
Molecular Drivers of Developmental Arrest in the Human Preimplantation Embryo: A Systematic Review and Critical Analysis Leading to Mapping Future Research

 ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
3. Results
3.1. Intrinsic Embryonic Factors Leading to Arrest
3.1.1. The Role of Embryonic Gene Variation and Expression
3.1.2. The Role of Mitochondrial DNA
3.1.3. The Role of Methylation Patterns
3.1.4. The Role of Small Noncoding RNA
3.1.5. The Role of Chromosomal Abnormalities
3.1.6. The Role of the Embryonic Metabolic Profile
3.2. The Association between Embryo Morphological Characteristics and Arrested Development
3.3. Maternal Factors Leading to Preimlantation Embryo Developmental Arrest
3.3.1. The Role of Genetic Factors
3.3.2. The Role of Follicular Markers
3.3.3. The Role of Female Infertility Etiology
3.4. Paternal Factors Leading to Preimplantation Embryo Developmental Arrest
3.4.1. The Role of Genetic Factors
3.4.2. The Role of DNA Fragmentation and Chromatin Condensation
3.4.3. The Role of Male Infertility Etiologies
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hammond, E.R.; Cree, L.M.; Morbeck, D.E. Should Extended Blastocyst Culture Include Day 7? Hum. Reprod. 2018, 33, 991–997. [Google Scholar] [CrossRef] [Green Version]
- Gardner, D.K.; Balaban, B. Assessment of Human Embryo Development Using Morphological Criteria in an Era of Time-Lapse, Algorithms and “OMICS”: Is Looking Good Still Important? Mol. Hum. Reprod 2016, 22, 704–718. [Google Scholar] [CrossRef]
- Simopoulou, M.; Sfakianoudis, K.; Maziotis, E.; Antoniou, N.; Rapani, A.; Anifandis, G.; Bakas, P.; Bolaris, S.; Pantou, A.; Pantos, K.; et al. Are Computational Applications the “Crystal Ball” in the IVF Laboratory? The Evolution from Mathematics to Artificial Intelligence. J. Assist. Reprod Genet. 2018, 35, 1545–1557. [Google Scholar] [CrossRef]
- Rafferty, A.R.; Reina, R.D. Arrested Embryonic Development: A Review of Strategies to Delay Hatching in Egg-Laying Reptiles. Proc. Biol. Sci. 2012, 279, 2299–2308. [Google Scholar] [CrossRef] [Green Version]
- Alpha Scientists in Reproductive Medicine and ESHRE Special Interest Group of Embryology The Istanbul Consensus Workshop on Embryo Assessment: Proceedings of an Expert Meeting. Hum. Reprod. 2011, 26, 1270–1283. [CrossRef] [PubMed] [Green Version]
- Mohebi, M.; Ghafouri-Fard, S. Embryo Developmental Arrest: Review of Genetic Factors and Pathways. Gene. Rep. 2019, 17, 100479. [Google Scholar] [CrossRef]
- Greco, E.; Minasi, M.G.; Fiorentino, F. Healthy Babies after Intrauterine Transfer of Mosaic Aneuploid Blastocysts. N. Engl. J. Med. 2015, 373, 2089–2090. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- ESHRE Special Interest Group of Embryology; Alpha Scientists in Reproductive Medicine The Vienna Consensus: Report of an Expert Meeting on the Development of Art Laboratory Performance Indicators†‡. Hum. Reprod. Open 2017, 2017. [CrossRef] [Green Version]
- Betts, D.H.; Madan, P. Permanent Embryo Arrest: Molecular and Cellular Concepts. Mol. Hum. Reprod. 2008, 14, 445–453. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Qian, Y.; Liu, Y.; Wang, Q.; Wang, R.; Zhou, Y.; Zhang, C.; Pang, Z.; Ye, H.; Xue, S.; et al. A Novel Homozygous Variant in NLRP5 Is Associate with Human Early Embryonic Arrest in a Consanguineous Chinese Family. Clin. Genet. 2020, 98, 69–73. [Google Scholar] [CrossRef]
- Bolton, V.N.; Hawes, S.M.; Taylor, C.T.; Parsons, J.H. Development of Spare Human Preimplantation Embryos in Vitro: An Analysis of the Correlations among Gross Morphology, Cleavage Rates, and Development to the Blastocyst. J. Vitr. Fert. Embryo Transf. 1989, 6, 30–35. [Google Scholar] [CrossRef]
- Cívico, S.; Agell, N.; Hernández, L.; Campo, E.; Bachs, O.; Balasch, J. Increased Messenger Ribonucleic Acid Expression of the Cyclin-Dependent Kinase Inhibitor P27Kip1 in Cleavage-Stage Human Embryos Exhibiting Developmental Arrest. Fertil. Steril. 2008, 89, 1557–1562. [Google Scholar] [CrossRef] [PubMed]
- Garrido, N.; Bellver, J.; Remohí, J.; Simón, C.; Pellicer, A. Cumulative Live-Birth Rates per Total Number of Embryos Needed to Reach Newborn in Consecutive in Vitro Fertilization (IVF) Cycles: A New Approach to Measuring the Likelihood of IVF Success. Fertil. Steril. 2011, 96, 40–46. [Google Scholar] [CrossRef]
- Meseguer, M.; Herrero, J.; Tejera, A.; Hilligsøe, K.M.; Ramsing, N.B.; Remohí, J. The Use of Morphokinetics as a Predictor of Embryo Implantation. Hum. Reprod. 2011, 26, 2658–2671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, S.T.; Shi, J.X.; Gong, F.; Zhang, S.P.; Lu, C.F.; Tan, K.; Leng, L.Z.; Hao, M.; He, H.; Gu, Y.F.; et al. Cleavage Pattern Predicts Developmental Potential of Day 3 Human Embryos Produced by IVF. Reprod. Biomed. Online 2015, 30, 625–634. [Google Scholar] [CrossRef] [Green Version]
- McCollin, A.; Swann, R.L.; Summers, M.C.; Handyside, A.H.; Ottolini, C.S. Abnormal Cleavage and Developmental Arrest of Human Preimplantation Embryos in Vitro. Eur. J. Med. Genet. 2020, 63, 103651. [Google Scholar] [CrossRef] [PubMed]
- Daughtry, B.L.; Chavez, S.L. Chromosomal Instability in Mammalian Pre-Implantation Embryos: Potential Causes, Detection Methods, and Clinical Consequences. Cell Tissue Res. 2016, 363, 201–225. [Google Scholar] [CrossRef] [Green Version]
- Maurer, M.; Ebner, T.; Puchner, M.; Mayer, R.B.; Shebl, O.; Oppelt, P.; Duba, H.-C. Chromosomal Aneuploidies and Early Embryonic Developmental Arrest. Int. J. Fertil. Steril. 2015, 9, 346–353. [Google Scholar] [CrossRef] [PubMed]
- Ottolini, C.S.; Kitchen, J.; Xanthopoulou, L.; Gordon, T.; Summers, M.C.; Handyside, A.H. Tripolar Mitosis and Partitioning of the Genome Arrests Human Preimplantation Development in Vitro. Sci. Rep. 2017, 7, 9744. [Google Scholar] [CrossRef] [Green Version]
- McCoy, R.C.; Demko, Z.P.; Ryan, A.; Banjevic, M.; Hill, M.; Sigurjonsson, S.; Rabinowitz, M.; Petrov, D.A. Evidence of Selection against Complex Mitotic-Origin Aneuploidy during Preimplantation Development. PLoS Genet. 2015, 11, e1005601. [Google Scholar] [CrossRef] [Green Version]
- Mantzouratou, A.; Delhanty, J.D.A. Aneuploidy in the Human Cleavage Stage Embryo. Cytogenet. Genome. Res. 2011, 133, 141–148. [Google Scholar] [CrossRef]
- Tšuiko, O.; Jatsenko, T.; Parameswaran Grace, L.K.; Kurg, A.; Vermeesch, J.R.; Lanner, F.; Altmäe, S.; Salumets, A. A Speculative Outlook on Embryonic Aneuploidy: Can Molecular Pathways Be Involved? Dev. Biol. 2019, 447, 3–13. [Google Scholar] [CrossRef]
- Cui, X.; Jing, X.; Wu, X.; Yan, M.; Li, Q.; Shen, Y.; Wang, Z. DNA Methylation in Spermatogenesis and Male Infertility (Review). Exp. Ther. Med. 2016, 12, 1973–1979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agarwal, A.; Roychoudhury, S.; Bjugstad, K.B.; Cho, C.-L. Oxidation-Reduction Potential of Semen: What Is Its Role in the Treatment of Male Infertility? Ther. Adv. Urol. 2016, 8, 302–318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frattarelli, J.L.; Miller, K.A.; Miller, B.T.; Elkind-Hirsch, K.; Scott, R.T. Male Age Negatively Impacts Embryo Development and Reproductive Outcome in Donor Oocyte Assisted Reproductive Technology Cycles. Fertil. Steril. 2008, 90, 97–103. [Google Scholar] [CrossRef] [PubMed]
- Qi, S.-T.; Liang, L.-F.; Xian, Y.-X.; Liu, J.-Q.; Wang, W. Arrested Human Embryos Are More Likely to Have Abnormal Chromosomes than Developing Embryos from Women of Advanced Maternal Age. J. Ovarian Res. 2014, 7, 65. [Google Scholar] [CrossRef] [Green Version]
- Simon, L.; Murphy, K.; Shamsi, M.B.; Liu, L.; Emery, B.; Aston, K.I.; Hotaling, J.; Carrell, D.T. Paternal Influence of Sperm DNA Integrity on Early Embryonic Development. Hum. Reprod. 2014, 29, 2402–2412. [Google Scholar] [CrossRef]
- Sha, Q.-Q.; Zheng, W.; Wu, Y.-W.; Li, S.; Guo, L.; Zhang, S.; Lin, G.; Ou, X.-H.; Fan, H.-Y. Dynamics and Clinical Relevance of Maternal MRNA Clearance during the Oocyte-to-Embryo Transition in Humans. Nat. Commun. 2020, 11, 4917. [Google Scholar] [CrossRef]
- Pellicer, A.; Oliveira, N.; Ruiz, A.; Remohí, J.; Simón, C. Exploring the Mechanism(s) of Endometriosis-Related Infertility: An Analysis of Embryo Development and Implantation in Assisted Reproduction. Hum. Reprod. 1995, 10 (Suppl. 2), 91–97. [Google Scholar] [CrossRef] [PubMed]
- Coticchio, G.; Barrie, A.; Lagalla, C.; Borini, A.; Fishel, S.; Griffin, D.; Campbell, A. Plasticity of the Human Preimplantation Embryo: Developmental Dogmas, Variations on Themes and Self-Correction. Hum. Reprod. Update 2021. [Google Scholar] [CrossRef]
- Cimadomo, D.; Capalbo, A.; Scarica, C.; Sosa Fernandez, L.; Rienzi, L.; Ciriminna, R.; Minasi, M.G.; Novelli, A.; De Santis, L.; Zuccarello, D. When Embryology Meets Genetics: The Definition of Developmentally Incompetent Preimplantation Embryos (DIPE)-the Consensus of Two Italian Scientific Societies. J. Assist. Reprod. Genet. 2021, 38, 319–331. [Google Scholar] [CrossRef]
- Paonessa, M.; Borini, A.; Coticchio, G. Genetic Causes of Preimplantation Embryo Developmental Failure. Mol. Reprod. Dev. 2021, 88, 338–348. [Google Scholar] [CrossRef]
- Canosa, S.; Bergandi, L.; Macrì, C.; Charrier, L.; Paschero, C.; Carosso, A.; Di Segni, N.; Silvagno, F.; Gennarelli, G.; Benedetto, C.; et al. Morphokinetic Analysis of Cleavage Stage Embryos and Assessment of Specific Gene Expression in Cumulus Cells Independently Predict Human Embryo Development to Expanded Blastocyst: A Preliminary Study. J. Assist. Reprod. Genet. 2020, 37, 1409–1420. [Google Scholar] [CrossRef]
- Yanez, L.Z.; Han, J.; Behr, B.B.; Pera, R.A.R.; Camarillo, D.B. Human Oocyte Developmental Potential Is Predicted by Mechanical Properties within Hours after Fertilization. Nat. Commun. 2016, 7, 10809. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Li, R.-Q.; Ou, S.-B.; Zhang, N.-F.; Ren, L.; Wei, L.-N.; Zhang, Q.-X.; Yang, D.-Z. Increased GDF9 and BMP15 MRNA Levels in Cumulus Granulosa Cells Correlate with Oocyte Maturation, Fertilization, and Embryo Quality in Humans. Reprod. Biol. Endocrinol. 2014, 12, 81. [Google Scholar] [CrossRef] [Green Version]
- Persani, L.; Rossetti, R.; Di Pasquale, E.; Cacciatore, C.; Fabre, S. The Fundamental Role of Bone Morphogenetic Protein 15 in Ovarian Function and Its Involvement in Female Fertility Disorders. Hum. Reprod. Update 2014, 20, 869–883. [Google Scholar] [CrossRef] [Green Version]
- Chang, H.-M.; Qiao, J.; Leung, P.C.K. Oocyte-Somatic Cell Interactions in the Human Ovary-Novel Role of Bone Morphogenetic Proteins and Growth Differentiation Factors. Hum. Reprod. Update 2016, 23, 1–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanfins, A.; Rodrigues, P.; Albertini, D.F. GDF-9 and BMP-15 Direct the Follicle Symphony. J. Assist. Reprod. Genet. 2018, 35, 1741–1750. [Google Scholar] [CrossRef] [PubMed]
- Bayne, R.A.L.; Kinnell, H.L.; Coutts, S.M.; He, J.; Childs, A.J.; Anderson, R.A. GDF9 Is Transiently Expressed in Oocytes before Follicle Formation in the Human Fetal Ovary and Is Regulated by a Novel NOBOX Transcript. PLoS ONE 2015, 10, e0119819. [Google Scholar] [CrossRef] [PubMed]
- Ho, J.R.; Arrach, N.; Rhodes-Long, K.; Salem, W.; McGinnis, L.K.; Chung, K.; Bendikson, K.A.; Paulson, R.J.; Ahmady, A. Blastulation Timing Is Associated with Differential Mitochondrial Content in Euploid Embryos. J. Assist. Reprod. Genet. 2018, 35, 711–720. [Google Scholar] [CrossRef]
- Stigliani, S.; Persico, L.; Lagazio, C.; Anserini, P.; Venturini, P.L.; Scaruffi, P. Mitochondrial DNA in Day 3 Embryo Culture Medium Is a Novel, Non-Invasive Biomarker of Blastocyst Potential and Implantation Outcome. Mol. Hum. Reprod. 2014, 20, 1238–1246. [Google Scholar] [CrossRef]
- Stigliani, S.; Anserini, P.; Venturini, P.L.; Scaruffi, P. Mitochondrial DNA Content in Embryo Culture Medium Is Significantly Associated with Human Embryo Fragmentation. Hum. Reprod. 2013, 28, 2652–2660. [Google Scholar] [CrossRef] [Green Version]
- Van Blerkom, J.; Davis, P.W.; Lee, J. ATP Content of Human Oocytes and Developmental Potential and Outcome after In-Vitro Fertilization and Embryo Transfer. Hum. Reprod. 1995, 10, 415–424. [Google Scholar] [CrossRef]
- Cummins, J. Mitochondrial DNA in Mammalian Reproduction. Rev. Reprod. 1998, 3, 172–182. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, R.-H.; Au, H.-K.; Yeh, T.-S.; Chang, S.-J.; Cheng, Y.-F.; Tzeng, C.-R. Decreased Expression of Mitochondrial Genes in Human Unfertilized Oocytes and Arrested Embryos. Fertil. Steril. 2004, 81, 912–918. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, R.-H.; Tsai, N.-M.; Au, H.-K.; Chang, S.-J.; Wei, Y.-H.; Tzeng, C.-R. Multiple Rearrangements of Mitochondrial DNA in Unfertilized Human Oocytes. Fertil. Steril. 2002, 77, 1012–1017. [Google Scholar] [CrossRef]
- Bergandi, L.; Basso, G.; Evangelista, F.; Canosa, S.; Dalmasso, P.; Aldieri, E.; Revelli, A.; Benedetto, C.; Ghigo, D. Inducible Nitric Oxide Synthase and Heme Oxygenase 1 Are Expressed in Human Cumulus Cells and May Be Used as Biomarkers of Oocyte Competence. Reprod. Sci. 2014, 21, 1370–1377. [Google Scholar] [CrossRef]
- Viotti, M.; Victor, A.R.; Zouves, C.G.; Barnes, F.L. Is Mitochondrial DNA Quantitation in Blastocyst Trophectoderm Cells Predictive of Developmental Competence and Outcome in Clinical IVF? J. Assist. Reprod. Genet. 2017, 34, 1581–1585. [Google Scholar] [CrossRef] [PubMed]
- Treff, N.R.; Zhan, Y.; Tao, X.; Olcha, M.; Han, M.; Rajchel, J.; Morrison, L.; Morin, S.J.; Scott, R.T. Levels of Trophectoderm Mitochondrial DNA Do Not Predict the Reproductive Potential of Sibling Embryos. Hum. Reprod. 2017, 32, 954–962. [Google Scholar] [CrossRef] [Green Version]
- Victor, A.; Griffin, D.; Gardner, D.K.; Brake, A.; Zouves, C.; Barnes, F.; Viotti, M. Births from Embryos with Highly Elevated Levels of Mitochondrial DNA. Reprod. Biomed. Online 2019, 39, 403–412. [Google Scholar] [CrossRef]
- Victor, A.R.; Brake, A.J.; Tyndall, J.C.; Griffin, D.K.; Zouves, C.G.; Barnes, F.L.; Viotti, M. Accurate Quantitation of Mitochondrial DNA Reveals Uniform Levels in Human Blastocysts Irrespective of Ploidy, Age, or Implantation Potential. Fertil. Steril. 2017, 107, 34–42.e3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fragouli, E.; McCaffrey, C.; Ravichandran, K.; Spath, K.; Grifo, J.A.; Munné, S.; Wells, D. Clinical Implications of Mitochondrial DNA Quantification on Pregnancy Outcomes: A Blinded Prospective Non-Selection Study. Hum. Reprod. 2017, 32, 2340–2347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Surani, M.A. Imprinting and the Initiation of Gene Silencing in the Germ Line. Cell 1998, 93, 309–312. [Google Scholar] [CrossRef] [Green Version]
- Hajkova, P.; Erhardt, S.; Lane, N.; Haaf, T.; El-Maarri, O.; Reik, W.; Walter, J.; Surani, M.A. Epigenetic Reprogramming in Mouse Primordial Germ Cells. Mech. Dev. 2002, 117, 15–23. [Google Scholar] [CrossRef] [Green Version]
- Reik, W. Stability and Flexibility of Epigenetic Gene Regulation in Mammalian Development. Nature 2007, 447, 425–432. [Google Scholar] [CrossRef]
- Surani, M.A.; Hayashi, K.; Hajkova, P. Genetic and Epigenetic Regulators of Pluripotency. Cell 2007, 128, 747–762. [Google Scholar] [CrossRef] [Green Version]
- Zemel, S.; Bartolomei, M.S.; Tilghman, S.M. Physical Linkage of Two Mammalian Imprinted Genes, H19 and Insulin-like Growth Factor 2. Nat. Genet. 1992, 2, 61–65. [Google Scholar] [CrossRef] [PubMed]
- Ibala-Romdhane, S.; Al-Khtib, M.; Khoueiry, R.; Blachère, T.; Guérin, J.-F.; Lefèvre, A. Analysis of H19 Methylation in Control and Abnormal Human Embryos, Sperm and Oocytes. Eur. J. Hum. Genet. 2011, 19, 1138–1143. [Google Scholar] [CrossRef]
- Al-Khtib, M.; Blachère, T.; Guérin, J.F.; Lefèvre, A. Methylation Profile of the Promoters of Nanog and Oct4 in ICSI Human Embryos. Hum. Reprod. 2012, 27, 2948–2954. [Google Scholar] [CrossRef] [Green Version]
- Chambers, I.; Colby, D.; Robertson, M.; Nichols, J.; Lee, S.; Tweedie, S.; Smith, A. Functional Expression Cloning of Nanog, a Pluripotency Sustaining Factor in Embryonic Stem Cells. Cell 2003, 113, 643–655. [Google Scholar] [CrossRef] [Green Version]
- Mitsui, K.; Tokuzawa, Y.; Itoh, H.; Segawa, K.; Murakami, M.; Takahashi, K.; Maruyama, M.; Maeda, M.; Yamanaka, S. The Homeoprotein Nanog Is Required for Maintenance of Pluripotency in Mouse Epiblast and ES Cells. Cell 2003, 113, 631–642. [Google Scholar] [CrossRef] [Green Version]
- Nichols, J.; Zevnik, B.; Anastassiadis, K.; Niwa, H.; Klewe-Nebenius, D.; Chambers, I.; Schöler, H.; Smith, A. Formation of Pluripotent Stem Cells in the Mammalian Embryo Depends on the POU Transcription Factor Oct4. Cell 1998, 95, 379–391. [Google Scholar] [CrossRef] [Green Version]
- Niwa, H.; Miyazaki, J.; Smith, A.G. Quantitative Expression of Oct-3/4 Defines Differentiation, Dedifferentiation or Self-Renewal of ES Cells. Nat. Genet. 2000, 24, 372–376. [Google Scholar] [CrossRef]
- Timofeeva, A.; Drapkina, Y.; Fedorov, I.; Chagovets, V.; Makarova, N.; Shamina, M.; Kalinina, E.; Sukhikh, G. Small Noncoding RNA Signatures for Determining the Developmental Potential of an Embryo at the Morula Stage. Int. J. Mol. Sci. 2020, 21, 9399. [Google Scholar] [CrossRef]
- Sahin, C.; Mamillapalli, R.; Yi, K.W.; Taylor, H.S. MicroRNA Let-7b: A Novel Treatment for Endometriosis. J. Cell Mol. Med. 2018, 22, 5346–5353. [Google Scholar] [CrossRef]
- Rapani, A.; Nikiforaki, D.; Karagkouni, D.; Sfakianoudis, K.; Tsioulou, P.; Grigoriadis, S.; Maziotis, E.; Pantou, A.; Voutsina, A.; Pantou, A.; et al. Reporting on the Role of MiRNAs and Affected Pathways on the Molecular Backbone of Ovarian Insufficiency: A Systematic Review and Critical Analysis Mapping of Future Research. Front. Cell Dev. Biol. 2021, 8. [Google Scholar] [CrossRef]
- Santos, M.A.; Teklenburg, G.; Macklon, N.S.; Van Opstal, D.; Schuring-Blom, G.H.; Krijtenburg, P.-J.; de Vreeden-Elbertse, J.; Fauser, B.C.; Baart, E.B. The Fate of the Mosaic Embryo: Chromosomal Constitution and Development of Day 4, 5 and 8 Human Embryos. Hum. Reprod. 2010, 25, 1916–1926. [Google Scholar] [CrossRef] [Green Version]
- Braude, P.; Bolton, V.; Moore, S. Human Gene Expression First Occurs between the Four- and Eight-Cell Stages of Preimplantation Development. Nature 1988, 332, 459–461. [Google Scholar] [CrossRef]
- Tesarík, J.; Kopecný, V.; Plachot, M.; Mandelbaum, J. Activation of Nucleolar and Extranucleolar RNA Synthesis and Changes in the Ribosomal Content of Human Embryos Developing in Vitro. J. Reprod. Fertil. 1986, 78, 463–470. [Google Scholar] [CrossRef] [Green Version]
- Evsikov, S.; Verlinsky, Y. Mosaicism in the Inner Cell Mass of Human Blastocysts. Hum. Reprod. 1998, 13, 3151–3155. [Google Scholar] [CrossRef] [PubMed]
- Ruangvutilert, P.; Delhanty, J.D.; Serhal, P.; Simopoulou, M.; Rodeck, C.H.; Harper, J.C. FISH Analysis on Day 5 Post-Insemination of Human Arrested and Blastocyst Stage Embryos. Prenat. Diagn. 2000, 20, 552–560. [Google Scholar] [CrossRef]
- Munné, S.; Grifo, J.; Cohen, J.; Weier, H.U. Chromosome Abnormalities in Human Arrested Preimplantation Embryos: A Multiple-Probe FISH Study. Am. J. Hum. Genet. 1994, 55, 150–159. [Google Scholar] [PubMed]
- Shahbazi, M.N.; Wang, T.; Tao, X.; Weatherbee, B.A.T.; Sun, L.; Zhan, Y.; Keller, L.; Smith, G.D.; Pellicer, A.; Scott, R.T.; et al. Developmental Potential of Aneuploid Human Embryos Cultured beyond Implantation. Nat. Commun. 2020, 11, 3987. [Google Scholar] [CrossRef] [PubMed]
- Delimitreva, S.M.; Zhivkova, R.S.; Vatev, I.T.S.; Toncheva, D.I. Chromosomal Disorders and Nuclear and Cell Destruction in Cleaving Human Embryos. Int. J. Dev. Biol. 2005, 49, 409–416. [Google Scholar] [CrossRef] [PubMed]
- Kort, D.H.; Chia, G.; Treff, N.R.; Tanaka, A.J.; Xing, T.; Vensand, L.B.; Micucci, S.; Prosser, R.; Lobo, R.A.; Sauer, M.V.; et al. Human Embryos Commonly Form Abnormal Nuclei during Development: A Mechanism of DNA Damage, Embryonic Aneuploidy, and Developmental Arrest. Hum. Reprod. 2016, 31, 312–323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magli, M.C.; Gianaroli, L.; Ferraretti, A.P.; Lappi, M.; Ruberti, A.; Farfalli, V. Embryo Morphology and Development Are Dependent on the Chromosomal Complement. Fertil. Steril. 2007, 87, 534–541. [Google Scholar] [CrossRef]
- Munné, S.; Alikani, M.; Tomkin, G.; Grifo, J.; Cohen, J. Embryo Morphology, Developmental Rates, and Maternal Age Are Correlated with Chromosome Abnormalities. Fertil. Steril. 1995, 64, 382–391. [Google Scholar] [CrossRef]
- Magli, M.C.; Gianaroli, L.; Ferraretti, A.P. Chromosomal Abnormalities in Embryos. Mol. Cell Endocrinol. 2001, 183, S29–S34. [Google Scholar] [CrossRef]
- Márquez, C.; Sandalinas, M.; Bahçe, M.; Alikani, M.; Munné, S. Chromosome Abnormalities in 1255 Cleavage-Stage Human Embryos. Reprod. Biomed. Online 2000, 1, 17–26. [Google Scholar] [CrossRef]
- Alikani, M.; Calderon, G.; Tomkin, G.; Garrisi, J.; Kokot, M.; Cohen, J. Cleavage Anomalies in Early Human Embryos and Survival after Prolonged Culture In-Vitro. Hum. Reprod. 2000, 15, 2634–2643. [Google Scholar] [CrossRef]
- Edwards, R.G. In Vitro Fertilization and Embryo Replacement: Opening Lecture. Ann. N.Y. Acad. Sci. 1985, 442, 1–22. [Google Scholar] [CrossRef]
- Mehta, A.; Haber, J.E. Sources of DNA Double-Strand Breaks and Models of Recombinational DNA Repair. Cold Spring Harb. Perspect. Biol. 2014, 6, a016428. [Google Scholar] [CrossRef] [Green Version]
- Girardi, L.; Serdarogullari, M.; Patassini, C.; Poli, M.; Fabiani, M.; Caroselli, S.; Coban, O.; Findikli, N.; Boynukalin, F.K.; Bahceci, M.; et al. Incidence, Origin, and Predictive Model for the Detection and Clinical Management of Segmental Aneuploidies in Human Embryos. Am. J. Hum. Genet. 2020, 106, 525–534. [Google Scholar] [CrossRef] [PubMed]
- Babariya, D.; Fragouli, E.; Alfarawati, S.; Spath, K.; Wells, D. The Incidence and Origin of Segmental Aneuploidy in Human Oocytes and Preimplantation Embryos. Hum. Reprod. 2017, 32, 2549–2560. [Google Scholar] [CrossRef]
- Capalbo, A.; Ubaldi, F.M.; Rienzi, L.; Scott, R.; Treff, N. Detecting Mosaicism in Trophectoderm Biopsies: Current Challenges and Future Possibilities. Hum. Reprod. 2017, 32, 492–498. [Google Scholar] [CrossRef]
- Mamas, T.; Gordon, A.; Brown, A.; Harper, J.; SenGupta, S. Detection of Aneuploidy by Array Comparative Genomic Hybridization Using Cell Lines to Mimic a Mosaic Trophectoderm Biopsy. Fertil. Steril. 2012, 97, 943–947. [Google Scholar] [CrossRef] [PubMed]
- Capalbo, A.; Wright, G.; Elliott, T.; Ubaldi, F.M.; Rienzi, L.; Nagy, Z.P. FISH Reanalysis of Inner Cell Mass and Trophectoderm Samples of Previously Array-CGH Screened Blastocysts Shows High Accuracy of Diagnosis and No Major Diagnostic Impact of Mosaicism at the Blastocyst Stage. Hum. Reprod. 2013, 28, 2298–2307. [Google Scholar] [CrossRef] [Green Version]
- Capalbo, A.; Treff, N.R.; Cimadomo, D.; Tao, X.; Upham, K.; Ubaldi, F.M.; Rienzi, L.; Scott, R.T. Comparison of Array Comparative Genomic Hybridization and Quantitative Real-Time PCR-Based Aneuploidy Screening of Blastocyst Biopsies. Eur. J. Hum. Genet. 2015, 23, 901–906. [Google Scholar] [CrossRef] [PubMed]
- Biricik, A.; Cotroneo, E.; Minasi, M.G.; Greco, P.F.; Bono, S.; Surdo, M.; Lecciso, F.; Sessa, M.; Fiorentino, F.; Spinella, F.; et al. Cross-Validation of Next-Generation Sequencing Technologies for Diagnosis of Chromosomal Mosaicism and Segmental Aneuploidies in Preimplantation Embryos Model. Life (Basel) 2021, 11, 340. [Google Scholar] [CrossRef] [PubMed]
- Goodrich, D.; Xing, T.; Tao, X.; Lonczak, A.; Zhan, Y.; Landis, J.; Zimmerman, R.; Scott, R.T.; Treff, N.R. Evaluation of Comprehensive Chromosome Screening Platforms for the Detection of Mosaic Segmental Aneuploidy. J. Assist. Reprod. Genet. 2017, 34, 975–981. [Google Scholar] [CrossRef] [Green Version]
- Gleicher, N.; Vidali, A.; Braverman, J.; Kushnir, V.A.; Barad, D.H.; Hudson, C.; Wu, Y.-G.; Wang, Q.; Zhang, L.; Albertini, D.F.; et al. Accuracy of Preimplantation Genetic Screening (PGS) Is Compromised by Degree of Mosaicism of Human Embryos. Reprod. Biol. Endocrinol. 2016, 14, 54. [Google Scholar] [CrossRef] [Green Version]
- Treff, N.R.; Franasiak, J.M. Detection of Segmental Aneuploidy and Mosaicism in the Human Preimplantation Embryo: Technical Considerations and Limitations. Fertil. Steril. 2017, 107, 27–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Popovic, M.; Dhaenens, L.; Taelman, J.; Dheedene, A.; Bialecka, M.; De Sutter, P.; Chuva de Sousa Lopes, S.M.; Menten, B.; Heindryckx, B. Extended in Vitro Culture of Human Embryos Demonstrates the Complex Nature of Diagnosing Chromosomal Mosaicism from a Single Trophectoderm Biopsy. Hum. Reprod. 2019, 34, 758–769. [Google Scholar] [CrossRef]
- Popovic, M.; Dheedene, A.; Christodoulou, C.; Taelman, J.; Dhaenens, L.; Van Nieuwerburgh, F.; Deforce, D.; Van den Abbeel, E.; De Sutter, P.; Menten, B.; et al. Chromosomal Mosaicism in Human Blastocysts: The Ultimate Challenge of Preimplantation Genetic Testing? Hum. Reprod. 2018, 33, 1342–1354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, L.; Jin, L.; Chen, W.; Liu, J.M.; Hu, J.; Yu, Q.; Ren, X.L.; Huang, B.; He, H. The True Incidence of Chromosomal Mosaicism after Preimplantation Genetic Testing Is Much Lower than That Indicated by Trophectoderm Biopsy. Hum. Reprod. 2021, 36, 1691–1701. [Google Scholar] [CrossRef] [PubMed]
- Goodrich, D.; Tao, X.; Bohrer, C.; Lonczak, A.; Xing, T.; Zimmerman, R.; Zhan, Y.; Scott Jr, R.T.; Treff, N.R. A Randomized and Blinded Comparison of QPCR and NGS-Based Detection of Aneuploidy in a Cell Line Mixture Model of Blastocyst Biopsy Mosaicism. J. Assist. Reprod. Genet. 2016, 33, 1473–1480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruttanajit, T.; Chanchamroen, S.; Cram, D.S.; Sawakwongpra, K.; Suksalak, W.; Leng, X.; Fan, J.; Wang, L.; Yao, Y.; Quangkananurug, W. Detection and Quantitation of Chromosomal Mosaicism in Human Blastocysts Using Copy Number Variation Sequencing. Prenat. Diagn. 2016, 36, 154–162. [Google Scholar] [CrossRef]
- Popovic, M.; Dhaenens, L.; Boel, A.; Menten, B.; Heindryckx, B. Chromosomal Mosaicism in Human Blastocysts: The Ultimate Diagnostic Dilemma. Hum. Reprod. Update 2020, 26, 313–334. [Google Scholar] [CrossRef]
- Besser, A.G.; Mounts, E.L. Counselling Considerations for Chromosomal Mosaicism Detected by Preimplantation Genetic Screening. Reprod. Biomed. Online 2017, 34, 369–374. [Google Scholar] [CrossRef] [Green Version]
- Orvieto, R.; Shimon, C.; Rienstein, S.; Jonish-Grossman, A.; Shani, H.; Aizer, A. Do Human Embryos Have the Ability of Self-Correction? Reprod. Biol. Endocrinol. 2020, 18, 98. [Google Scholar] [CrossRef]
- Doseff, A.I. Apoptosis: The Sculptor of Development. Stem. Cells Dev. 2004, 13, 473–483. [Google Scholar] [CrossRef]
- Liu, X.; Zou, H.; Slaughter, C.; Wang, X. DFF, a Heterodimeric Protein That Functions Downstream of Caspase-3 to Trigger DNA Fragmentation during Apoptosis. Cell 1997, 89, 175–184. [Google Scholar] [CrossRef] [Green Version]
- Kaihola, H.; Yaldir, F.G.; Bohlin, T.; Samir, R.; Hreinsson, J.; Åkerud, H. Levels of Caspase-3 and Histidine-Rich Glycoprotein in the Embryo Secretome as Biomarkers of Good-Quality Day-2 Embryos and High-Quality Blastocysts. PLoS ONE 2019, 14, e0226419. [Google Scholar] [CrossRef] [Green Version]
- Lindgren, K.E.; Gülen Yaldir, F.; Hreinsson, J.; Holte, J.; Kårehed, K.; Sundström-Poromaa, I.; Kaihola, H.; Åkerud, H. Differences in Secretome in Culture Media When Comparing Blastocysts and Arrested Embryos Using Multiplex Proximity Assay. Ups. J. Med. Sci. 2018, 123, 143–152. [Google Scholar] [CrossRef] [Green Version]
- Ubeda, M.; Habener, J.F. The Large Subunit of the DNA Replication Complex C (DSEB/RF-C140) Cleaved and Inactivated by Caspase-3 (CPP32/YAMA) during Fas-Induced Apoptosis. J. Biol. Chem. 1997, 272, 19562–19568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samejima, K.; Villa, P.; Earnshaw, W.C. Role of Factors Downstream of Caspases in Nuclear Disassembly during Apoptotic Execution. Philos. Trans. R Soc. Lond. B Biol. Sci. 1999, 354, 1591–1598; discussion 1598–1599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seshagiri, P.B.; Vani, V.; Madhulika, P. Cytokines and Blastocyst Hatching. Am. J. Reprod. Immunol. 2016, 75, 208–217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jurisicova, A.; Antenos, M.; Varmuza, S.; Tilly, J.L.; Casper, R.F. Expression of Apoptosis-Related Genes during Human Preimplantation Embryo Development: Potential Roles for the Harakiri Gene Product and Caspase-3 in Blastomere Fragmentation. Mol. Hum. Reprod. 2003, 9, 133–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leung, L.L. Interaction of Histidine-Rich Glycoprotein with Fibrinogen and Fibrin. J. Clin. Invest. 1986, 77, 1305–1311. [Google Scholar] [CrossRef]
- Simantov, R.; Febbraio, M.; Crombie, R.; Asch, A.S.; Nachman, R.L.; Silverstein, R.L. Histidine-Rich Glycoprotein Inhibits the Antiangiogenic Effect of Thrombospondin-1. J. Clin. Invest. 2001, 107, 45–52. [Google Scholar] [CrossRef] [Green Version]
- Armstrong, L.C.; Bornstein, P. Thrombospondins 1 and 2 Function as Inhibitors of Angiogenesis. Matrix Biol. 2003, 22, 63–71. [Google Scholar] [CrossRef]
- Mains, L.M.; Christenson, L.; Yang, B.; Sparks, A.E.T.; Mathur, S.; Van Voorhis, B.J. Identification of Apolipoprotein A1 in the Human Embryonic Secretome. Fertil. Steril. 2011, 96, 422–427.e2. [Google Scholar] [CrossRef]
- Santos-Gallego, C.G.; Ibanez, B.; Badimon, J.J. HDL-Cholesterol: Is It Really Good? Differences between ApoA-I and HDL. Biochem. Pharm. 2008, 76, 443–452. [Google Scholar] [CrossRef]
- Browne, R.W.; Bloom, M.S.; Shelly, W.B.; Ocque, A.J.; Huddleston, H.G.; Fujimoto, V.Y. Follicular Fluid High Density Lipoprotein-Associated Micronutrient Levels Are Associated with Embryo Fragmentation during IVF. J. Assist. Reprod. Genet. 2009, 26, 557–560. [Google Scholar] [CrossRef] [Green Version]
- Telford, N.A.; Watson, A.J.; Schultz, G.A. Transition from Maternal to Embryonic Control in Early Mammalian Development: A Comparison of Several Species. Mol. Reprod. Dev. 1990, 26, 90–100. [Google Scholar] [CrossRef] [PubMed]
- Gott, A.L.; Hardy, K.; Winston, R.M.; Leese, H.J. Non-Invasive Measurement of Pyruvate and Glucose Uptake and Lactate Production by Single Human Preimplantation Embryos. Hum. Reprod. 1990, 5, 104–108. [Google Scholar] [CrossRef] [PubMed]
- Ji, H.; Shi, X.; Wang, J.; Cao, S.; Ling, X.; Zhang, J.; Shen, R.; Zhao, C. Peptidomic Analysis of Blastocyst Culture Medium and the Effect of Peptide Derived from Blastocyst Culture Medium on Blastocyst Formation and Viability. Mol. Reprod. Dev. 2020, 87, 191–201. [Google Scholar] [CrossRef] [PubMed]
- Absalón-Medina, V.A.; Butler, W.R.; Gilbert, R.O. Preimplantation Embryo Metabolism and Culture Systems: Experience from Domestic Animals and Clinical Implications. J. Assist. Reprod. Genet. 2014, 31, 393–409. [Google Scholar] [CrossRef] [Green Version]
- Tasaki, T.; Kim, S.T.; Zakrzewska, A.; Lee, B.E.; Kang, M.J.; Yoo, Y.D.; Cha-Molstad, H.J.; Hwang, J.; Soung, N.K.; Sung, K.S.; et al. UBR Box N-Recognin-4 (UBR4), an N-Recognin of the N-End Rule Pathway, and Its Role in Yolk Sac Vascular Development and Autophagy. Proc. Natl. Acad. Sci. USA 2013, 110, 3800–3805. [Google Scholar] [CrossRef] [Green Version]
- Cubillos-Rojas, M.; Amair-Pinedo, F.; Peiró-Jordán, R.; Bartrons, R.; Ventura, F.; Rosa, J.L. The E3 Ubiquitin Protein Ligase HERC2 Modulates the Activity of Tumor Protein P53 by Regulating Its Oligomerization. J. Biol. Chem. 2014, 289, 14782–14795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bekker-Jensen, S.; Rendtlew Danielsen, J.; Fugger, K.; Gromova, I.; Nerstedt, A.; Lukas, C.; Bartek, J.; Lukas, J.; Mailand, N. HERC2 Coordinates Ubiquitin-Dependent Assembly of DNA Repair Factors on Damaged Chromosomes. Nat. Cell Biol. 2010, 12, 80–86. [Google Scholar] [CrossRef]
- Morice-Picard, F.; Benard, G.; Rezvani, H.R.; Lasseaux, E.; Simon, D.; Moutton, S.; Rooryck, C.; Lacombe, D.; Baumann, C.; Arveiler, B. Complete Loss of Function of the Ubiquitin Ligase HERC2 Causes a Severe Neurodevelopmental Phenotype. Eur. J. Hum. Genet. 2016, 25, 52–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muramatsu, T. Basigin (CD147), a Multifunctional Transmembrane Glycoprotein with Various Binding Partners. J. Biochem. 2016, 159, 481–490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sherr, C.J. G1 Phase Progression: Cycling on Cue. Cell 1994, 79, 551–555. [Google Scholar] [CrossRef]
- Sherr, C.J.; Roberts, J.M. Inhibitors of Mammalian G1 Cyclin-Dependent Kinases. Genes Dev. 1995, 9, 1149–1163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pagano, M.; Tam, S.W.; Theodoras, A.M.; Beer-Romero, P.; Del Sal, G.; Chau, V.; Yew, P.R.; Draetta, G.F.; Rolfe, M. Role of the Ubiquitin-Proteasome Pathway in Regulating Abundance of the Cyclin-Dependent Kinase Inhibitor P27. Science 1995, 269, 682–685. [Google Scholar] [CrossRef]
- Hara, T.; Kamura, T.; Nakayama, K.; Oshikawa, K.; Hatakeyama, S.; Nakayama, K. Degradation of P27(Kip1) at the G(0)-G(1) Transition Mediated by a Skp2-Independent Ubiquitination Pathway. J. Biol. Chem. 2001, 276, 48937–48943. [Google Scholar] [CrossRef] [Green Version]
- Civico, S.; Agell, N.; Bachs, O.; Vanrell, J.A.; Balasch, J. Increased Expression of the Cyclin-Dependent Kinase Inhibitor P27 in Cleavage-Stage Human Embryos Exhibiting Developmental Arrest. Mol. Hum. Reprod. 2002, 8, 919–922. [Google Scholar] [CrossRef] [Green Version]
- Houghton, F.D.; Hawkhead, J.A.; Humpherson, P.G.; Hogg, J.E.; Balen, A.H.; Rutherford, A.J.; Leese, H.J. Non-Invasive Amino Acid Turnover Predicts Human Embryo Developmental Capacity. Hum. Reprod. 2002, 17, 999–1005. [Google Scholar] [CrossRef] [Green Version]
- Hammer, M.A.; Kolajova, M.; Léveillé, M.; Claman, P.; Baltz, J.M. Glycine Transport by Single Human and Mouse Embryos. Hum. Reprod. 2000, 15, 419–426. [Google Scholar] [CrossRef] [Green Version]
- Anthony, J.C.; Yoshizawa, F.; Anthony, T.G.; Vary, T.C.; Jefferson, L.S.; Kimball, S.R. Leucine Stimulates Translation Initiation in Skeletal Muscle of Postabsorptive Rats via a Rapamycin-Sensitive Pathway. J. Nutr. 2000, 130, 2413–2419. [Google Scholar] [CrossRef] [PubMed]
- Lynch, C.J.; Fox, H.L.; Vary, T.C.; Jefferson, L.S.; Kimball, S.R. Regulation of Amino Acid-Sensitive TOR Signaling by Leucine Analogues in Adipocytes. J. Cell Biochem. 2000, 77, 234–251. [Google Scholar] [CrossRef]
- Kimball, S.R.; Shantz, L.M.; Horetsky, R.L.; Jefferson, L.S. Leucine Regulates Translation of Specific MRNAs in L6 Myoblasts through MTOR-Mediated Changes in Availability of EIF4E and Phosphorylation of Ribosomal Protein S6. J. Biol. Chem. 1999, 274, 11647–11652. [Google Scholar] [CrossRef] [Green Version]
- Conaghan, J.; Hardy, K.; Handyside, A.H.; Winston, R.M.; Leese, H.J. Selection Criteria for Human Embryo Transfer: A Comparison of Pyruvate Uptake and Morphology. J. Assist. Reprod. Genet. 1993, 10, 21–30. [Google Scholar] [CrossRef] [PubMed]
- Baumann, C.G.; Morris, D.G.; Sreenan, J.M.; Leese, H.J. The Quiet Embryo Hypothesis: Molecular Characteristics Favoring Viability. Mol. Reprod. Dev. 2007, 74, 1345–1353. [Google Scholar] [CrossRef] [PubMed]
- Leese, H.J. Quiet Please, Do Not Disturb: A Hypothesis of Embryo Metabolism and Viability. Bioessays 2002, 24, 845–849. [Google Scholar] [CrossRef] [PubMed]
- Leese, H.J.; Sturmey, R.G.; Baumann, C.G.; McEvoy, T.G. Embryo Viability and Metabolism: Obeying the Quiet Rules. Hum. Reprod. 2007, 22, 3047–3050. [Google Scholar] [CrossRef] [PubMed]
- Sturmey, R.G.; Hawkhead, J.A.; Barker, E.A.; Leese, H.J. DNA Damage and Metabolic Activity in the Preimplantation Embryo. Hum. Reprod. 2009, 24, 81–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siristatidis, C.S.; Sertedaki, E.; Vaidakis, D.; Varounis, C.; Trivella, M. Metabolomics for Improving Pregnancy Outcomes in Women Undergoing Assisted Reproductive Technologies. Cochrane Database Syst. Rev. 2018. [Google Scholar] [CrossRef]
- Sanders, E.J.; Wride, M.A. Ultrastructural Identification of Apoptotic Nuclei Using the TUNEL Technique. Histochem. J. 1996, 28, 275–281. [Google Scholar] [CrossRef]
- Koopman, G.; Reutelingsperger, C.P.; Kuijten, G.A.; Keehnen, R.M.; Pals, S.T.; van Oers, M.H. Annexin V for Flow Cytometric Detection of Phosphatidylserine Expression on B Cells Undergoing Apoptosis. Blood 1994, 84, 1415–1420. [Google Scholar] [CrossRef] [Green Version]
- Homburg, C.H.; de Haas, M.; von dem Borne, A.E.; Verhoeven, A.J.; Reutelingsperger, C.P.; Roos, D. Human Neutrophils Lose Their Surface Fc Gamma RIII and Acquire Annexin V Binding Sites during Apoptosis in Vitro. Blood 1995, 85, 532–540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levy, R.; Benchaib, M.; Cordonier, H.; Souchier, C.; Guerin, J.F. Annexin V Labelling and Terminal Transferase-Mediated DNA End Labelling (TUNEL) Assay in Human Arrested Embryos. Mol. Hum. Reprod. 1998, 4, 775–783. [Google Scholar] [CrossRef] [PubMed]
- Görlich, D.; Mattaj, I.W. Nucleocytoplasmic Transport. Science 1996, 271, 1513–1518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benavente, R.; Dabauvalle, M.C.; Scheer, U.; Chaly, N. Functional Role of Newly Formed Pore Complexes in Postmitotic Nuclear Reorganization. Chromosoma 1989, 98, 233–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panté, N.; Aebi, U. The Nuclear Pore Complex. J. Cell Biol. 1993, 122, 977–984. [Google Scholar] [CrossRef] [Green Version]
- Raghunayakula, S.; Subramonian, D.; Dasso, M.; Kumar, R.; Zhang, X.-D. Molecular Characterization and Functional Analysis of Annulate Lamellae Pore Complexes in Nuclear Transport in Mammalian Cells. PLoS ONE 2015, 10, e0144508. [Google Scholar] [CrossRef] [Green Version]
- Rawe, V.Y.; Olmedo, S.B.; Nodar, F.N.; Ponzio, R.; Sutovsky, P. Abnormal Assembly of Annulate Lamellae and Nuclear Pore Complexes Coincides with Fertilization Arrest at the Pronuclear Stage of Human Zygotic Development. Hum. Reprod. 2003, 18, 576–582. [Google Scholar] [CrossRef] [Green Version]
- Yamanaka, M.; Hashimoto, S.; Amo, A.; Ito-Sasaki, T.; Abe, H.; Morimoto, Y. Developmental Assessment of Human Vitrified-Warmed Blastocysts Based on Oxygen Consumption. Hum. Reprod. 2011, 26, 3366–3371. [Google Scholar] [CrossRef] [Green Version]
- Watson, A.J. The Cell Biology of Blastocyst Development. Mol. Reprod. Dev. 1992, 33, 492–504. [Google Scholar] [CrossRef]
- Kurosawa, H.; Utsunomiya, H.; Shiga, N.; Takahashi, A.; Ihara, M.; Ishibashi, M.; Nishimoto, M.; Watanabe, Z.; Abe, H.; Kumagai, J.; et al. Development of a New Clinically Applicable Device for Embryo Evaluation Which Measures Embryo Oxygen Consumption. Hum. Reprod. 2016, 31, 2321–2330. [Google Scholar] [CrossRef]
- Thompson, J.G.; Partridge, R.J.; Houghton, F.D.; Cox, C.I.; Leese, H.J. Oxygen Uptake and Carbohydrate Metabolism by in Vitro Derived Bovine Embryos. J. Reprod. Fertil. 1996, 106, 299–306. [Google Scholar] [CrossRef] [Green Version]
- Trimarchi, J.R.; Liu, L.; Porterfield, D.M.; Smith, P.J.; Keefe, D.L. Oxidative Phosphorylation-Dependent and -Independent Oxygen Consumption by Individual Preimplantation Mouse Embryos. Biol. Reprod. 2000, 62, 1866–1874. [Google Scholar] [CrossRef]
- Harvey, A.J.; Kind, K.L.; Thompson, J.G. REDOX Regulation of Early Embryo Development. Reproduction 2002, 123, 479–486. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, A.; Said, T.M.; Bedaiwy, M.A.; Banerjee, J.; Alvarez, J.G. Oxidative Stress in an Assisted Reproductive Techniques Setting. Fertil. Steril. 2006, 86, 503–512. [Google Scholar] [CrossRef] [PubMed]
- Guérin, P.; El Mouatassim, S.; Ménézo, Y. Oxidative Stress and Protection against Reactive Oxygen Species in the Pre-Implantation Embryo and Its Surroundings. Hum. Reprod. Update 2001, 7, 175–189. [Google Scholar] [CrossRef]
- Lan, K.-C.; Lin, Y.-C.; Chang, Y.-C.; Lin, H.-J.; Tsai, Y.-R.; Kang, H.-Y. Limited Relationships between Reactive Oxygen Species Levels in Culture Media and Zygote and Embryo Development. J. Assist. Reprod. Genet. 2019, 36, 325–334. [Google Scholar] [CrossRef] [Green Version]
- Bedaiwy, M.A.; Falcone, T.; Mohamed, M.S.; Aleem, A.A.N.; Sharma, R.K.; Worley, S.E.; Thornton, J.; Agarwal, A. Differential Growth of Human Embryos in Vitro: Role of Reactive Oxygen Species. Fertil. Steril. 2004, 82, 593–600. [Google Scholar] [CrossRef]
- Gardner, D.K.; Wale, P.L.; Collins, R.; Lane, M. Glucose Consumption of Single Post-Compaction Human Embryos Is Predictive of Embryo Sex and Live Birth Outcome. Hum. Reprod. 2011, 26, 1981–1986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gardner, D.K.; Wale, P.L. Analysis of Metabolism to Select Viable Human Embryos for Transfer. Fertil. Steril. 2013, 99, 1062–1072. [Google Scholar] [CrossRef]
- Desch, L.; Bruno, C.; Luu, M.; Barberet, J.; Choux, C.; Lamotte, M.; Schmutz, E.; Sagot, P.; Fauque, P. Embryo Multinucleation at the Two-Cell Stage Is an Independent Predictor of Intracytoplasmic Sperm Injection Outcomes. Fertil. Steril. 2017, 107, 97–103.e4. [Google Scholar] [CrossRef] [Green Version]
- Wallbutton, S.; Kasraie, J. Vacuolated Oocytes: Fertilization and Embryonic Arrest Following Intra-Cytoplasmic Sperm Injection in a Patient Exhibiting Persistent Oocyte Macro Vacuolization—Case Report. J. Assist. Reprod. Genet. 2010, 27, 183–188. [Google Scholar] [CrossRef] [Green Version]
- Van Blerkom, J. Occurrence and Developmental Consequences of Aberrant Cellular Organization in Meiotically Mature Human Oocytes after Exogenous Ovarian Hyperstimulation. J. Electron. Microsc. Tech. 1990, 16, 324–346. [Google Scholar] [CrossRef]
- Veeck, L.L.; Clark, R.N. El Shafie, M.; Sousa, M.; Windt, M.-L.; Kruger, T.F. An Atlas of the Ultrastructure of Human Oocytes: A Guide for Assisted Reproduction, First Edition. New York: The Parthenon Publishing Group, 2000. Fertil. Steril. 2001, 75, 838–839. [Google Scholar] [CrossRef]
- Ebner, T.; Moser, M.; Sommergruber, M.; Gaiswinkler, U.; Shebl, O.; Jesacher, K.; Tews, G. Occurrence and Developmental Consequences of Vacuoles throughout Preimplantation Development. Fertil. Steril. 2005, 83, 1635–1640. [Google Scholar] [CrossRef] [PubMed]
- Landry, D.W.; Zucker, H.A.; Sauer, M.V.; Reznik, M.; Wiebe, L. Hypocellularity and Absence of Compaction as Criteria for Embryonic Death. Regen. Med. 2006, 1, 367–371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kong, X.; Yang, S.; Gong, F.; Lu, C.; Zhang, S.; Lu, G.; Lin, G. The Relationship between Cell Number, Division Behavior and Developmental Potential of Cleavage Stage Human Embryos: A Time-Lapse Study. PLoS ONE 2016, 11. [Google Scholar] [CrossRef] [Green Version]
- Schulman, A. The Search for Alternative Sources of Human Pluripotent Stem Cells. Stem. Cell Rev. 2005, 1, 291–292. [Google Scholar] [CrossRef]
- Alghamdi, M.; Alasmari, D.; Assiri, A.; Mattar, E.; Aljaddawi, A.A.; Alattas, S.G.; Redwan, E.M. An Overview of the Intrinsic Role of Citrullination in Autoimmune Disorders. J. Immunol. Res. 2019, 2019. [Google Scholar] [CrossRef] [Green Version]
- Kan, R.; Jin, M.; Subramanian, V.; Causey, C.P.; Thompson, P.R.; Coonrod, S.A. Potential Role for PADI-Mediated Histone Citrullination in Preimplantation Development. BMC Dev. Biol. 2012, 12, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Liu, X.; Zhang, M.; Li, T.; Muth, A.; Thompson, P.R.; Coonrod, S.A.; Zhang, X. Peptidylarginine Deiminase 1-Catalyzed Histone Citrullination Is Essential for Early Embryo Development. Sci. Rep. 2016, 6, 38727. [Google Scholar] [CrossRef] [Green Version]
- Yurttas, P.; Vitale, A.M.; Fitzhenry, R.J.; Cohen-Gould, L.; Wu, W.; Gossen, J.A.; Coonrod, S.A. Role for PADI6 and the Cytoplasmic Lattices in Ribosomal Storage in Oocytes and Translational Control in the Early Mouse Embryo. Development 2008, 135, 2627–2636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Y.; Shi, Y.; Fu, J.; Yu, M.; Feng, R.; Sang, Q.; Liang, B.; Chen, B.; Qu, R.; Li, B.; et al. Mutations in PADI6 Cause Female Infertility Characterized by Early Embryonic Arrest. Am. J. Hum. Genet. 2016, 99, 744–752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qian, J.; Nguyen, N.M.P.; Rezaei, M.; Huang, B.; Tao, Y.; Zhang, X.; Cheng, Q.; Yang, H.; Asangla, A.; Majewski, J.; et al. Biallelic PADI6 Variants Linking Infertility, Miscarriages, and Hydatidiform Moles. Eur. J. Hum. Genet. 2018, 26, 1007–1013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dean, J. Exacting Requirements for Development of the Egg. Available online: https://www.nejm.org/doi/pdf/10.1056/NEJMe1515512 (accessed on 17 March 2021).
- Feng, R.; Sang, Q.; Kuang, Y.; Sun, X.; Yan, Z.; Zhang, S.; Shi, J.; Tian, G.; Luchniak, A.; Fukuda, Y.; et al. Mutations in TUBB8 and Human Oocyte Meiotic Arrest. N. Engl. J. Med. 2016, 374, 223–232. [Google Scholar] [CrossRef]
- Chen, B.; Wang, W.; Peng, X.; Jiang, H.; Zhang, S.; Li, D.; Li, B.; Fu, J.; Kuang, Y.; Sun, X.; et al. The Comprehensive Mutational and Phenotypic Spectrum of TUBB8 in Female Infertility. Eur. J. Hum. Genet. 2019, 27, 300–307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, P.; Zheng, L.; Liang, H.; Li, Y.; Zhao, H.; Li, R.; Lai, L.; Zhang, Q.; Wang, W. A Novel Mutation in the TUBB8 Gene Is Associated with Complete Cleavage Failure in Fertilized Eggs. J. Assist. Reprod. Genet. 2018, 35, 1349–1356. [Google Scholar] [CrossRef]
- Li, L.; Baibakov, B.; Dean, J. A Subcortical Maternal Complex Essential for Preimplantation Mouse Embryogenesis. Dev. Cell 2008, 15, 416–425. [Google Scholar] [CrossRef] [Green Version]
- Bebbere, D.; Masala, L.; Albertini, D.F.; Ledda, S. The Subcortical Maternal Complex: Multiple Functions for One Biological Structure? J. Assist. Reprod. Genet. 2016, 33, 1431–1438. [Google Scholar] [CrossRef] [Green Version]
- Zhu, K.; Yan, L.; Zhang, X.; Lu, X.; Wang, T.; Yan, J.; Liu, X.; Qiao, J.; Li, L. Identification of a Human Subcortical Maternal Complex. Mol. Hum. Reprod. 2015, 21, 320–329. [Google Scholar] [CrossRef] [Green Version]
- Tashiro, F.; Kanai-Azuma, M.; Miyazaki, S.; Kato, M.; Tanaka, T.; Toyoda, S.; Yamato, E.; Kawakami, H.; Miyazaki, T.; Miyazaki, J.-I. Maternal-Effect Gene Ces5/OOEP/Moep19/Floped Is Essential for Oocyte Cytoplasmic Lattice Formation and Embryonic Development at the Maternal-Zygotic Stage Transition. Genes. Cells 2010, 15, 813–828. [Google Scholar] [CrossRef] [PubMed]
- Tong, Z.B.; Gold, L.; Pfeifer, K.E.; Dorward, H.; Lee, E.; Bondy, C.A.; Dean, J.; Nelson, L.M. Mater, a Maternal Effect Gene Required for Early Embryonic Development in Mice. Nat. Genet. 2000, 26, 267–268. [Google Scholar] [CrossRef] [PubMed]
- Dean, J. Oocyte-Specific Genes Regulate Follicle Formation, Fertility and Early Mouse Development. J. Reprod. Immunol. 2002, 53, 171–180. [Google Scholar] [CrossRef]
- Zheng, P.; Dean, J. Role of Filia, a Maternal Effect Gene, in Maintaining Euploidy during Cleavage-Stage Mouse Embryogenesis. Proc. Natl. Acad. Sci. USA 2009, 106, 7473–7478. [Google Scholar] [CrossRef] [Green Version]
- Niakan, K.K.; Eggan, K. Analysis of Human Embryos from Zygote to Blastocyst Reveals Distinct Gene Expression Patterns Relative to the Mouse. Dev. Biol. 2013, 375, 54–64. [Google Scholar] [CrossRef] [Green Version]
- Ting, J.P.-Y.; Lovering, R.C.; Alnemri, E.S.; Bertin, J.; Boss, J.M.; Davis, B.K.; Flavell, R.A.; Girardin, S.E.; Godzik, A.; Harton, J.A.; et al. The NLR Gene Family: A Standard Nomenclature. Immunity 2008, 28, 285–287. [Google Scholar] [CrossRef] [Green Version]
- Davis, B.K.; Wen, H.; Ting, J.P.-Y. The Inflammasome NLRs in Immunity, Inflammation, and Associated Diseases. Annu. Rev. Immunol. 2011, 29, 707–735. [Google Scholar] [CrossRef] [Green Version]
- Ting, J.P.Y.; Duncan, J.A.; Lei, Y. How the Noninflammasome NLRs Function in the Innate Immune System. Science 2010, 327, 286–290. [Google Scholar] [CrossRef] [Green Version]
- Ting, J.P.-Y.; Willingham, S.B.; Bergstralh, D.T. NLRs at the Intersection of Cell Death and Immunity. Nat. Rev. Immunol. 2008, 8, 372–379. [Google Scholar] [CrossRef]
- Lamkanfi, M.; Dixit, V.M. Inflammasomes: Guardians of Cytosolic Sanctity. Immunol. Rev. 2009, 227, 95–105. [Google Scholar] [CrossRef]
- Allen, I.C.; TeKippe, E.M.; Woodford, R.-M.T.; Uronis, J.M.; Holl, E.K.; Rogers, A.B.; Herfarth, H.H.; Jobin, C.; Ting, J.P.-Y. The NLRP3 Inflammasome Functions as a Negative Regulator of Tumorigenesis during Colitis-Associated Cancer. J. Exp. Med. 2010, 207, 1045–1056. [Google Scholar] [CrossRef] [PubMed]
- Lei, Y.; Wen, H.; Ting, J.P.Y. The NLR Protein, NLRX1, and Its Partner, TUFM, Reduce Type I Interferon, and Enhance Autophagy. Autophagy 2013, 9, 432–433. [Google Scholar] [CrossRef] [Green Version]
- Mu, J.; Wang, W.; Chen, B.; Wu, L.; Li, B.; Mao, X.; Zhang, Z.; Fu, J.; Kuang, Y.; Sun, X.; et al. Mutations in NLRP2 and NLRP5 Cause Female Infertility Characterised by Early Embryonic Arrest. J. Med. Genet. 2019, 56, 471–480. [Google Scholar] [CrossRef] [PubMed]
- Murdoch, S.; Djuric, U.; Mazhar, B.; Seoud, M.; Khan, R.; Kuick, R.; Bagga, R.; Kircheisen, R.; Ao, A.; Ratti, B.; et al. Mutations in NALP7 Cause Recurrent Hydatidiform Moles and Reproductive Wastage in Humans. Nat. Genet. 2006, 38, 300–302. [Google Scholar] [CrossRef] [PubMed]
- Maddirevula, S.; Awartani, K.; Coskun, S.; AlNaim, L.; Ibrahim, N.; Abdulwahab, F.; Hashem, M.; Alhassan, S.; Alkuraya, F. A Genomics Approach to Females with Infertility and Recurrent Pregnancy Loss. Hum. Genet. 2020, 139. [Google Scholar] [CrossRef]
- Duncan, F.E.; Padilla-Banks, E.; Bernhardt, M.L.; Ord, T.S.; Jefferson, W.N.; Moss, S.B.; Williams, C.J. Transducin-like Enhancer of Split-6 (TLE6) Is a Substrate of Protein Kinase A Activity during Mouse Oocyte Maturation. Biol. Reprod. 2014, 90, 63. [Google Scholar] [CrossRef]
- Alazami, A.M.; Awad, S.M.; Coskun, S.; Al-Hassan, S.; Hijazi, H.; Abdulwahab, F.M.; Poizat, C.; Alkuraya, F.S. TLE6 Mutation Causes the Earliest Known Human Embryonic Lethality. Genome. Biol. 2015, 16. [Google Scholar] [CrossRef] [Green Version]
- Lin, J.; Xu, H.; Chen, B.; Wang, W.; Wang, L.; Sun, X.; Sang, Q. Expanding the Genetic and Phenotypic Spectrum of Female Infertility Caused by TLE6 Mutations. J. Assist. Reprod. Genet. 2020, 37, 437–442. [Google Scholar] [CrossRef]
- Wu, L.; Chen, H.; Li, D.; Song, D.; Chen, B.; Yan, Z.; Lyu, Q.; Wang, L.; Kuang, Y.; Li, B.; et al. Novel Mutations in PATL2: Expanding the Mutational Spectrum and Corresponding Phenotypic Variability Associated with Female Infertility. J. Hum. Genet. 2019, 64, 379–385. [Google Scholar] [CrossRef]
- Suzuki, T.; Matsushima, C.; Nishimura, S.; Higashiyama, T.; Sasabe, M.; Machida, Y. Identification of Phosphoinositide-Binding Protein PATELLIN2 as a Substrate of Arabidopsis MPK4 MAP Kinase during Septum Formation in Cytokinesis. Plant Cell Physiol. 2016, 57, 1744–1755. [Google Scholar] [CrossRef] [Green Version]
- Christou-Kent, M.; Kherraf, Z.; Amiri-Yekta, A.; Le Blévec, E.; Karaouzène, T.; Conne, B.; Escoffier, J.; Assou, S.; Guttin, A.; Lambert, E.; et al. PATL2 Is a Key Actor of Oocyte Maturation Whose Invalidation Causes Infertility in Women and Mice. EMBO Mol. Med. 2018, 10. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Song, D.; Mykytenko, D.; Kuang, Y.; Lv, Q.; Li, B.; Chen, B.; Mao, X.; Xu, Y.; Zukin, V.; et al. Novel Mutations in Genes Encoding Subcortical Maternal Complex Proteins May Cause Human Embryonic Developmental Arrest. Reprod. Biomed. Online 2018, 36, 698–704. [Google Scholar] [CrossRef]
- Zhang, W.; Chen, Z.; Zhang, D.; Zhao, B.; Liu, L.; Xie, Z.; Yao, Y.; Zheng, P. KHDC3L Mutation Causes Recurrent Pregnancy Loss by Inducing Genomic Instability of Human Early Embryonic Cells. PLoS Biol. 2019, 17, e3000468. [Google Scholar] [CrossRef] [PubMed]
- Zheng, P.; Dean, J. Oocyte-Specific Genes Affect Folliculogenesis, Fertilization, and Early Development. Semin. Reprod. Med. 2007, 25, 243–251. [Google Scholar] [CrossRef]
- Wu, X.; Viveiros, M.M.; Eppig, J.J.; Bai, Y.; Fitzpatrick, S.L.; Matzuk, M.M. Zygote Arrest 1 (ZAR1) Is a Novel Maternal-Effect Gene Critical for the Oocyte-to-Embryo Transition. Nat. Genet. 2003, 33, 187–191. [Google Scholar] [CrossRef] [PubMed]
- Uzbekova, S.; Roy-Sabau, M.; Dalbiès-Tran, R.; Perreau, C.; Papillier, P.; Mompart, F.; Thelie, A.; Pennetier, S.; Cognie, J.; Cadoret, V.; et al. Zygote Arrest 1 Gene in Pig, Cattle and Human: Evidence of Different Transcript Variants in Male and Female Germ Cells. Reprod. Biol. Endocrinol. 2006, 4, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, Y.; Yang, J.; Peng, Y.; Chen, T.; Huang, T.; Zhang, C.; Zhao, H. Variation Screening of Zygote Arrest 1(ZAR1) in Women with Recurrent Zygote Arrest During IVF/ICSI Programs. Reprod. Sci. 2020, 27, 2265–2270. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Dong, J.; Chen, B.; Du, J.; Kuang, Y.; Sun, X.; Fu, J.; Li, B.; Mu, J.; Zhang, Z.; et al. Homozygous Mutations in REC114 Cause Female Infertility Characterised by Multiple Pronuclei Formation and Early Embryonic Arrest. J. Med. Genet. 2020, 57, 187–194. [Google Scholar] [CrossRef]
- Kumar, R.; Oliver, C.; Brun, C.; Juarez-Martinez, A.B.; Tarabay, Y.; Kadlec, J.; de Massy, B. Mouse REC114 Is Essential for Meiotic DNA Double-Strand Break Formation and Forms a Complex with MEI4. Life Sci. Alliance 2018, 1, e201800259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pittman, D.; Lu, W.; Malone, R.E. Genetic and Molecular Analysis of REC114, an Early Meiotic Recombination Gene in Yeast. Curr. Genet. 1993, 23, 295–304. [Google Scholar] [CrossRef]
- Kordus, R.J.; Hossain, A.; Corso, M.C.; Chakraborty, H.; Whitman-Elia, G.F.; LaVoie, H.A. Cumulus Cell Pappalysin-1, Luteinizing Hormone/Choriogonadotropin Receptor, Amphiregulin and Hydroxy-Delta-5-Steroid Dehydrogenase, 3 Beta- and Steroid Delta-Isomerase 1 MRNA Levels Associate with Oocyte Developmental Competence and Embryo Outcomes. J. Assist. Reprod. Genet. 2019, 36, 1457–1469. [Google Scholar] [CrossRef] [PubMed]
- Kahraman, S.; Çetinkaya, C.P.; Çetinkaya, M.; Tüfekçi, M.A.; Ekmekçi, C.G.; Montag, M. Is There a Correlation between Follicle Size and Gene Expression in Cumulus Cells and Is Gene Expression an Indicator of Embryo Development? Reprod. Biol. Endocrinol. 2018, 16, 69. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.-H.; Chang, C.-L.; Wu, H.-M.; Chiu, Y.-M.; Chen, C.-K.; Wang, H.-S. Insulin-like Growth Factor-II (IGF-II), IGF-Binding Protein-3 (IGFBP-3), and IGFBP-4 in Follicular Fluid Are Associated with Oocyte Maturation and Embryo Development. Fertil. Steril. 2006, 86, 1392–1401. [Google Scholar] [CrossRef] [PubMed]
- Das, S.; Chattopadhyay, R.; Ghosh, S.; Ghosh, S.; Goswami, S.K.; Chakravarty, B.N.; Chaudhury, K. Reactive Oxygen Species Level in Follicular Fluid--Embryo Quality Marker in IVF? Hum. Reprod. 2006, 21, 2403–2407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Revelli, A.; Canosa, S.; Bergandi, L.; Skorokhod, O.A.; Biasoni, V.; Carosso, A.; Bertagna, A.; Maule, M.; Aldieri, E.; D’Eufemia, M.D.; et al. Oocyte Polarized Light Microscopy, Assay of Specific Follicular Fluid Metabolites, and Gene Expression in Cumulus Cells as Different Approaches to Predict Fertilization Efficiency after ICSI. Reprod. Biol. Endocrinol. 2017, 15, 47. [Google Scholar] [CrossRef]
- Janny, L.; Menezo, Y.J. Maternal Age Effect on Early Human Embryonic Development and Blastocyst Formation. Mol. Reprod. Dev. 1996, 45, 31–37. [Google Scholar] [CrossRef]
- Rodrigo, L.; Peinado, V.; Vera-Rodriguez, M.; Lopez-Iglesias, P.; Navarro, R.; Harton, G.L.; Simon, C.; Rubio, C. Different Contribution of Maternal and Paternal Age to Mosaicism and Partial Duplication/Deletions in Blastocyst Biopsies Assessed by next Generation Sequencing. Fertil. Steril. 2017, 108, e274–e275. [Google Scholar] [CrossRef] [Green Version]
- Sekhon, L.; Feuerstein, J.; Nazem, T.G.; Briton-Jones, C.; Lee, J.A.; Grunfeld, L.; Copperman, A.B. The Incidence of Mosaicism Is Not Associated with Advanced Maternal Age or Diminished Ovarian Reserve. Fertil. Steril. 2017, 108, e217. [Google Scholar] [CrossRef]
- Yeoh, M.H.; Chen, J.J.; Sinthamoney, E.; Wong, P.S. Clinical Outcome: The Relationship between Mosaicism and Advanced Maternal Age with the Use of Next Generation Sequencing (NGS). Reprod. Biomed. Online 2019, 38, e48. [Google Scholar] [CrossRef]
- Reich, J.; Blakemore, J.K.; Besser, A.G.; Hodes-Wertz, B.; Grifo, J.A. THE EFFECT OF MATERNAL AGE ON CHROMOSOMAL MOSAICISM: AN ANALYSIS BY CHROMOSOME TYPE AND MOSAIC RESULT. Fertil. Steril. 2020, 114, e419–e420. [Google Scholar] [CrossRef]
- Mikwar, M.; MacFarlane, A.J.; Marchetti, F. Mechanisms of Oocyte Aneuploidy Associated with Advanced Maternal Age. Mutat. Res. Rev. Mutat. Res. 2020, 785, 108320. [Google Scholar] [CrossRef] [PubMed]
- Boynukalin, F.K.; Serdarogullari, M.; Gultomruk, M.; Coban, O.; Findikli, N.; Bahceci, M. The Impact of Endometriosis on Early Embryo Morphokinetics: A Case-Control Study. Syst. Biol. Reprod. Med. 2019, 65, 250–257. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, A.M.; Somigliana, E.; Vercellini, P.; Pagliardini, L.; Candiani, M.; Vigano, P. Endometriosis as a Detrimental Condition for Granulosa Cell Steroidogenesis and Development: From Molecular Alterations to Clinical Impact. J. Steroid. Biochem. Mol. Biol. 2016, 155, 35–46. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, A.M.; Vanni, V.S.; Bartiromo, L.; Papaleo, E.; Zilberberg, E.; Candiani, M.; Orvieto, R.; Viganò, P. Is the Oocyte Quality Affected by Endometriosis? A Review of the Literature. J. Ovarian Res. 2017, 10, 43. [Google Scholar] [CrossRef] [Green Version]
- Boucret, L.; Bouet, P.-E.; Riou, J.; Legendre, G.; Delbos, L.; Hachem, H.E.; Descamps, P.; Reynier, P.; May-Panloup, P. Endometriosis Lowers the Cumulative Live Birth Rates in IVF by Decreasing the Number of Embryos but Not Their Quality. J. Clin. Med. 2020, 9, 2478. [Google Scholar] [CrossRef]
- Sapkota, Y.; Steinthorsdottir, V.; Morris, A.P.; Fassbender, A.; Rahmioglu, N.; De Vivo, I.; Buring, J.E.; Zhang, F.; Edwards, T.L.; Jones, S.; et al. Meta-Analysis Identifies Five Novel Loci Associated with Endometriosis Highlighting Key Genes Involved in Hormone Metabolism. Nat. Commun. 2017, 8, 15539. [Google Scholar] [CrossRef] [Green Version]
- Méar, L.; Herr, M.; Fauconnier, A.; Pineau, C.; Vialard, F. Polymorphisms and Endometriosis: A Systematic Review and Meta-Analyses. Hum. Reprod. Update 2020, 26, 73–103. [Google Scholar] [CrossRef]
- Hernandez Gifford, J.A. The Role of WNT Signaling in Adult Ovarian Folliculogenesis. Reproduction 2015, 150, R137–R148. [Google Scholar] [CrossRef] [Green Version]
- Tepekoy, F.; Akkoyunlu, G.; Demir, R. The Role of Wnt Signaling Members in the Uterus and Embryo during Pre-Implantation and Implantation. J. Assist. Reprod. Genet. 2015, 32, 337–346. [Google Scholar] [CrossRef] [Green Version]
- Sidrat, T.; Khan, A.A.; Idrees, M.; Joo, M.-D.; Xu, L.; Lee, K.-L.; Kong, I.-K. Role of Wnt Signaling During In-Vitro Bovine Blastocyst Development and Maturation in Synergism with PPARδ Signaling. Cells 2020, 9, 923. [Google Scholar] [CrossRef] [Green Version]
- Simopoulou, M.; Rapani, A.; Grigoriadis, S.; Pantou, A.; Tsioulou, P.; Maziotis, E.; Tzanakaki, D.; Triantafyllidou, O.; Kalampokas, T.; Siristatidis, C.; et al. Getting to Know Endometriosis-Related Infertility Better: A Review on How Endometriosis Affects Oocyte Quality and Embryo Development. Biomedicines 2021, 9, 273. [Google Scholar] [CrossRef]
- Konstantinos, S.; Tsioulou, P.; Maziotis, E.; Polina, G.; Argyro, G.; Grigoriadis, S.; Rapani, A.; Nezos, A.; Agni, P.; Koutsilieris, M.; et al. Assessing the Practice of LuPOR for Poor Responders: A Prospective Study Evaluating Follicular Fluid CfDNA Levels during Natural IVF Cycles. J. Assist. Reprod. Genet. 2020, 37. [Google Scholar] [CrossRef]
- Hojnik, N.; Vlaisavljević, V.; Kovačič, B. Morphokinetic Characteristics and Developmental Potential of In Vitro Cultured Embryos from Natural Cycles in Patients with Poor Ovarian Response. Biomed. Res. Int. 2016, 2016, 4286528. [Google Scholar] [CrossRef] [Green Version]
- Nesbit, C.B.; Huang, J.; Singh, B.; Maher, J.Y.; Pastore, L.M.; Segars, J. New Perspectives on the Genetic Causes of Diminished Ovarian Reserve and Opportunities for Genetic Screening: Systematic Review and Meta-Analysis. FS Rev. 2020, 1, 1–15. [Google Scholar] [CrossRef]
- Mila, M.; Alvarez-Mora, M.I.; Madrigal, I.; Rodriguez-Revenga, L. Fragile X Syndrome: An Overview and Update of the FMR1 Gene. Clin. Genet. 2018, 93, 197–205. [Google Scholar] [CrossRef] [PubMed]
- Monti, M.; Redi, C. Oogenesis Specific Genes (Nobox, Oct4, Bmp15, Gdf9, Oogenesin1 and Oogenesin2) Are Differentially Expressed during Natural and Gonadotropin-Induced Mouse Follicular Development. Mol. Reprod. Dev. 2009, 76, 994–1003. [Google Scholar] [CrossRef] [PubMed]
- Barasoain, M.; Barrenetxea, G.; Huerta, I.; Télez, M.; Criado, B.; Arrieta, I. Study of the Genetic Etiology of Primary Ovarian Insufficiency: FMR1 Gene. Genes (Basel) 2016, 7, 123. [Google Scholar] [CrossRef] [Green Version]
- Greene, A.D.; Patounakis, G.; Segars, J.H. Genetic Associations with Diminished Ovarian Reserve: A Systematic Review of the Literature. J. Assist. Reprod. Genet. 2014, 31, 935–946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Society for Assisted Reproductive Technology National Summary Report. Available online: https://www.sartcorsonline.com/rptCSR_PublicMultYear.aspx?reportingYear=2014 (accessed on 18 July 2021).
- Dumasia, K.; Kumar, A.; Deshpande, S.; Balasinor, N.H. Estrogen Signaling, through Estrogen Receptor β, Regulates DNA Methylation and Its Machinery in Male Germ Line in Adult Rats. Epigenetics 2017, 12, 476–483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Q.F.; Feng, H.L.; Zhao, L.; Liu, P.; Li, L.; Yan, J.; Qiao, J. Alteration of ERβ Gene RsaI Polymorphism May Contribute to Reduced Fertilization Rate and Embryonic Developmental Competence. Asian J. 2010, 13, 317–321. [Google Scholar] [CrossRef] [Green Version]
- Aarabi, M.; Balakier, H.; Bashar, S.; Moskovtsev, S.I.; Sutovsky, P.; Librach, C.L.; Oko, R. Sperm Content of Postacrosomal WW Binding Protein Is Related to Fertilization Outcomes in Patients Undergoing Assisted Reproductive Technology. Fertil. Steril. 2014, 102, 440–447. [Google Scholar] [CrossRef]
- Aarabi, M.; Balakier, H.; Bashar, S.; Moskovtsev, S.I.; Sutovsky, P.; Librach, C.L.; Oko, R. Sperm-Derived WW Domain-Binding Protein, PAWP, Elicits Calcium Oscillations and Oocyte Activation in Humans and Mice. Faseb J. 2014, 28, 4434–4440. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.-Y.; Minahan, K.; Merriman, J.A.; Jones, K.T. Calmodulin-Dependent Protein Kinase Gamma 3 (CamKIIgamma3) Mediates the Cell Cycle Resumption of Metaphase II Eggs in Mouse. Development 2009, 136, 4077–4081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miao, Y.-L.; Williams, C.J. Calcium Signaling in Mammalian Egg Activation and Embryo Development: The Influence of Subcellular Localization. Mol. Reprod. Dev. 2012, 79, 742–756. [Google Scholar] [CrossRef] [Green Version]
- Nomikos, M.; Sanders, J.R.; Theodoridou, M.; Kashir, J.; Matthews, E.; Nounesis, G.; Lai, F.A.; Swann, K. Sperm-Specific Post-Acrosomal WW-Domain Binding Protein (PAWP) Does Not Cause Ca2+ Release in Mouse Oocytes. Mol. Hum. Reprod. 2014, 20, 938–947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyazaki, S.; Shirakawa, H.; Nakada, K.; Honda, Y. Essential Role of the Inositol 1,4,5-Trisphosphate Receptor/Ca2+ Release Channel in Ca2+ Waves and Ca2+ Oscillations at Fertilization of Mammalian Eggs. Dev. Biol. 1993, 158, 62–78. [Google Scholar] [CrossRef]
- Wu, A.T.H.; Sutovsky, P.; Manandhar, G.; Xu, W.; Katayama, M.; Day, B.N.; Park, K.-W.; Yi, Y.-J.; Xi, Y.W.; Prather, R.S.; et al. PAWP, a Sperm-Specific WW Domain-Binding Protein, Promotes Meiotic Resumption and Pronuclear Development during Fertilization. J. Biol. Chem. 2007, 282, 12164–12175. [Google Scholar] [CrossRef] [Green Version]
- Kennedy, C.E.; Krieger, K.B.; Sutovsky, M.; Xu, W.; Vargovič, P.; Didion, B.A.; Ellersieck, M.R.; Hennessy, M.E.; Verstegen, J.; Oko, R.; et al. Protein Expression Pattern of PAWP in Bull Spermatozoa Is Associated with Sperm Quality and Fertility Following Artificial Insemination. Mol. Reprod. Dev. 2014, 81, 436–449. [Google Scholar] [CrossRef] [PubMed]
- Panner Selvam, M.K.; Agarwal, A. A Systematic Review on Sperm DNA Fragmentation in Male Factor Infertility: Laboratory Assessment. Arab. J. Urol. 2018, 16, 65–76. [Google Scholar] [CrossRef] [Green Version]
- Vaughan, D.A.; Tirado, E.; Garcia, D.; Datta, V.; Sakkas, D. DNA Fragmentation of Sperm: A Radical Examination of the Contribution of Oxidative Stress and Age in 16 945 Semen Samples. Hum. Reprod. 2020, 35, 2188–2196. [Google Scholar] [CrossRef]
- Jiang, W.; Jin, F.; Zhou, L. Influence of the DNA integrity of optimized sperm on the embryonic development and clinical outcomes of in vitro fertilization and embryo transfer. Zhonghua Nan Ke Xue 2016, 22, 425–431. [Google Scholar] [PubMed]
- Qiu, Y.; Yang, H.; Li, C.; Xu, C. Progress in Research on Sperm DNA Fragmentation. Med. Sci. Monit. 2020, 26, e918746-1–e918746-11. [Google Scholar] [CrossRef] [PubMed]
- Lazaros, L.; Vartholomatos, G.; Pamporaki, C.; Kosmas, I.; Takenaka, A.; Makrydimas, G.; Sofikitis, N.; Stefos, T.; Zikopoulos, K.; Hatzi, E.; et al. Sperm Flow Cytometric Parameters Are Associated with ICSI Outcome. Reprod. Biomed. Online 2013, 26, 611–618. [Google Scholar] [CrossRef] [PubMed]
- Virant-Klun, I.; Tomazevic, T.; Meden-Vrtovec, H. Sperm Single-Stranded DNA, Detected by Acridine Orange Staining, Reduces Fertilization and Quality of ICSI-Derived Embryos. J. Assist. Reprod. Genet. 2002, 19, 319–328. [Google Scholar] [CrossRef]
- Slameňová, D.; Dušinská, M.; Gábelová, A.; Bohušová, T.; Ruppová, K. Decemtione (Imidan)-Induced Single-Strand Breaks to Human Dna, Mutations at the Hgprt Locus of V79 Cells, and Morphological Transformations of Embryo Cells. Environ. Mol. Mutagenesis 1992, 20, 73–78. [Google Scholar] [CrossRef]
- Anindya, R. Single-Stranded DNA Damage: Protecting the Single-Stranded DNA from Chemical Attack. DNA Repair 2020, 87, 102804. [Google Scholar] [CrossRef]
- Deng, C.; Li, T.; Xie, Y.; Guo, Y.; Yang, Q.-Y.; Liang, X.; Deng, C.-H.; Liu, G.-H. Sperm DNA Fragmentation Index Influences Assisted Reproductive Technology Outcome: A Systematic Review and Meta-Analysis Combined with a Retrospective Cohort Study. Andrologia 2019, 51, e13263. [Google Scholar] [CrossRef]
- Simon, L.; Zini, A.; Dyachenko, A.; Ciampi, A.; Carrell, D.T. A Systematic Review and Meta-Analysis to Determine the Effect of Sperm DNA Damage on in Vitro Fertilization and Intracytoplasmic Sperm Injection Outcome. Asian J. 2017, 19, 80–90. [Google Scholar] [CrossRef]
- Diagnostic Evaluation of the Infertile Male: A Committee Opinion. Fertil. Steril. 2015, 103, e18–e25. [CrossRef]
- Rienzi, L.; Mazzilli, R.; Ubaldi, F.M. Sperm DNA Fragmentation to Predict Embryo Development, Implantation, and Miscarriage: Still an Open Question. Fertil. Steril. 2019, 112, 466. [Google Scholar] [CrossRef] [Green Version]
- Green, K.A.; Patounakis, G.; Dougherty, M.P.; Werner, M.D.; Scott, R.T.; Franasiak, J.M. Sperm DNA Fragmentation on the Day of Fertilization Is Not Associated with Embryologic or Clinical Outcomes after IVF/ICSI. J. Assist. Reprod. Genet. 2020, 37, 71–76. [Google Scholar] [CrossRef] [PubMed]
- Cissen, M.; van Wely, M.; Scholten, I.; Mansell, S.; de Bruin, J.P.; Mol, B.W.; Braat, D.; Repping, S.; Hamer, G. Measuring Sperm DNA Fragmentation and Clinical Outcomes of Medically Assisted Reproduction: A Systematic Review and Meta-Analysis. PLoS ONE 2016, 11, e0165125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Virant-Klun, I.; Tomazevic, T.; Zorn, B.; Bacer-Kermavner, L.; Mivsek, J.; Meden-Vrtovec, H. Blastocyst Formation—Good Indicator of Clinical Results after ICSI with Testicular Spermatozoa. Hum. Reprod. 2003, 18, 1070–1076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desai, N.; Gill, P.; Tadros, N.N.; Goldberg, J.M.; Sabanegh, E.; Falcone, T. Azoospermia and Embryo Morphokinetics: Testicular Sperm-Derived Embryos Exhibit Delays in Early Cell Cycle Events and Increased Arrest Prior to Compaction. J. Assist. Reprod. Genet. 2018, 35, 1339–1348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghieh, F.; Mitchell, V.; Mandon-Pepin, B.; Vialard, F. Genetic Defects in Human Azoospermia. Basic Clin. 2019, 29. [Google Scholar] [CrossRef] [Green Version]
- Daneshmandpour, Y.; Bahmanpour, Z.; Hamzeiy, H.; Mazaheri Moghaddam, M.; Mazaheri Moghaddam, M.; Khademi, B.; Sakhinia, E. MicroRNAs Association with Azoospermia, Oligospermia, Asthenozoospermia, and Teratozoospermia: A Systematic Review. J. Assist. Reprod. Genet. 2020, 37, 763–775. [Google Scholar] [CrossRef] [PubMed]
- Colpi, G.M.; Francavilla, S.; Haidl, G.; Link, K.; Behre, H.M.; Goulis, D.G.; Krausz, C.; Giwercman, A. European Academy of Andrology Guideline Management of Oligo-Astheno-Teratozoospermia. Andrology 2018, 6, 513–524. [Google Scholar] [CrossRef] [Green Version]
- Pantos, K.; Sfakianoudis, K.; Maziotis, E.; Rapani, A.; Karantzali, E.; Gounari-Papaioannou, A.; Vaxevanoglou, T.; Koutsilieris, M.; Simopoulou, M. Abnormal Fertilization in ICSI and Its Association with Abnormal Semen Parameters: A Retrospective Observational Study on 1855 Cases. Asian J. 2021. [Google Scholar] [CrossRef]
- Dam, A.H.D.M.; Koscinski, I.; Kremer, J.A.M.; Moutou, C.; Jaeger, A.-S.; Oudakker, A.R.; Tournaye, H.; Charlet, N.; Lagier-Tourenne, C.; van Bokhoven, H.; et al. Homozygous Mutation in SPATA16 Is Associated with Male Infertility in Human Globozoospermia. Am. J. Hum. Genet. 2007, 81, 813–820. [Google Scholar] [CrossRef] [Green Version]
- Canepa, P.; Casciano, I.; De Leo, C.; Massarotti, C.; Anserini, P.; Remorgida, V.; Scaruffi, P. A Successful Healthy Childbirth and an Ongoing Evolutive Pregnancy in a Case of Partial Globozoospermia by Hyaluronic Acid Sperm Selection. Andrologia 2019, 51, e13178. [Google Scholar] [CrossRef]
- Perrin, A.; Coat, C.; Nguyen, M.H.; Talagas, M.; Morel, F.; Amice, J.; De Braekeleer, M. Molecular Cytogenetic and Genetic Aspects of Globozoospermia: A Review. Andrologia 2013, 45, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Celse, T.; Cazin, C.; Mietton, F.; Martinez, G.; Martinez, D.; Thierry-Mieg, N.; Septier, A.; Guillemain, C.; Beurois, J.; Clergeau, A.; et al. Genetic Analyses of a Large Cohort of Infertile Patients with Globozoospermia, DPY19L2 Still the Main Actor, GGN Confirmed as a Guest Player. Hum. Genet. 2021, 140, 43–57. [Google Scholar] [CrossRef] [PubMed]
- De Braekeleer, M.; Nguyen, M.H.; Morel, F.; Perrin, A. Genetic Aspects of Monomorphic Teratozoospermia: A Review. J. Assist. Reprod. Genet. 2015, 32, 615–623. [Google Scholar] [CrossRef] [Green Version]
- Miller, J.E.; Smith, T.T. The Effect of Intracytoplasmic Sperm Injection and Semen Parameters on Blastocyst Development in Vitro. Hum. Reprod. 2001, 16, 918–924. [Google Scholar] [CrossRef] [Green Version]
- Abbas, A.M.; Hussein, R.S.; Elsenity, M.A.; Samaha, I.I.; El Etriby, K.A.; Abd El-Ghany, M.F.; Khalifa, M.A.; Abdelrheem, S.S.; Ahmed, A.A.; Khodry, M.M. Higher Clinical Pregnancy Rate with In-Vitro Fertilization versus Intracytoplasmic Sperm Injection in Treatment of Non-Male Factor Infertility: Systematic Review and Meta-Analysis. J. Gynecol. Obs. Hum. Reprod. 2020, 49, 101706. [Google Scholar] [CrossRef]
- Palermo, G.D.; Neri, Q.V.; Takeuchi, T.; Squires, J.; Moy, F.; Rosenwaks, Z. Genetic and Epigenetic Characteristics of ICSI Children. Reprod. Biomed. Online 2008, 17, 820–833. [Google Scholar] [CrossRef]
- Whitelaw, N.; Bhattacharya, S.; Hoad, G.; Horgan, G.W.; Hamilton, M.; Haggarty, P. Epigenetic Status in the Offspring of Spontaneous and Assisted Conception. Hum. Reprod. 2014, 29, 1452–1458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Novakovic, B.; Lewis, S.; Halliday, J.; Kennedy, J.; Burgner, D.P.; Czajko, A.; Kim, B.; Sexton-Oates, A.; Juonala, M.; Hammarberg, K.; et al. Assisted Reproductive Technologies Are Associated with Limited Epigenetic Variation at Birth That Largely Resolves by Adulthood. Nat. Commun. 2019, 10, 3922. [Google Scholar] [CrossRef]
- Darwish, E.; Magdi, Y. A Preliminary Report of Successful Cleavage after Calcium Ionophore Activation at ICSI in Cases with Previous Arrest at the Pronuclear Stage. Reprod. Biomed. Online 2015, 31, 799–804. [Google Scholar] [CrossRef] [Green Version]
- Ebner, T.; Oppelt, P.; Wöber, M.; Staples, P.; Mayer, R.B.; Sonnleitner, U.; Bulfon-Vogl, S.; Gruber, I.; Haid, A.E.; Shebl, O. Treatment with Ca2+ Ionophore Improves Embryo Development and Outcome in Cases with Previous Developmental Problems: A Prospective Multicenter Study. Hum. Reprod. 2015, 30, 97–102. [Google Scholar] [CrossRef] [Green Version]
- Murugesu, S.; Saso, S.; Jones, B.P.; Bracewell-Milnes, T.; Athanasiou, T.; Mania, A.; Serhal, P.; Ben-Nagi, J. Does the Use of Calcium Ionophore during Artificial Oocyte Activation Demonstrate an Effect on Pregnancy Rate? A Meta-Analysis. Fertil. Steril. 2017, 108, 468–482.e3. [Google Scholar] [CrossRef] [Green Version]
- Long, R.; Wang, M.; Yang, Q.Y.; Hu, S.Q.; Zhu, L.X.; Jin, L. Risk of Birth Defects in Children Conceived by Artificial Oocyte Activation and Intracytoplasmic Sperm Injection: A Meta-Analysis. Reprod. Biol. Endocrinol. 2020, 18, 123. [Google Scholar] [CrossRef]
- Maheshwari, A.; Hamilton, M.; Bhattacharya, S. Effect of Female Age on the Diagnostic Categories of Infertility. Hum. Reprod. 2008, 23, 538–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mazur, D.J.; Lipshultz, L.I. Infertility in the Aging Male. Curr. Urol. Rep. 2018, 19, 54. [Google Scholar] [CrossRef] [PubMed]
- Schulz, K.N.; Harrison, M.M. Mechanisms Regulating Zygotic Genome Activation. Nat. Rev. Genet. 2019, 20, 221–234. [Google Scholar] [CrossRef] [PubMed]
- Wong, C.C.; Loewke, K.E.; Bossert, N.L.; Behr, B.; De Jonge, C.J.; Baer, T.M.; Reijo Pera, R.A. Non-Invasive Imaging of Human Embryos before Embryonic Genome Activation Predicts Development to the Blastocyst Stage. Nat. Biotechnol. 2010, 28, 1115–1121. [Google Scholar] [CrossRef]
- Rienzi, L.; Vajta, G.; Ubaldi, F. Predictive Value of Oocyte Morphology in Human IVF: A Systematic Review of the Literature. Hum. Reprod. Update 2011, 17, 34–45. [Google Scholar] [CrossRef] [Green Version]
- Lazzaroni-Tealdi, E.; Barad, D.H.; Albertini, D.F.; Yu, Y.; Kushnir, V.A.; Russell, H.; Wu, Y.-G.; Gleicher, N. Oocyte Scoring Enhances Embryo-Scoring in Predicting Pregnancy Chances with IVF Where It Counts Most. PLoS ONE 2015, 10, e0143632. [Google Scholar] [CrossRef]
- Maziotis, E.; Sfakianoudis, K.; Giannelou, P.; Grigoriadis, S.; Rapani, A.; Tsioulou, P.; Nikolettos, K.; Pantou, A.; Tiptiri-Kourpeti, A.; Koutsilieris, M.; et al. Evaluating the Value of Day 0 of an ICSI Cycle on Indicating Laboratory Outcome. Sci. Rep. 2020, 10, 19325. [Google Scholar] [CrossRef]
- Bjorkman, S.; Taylor, H.S. MicroRNAs in Endometriosis: Biological Function and Emerging Biomarker Candidates. Biol. Reprod. 2019, 100, 1135–1146. [Google Scholar] [CrossRef]
- Khazaie, Y.; Nasr Esfahani, M.H. MicroRNA and Male Infertility: A Potential for Diagnosis. Int. J. Fertil. Steril. 2014, 8, 113–118. [Google Scholar]
- Kim, J.; Lee, J.; Jun, J.H. Identification of Differentially Expressed MicroRNAs in Outgrowth Embryos Compared with Blastocysts and Non-Outgrowth Embryos in Mice. Reprod. Fertil. Dev. 2019, 31, 645–657. [Google Scholar] [CrossRef]
- Campisi, J.; d’Adda di Fagagna, F. Cellular Senescence: When Bad Things Happen to Good Cells. Nat. Rev. Mol. Cell Biol. 2007, 8, 729–740. [Google Scholar] [CrossRef]
- Gorgoulis, V.; Adams, P.D.; Alimonti, A.; Bennett, D.C.; Bischof, O.; Bishop, C.; Campisi, J.; Collado, M.; Evangelou, K.; Ferbeyre, G.; et al. Cellular Senescence: Defining a Path Forward. Cell 2019, 179, 813–827. [Google Scholar] [CrossRef] [PubMed]
- Costa-Borges, N.; Spath, K.; Miguel-Escalada, I.; Mestres, E.; Balmaseda, R.; Serafín, A.; Garcia-Jiménez, M.; Vanrell, I.; González, J.; Rink, K.; et al. Maternal Spindle Transfer Overcomes Embryo Developmental Arrest Caused by Ooplasmic Defects in Mice. eLife 2020, 9, e48591. [Google Scholar] [CrossRef] [PubMed]
- Loutradis, D.; Drakakis, P.; Kallianidis, K.; Sofikitis, N.; Kallipolitis, G.; Milingos, S.; Makris, N.; Michalas, S. Biological Factors in Culture Media Affecting in Vitro Fertilization, Preimplantation Embryo Development, and Implantation. Ann. N.Y. Acad. Sci. 2000, 900, 325–335. [Google Scholar] [CrossRef]
- Sfontouris, I.A.; Martins, W.P.; Nastri, C.O.; Viana, I.G.R.; Navarro, P.A.; Raine-Fenning, N.; van der Poel, S.; Rienzi, L.; Racowsky, C. Blastocyst Culture Using Single versus Sequential Media in Clinical IVF: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. J. Assist. Reprod. Genet. 2016, 33, 1261–1272. [Google Scholar] [CrossRef]
- Simopoulou, M.; Sfakianoudis, K.; Rapani, A.; Giannelou, P.; Anifandis, G.; Bolaris, S.; Pantou, A.; Lambropoulou, M.; Pappas, A.; Deligeoroglou, E.; et al. Considerations Regarding Embryo Culture Conditions: From Media to Epigenetics. Vivo 2018, 32, 451–460. [Google Scholar] [CrossRef] [Green Version]
- Estill, M.S.; Bolnick, J.M.; Waterland, R.A.; Bolnick, A.D.; Diamond, M.P.; Krawetz, S.A. Assisted Reproductive Technology Alters Deoxyribonucleic Acid Methylation Profiles in Bloodspots of Newborn Infants. Fertil. Steril. 2016, 106, 629–639.e10. [Google Scholar] [CrossRef]
- El Hajj, N.; Haertle, L.; Dittrich, M.; Denk, S.; Lehnen, H.; Hahn, T.; Schorsch, M.; Haaf, T. DNA Methylation Signatures in Cord Blood of ICSI Children. Hum. Reprod. 2017, 32, 1761–1769. [Google Scholar] [CrossRef] [Green Version]
- Barberet, J.; Barry, F.; Choux, C.; Guilleman, M.; Karoui, S.; Simonot, R.; Bruno, C.; Fauque, P. What Impact Does Oocyte Vitrification Have on Epigenetics and Gene Expression? Clin. Epigenetics 2020, 12, 121. [Google Scholar] [CrossRef]
- Chatterjee, A.; Saha, D.; Niemann, H.; Gryshkov, O.; Glasmacher, B.; Hofmann, N. Effects of Cryopreservation on the Epigenetic Profile of Cells. Cryobiology 2017, 74, 1–7. [Google Scholar] [CrossRef]
- Wu, Y.; Lv, Z.; Yang, Y.; Dong, G.; Yu, Y.; Cui, Y.; Tong, M.; Wang, L.; Zhou, Z.; Zhu, H.; et al. Blastomere Biopsy Influences Epigenetic Reprogramming during Early Embryo Development, Which Impacts Neural Development and Function in Resulting Mice. Cell Mol. Life Sci. 2014, 71, 1761–1774. [Google Scholar] [CrossRef]
- Simopoulou, M.; Sfakianoudis, K.; Maziotis, E.; Tsioulou, P.; Grigoriadis, S.; Rapani, A.; Giannelou, P.; Asimakopoulou, M.; Kokkali, G.; Pantou, A.; et al. PGT-A: Who and When? A Systematic Review and Network Meta-Analysis of RCTs. J. Assist. Reprod. Genet. 2021. [Google Scholar] [CrossRef]
- Consensus Group, C. “There Is Only One Thing That Is Truly Important in an IVF Laboratory: Everything” Cairo Consensus Guidelines on IVF Culture Conditions. Reprod. Biomed. Online 2020, 40, 33–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sunde, A.; Brison, D.; Dumoulin, J.; Harper, J.; Lundin, K.; Magli, M.C.; Van den Abbeel, E.; Veiga, A. Time to Take Human Embryo Culture Seriously. Hum. Reprod. 2016, 31, 2174–2182. [Google Scholar] [CrossRef] [PubMed]




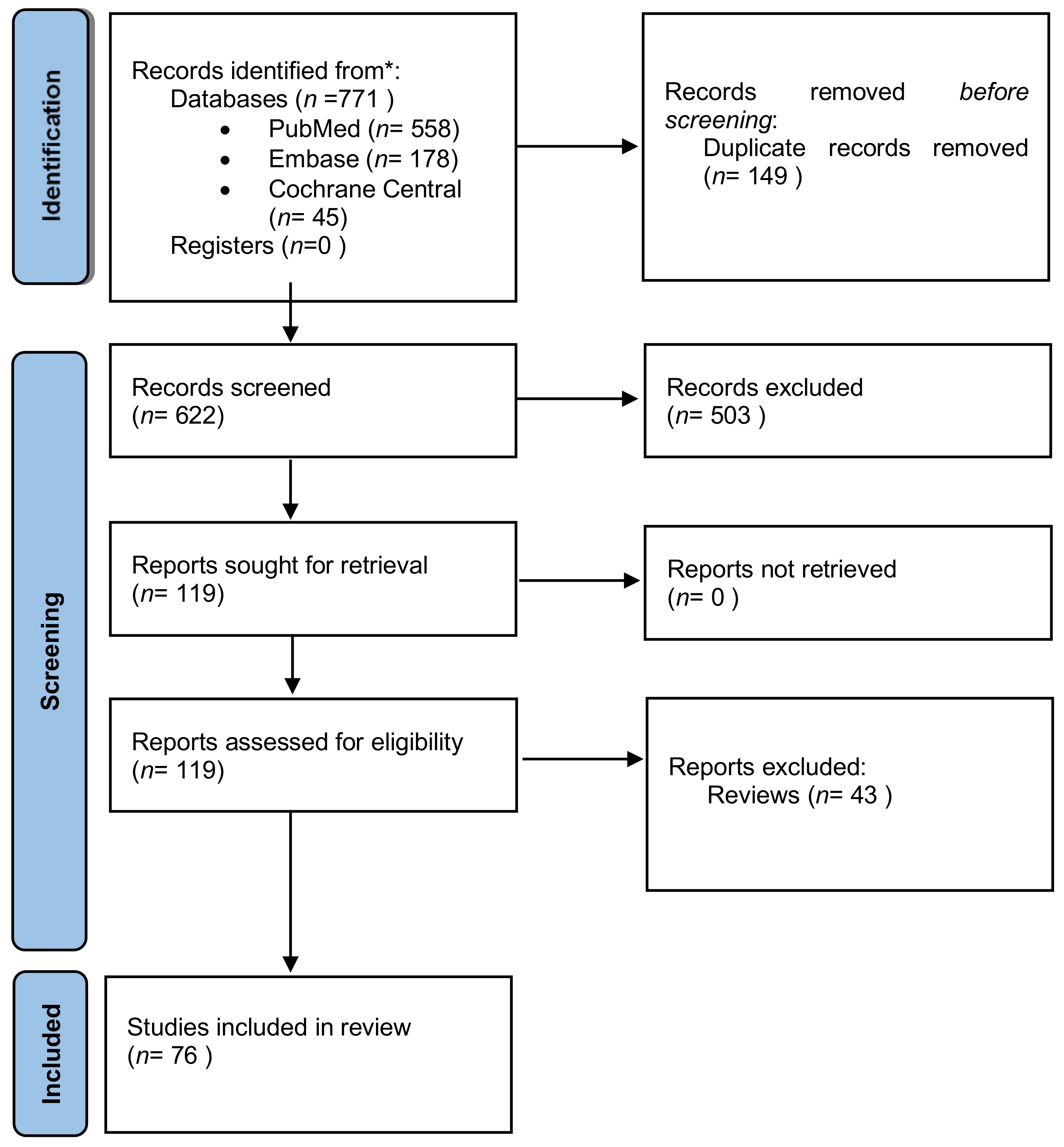{kind=link}
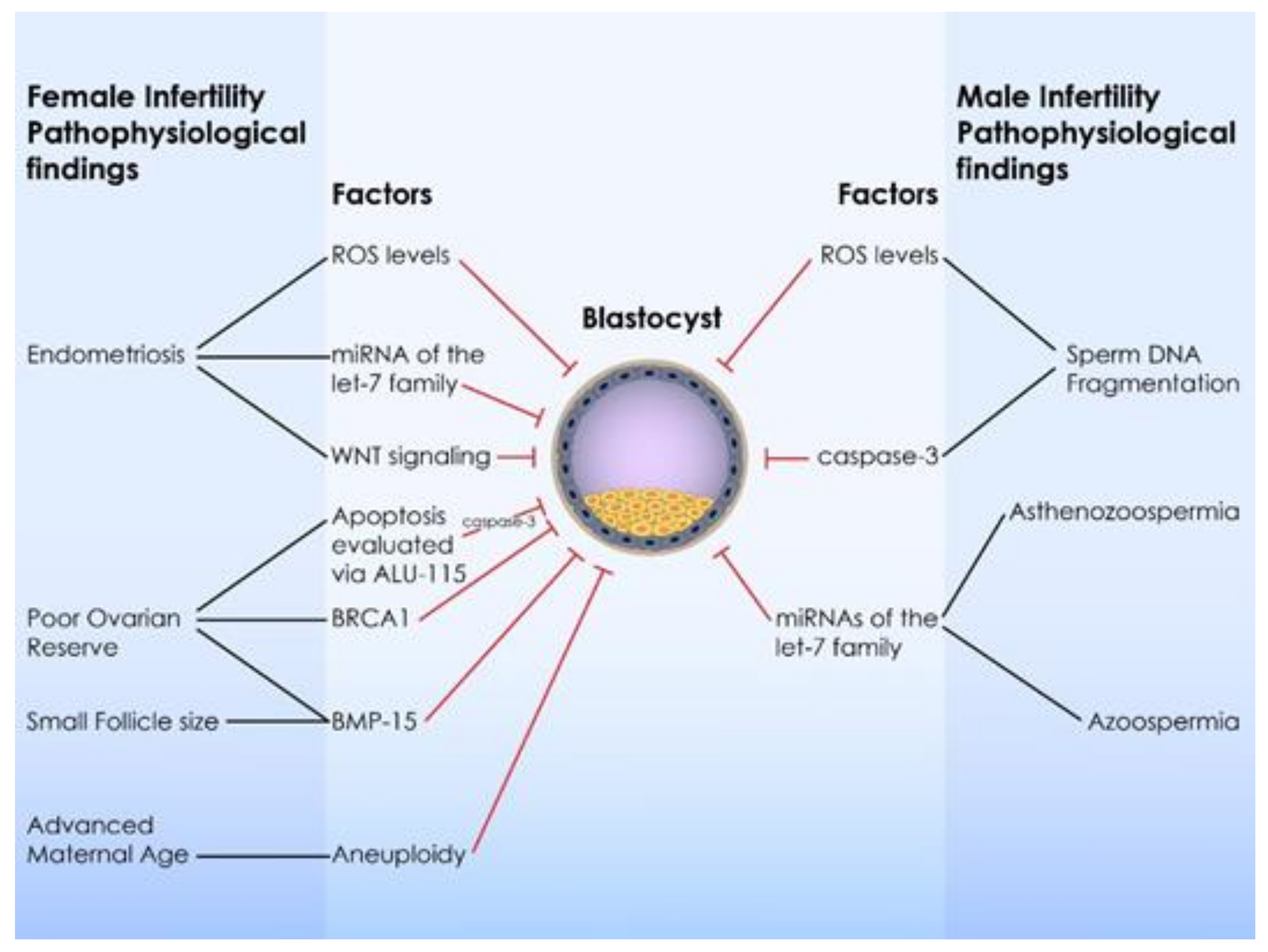{kind=link}
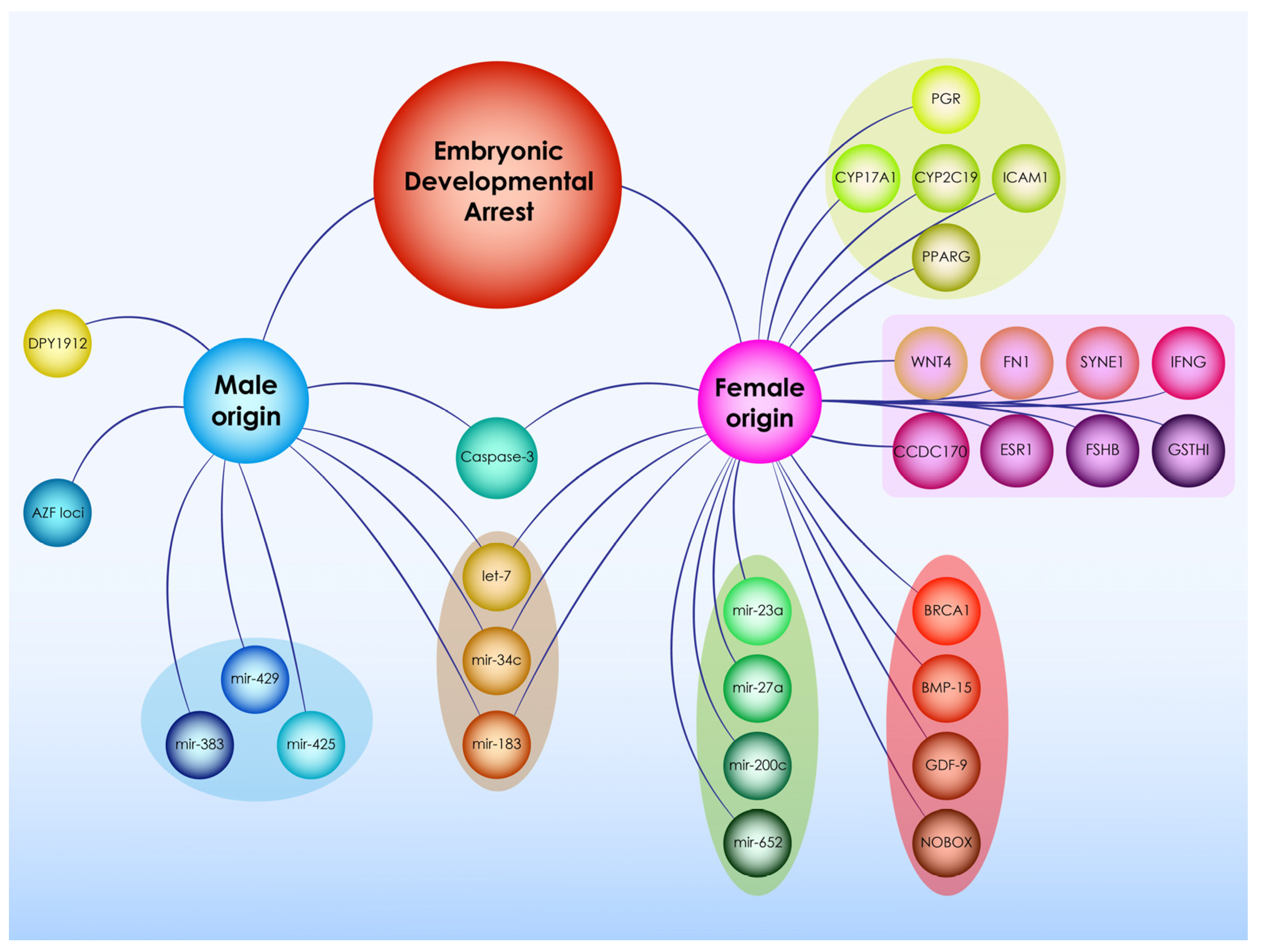{kind=link}
| Study ID | GENE | Variations | Study Origin | Sample Size | |
|---|---|---|---|---|---|
| Tian | ZAR1 | rs117545505 and rs17609740 | China | 47 women with cleavage failure | Controls: 93 women undergoing IVF/ICSI and 188 women with natural conception |
| Sha | TUBB8 | NP | China | 66 women | |
| Chen | TUBB8 | 28 variants | China | 30 families | |
| Yuan | TUBB8 | c.322G>A | China | 1 woman and 178 controls | |
| Wang | REC114 | c.546 + 5G>A | China | 2 families | |
| Zheng | PADI6 | c.1521dupC, c.A1117C and c.C1708T | China | 2 women | |
| Quian | PADI6 | c.1793A>G and c.2045 G>A, | China | 1 family | |
| Wang 2018 | PADI6 | c.866C>T, c.1895C>T and c.1124dupT | China | 2 families and 80 infertile controls | |
| Xu | PADI6 | c.2009_2010del c.633T>A c.1618G>A c.970C>T | China | 2 out of 36 women | |
| Mu | NLRP2 | c.1961C>A, c.773T>C, c.2254C>T, c.525G>C, c.2544A>T, c.662C>T, c.1847A>T, c.1469C>T | China | 5 families | |
| Mu | NLRP5 | c.292C>T, c.2081C>T, c.866G>A, c.3320C>T | China | 2 families | |
| Wu | PATL2 | p.V260M, p.Q300*, p.T425P, p.D293Y p.N239Tfs*9 p.R75Vfs*21 | China | 5 families | |
| Wang 2018 | TLE6 | c.1133delC | China, however the family was from Ukraine | 1 family and 80 infertile controls | |
| Wang 2018 | KHDC3L | c.44delA | China | 1 family and 80 infertile controls | |
| Factor | Type | Effect | Possible Underlying Infertility Pathophysiology | Screening | |
|---|---|---|---|---|---|
| Cyclins, Cyclin-dependent kinases and their inhibitors (CCNA2, CDK1, CDK2, CDK4, p27) | Protein expression levels | Embryo | NP | Proteomic analysis | |
| DNMT3B, HDAC1, TET3 and YY1 | Gene variation | Embryo | NP | Genetic analysis | |
| ATG5, CUL3, USP11 and USP2 | Gene variation | Embryo | NP | Genetic analysis | |
| TERF1, ERCC1 and XRCC6 | Gene variation | Embryo | NP | Genetic analysis | |
| BRCA1 | Gene variation | Both | POR | Genetic analysis | |
| ITPR1 | Gene variation | Maternal | NP | Genetic analysis | |
| 4977-bp mtDNA deletion | Copy numbers | Parental | NP | Genetic analysis | |
| HO-1 | Gene variation | Parental | NP | Genetic analysis | |
| H19DMR | ICRs | Parental | NP | Transcriptomic analysis | |
| Nanog and Oct4 transcription factors | Protein expression levels | Embryo | NP | Methylome analysis | |
| piRNA | sncRNA | Embryo | NP | Transcriptomic analysis | |
| miRNA mainly of the let-7 family | sncRNA | Both | In females, miRNAs have been associated with endometriosis | Transcriptomic analysis | |
| In males, miRNAs have been associated with azoospermia and asthenozoospermia | |||||
| Aneuploidy, mosaicism | Chromosomal abnormalities | Embryo | Advanced maternal age | Genetic analysis | |
| Caspase-3 | Protein expression levels | Both | sperm DNA fragmentation | Proteomic analysis | |
| IL-6 | Protein expression levels | Embryo | NP | Proteomic analysis | |
| HRG | Protein expression levels | Embryo | NP | Proteomic analysis | |
| HDL and ApoA1 | Protein expression levels | Embryo | NP | Proteomic analysis | |
| ABCC8 | Gene expression levels | Embryo | NP | Transcriptomic analysis | |
| UBR4 | Gene expression levels | Embryo | NP | Transcriptomic analysis | |
| HERC2 | Gene expression levels | Embryo | NP | Transcriptomic analysis | |
| EMMPRIN | Gene expression levels | Embryo | NP | Transcriptomic analysis | |
| Oxygen consumption | Metabolic biomarker | Embryo | NP | Metabolomic analysis | |
| ROS | Metabolic biomarker | Both | ROS in the follicular fluid is related to poor oocyte quality and fertilization failure. | Metabolomic analysis | |
| Seminal ROS levels are associated with sperm DNA fragmentation. | |||||
| PADI6 | Gene variation | Maternal | oocyte activation failure | Genetic analysis in parents | |
| TUBB8 | Gene variation | Both | oocyte meiotic spindle assembly | Genetic analysis in parents | |
| NLRP7, TLE6, KHDC3, OOEP | Gene variation | Maternal | Formation of subcortical maternal complex | Genetic analysis | |
| NRLP5 | Gene variation | Maternal | Primary infertility | Genetic analysis | |
| PATL2 | Gene variation | Maternal | NP | Genetic analysis | |
| ZAR1 | Gene variation | Both | Affects MZT | Genetic analysis | |
| REC114 | Protein expression levels | Maternal | NP | Proteomic analysis in parents | |
| IGF-1, -2 and their respective receptors | Protein or mRNA expression levels | Maternal | POR | Proteomic or transcriptomic analysis in parents | |
| IGFBPs, and PAPP-A | Protein expression levels | Maternal | NP | Proteomic analysis in parents | |
| FSH, LH and their receptors | Gene variation or Expression levels | Parental | In females: follicular growth | In males: DNA fragmentation, azoospermia | Genetic or Proteomic analysis in parents |
| Activin a | Protein expression levels | Parental | Follicular growth | Proteomic analysis in parents | |
| WNT4 | Gene variation | Maternal | Endometriosis | Genetic analysis in parents | |
| FMR1, GDF-9 and NOBOX | Gene variation | Maternal | POR | Genetic analysis in parents | |
| BMP-15 | Gene variation | Both | POR and follicular growth | Genetic analysis in parents | |
| PAWP | Protein expression levels | Paternal | Oocyte activation | Proteomic analysis in parents | |
| AZF | Gene variation | Paternal | Azoospermia, oligozoospermia and oligoasthenoteratzoospermia | Genetic analysis in parents | |
| DPY19L2, SPATA16, PICK1 and GGN | Gene variation | Paternal | Globozoospermia | Genetic analysis in parents | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sfakianoudis, K.; Maziotis, E.; Karantzali, E.; Kokkini, G.; Grigoriadis, S.; Pantou, A.; Giannelou, P.; Petroutsou, K.; Markomichali, C.; Fakiridou, M.; et al. Molecular Drivers of Developmental Arrest in the Human Preimplantation Embryo: A Systematic Review and Critical Analysis Leading to Mapping Future Research. Int. J. Mol. Sci. 2021, 22, 8353. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22158353
Sfakianoudis K, Maziotis E, Karantzali E, Kokkini G, Grigoriadis S, Pantou A, Giannelou P, Petroutsou K, Markomichali C, Fakiridou M, et al. Molecular Drivers of Developmental Arrest in the Human Preimplantation Embryo: A Systematic Review and Critical Analysis Leading to Mapping Future Research. International Journal of Molecular Sciences. 2021; 22(15):8353. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22158353
Chicago/Turabian StyleSfakianoudis, Konstantinos, Evangelos Maziotis, Eleni Karantzali, Georgia Kokkini, Sokratis Grigoriadis, Amelia Pantou, Polina Giannelou, Konstantina Petroutsou, Christina Markomichali, Maria Fakiridou, and et al. 2021. "Molecular Drivers of Developmental Arrest in the Human Preimplantation Embryo: A Systematic Review and Critical Analysis Leading to Mapping Future Research" International Journal of Molecular Sciences 22, no. 15: 8353. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22158353