
G2-S16 Polyanionic Carbosilane Dendrimer Can Reduce HIV-1 Reservoir Formation by Inhibiting Macrophage Cell to Cell Transmission

 , and
, and 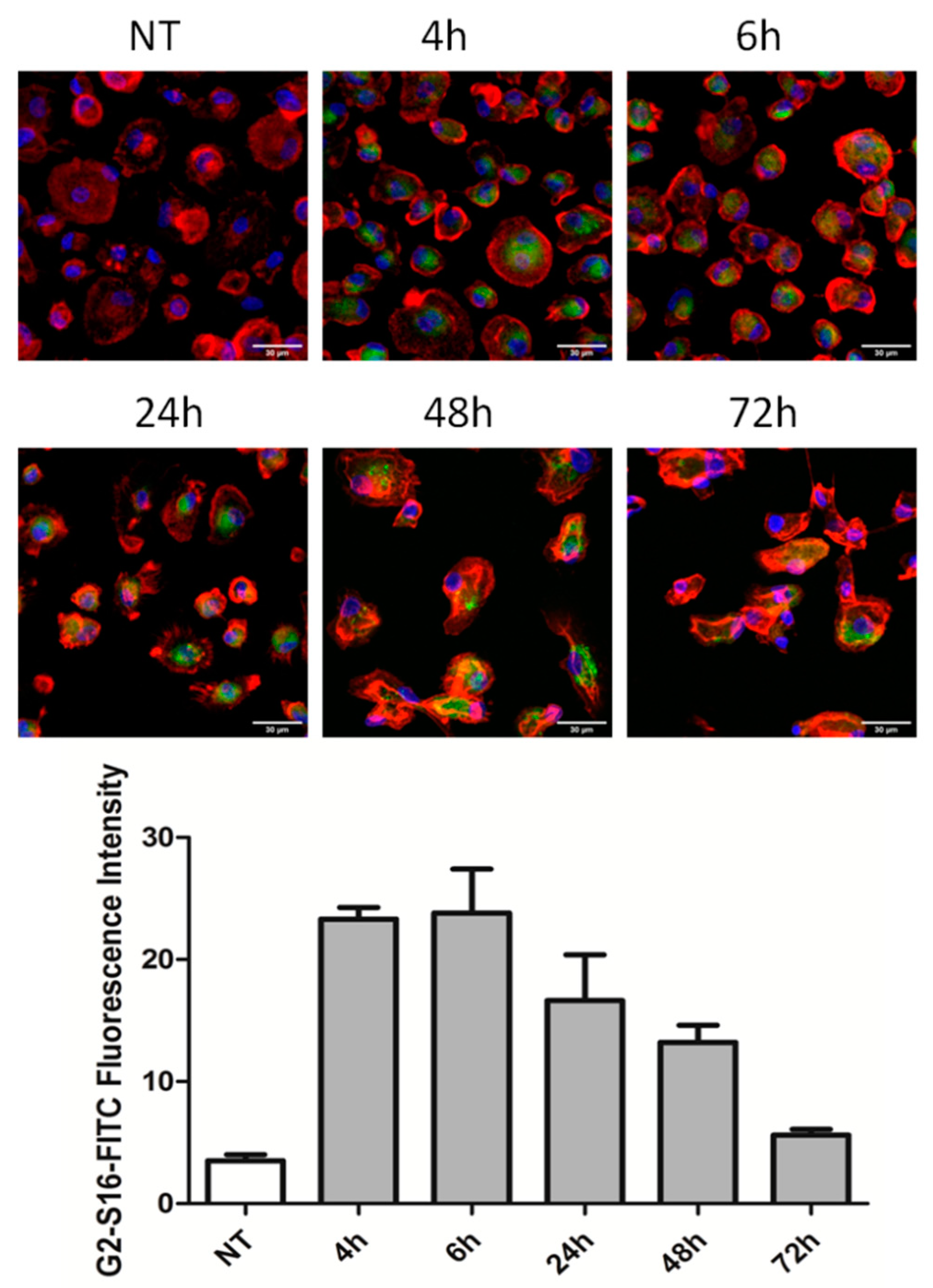{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
2.1. Biocompatibility of G2-S16 Dendrimer
2.2. G2-S16 Dendrimer Internalizes into Monocyte-Derived Macrophages and Is Stable for 48 h
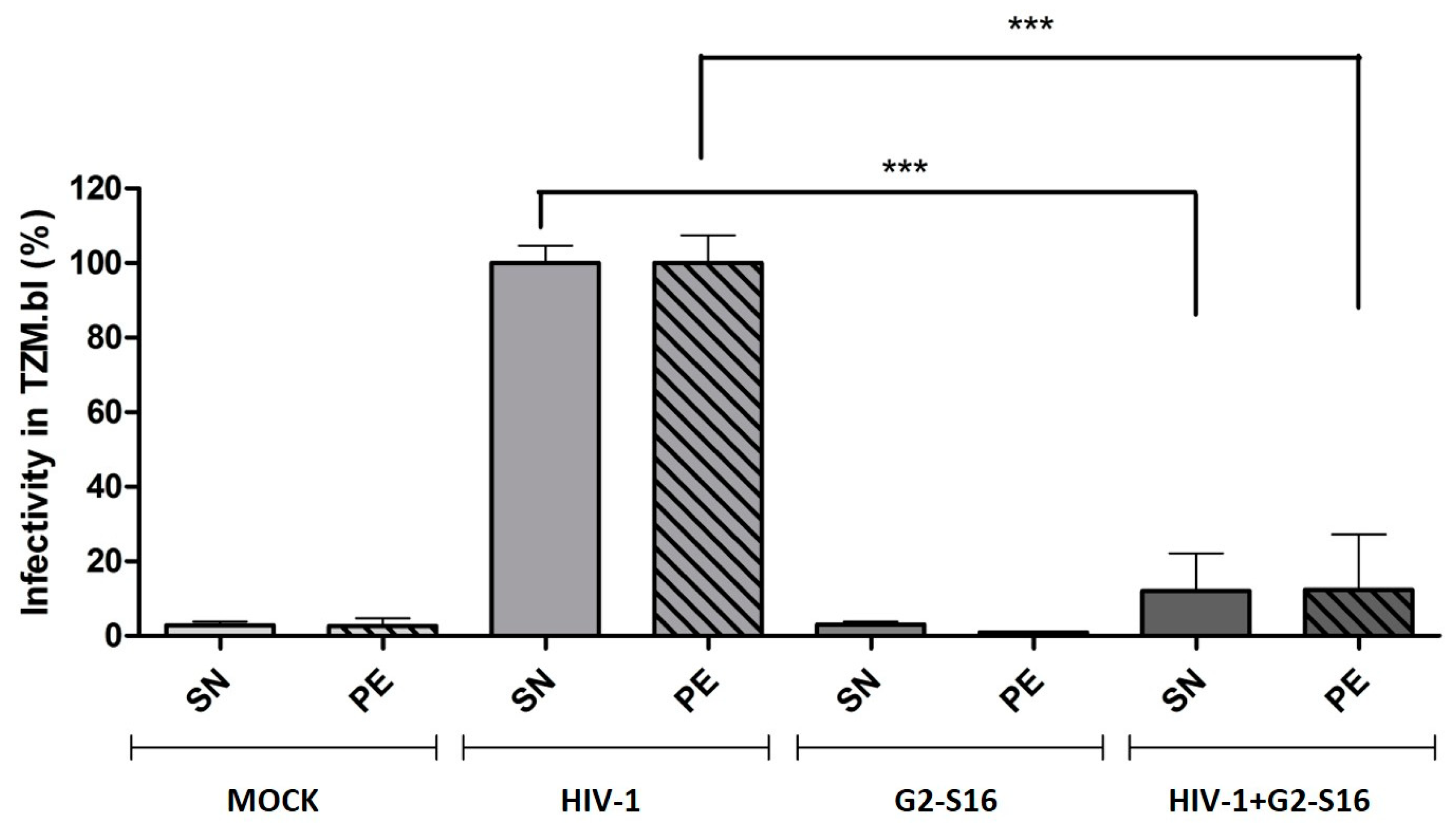
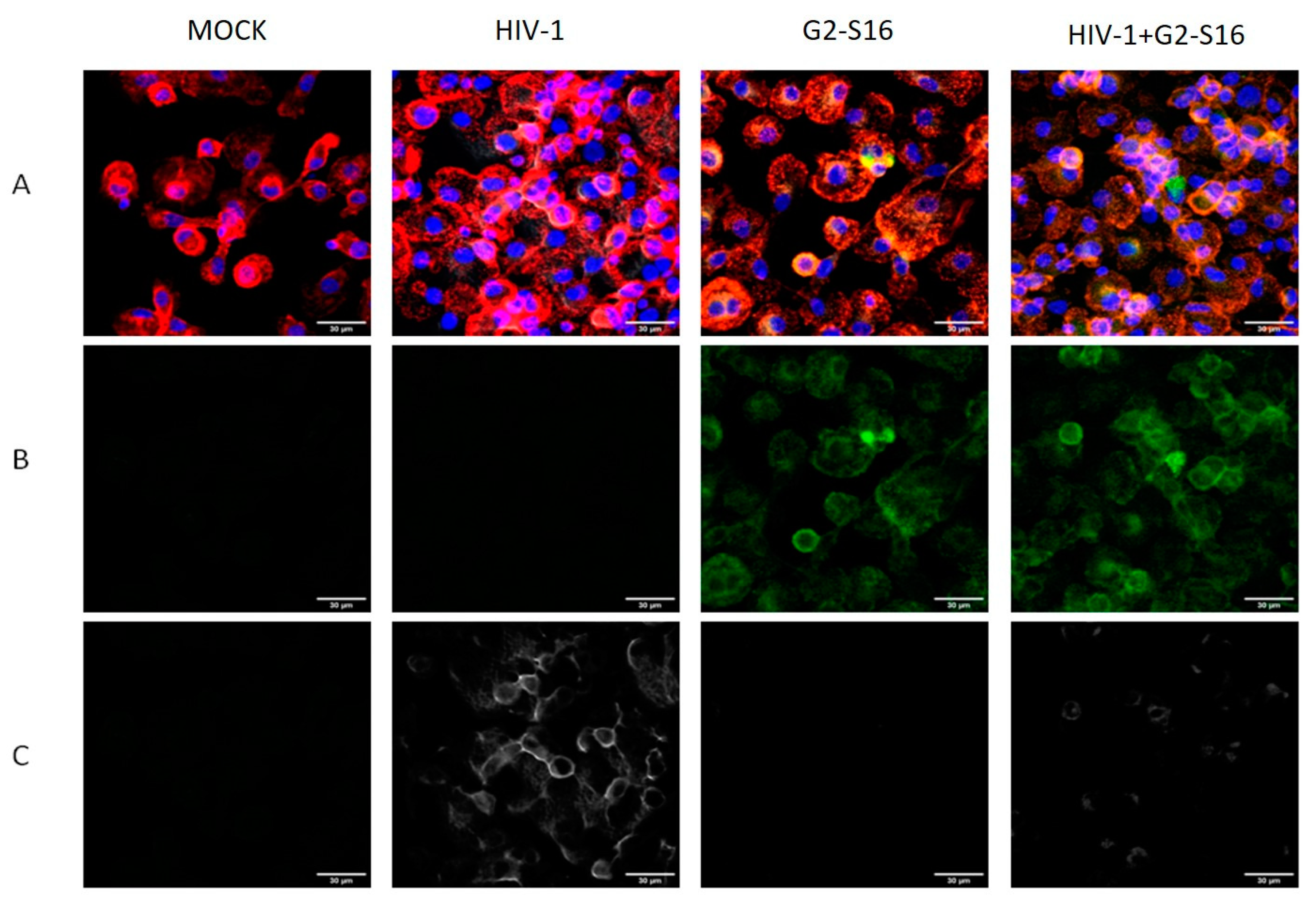
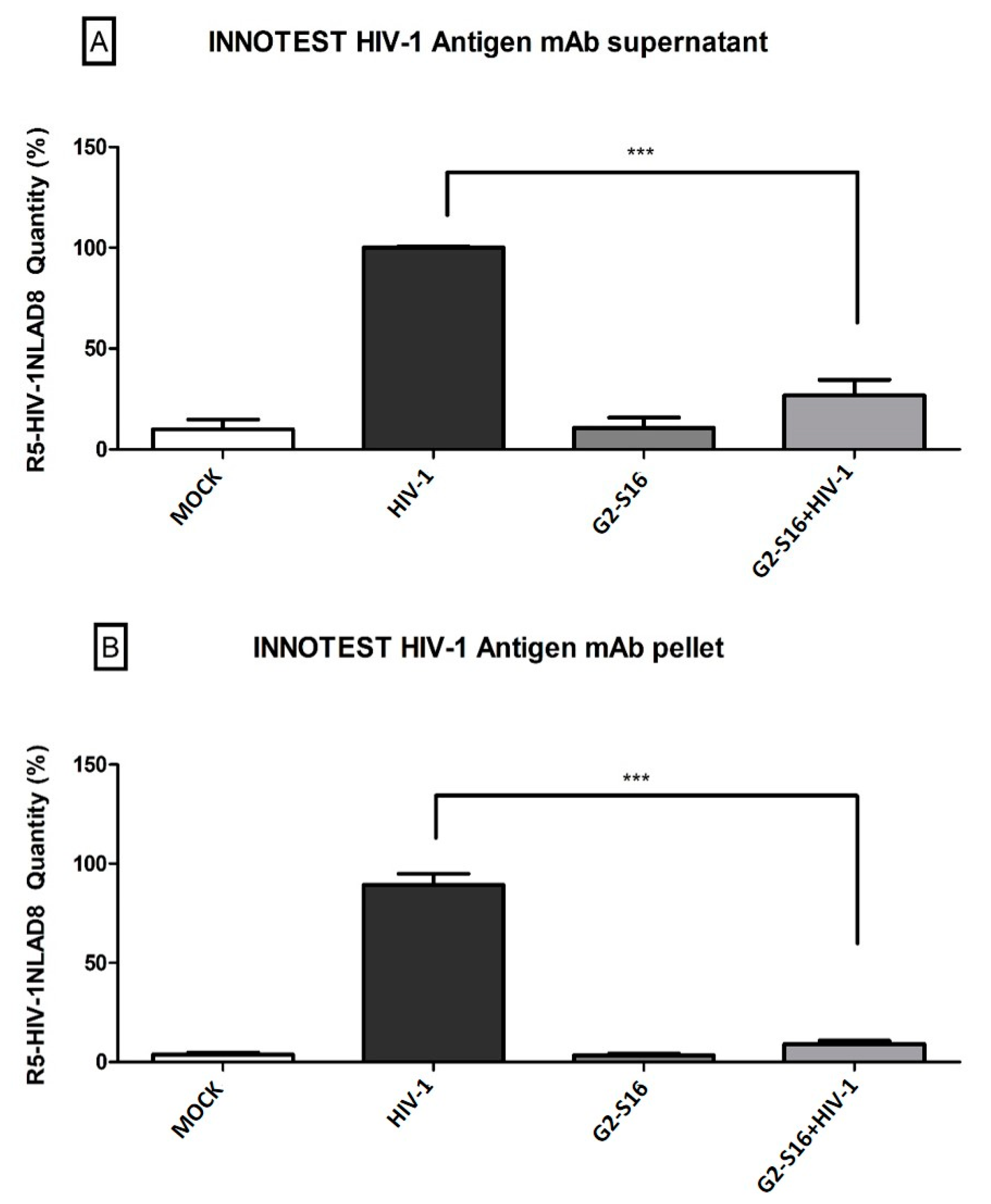
2.3. G2-S16 Dendrimer Internalized into Monocyte-Derived Macrophages Eliminates HIV-1 Infectivity
2.4. Infected Monocyte-Derived Macrophages Do Not Spread HIV-1 Infection to T CD4/CD8 Lymphocytes after G2-S16 Dendrimer Treatment
3. Materials and Methods
3.1. Reagents
3.2. Cell Culture and Viral Strains
3.3. Primary Cell Cultures, Purification, and Differentiation
3.4. Immuno Fluorescence and Confocal Images
3.5. Inhibition Assay
3.6. Inhibition Assay Cell–Cell
3.7. Infectivity of HIV-1
3.8. Determination of HIV-1 Quantity
3.9. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mahy, M.; Marsh, K.; Sabin, K.; Wanyeki, I.; Daher, J.; Ghys, P.D. Hiv estimates through 2018: Data for decision-making. AIDS 2019, 33 (Suppl. 3), S203–S211. [Google Scholar] [CrossRef]
- Roy, U.; Rodriguez, J.; Barber, P.; das Neves, J.; Sarmento, B.; Nair, M. The potential of hiv-1 nanotherapeutics: From in vitro studies to clinical trials. Nanomedicine 2015, 10, 3597–3609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Relano-Rodriguez, I.; Juarez-Sanchez, R.; Pavicic, C.; Munoz, E.; Munoz-Fernandez, M.A. Polyanionic carbosilane dendrimers as a new adjuvant in combination with latency reversal agents for hiv treatment. J. Nanobiotechnol. 2019, 17, 69. [Google Scholar] [CrossRef] [Green Version]
- Royo-Rubio, E.; Rodriguez-Izquierdo, I.; Moreno-Domene, M.; Lozano-Cruz, T.; de la Mata, F.J.; Gomez, R.; Munoz-Fernandez, M.A.; Jimenez, J.L. Promising pegylated cationic dendrimers for delivery of mirnas as a possible therapy against hiv-1 infection. J. Nanobiotechnol. 2021, 19, 158. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Izquierdo, I.; Natalia, C.; Garcia, F.; Los Angeles Munoz-Fernandez, M. G2-s16 sulfonate dendrimer as new therapy for treatment failure in hiv-1 entry inhibitors. Nanomedicine 2019, 14, 1095–1107. [Google Scholar] [CrossRef]
- Guerrero-Beltran, C.; Prieto, A.; Leal, M.; Jimenez, J.L.; Munoz-Fernandez, M.A. Combination of g2-s16 dendrimer/dapivirine antiretroviral as a new hiv-1 microbicide. Future Med. Chem. 2019, 11, 3005–3013. [Google Scholar] [CrossRef] [PubMed]
- Guerrero-Beltran, C.; Rodriguez-Izquierdo, I.; Serramia, M.J.; Araya-Duran, I.; Marquez-Miranda, V.; Gomez, R.; de la Mata, F.J.; Leal, M.; Gonzalez-Nilo, F.; Munoz-Fernandez, M.A. Anionic carbosilane dendrimers destabilize the gp120-cd4 complex blocking hiv-1 entry and cell to cell fusion. Bioconjug. Chem. 2018, 29, 1584–1594. [Google Scholar] [CrossRef] [PubMed]
- Sepulveda-Crespo, D.; Serramia, M.J.; Tager, A.M.; Vrbanac, V.; Gomez, R.; De La Mata, F.J.; Jimenez, J.L.; Munoz-Fernandez, M.A. Prevention vaginally of hiv-1 transmission in humanized blt mice and mode of antiviral action of polyanionic carbosilane dendrimer g2-s16. Nanomedicine 2015, 11, 1299–1308. [Google Scholar] [CrossRef] [PubMed]
- Relano-Rodriguez, I.; Munoz-Fernandez, M.A. Emergence of nanotechnology to fight hiv sexual transmission: The trip of g2-s16 polyanionic carbosilane dendrimer to possible pre-clinical trials. Int. J. Mol. Sci. 2020, 21, 9403. [Google Scholar] [CrossRef] [PubMed]
- Breton, Y.; Desrosiers, V.; Ouellet, M.; Deshiere, A.; Torresilla, C.; Cohen, E.A.; Tremblay, M.J. Expression of mdm2 in macrophages promotes the early postentry steps of hiv-1 infection through inhibition of p53. J. Virol. 2019, 93, e01871-18. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, I.; Porterfield, J.Z.; Gupta, R.K.; Mlcochova, P. Cell cycle regulation in macrophages and susceptibility to hiv-1. Viruses 2020, 12, 839. [Google Scholar] [CrossRef]
- Hughes, K.; Akturk, G.; Gnjatic, S.; Chen, B.; Klotman, M.; Blasi, M. Proliferation of hiv-infected renal epithelial cells following virus acquisition from infected macrophages. AIDS 2020, 34, 1581–1591. [Google Scholar] [CrossRef]
- Burdo, T.H.; Lackner, A.; Williams, K.C. Monocyte/macrophages and their role in hiv neuropathogenesis. Immunol. Rev. 2013, 254, 102–113. [Google Scholar] [CrossRef] [Green Version]
- Wallet, C.; De Rovere, M.; Van Assche, J.; Daouad, F.; De Wit, S.; Gautier, V.; Mallon, P.W.G.; Marcello, A.; Van Lint, C.; Rohr, O.; et al. Microglial cells: The main hiv-1 reservoir in the brain. Front. Cell. Infect. Microbiol. 2019, 9, 362. [Google Scholar] [CrossRef] [Green Version]
- Wong, M.E.; Jaworowski, A.; Hearps, A.C. The hiv reservoir in monocytes and macrophages. Front. Immunol. 2019, 10, 1435. [Google Scholar] [CrossRef] [Green Version]
- Bayliss, R.; Wheeldon, J.; Caucheteux, S.M.; Niessen, C.M.; Piguet, V. Identification of host trafficking genes required for hiv-1 virological synapse formation in dendritic cells. J. Virol. 2020, 94, e01597-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez, P.; Koh, W.H.; Hnatiuk, R.; Murooka, T.T. Hiv infection stabilizes macrophage-t cell interactions to promote cell-cell hiv spread. J. Virol. 2019, 93, e00805-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dupont, M.; Sattentau, Q.J. Macrophage cell-cell interactions promoting hiv-1 infection. Viruses 2020, 12, 492. [Google Scholar] [CrossRef] [PubMed]
- Woodham, A.W.; Skeate, J.G.; Sanna, A.M.; Taylor, J.R.; Da Silva, D.M.; Cannon, P.M.; Kast, W.M. Human immunodeficiency virus immune cell receptors, coreceptors, and cofactors: Implications for prevention and treatment. AIDS Patient Care STDS 2016, 30, 291–306. [Google Scholar] [CrossRef] [Green Version]
- Bracq, L.; Xie, M.; Benichou, S.; Bouchet, J. Mechanisms for cell-to-cell transmission of hiv-1. Front. Immunol. 2018, 9, 260. [Google Scholar] [CrossRef] [PubMed]
- Cena-Diez, R.; Garcia-Broncano, P.; Javier de la Mata, F.; Gomez, R.; Resino, S.; Munoz-Fernandez, M. G2-s16 dendrimer as a candidate for a microbicide to prevent hiv-1 infection in women. Nanoscale 2017, 9, 9732–9742. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Rodriguez, J.; Diaz, L.; Galan, M.; Maly, M.; Gomez, R.; Javier de la Mata, F.; Jimenez, J.L.; Munoz-Fernandez, M.A. Anti-human immunodeficiency virus activity of thiol-ene carbosilane dendrimers and their potential development as a topical microbicide. J. Biomed. Nanotechnol. 2015, 11, 1783–1798. [Google Scholar] [CrossRef]
- Sanchez-Rodriguez, J.; Vacas-Cordoba, E.; Gomez, R.; De La Mata, F.J.; Munoz-Fernandez, M.A. Nanotech-derived topical microbicides for hiv prevention: The road to clinical development. Antiviral Res. 2015, 113, 33–48. [Google Scholar] [CrossRef]
- Abreu, C.; Shirk, E.N.; Queen, S.E.; Beck, S.E.; Mangus, L.M.; Pate, K.A.M.; Mankowski, J.L.; Gama, L.; Clements, J.E. Brain macrophages harbor latent, infectious simian immunodeficiency virus. AIDS 2019, 33 (Suppl. 2), S181–S188. [Google Scholar] [CrossRef] [PubMed]
- Honeycutt, J.B.; Wahl, A.; Baker, C.; Spagnuolo, R.A.; Foster, J.; Zakharova, O.; Wietgrefe, S.; Caro-Vegas, C.; Madden, V.; Sharpe, G.; et al. Macrophages sustain hiv replication in vivo independently of t cells. J. Clin. Investig. 2016, 126, 1353–1366. [Google Scholar] [CrossRef] [PubMed]
- Akiyama, H.; Gummuluru, S. Hiv-1 persistence and chronic induction of innate immune responses in macrophages. Viruses 2020, 12, 711. [Google Scholar] [CrossRef]
- Duncan, C.J.; Williams, J.P.; Schiffner, T.; Gartner, K.; Ochsenbauer, C.; Kappes, J.; Russell, R.A.; Frater, J.; Sattentau, Q.J. High-multiplicity hiv-1 infection and neutralizing antibody evasion mediated by the macrophage-t cell virological synapse. J. Virol. 2014, 88, 2025–2034. [Google Scholar] [CrossRef] [Green Version]
- Bayliss, R.J.; Piguet, V. Masters of manipulation: Viral modulation of the immunological synapse. Cell. Microbiol. 2018, 20, e12944. [Google Scholar] [CrossRef] [Green Version]
- Vargas-Garcia, C.; Zurakowski, R.; Singh, A. Synaptic transmission may provide an evolutionary benefit to hiv through modulation of latency. J. Theor. Biol. 2018, 455, 261–268. [Google Scholar] [CrossRef]
- Vacas-Cordoba, E.; Maly, M.; De la Mata, F.J.; Gomez, R.; Pion, M.; Munoz-Fernandez, M.A. Antiviral mechanism of polyanionic carbosilane dendrimers against hiv-1. Int. J. Nanomed. 2016, 11, 1281–1294. [Google Scholar]
- Gutiérrez-Ulloa, C.; Peña-González, C.E.; Barrios-Gumiel, A.; Ceña-Díez, R.; Serramía-Lobera, M.J.; Muñoz-Fernández, M.Á.; Javier de la Mata, F.; Sánchez-Nieves, J.; Gómez, R. New synthetic procedure for the antiviral sulfonate carbosilane dendrimer g2-s16 and its fluorescein-labelled derivative for biological studies. RSC Adv. 2020, 10, 20083–20088. [Google Scholar] [CrossRef]
- Consuegra, I.; Rodriguez-Aierbe, C.; Santiuste, I.; Bosch, A.; Martinez-Marin, R.; Fortuto, M.A.; Diaz, T.; Marti, S.; Munoz-Fernandez, M.A. Isolation methods of peripheral blood mononuclear cells in spanish biobanks: An overview. Biopreserv. Biobank. 2017, 15, 305–309. [Google Scholar] [CrossRef] [PubMed]
- Martin-Moreno, A.; Sepulveda-Crespo, D.; Serramia-Lobera, M.J.; Perise-Barrios, A.J.; Munoz-Fernandez, M.A. G2-s16 dendrimer microbicide does not interfere with the vaginal immune system. J. Nanobiotechnology 2019, 17, 65. [Google Scholar] [CrossRef] [PubMed]
- Cena-Diez, R.; Martin-Moreno, A.; de la Mata, F.J.; Gomez-Ramirez, R.; Munoz, E.; Ardoy, M.; Munoz-Fernandez, M.A. G1-s4 or g2-s16 carbosilane dendrimer in combination with platycodin d as a promising vaginal microbicide candidate with contraceptive activity. Int. J. Nanomed. 2019, 14, 2371–2381. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-Izquierdo, I.; Gasco, S.; Munoz-Fernandez, M.A. High preventive effect of g2-s16 anionic carbosilane dendrimer against sexually transmitted hsv-2 infection. Molecules 2020, 25, 2965. [Google Scholar] [CrossRef]
- Guerrero-Beltran, C.; Garcia-Heredia, I.; Cena-Diez, R.; Rodriguez-Izquierdo, I.; Serramia, M.J.; Martinez-Hernandez, F.; Lluesma-Gomez, M.; Martinez-Garcia, M.; Munoz-Fernandez, M.A. Cationic dendrimer g2-s16 inhibits herpes simplex type 2 infection and protects mice vaginal microbiome. Pharmaceutics 2020, 12, 515. [Google Scholar] [CrossRef]
- Cena-Diez, R.; Garcia-Broncano, P.; de la Mata, F.J.; Gomez, R.; Munoz-Fernandez, M.A. Efficacy of hiv antiviral polyanionic carbosilane dendrimer g2-s16 in the presence of semen. Int. J. Nanomed. 2016, 11, 2443–2450. [Google Scholar]
- Sepulveda-Crespo, D.; Jimenez, J.L.; Gomez, R.; De La Mata, F.J.; Majano, P.L.; Munoz-Fernandez, M.A.; Gastaminza, P. Polyanionic carbosilane dendrimers prevent hepatitis c virus infection in cell culture. Nanomedicine 2017, 13, 49–58. [Google Scholar] [CrossRef]
- Dutartre, H.; Claviere, M.; Journo, C.; Mahieux, R. Cell-free versus cell-to-cell infection by human immunodeficiency virus type 1 and human t-lymphotropic virus type 1: Exploring the link among viral source, viral trafficking, and viral replication. J. Virol. 2016, 90, 7607–7617. [Google Scholar] [CrossRef] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Relaño-Rodríguez, I.; Espinar-Buitrago, M.d.l.S.; Martín-Cañadilla, V.; Gómez-Ramírez, R.; Muñoz-Fernández, M.Á. G2-S16 Polyanionic Carbosilane Dendrimer Can Reduce HIV-1 Reservoir Formation by Inhibiting Macrophage Cell to Cell Transmission. Int. J. Mol. Sci. 2021, 22, 8366. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22168366
Relaño-Rodríguez I, Espinar-Buitrago MdlS, Martín-Cañadilla V, Gómez-Ramírez R, Muñoz-Fernández MÁ. G2-S16 Polyanionic Carbosilane Dendrimer Can Reduce HIV-1 Reservoir Formation by Inhibiting Macrophage Cell to Cell Transmission. International Journal of Molecular Sciences. 2021; 22(16):8366. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22168366
Chicago/Turabian StyleRelaño-Rodríguez, Ignacio, María de la Sierra Espinar-Buitrago, Vanessa Martín-Cañadilla, Rafael Gómez-Ramírez, and María Ángeles Muñoz-Fernández. 2021. "G2-S16 Polyanionic Carbosilane Dendrimer Can Reduce HIV-1 Reservoir Formation by Inhibiting Macrophage Cell to Cell Transmission" International Journal of Molecular Sciences 22, no. 16: 8366. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22168366