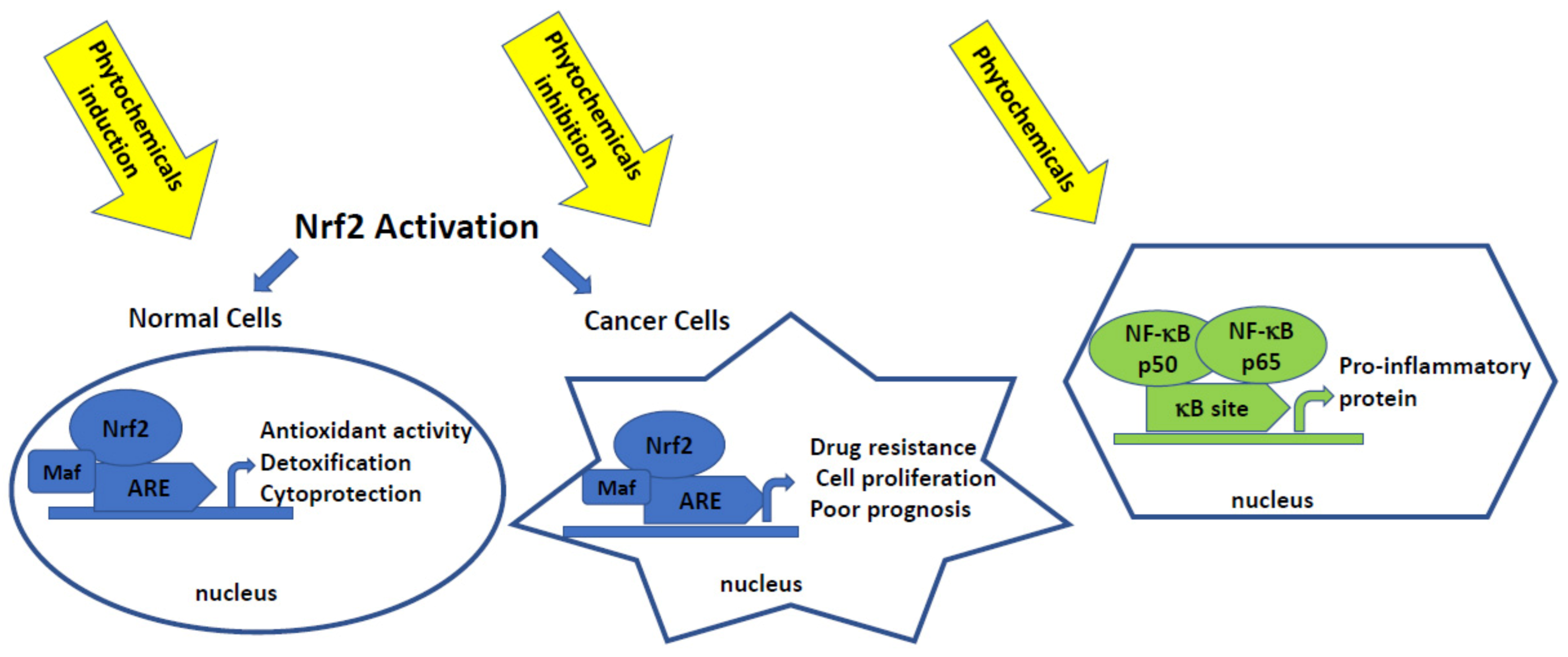
3.1. Phytochemicals and Nrf2-Handling Its Double Faces toward Cancer
3.2. Phytochemicals and NF-κB–Inhibitors Are Needed in Cancer Chemoprevention and Therapy
3.3. The Effect of Food Matrix-A Prototype of Combinations Strategy on Nrf2 and NF-κB Activities
3.4. Phytochemical Combinations Affecting Nrf2 and NF-κB Signaling Pathways

{kind=link}
{kind=link}
| Phytochemicals Combination | Phytochemicals Interactions | Experimental Model | Concentrations | Effect on NF-ĸB | Effect on Nrf2 | Ref. |
|---|---|---|---|---|---|---|
| Resveratrol and Phenethyl isothiocyanate | Synergism | Human Pancreatic cancer cells (Mia-Pa-Ca-2 cells) | * Resveratrol 10 µM; Phenethyl isothiocyanate 10 µM | ↑ expression of Nrf2 and binding Nrf2 to DNA, and expression of SOD, NQO1, GSTP | [87] | |
| Synergism | PANC-1 cells | Resveratrol 10 µM; Phenethyl isothiocyanate 10 µM | ↓ binding NF-ĸBp65 to DNA and expression of NF-ĸBp65 and COX-2 | [53] | ||
| Xanthohumol and Phenethyl isothiocyanate | Synergism | PANC-1 cells | Xanthohumol 10 µM; Phenethyl isothiocyanate 10 µM | ↑nuclear translocation of Nrf2, and binding Nrf2 to DNA, and expression of Nrf2, SOD, NQO1, GSTP | [53] | |
| Synergism | PANC-1 cells | Xanthohumol 10 µM; Phenethyl isothiocyanate 10 µM | ↓ nuclear translocation NF-ĸB, and binding NF-ĸBp65 and NF-ĸBp50 to DNA, and expression of NF-ĸB and COX-2 | [53] | ||
| Curcumin and Arctigenin | Synergism | Human prostate adenocarcinoma cells (LNCaP cells); MCF-7 cells | Curcumin 5 μM, Arctigenin 1μM | ↓ phosphorylation of NF-ĸB; and p-IκB levels | [88] | |
| Curcumin and Epigallocatechin gallate | Synergism | LNCaPcells; MCF-7 cells | Curcumin 5 μM; EGCG 40 μM | ↓ phosphorylation NF-ĸB; ↓ p-IκB levels | [88] | |
| 3,3′-Diindolylmethane and Sulforaphane | Additive | Human liver hepatoma cells (HepG2-C8 cells) | 3,3′-diindolylmethane 6.25µM; Sulforaphane 1µM | ↑ expression of Nrf2 and SOD | [89] | |
| Sulforaphane and Curcumin | Synergism | RAW264.7 cells | Sulforaphane 0.4 μM; Curcumin 2 μM | ↓ expression of iNOS; COX-2; PGE2 | [90] | |
| Synergism | RAW264.7 cells | Sulforaphane 0.4 μM; Curcumin 2 μM | ↑ expression of Nrf2 and NQO1, HO-1 | [90] | ||
| Sulforaphane and Phenethyl isothiocyanate | Synergism | RAW264.7 cells | Sulforaphane 0.4 μM; Phenethyl isothiocyanate 2 μM | ↓ expression of iNOS; COX-2; PGE2 | [90] | |
| Synergism | RAW264.7 cells | Sulforaphane 0.4 μM; Phenethyl isothiocyanate 2 μM | ↑ expression of Nrf2 and NQO1, HO-1 | [90] | ||
| Curcumin and Resveratrol | Synergism | Human hypopharyngeal carcinoma cells (Fadu cells) Human oral adenosquamous carcinoma cells (Cal-27 cells) | Curcumin 25 μM; Resveratrol 25 μM | ↓ nuclear translocation of NF-ĸB | [91] | |
| Synergism | Xenografts SCID mouse Spinal cord injury model | ** Curcumin 500 mg/kg; Resveratrol 150 mg/kg gavage | ↓ NF-ĸB binding to DNA | [92] | ||
| Curcumin and Piperine | Lack of Synergism | Holtzman rats Periodontitis model | Curcumin 400 mg/kg; Piperine 20 mg/kg gavage | ↓ phosphorylation and activation of NF-ĸB | [93] |
4. Conclusions and Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Albini, A.; Tosetti, F.; Li, V.W.; Noonan, D.M.; Li, W.W. Cancer Prevention by Targeting Angiogenesis. Nat. Rev. Clin. Oncol. 2012, 9, 498–509. [Google Scholar] [CrossRef]
- Hoesel, B.; Schmid, J.A. The Complexity of NF-ΚB Signaling in Inflammation and Cancer. Mol. Cancer 2013, 12, 86. [Google Scholar] [CrossRef] [Green Version]
- Nicolas, A.; Carré, M.; Pasquier, E. Metronomics: Intrinsic Anakoinosis Modulator? Front. Pharmacol. 2018, 9. [Google Scholar] [CrossRef] [Green Version]
- Dinkova-Kostova, A.T.; Liby, K.T.; Stephenson, K.K.; Holtzclaw, W.D.; Gao, X.; Suh, N.; Williams, C.; Risingsong, R.; Honda, T.; Gribble, G.W.; et al. Extremely Potent Triterpenoid Inducers of the Phase 2 Response: Correlations of Protection against Oxidant and Inflammatory Stress. Proc. Natl. Acad. Sci. USA 2005, 102, 4584–4589. [Google Scholar] [CrossRef] [Green Version]
- Singh, S.; Aggarwal, B.B. Activation of Transcription Factor NF-ΚB Is Suppressed by Curcumin (Diferuloylmethane) (∗). J. Biol. Chem. 1995, 270, 24995–25000. [Google Scholar] [CrossRef] [Green Version]
- Haque, A.; Brazeau, D.; Amin, A.R. Perspectives on Natural Compounds in Chemoprevention and Treatment of Cancer: An Update with New Promising Compounds. Eur. J. Cancer 2021, 149, 165–183. [Google Scholar] [CrossRef] [PubMed]
- Jung, B.-J.; Yoo, H.-S.; Shin, S.; Park, Y.-J.; Jeon, S.-M. Dysregulation of NRF2 in Cancer: From Molecular Mechanisms to Therapeutic Opportunities. Biomol. Ther. 2018, 26, 57–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saha, S.; Buttari, B.; Panieri, E.; Profumo, E.; Saso, L. An Overview of Nrf2 Signaling Pathway and Its Role in Inflammation. Molecules 2020, 25, 5474. [Google Scholar] [CrossRef] [PubMed]
- Krajka-Kuźniak, V.; Paluszczak, J.; Baer-Dubowska, W. The Nrf2-ARE Signaling Pathway: An Update on Its Regulation and Possible Role in Cancer Prevention and Treatment. Pharmacol. Rep. 2017, 69, 393–402. [Google Scholar] [CrossRef] [PubMed]
- Silva-Islas, C.A.; Maldonado, P.D. Canonical and Non-Canonical Mechanisms of Nrf2 Activation. Pharmacol. Res. 2018, 134, 92–99. [Google Scholar] [CrossRef]
- Shah, S.Z.A.; Zhao, D.; Hussain, T.; Sabir, N.; Mangi, M.H.; Yang, L. P62-Keap1-NRF2-ARE Pathway: A Contentious Player for Selective Targeting of Autophagy, Oxidative Stress and Mitochondrial Dysfunction in Prion Diseases. Front. Mol. Neurosci. 2018, 11, 310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, X.; Ou, Z.; Chen, R.; Niu, X.; Chen, D.; Kang, R.; Tang, D. Activation of the P62-Keap1-NRF2 Pathway Protects against Ferroptosis in Hepatocellular Carcinoma Cells. Hepatology 2016, 63, 173–184. [Google Scholar] [CrossRef] [PubMed]
- Raghunath, A.; Sundarraj, K.; Arfuso, F.; Sethi, G.; Perumal, E. Dysregulation of Nrf2 in Hepatocellular Carcinoma: Role in Cancer Progression and Chemoresistance. Cancers 2018, 10, 481. [Google Scholar] [CrossRef] [Green Version]
- Panieri, E.; Saso, L. Potential Applications of NRF2 Inhibitors in Cancer Therapy. Oxid. Med. Cell Longev. 2019, 2019, 8592348. [Google Scholar] [CrossRef] [Green Version]
- Song, M.-Y.; Lee, D.-Y.; Chun, K.-S.; Kim, E.-H. The Role of NRF2/KEAP1 Signaling Pathway in Cancer Metabolism. Int. J. Mol. Sci. 2021, 22, 4376. [Google Scholar] [CrossRef]
- Pires, B.R.B.; Silva, R.C.M.C.; Ferreira, G.M.; Abdelhay, E. NF-KappaB: Two Sides of the Same Coin. Genes 2018, 9, 24. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Xu, H.; Huang, K. Selenium in the Prevention of Atherosclerosis and Its Underlying Mechanisms. Metallomics 2017, 9, 21–37. [Google Scholar] [CrossRef]
- Giridharan, S.; Srinivasan, M. Mechanisms of NF-ΚB P65 and Strategies for Therapeutic Manipulation. J. Inflamm. Res. 2018, 11, 407–419. [Google Scholar] [CrossRef] [Green Version]
- Yu, H.; Lin, L.; Zhang, Z.; Zhang, H.; Hu, H. Targeting NF-ΚB Pathway for the Therapy of Diseases: Mechanism and Clinical Study. Signal Transduct. Target Ther. 2020, 5, 209. [Google Scholar] [CrossRef] [PubMed]
- Wan, F.; Lenardo, M.J. Specification of DNA Binding Activity of NF-KappaB Proteins. Cold Spring Harb. Perspect. Biol. 2009, 1, a000067. [Google Scholar] [CrossRef] [Green Version]
- Rius-Pérez, S.; Pérez, S.; Martí-Andrés, P.; Monsalve, M.; Sastre, J. Nuclear Factor Kappa B Signaling Complexes in Acute Inflammation. Antioxid. Redox Signal. 2020, 33, 145–165. [Google Scholar] [CrossRef]
- Verzella, D.; Pescatore, A.; Capece, D.; Vecchiotti, D.; Ursini, M.V.; Franzoso, G.; Alesse, E.; Zazzeroni, F. Life, Death, and Autophagy in Cancer: NF-ΚB Turns up Everywhere. Cell Death Dis. 2020, 11, 210. [Google Scholar] [CrossRef] [Green Version]
- Sandberg, M.; Patil, J.; D’Angelo, B.; Weber, S.G.; Mallard, C. NRF2-Regulation in Brain Health and Disease: Implication of Cerebral Inflammation. Neuropharmacology 2014, 79, 298–306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wardyn, J.D.; Ponsford, A.H.; Sanderson, C.M. Dissecting Molecular Cross-Talk between Nrf2 and NF-ΚB Response Pathways. Biochem. Soc. Trans 2015, 43, 621–626. [Google Scholar] [CrossRef] [Green Version]
- Liu, G.-H.; Qu, J.; Shen, X. NF-ΚB/P65 Antagonizes Nrf2-ARE Pathway by Depriving CBP from Nrf2 and Facilitating Recruitment of HDAC3 to MafK. Biochim. Biophys. Acta Mol. Cell Res. 2008, 1783, 713–727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaspar, J.W.; Niture, S.K.; Jaiswal, A.K. Nrf2:INrf2 (Keap1) Signaling in Oxidative Stress. Free Radic. Biol. Med. 2009, 47, 1304–1309. [Google Scholar] [CrossRef] [Green Version]
- González-Vallinas, M.; González-Castejón, M.; Rodríguez-Casado, A.; Ramírez de Molina, A. Dietary Phytochemicals in Cancer Prevention and Therapy: A Complementary Approach with Promising Perspectives. Nutr. Rev. 2013, 71, 585–599. [Google Scholar] [CrossRef]
- Rue, E.A.; Rush, M.D.; van Breemen, R.B. Procyanidins: A Comprehensive Review Encompassing Structure Elucidation via Mass Spectrometry. Phytochem. Rev. 2018, 17, 1–16. [Google Scholar] [CrossRef]
- Rasouli, H.; Farzaei, M.H.; Khodarahmi, R. Polyphenols and Their Benefits: A Review. Int. J. Food Prop. 2017, 20, 1700–1741. [Google Scholar] [CrossRef] [Green Version]
- Galluzzi, L.; Vitale, I.; Aaronson, S.A.; Abrams, J.M.; Adam, D.; Agostinis, P.; Alnemri, E.S.; Altucci, L.; Amelio, I.; Andrews, D.W.; et al. Molecular Mechanisms of Cell Death: Recommendations of the Nomenclature Committee on Cell Death 2018. Cell Death Differ. 2018, 25, 486–541. [Google Scholar] [CrossRef]
- Shin, S.A.; Joo, B.J.; Lee, J.S.; Ryu, G.; Han, M.; Kim, W.Y.; Park, H.H.; Lee, J.H.; Lee, C.S. Phytochemicals as Anti-Inflammatory Agents in Animal Models of Prevalent Inflammatory Diseases. Molecules 2020, 25, 5932. [Google Scholar] [CrossRef] [PubMed]
- Clifford, T.; Acton, J.P.; Cocksedge, S.P.; Davies, K.A.B.; Bailey, S.J. The Effect of Dietary Phytochemicals on Nuclear Factor Erythroid 2-Related Factor 2 (Nrf2) Activation: A Systematic Review of Human Intervention Trials. Mol. Biol. Rep. 2021, 48, 1745–1761. [Google Scholar] [CrossRef] [PubMed]
- Gugliandolo, A.; Bramanti, P.; Mazzon, E. Activation of Nrf2 by Natural Bioactive Compounds: A Promising Approach for Stroke? Int. J. Mol. Sci. 2020, 21, 4875. [Google Scholar] [CrossRef] [PubMed]
- Baer-Dubowska, W.; Szaefer, H.; Majchrzak-Celińska, A.; Krajka-Kuźniak, V. Tannic Acid: Specific Form of Tannins in Cancer Chemoprevention and Therapy-Old and New Applications. Curr. Pharmacol. Rep. 2020, 6, 28–37. [Google Scholar] [CrossRef] [Green Version]
- Clifford, T.; Howatson, G.; West, D.J.; Stevenson, E.J. The Potential Benefits of Red Beetroot Supplementation in Health and Disease. Nutrients 2015, 7, 2801–2822. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, D.V.T.; Baião, D.D.S.; Ferreira, V.F.; Paschoalin, V.M.F. Betanin as a Multipath Oxidative Stress and Inflammation Modulator: A Beetroot Pigment with Protective Effects on Cardiovascular Disease Pathogenesis. Crit. Rev. Food Sci. Nutr. 2020, 1–16. [Google Scholar] [CrossRef]
- Krajka-Kuźniak, V.; Paluszczak, J.; Szaefer, H.; Baer-Dubowska, W. Betanin, a Beetroot Component, Induces Nuclear Factor Erythroid-2-Related Factor 2-Mediated Expression of Detoxifying/Antioxidant Enzymes in Human Liver Cell Lines. Br. J. Nutr. 2013, 110, 2138–2149. [Google Scholar] [CrossRef] [Green Version]
- Harish, V.; Haque, E.; Śmiech, M.; Taniguchi, H.; Jamieson, S.; Tewari, D.; Bishayee, A. Xanthohumol for Human Malignancies: Chemistry, Pharmacokinetics and Molecular Targets. Int. J. Mol. Sci. 2021, 22, 4478. [Google Scholar] [CrossRef]
- Krajka-Kuźniak, V.; Paluszczak, J.; Baer-Dubowska, W. Xanthohumol Induces Phase II Enzymes via Nrf2 in Human Hepatocytes in Vitro. Toxicol. Vitro 2013, 27, 149–156. [Google Scholar] [CrossRef]
- Lee, I.-S.; Lim, J.; Gal, J.; Kang, J.C.; Kim, H.J.; Kang, B.Y.; Choi, H.J. Anti-Inflammatory Activity of Xanthohumol Involves Heme Oxygenase-1 Induction via NRF2-ARE Signaling in Microglial BV2 Cells. Neurochem. Int. 2011, 58, 153–160. [Google Scholar] [CrossRef]
- Lv, H.; Liu, Q.; Wen, Z.; Feng, H.; Deng, X.; Ci, X. Xanthohumol Ameliorates Lipopolysaccharide (LPS)-Induced Acute Lung Injury via Induction of AMPK/GSK3β-Nrf2 Signal Axis. Redox Biol. 2017, 12, 311–324. [Google Scholar] [CrossRef]
- Lněničková, K.; Šadibolová, M.; Matoušková, P.; Szotáková, B.; Skálová, L.; Boušová, I. The Modulation of Phase II Drug-Metabolizing Enzymes in Proliferating and Differentiated CaCo-2 Cells by Hop-Derived Prenylflavonoids. Nutrients 2020, 12, 2138. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Ramirez, C.N.; Su, Z.-Y.; Kong, A.-N.T. Epigenetic Modifications of Triterpenoid Ursolic Acid in Activating Nrf2 and Blocking Cellular Transformation of Mouse Epidermal Cells. J. Nutr. Biochem. 2016, 33, 54–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chian, S.; Thapa, R.; Chi, Z.; Wang, X.J.; Tang, X. Luteolin Inhibits the Nrf2 Signaling Pathway and Tumor Growth in Vivo. Biochem. Biophys. Res. Commun. 2014, 447, 602–608. [Google Scholar] [CrossRef]
- Arlt, A.; Sebens, S.; Geismann, C.; Grossmann, M.; Krise, M.L.; Schreiber, S.; Schäfer, H. Inhibition of the Nrf2 Transcription Factor by the Alkaloid Through Decreased Proteosomal Gene Expression and Proteosome Activity. Oncogene 2013, 32, 4825–4835. [Google Scholar] [CrossRef]
- Ohnuma, T.; Matsumoto, T.; Itoi, A.; Kawana, A.; Nishiyama, T.; Ogura, K.; Hiratsuka, A. Enhanced Sensitivity of A549 Cells to the Cytotoxic Action of Anticancer Drugs via Suppression of Nrf2 by Procyanidins from Cinnamomi Cortex Extract. Biochem. Biophys. Res. Commun. 2011, 413, 623–629. [Google Scholar] [CrossRef] [PubMed]
- De Prax, M.C.A.; Ferro, K.P.V.; Santos, I.; Torello, C.O.; Salazar-Terreros, M.; Olalla Saad, S.T. NRF2 Is Targeted By the Polyphenol Quercetin and Induces Apoptosis, in Part, through up Regulation of Pro Apoptotic Mirs. Blood 2019, 134, 2529. [Google Scholar] [CrossRef]
- Bhattacharjee, S.; Dashwood, R.H. Epigenetic Regulation of NRF2/KEAP1 by Phytochemicals. Antioxidants 2020, 9, 865. [Google Scholar] [CrossRef]
- Pulido-Moran, M.; Moreno-Fernandez, J.; Ramirez-Tortosa, C.; Ramirez-Tortosa, M. Curcumin and Health. Molecules 2016, 21, 264. [Google Scholar] [CrossRef]
- Nelson, K.M.; Dahlin, J.L.; Bisson, J.; Graham, J.; Pauli, G.F.; Walters, M.A. The Essential Medicinal Chemistry of Curcumin. J. Med. Chem. 2017, 60, 1620–1637. [Google Scholar] [CrossRef]
- Cichocki, M.; Blumczyńska, J.; Baer-Dubowska, W. Naturally Occurring Phenolic Acids Inhibit 12-O-Tetradecanoylphorbol-13-Acetate Induced NF-ΚB, INOS and COX-2 Activation in Mouse Epidermis. Toxicology 2010, 268, 118–124. [Google Scholar] [CrossRef] [PubMed]
- Reddy, M.K.; Alexander-Lindo, R.L.; Nair, M.G. Relative Inhibition of Lipid Peroxidation, Cyclooxygenase Enzymes, and Human Tumor Cell Proliferation by Natural Food Colors. J. Agric. Food Chem. 2005, 53, 9268–9273. [Google Scholar] [CrossRef] [PubMed]
- Krajka-Kuźniak, V.; Cykowiak, M.; Szaefer, H.; Kleszcz, R.; Baer-Dubowska, W. Combination of Xanthohumol and Phenethyl Isothiocyanate Inhibits NF-ΚB and Activates Nrf2 in Pancreatic Cancer Cells. Toxicol. Vitro 2020, 65, 104799. [Google Scholar] [CrossRef]
- Guo, W.; Xu, B.; Zheng, B.; Du, J.; Liu, S. The Analysis of the Anti-Tumor Mechanism of Ursolic Acid Using Connectively Map Approach in Breast Cancer Cells Line MCF-7. Cancer Manag. Res. 2020, 12, 3469–3476. [Google Scholar] [CrossRef]
- Paredes-Gonzalez, X.; Fuentes, F.; Jeffery, S.; Saw, C.L.L.; Shu, L.; Su, Z.Y.; Kong, A.N.T. Induction of Nrf2-Mediated Gene Expression by Dietary Phytochemical Flavones Apigenin and Luteolin. Biopharm. Drug Dispos. 2015, 36, 440–451. [Google Scholar] [CrossRef]
- Witkowska-Banaszczak, E.; Krajka-Kuźniak, V.; Papierska, K. The Effect of Luteolin 7-Glucoside, Apigenin 7-Glucoside and Succisa Pratensis Extracts on NF-ΚB Activation and α-Amylase Activity in HepG2 Cells. Acta Biochim. Pol. 2020, 67, 41–47. [Google Scholar] [CrossRef]
- Licznerska, B.; Szaefer, H.; Krajka-Kuźniak, V. R-Sulforaphane Modulates the Expression Profile of AhR, ERα, Nrf2, NQO1, and GSTP in Human Breast Cell Lines. Mol. Cell Biochem. 2021, 476, 525–533. [Google Scholar] [CrossRef] [PubMed]
- Heiss, E.; Herhaus, C.; Klimo, K.; Bartsch, H.; Gerhäuser, C. Nuclear Factor Kappa B Is a Molecular Target for Sulforaphane-Mediated Anti-Inflammatory Mechanisms. J. Biol. Chem. 2001, 276, 32008–32015. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.; Choi, S.Y.; Choo, Y.Y.; Kim, O.; Tran, P.T.; Dao, C.T.; Min, B.S.; Lee, J.H. Sappanone A Exhibits Anti-Inflammatory Effects via Modulation of Nrf2 and NF-κB. Int. Immunopharmacol. 2015, 28, 328–336. [Google Scholar] [CrossRef]
- Rogati, F.; Millán, E.; Appendino, G.; Correa, A.; Caprioglio, D.; Minassi, A.; Muňoz, E. Identification of a Strigoterpenoid with Dual Nrf2 and NF-ĸB Modulatory Activity. ACS Med. Chem. Lett. 2019, 8, 606–610. [Google Scholar] [CrossRef]
- Xu, X.; Zhang, X.; Zhang, Y.; Yang, L.; Liu, Y.; Huang, S.; Lu, L.; Kong, L.; Li, Z.; Guo, Q.; et al. Wogonin Reserved Resistant Human Myelogenous Leukemia Cells via Inhibiting Nrf2 Signaling by Stat3/NF-ĸB Inactivation. Sci. Rep. 2017, 7, 39950. [Google Scholar] [CrossRef] [Green Version]
- Zhong, Y.; Zhang, F.; Sun, Z.; Zhou, W.; Li, Z.Y.; You, Q.D.; Guo, Q.L.; Hu, R. Drug Resistance Associates with Activation of Nrf2 in MCF-7/DOX Cells, and Wogonin Reserves it By Down-Regulating Nrf2-Mediated Cellular Defense Response. Mol. Carcinog. 2013, 52, 824–834. [Google Scholar] [CrossRef]
- Chen, B.; Zhang, Y.; Wang, Y.; Rao, J.; Jiang, X.; Xu, Z. Curcumin Inhibits Proliferation of Breast Cancer Cells Through Nrf2-Mediated Down-Regulation of Fen1 Expression. J. Steroid Biochem. Mol. Biol. 2014, 143, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Pimentel-Gutiérrez, H.J.; Bobadilla-Morales, L.; Barba-Barba, C.C.; Ortega-De-La-Torre, C.; Sánchez-Zubieta, F.A.; Corona-Rivera, J.R.; González-Quezada, B.A.; Armendáriz-Borunda, J.S.; Silva-Cruz, R.; Corona-Rivera, A. Curcumin Potentiates the Effect of Chemotherapy Against Acute Lymphoblastic Leukemia Cells Via Downregulation of NF-ĸB. Oncol. Lett. 2016, 5, 4117–4124. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.; Sun, Y.; Zhu, J.; Yu, L.; Jiang, X.; Zhang, J.; Dong, X.; Ma, B.; Zhang, Q. Oridonin Exerts Anticancer Effect on Osteosarcoma by Activating PPAR-γ and Inhibiting Nrf2 Pathway. Cell Death Dis. 2018, 9, 15. [Google Scholar] [CrossRef] [Green Version]
- Krajka-Kuźniak, V.; Szaefer, H.; Stefański, T.; Sobiak, S.; Cichocki, M.; Baer-Dubowska, W. The Effect of Resveratrol and Its Methylthioderivatives on the Nrf2-ARE Pathway in Mouse Epidermis and HaCaT Keratinocytes. Cell Mol. Biol. Lett. 2014, 19, 500–516. [Google Scholar] [CrossRef] [PubMed]
- Szaefer, H.; Cichocki, M.; Krajka-Kuźniak, V.; Stefański, T.; Sobiak, S.; Licznerska, B.; Baer-Dubowska, W. The Effect of Resveratrol and Its Methylthioderivatives on NF-ĸB and AP-1 signaling Pathways in HaCaT Keratinocytes. Pharmacol. Rep. 2014, 66, 732–740. [Google Scholar] [CrossRef]
- Kundu, J.K.; Shin, Y.K.; Kim, S.H.; Surh, Y.-J. Resveratrol Inhibits Phorbol Ester-Induced Expression of COX-2 and Activation of NF-KappaB in Mouse Skin by Blocking IkappaB Kinase Activity. Carcinogenesis 2006, 27, 1465–1474. [Google Scholar] [CrossRef] [PubMed]
- Singh, B.; Shoulson, R.; Chatterjee, A.; Ronghe, A.; Bhat, N.K.; Dim, D.C.; Bhat, H.K. Resveratrol Inhibits Estrogen-Induced Breast Carcinogenesis Through Induction of Nrf2-Mediated Protective Pathways. Carcinogenesis 2014, 35, 1872–1880. [Google Scholar] [CrossRef] [Green Version]
- Gong, L.; Li, Y.; Nedeljkovic-Kurepa, A.; Sarkar, F.H. Inactivation of NF-KappaB by Genistein Is Mediated via Akt Signaling Pathway in Breast Cancer Cells. Oncogene 2003, 22, 4702–4709. [Google Scholar] [CrossRef] [Green Version]
- Sahin, K.; Yenice, E.; Bilir, B.; Orhan, C.; Tuzcu, M.; Sahin, N.; Ozercan, I.H.; Kabil, N.; Ozpolat, B.; Kucuk, O. Genistein Prevents Development of Spontaneous Ovarian Cancer and Inhibits Tumor Growth in Hen Model. Cancer Prev. Res. 2019, 12, 135–146. [Google Scholar] [CrossRef] [Green Version]
- Mukherjee, S.; Siddiqui, M.A.; Dayal, S.; Ayoub, Y.Z.; Malathi, K. Epigallocatechin-3-gallate Suppresses Proinflammatory Cytokines and Chemokines Induced by Toll-like Receptor 9 Agonists in Prostate Cancer Cells. J. Inflamm. Res. 2014, 7, 89–101. [Google Scholar] [CrossRef] [Green Version]
- Han, J.; Wang, M.; Jing, X.; Shi, H.; Ren, M.; Lou, H. (-)-Epigallocatechin Gallate Protects against Cerebral Ischemia-Induced Oxidative Stress via Nrf2/ARE Signaling. Neurochem Res. 2014, 39, 1292–1299. [Google Scholar] [CrossRef]
- Zhang, N.; Dou, D.; Ran, X.; Kang, T. Neuroprotective Effect of Arctigenin against Neuroinflammation and Oxidative Stress Induced by Rotenone. RSC Adv. 2018, 8, 2280–2292. [Google Scholar] [CrossRef] [Green Version]
- Rehman, M.U.; Rashid, S.; Arafah, A.; Qamar, W.; Alsaffar, R.M.; Ahmad, A.; Almatroudi, N.M.; Alqahtani, S.M.A.; Rashid, S.M.; Ahmad, S.B. Piperine Regulates Nrf-2/Keap-1 Signalling and Exhibits Anticancer Effect in Experimental Colon Carcinogenesis in Wistar Rats. Biology 2020, 9, 302. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.-M.; Ma, J.-Q.; Xie, W.-R.; Liu, S.-S.; Feng, Z.-J.; Zheng, G.-H.; Wang, A.-M. Quercetin Protects Mouse Liver against Nickel-Induced DNA Methylation and Inflammation Associated with the Nrf2/HO-1 and P38/STAT1/NF-ΚB Pathway. Food Chem. Toxicol. 2015, 82, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Szaefer, H.; Krajka-Kuźniak, V.; Ignatowicz, E.; Adamska, T.; Baer-Dubowska, W. Evaluation of the Effect of Beetroot Juice on DMBA-Induced Damage in Liver and Mammary Gland of Female Sprague-Dawley Rats. Phytother. Res. 2014, 28, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Tan, M.L.; Hamid, S.B.S. Beetroot as a Potential Functional Food for Cancer Chemoprevention, a Narrative Review. J. Cancer Prev. 2021, 26, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Krajka-Kuźniak, V.; Szaefer, H.; Ignatowicz, E.; Adamska, T.; Baer-Dubowska, W. Beetroot Juice Protects against N-Nitrosodiethylamine-Induced Liver Injury in Rats. Food Chem. Toxicol. 2012, 50, 2027–2033. [Google Scholar] [CrossRef]
- El Gamal, A.A.; Al Said, M.S.; Raish, M.; Al-Sohaibani, M.; Al-Massarani, S.M.; Ahmad, A.; Hefnawy, M.; Al-Yahya, M.; Basoudan, O.A.; Rafatullah, S. Beetroot (Beta vulgaris L.) Extract Ameliorates Gentamicin-Induced Nephrotoxicity Associated Oxidative Stress, Iflammation, and Apoptosis in Rodent Model. Mediat. Inflamm. 2014, 2014, 983952. [Google Scholar] [CrossRef] [Green Version]
- Hyun, T.K.; Jang, K.-I. Apple as a Source of Dietary Phytonutrients: An Update on the Potential Health Benefits of Apple. EXCLI J. 2016, 15, 565–569. [Google Scholar] [CrossRef]
- Lodi, A.; Saha, A.; Lu, X.; Wang, B.; Sentandreu, E.; Collins, M.; Kolonin, M.G.; DiGiovanni, J.; Tiziani, S. Combinatorial Treatment with Natural Compounds in Prostate Cancer Inhibits Prostate Tumor Growth and Leads to Key Modulations of Cancer Cell Metabolism. NPJ Precis. Onc. 2017, 1, 1–12. [Google Scholar] [CrossRef]
- Tremmel, L.; Rho, O.; Slaga, T.J.; DiGiovanni, J. Inhibition of Skin Tumor Promotion by TPA Using a Combination of Topically Applied Ursolic Acid and Curcumin. Mol. Carcinog. 2019, 58, 185–195. [Google Scholar] [CrossRef]
- Saw, C.L.L.; Guo, Y.; Yang, A.Y.; Paredes-Gonzalez, X.; Ramirez, C.; Pung, D.; Kong, A.-N.T. The Berry Constituents Quercetin, Kaempferol, and Pterostilbene Synergistically Attenuate Reactive Oxygen Species: Involvement of the Nrf2-ARE Signaling Pathway. Food Chem Toxicol. 2014, 72, 303–311. [Google Scholar] [CrossRef]
- Hackman, G.L.; Collins, M.; Lu, X.; Lodi, A.; DiGiovanni, J.; Tiziani, S. Predicting and Quantifying Antagonistic Effects of Natural Compounds Given with Chemotherapeutic Agents: Applications for High-Throughput Screening. Cancers 2020, 12, 3714. [Google Scholar] [CrossRef]
- Probst, B.L.; McCauley, L.; Trevino, I.; Wigley, W.C.; Ferguson, D.A. Cancer Cell Growth Is Differentially Affected by Constitutive Activation of NRF2 by KEAP1 Deletion and Pharmacological Activation of NRF2 by the Synthetic Triterpenoid, RTA 405. PLoS ONE 2015, 10, e0135257. [Google Scholar] [CrossRef] [Green Version]
- Cykowiak, M.; Krajka-Kuźniak, V.; Baer-Dubowska, W. Combinations of Phytochemicals More Efficiently than Single Components Activate Nrf2 and Induce the Expression of Antioxidant Enzymes in Pancreatic Cancer Cells. Nutr. Cancer 2021, 1–16. [Google Scholar] [CrossRef]
- Wang, P.; Wang, B.; Chung, S.; Wu, Y.; Henning, S.M.; Vadgama, J.V. Increased Chemopreventive Effect by Combining Arctigenin, Green Tea Polyphenol and Curcumin in Prostate and Breast Cancer Cells. RSC Adv. 2014, 4, 35242–35250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saw, C.L.L.; Cintron, M.; Wu, T.Y.; Guo, Y.; Huang, Y.; Jeong, W.S.; Kong, A.N.T. Pharmacodynamics of dietary phytochemical indoles I3C and DIM: Induction of Nrf2-mediated Phase II Drug Metabolizing and Antioxidant Genes and Synergism with isothiocyanates. Biopharm. Drug Dispos. 2011, 35, 289–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheung, K.L.; Khor, T.O.; Kong, A.-N. Synergistic Effect of Combination of Phenethyl Isothiocyanate and Sulforaphane or Curcumin and Sulforaphane in the Inhibition of Inflammation. Pharm. Res. 2009, 26, 224–231. [Google Scholar] [CrossRef] [PubMed]
- Masuelli, L.; Di Stefano, E.; Fantini, M.; Mattera, R.; Benvenuto, M.; Marzocchella, L.; Sacchetti, P.; Focaccetti, C.; Bernardini, R.; Tresoldi, I.; et al. Resveratrol Potentiates the in Vitro and in Vivo Anti-Tumoral Effects of Curcumin in Head and Neck Carcinomas. Oncotarget 2014, 5, 10745–10762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Majumdar, A.P.N.; Banerjee, S.; Nautiyal, J.; Patel, B.B.; Patel, V.; Du, J.; Yu, Y.; Elliott, A.A.; Levi, E.; Sarkar, F.H. Curcumin Synergizes with Resveratrol to Inhibit Colon Cancer. Nutr. Cancer 2009, 61, 544–553. [Google Scholar] [CrossRef] [PubMed]
- Guimaraes-Stabili, M.R.; de Aquino, S.G.; de Almeida Curylofo, F.; Tasso, C.O.; Rocha, F.R.G.; de Medeiros, M.C.; de Pizzol, J.P.; Cerri, P.S.; Romito, G.A.; Rossa, C. Systemic Administration of Curcumin or Piperine Enhances the Periodontal Repair: A Preliminary Study in Rats. Clin Oral. Investig. 2019, 23, 3297–3306. [Google Scholar] [CrossRef] [PubMed]


| Phytochemical | Source | Experimental Model | Concentrations | Effect on NF-ĸB | Effect on Nrf2 | Ref. |
|---|---|---|---|---|---|---|
| Xanthohumol/Chalcone | Humulus lupulus | Human pancreatic cancer cells (PANC-1 cells) | * 5 µM and 10 µM | ↑ expression of Nrf2, binding Nrf2 to DNA and expression of antioxidant enzymes (SOD, NQO1, GSTP) | [53] | |
| PANC-1 cells | 5 µM and 10 µM | ↓ expression of NF-ĸB, binding NF-ĸBp65 and NF-ĸBp50 to DNA, and expression of COX-2 | [53] | |||
| Apigenin/Flavone | Matricaria chamomilla | Human hepatocellular liver carcinoma cells (HepG2 cells) | 6.25 µM | ↓ mRNA and protein levels of Nrf2, HO-1 | [55] | |
| HepG2 cells | 10 µg/mL | ↓ mRNA and protein levels of NF-ĸBp50 and NF-ĸBp65 | [56] | |||
| Phenethyl isothiocyanate/Isothiocyanate | Brassicaceae | PANC-1 cells | 5 µM and 10 µM | ↑ expression of Nrf2, binding Nrf2 to DNA, expression of SOD, NQO1, GSTP | [53] | |
| PANC-1 cells | 5 µM and 10 µM | ↓ expression of NF-ĸB, binding NF-ĸBp65 and NF-ĸBp50 to DNA, and expression of COX-2 | [53] | |||
| Sulforaphane/Isothiocyanate | Brassicaceae | Human breast adenocarcinoma cells (MCF-7 cells, MDA-MB231 cells) | 5 µM and 10 µM 20 µM | ↑ expression of Nrf2, and GSTP, NQO1 | [57] | |
| Murine macrophage (RAW264.7 cells) | 5 µM and 15 µM | ↓ translocation of NF-κB and expression of COX-2 | [58] | |||
| Sappanone/Isoflavone | Caesalpinia sappan | RAW264.7 cells | 30 µM | ↑ nuclear translocation of Nrf2 and expression of HO-1, NQO1 | [59] | |
| RAW264.7 cells | 30 µM | ↓ nuclear translocation of NF-ĸBp65 | [59] | |||
| Strigoterpenoid/Terpenoid | Pistachia terebinthus | Human immortalized keratinocytes (HaCaT cells) | 10 µM | ↑ activation of Nrf2 | [60] | |
| HaCaT cells | 10 µM | ↓ expression of NF-ĸB p65 ↑ expression of IKKβ | [60] | |||
| Wogonin/Flavone | Scutelaria baicalensis | Human erythroleukemic cells (K562 cells) | 40 µM | ↓ NF-ĸB activation and NF-ĸBp65 binding to DNA | [61] | |
| K562 cells | 40 µM | ↓ expression of Nrf2 and nuclear translocation of Nrf2 | [61] | |||
| MCF-7 cells | 40 µM 60 µM | ↓ expression of Nrf2 ↑ Keap1 protein level; ↓HO-1 and NQO1 protein level | [62] | |||
| Curcumin/diferuloylmethane | Curcuma longa rhizomes | MCF-7 cells | 20 µM and 25 µM and 40 µM | ↑ expression and protein of Nrf2 | [63] | |
|
Human lymphocytic leukemia cells (REH cells) | 20 µM | ↓expression of NF-ĸB | [64] | |||
| Oridinin/Diterpenoid | Rabdosia rubescens | Human osteosarcoma cells (MG-63 cells HOS cells) | 15 µM | ↓ NF-ĸB activation and NF-ĸB p65 binding to DNA | ↓ nuclear translocation of Nrf2, expression of HO-1 and NQO1 | [65] |
| Male BALB/c nude mice | 30 mg/kg intraperitoneally | ↓ nuclear translocation of NF-ĸB | ↓ nuclear translocation of Nrf2 | [65] | ||
| Resveratrol/Stilbene | Vitis vinifera | HaCaT cells | 60 µM | ↑ nuclear level of Nrf2 and GSTP protein level | [66] | |
| HaCaT cells | 60 µM | ↓NF-ĸBp65, IĸB kinase | [67] | |||
| Female ICR mice | 0.25 µmol and 1 µmol topically applied | ↓expression of NF-ĸBp65 and COX-2, and IKK activity | [68] | |||
| Female ACI rats | 50 mg as a subcutaneous pellet | ↑ expression and protein levels of Nrf2 and HO-1, NQO1 | [69] | |||
| Genistein/Isoflavone | Genista tinctoria | MDA-MB-231 cells | 50 µM | ↓ expression of NF-ĸBp65 and p50, COX-2 | [70] | |
| Laying Hen model ovarian cancer | ** 52.48 mg/hen and 106.26 mg/hen | ↑ expression of Nrf2 and HO-1 | [71] | |||
| Laying Hen model ovarian cancer | 52.48 mg/hen and 106.26 mg/hen | ↓expression of NF-ĸB | [71] | |||
| Epigallocatechin gallate/Flavonoid | Camellia sinensis | Human prostate cancer cells (DU145 cells) | 40 µg/mL | ↓ nuclear translocation of NF-ĸBp65 | [72] | |
| Sprague-Dawley rats | 40 mg/kg intraperitoneally | ↑ expression and protein level of Nrf2 and HO-1 | [73] | |||
| Arctigenin/Lignan | Fructus arctii | SD male rats | 20 mg/kg orally | ↓ expression of NF-ĸB, COX-2 | [74] | |
| SD male rats | 20 mg/kg orally | ↑ expression of SOD | [74] | |||
| Piperine/Alkaloid | Piper nigrum | Male Wistar rats | 30 mg/kg and 60 mg/kg orally | ↑ expression of Nrf2, HO-1, NOQ1 | [75] | |
| Male Wistar rats | 30 mg/kg and 60 mg/kg orally | ↓ expression of NF-ĸB, iNOS, COX-2 | [75] | |||
| Quercetin/Flavonoid | Pinus banksiana | Male ICR mice | 40 mg/kg and 80 mg/kg gavage | ↑ nuclear translocation of Nrf2 and expression of HO-1 | [76] | |
| Male ICR mice | 40 mg/kg and 80 mg/kg gavage | ↓ nuclear translocation of NF-ĸBp65 | [76] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Krajka-Kuźniak, V.; Baer-Dubowska, W. Modulation of Nrf2 and NF-κB Signaling Pathways by Naturally Occurring Compounds in Relation to Cancer Prevention and Therapy. Are Combinations Better Than Single Compounds? Int. J. Mol. Sci. 2021, 22, 8223. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22158223
Krajka-Kuźniak V, Baer-Dubowska W. Modulation of Nrf2 and NF-κB Signaling Pathways by Naturally Occurring Compounds in Relation to Cancer Prevention and Therapy. Are Combinations Better Than Single Compounds? International Journal of Molecular Sciences. 2021; 22(15):8223. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22158223
Chicago/Turabian StyleKrajka-Kuźniak, Violetta, and Wanda Baer-Dubowska. 2021. "Modulation of Nrf2 and NF-κB Signaling Pathways by Naturally Occurring Compounds in Relation to Cancer Prevention and Therapy. Are Combinations Better Than Single Compounds?" International Journal of Molecular Sciences 22, no. 15: 8223. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22158223