
LncRNA MIR31HG Drives Oncogenicity by Inhibiting the Limb-Bud and Heart Development Gene (LBH) during Oral Carcinoma

 and
and 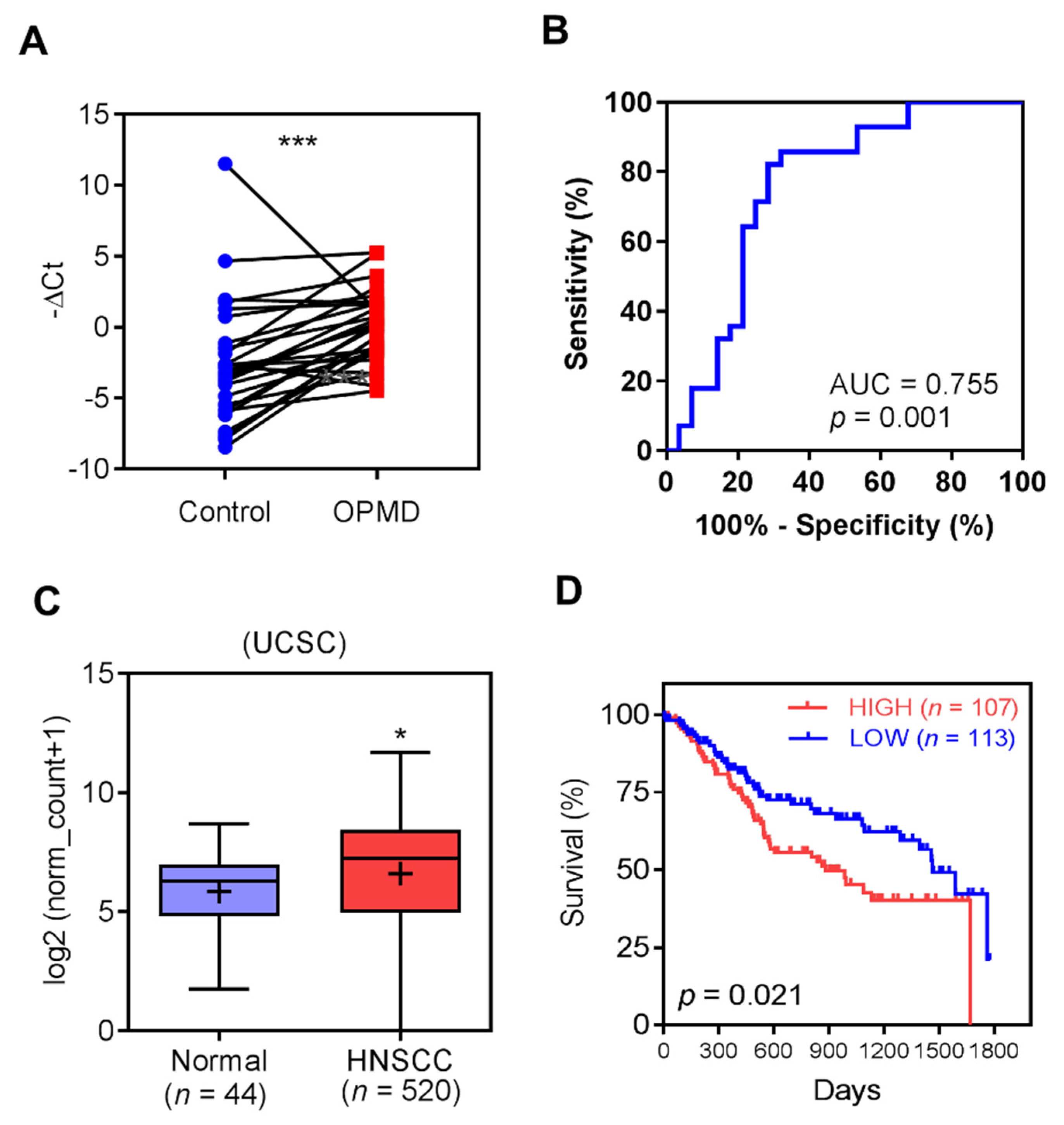{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Up-Regulation of MIR31HG in OPMD and OSCC
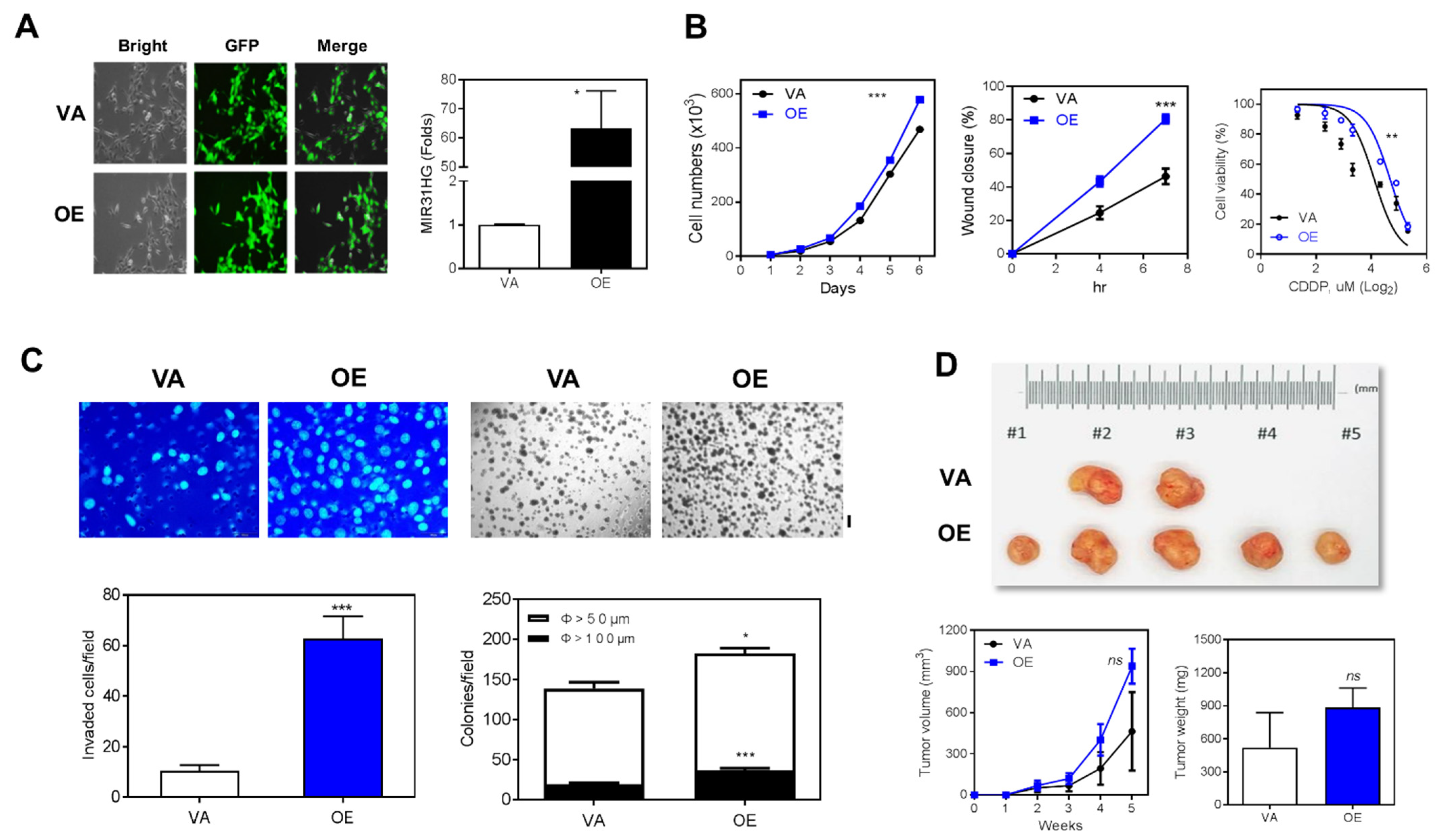
2.2. Overexpression of MIR31HG Enhances Various Oncogenic Phenotypes
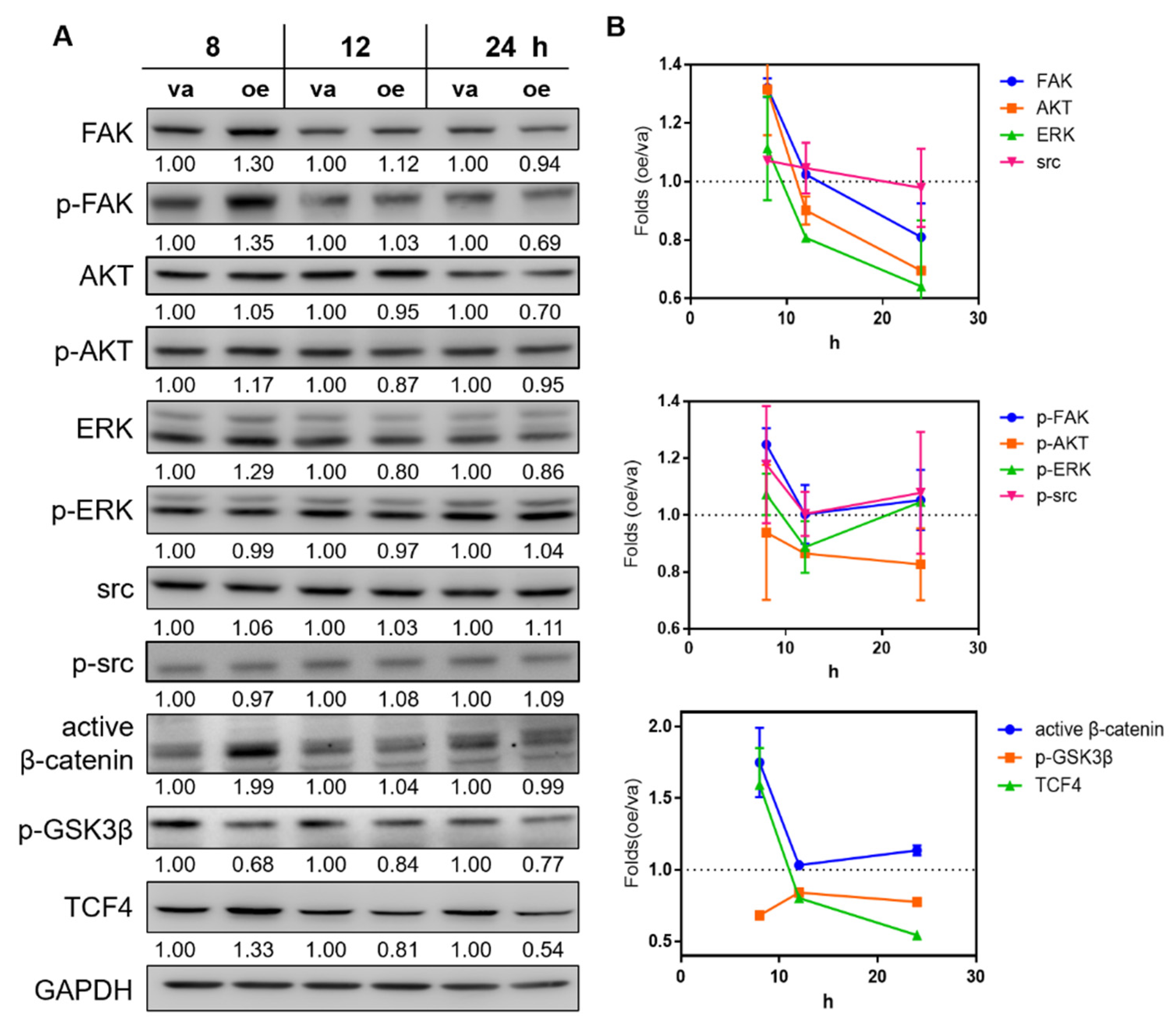
2.3. Overexpression of MIR31HG Activates the Wnt Pathway
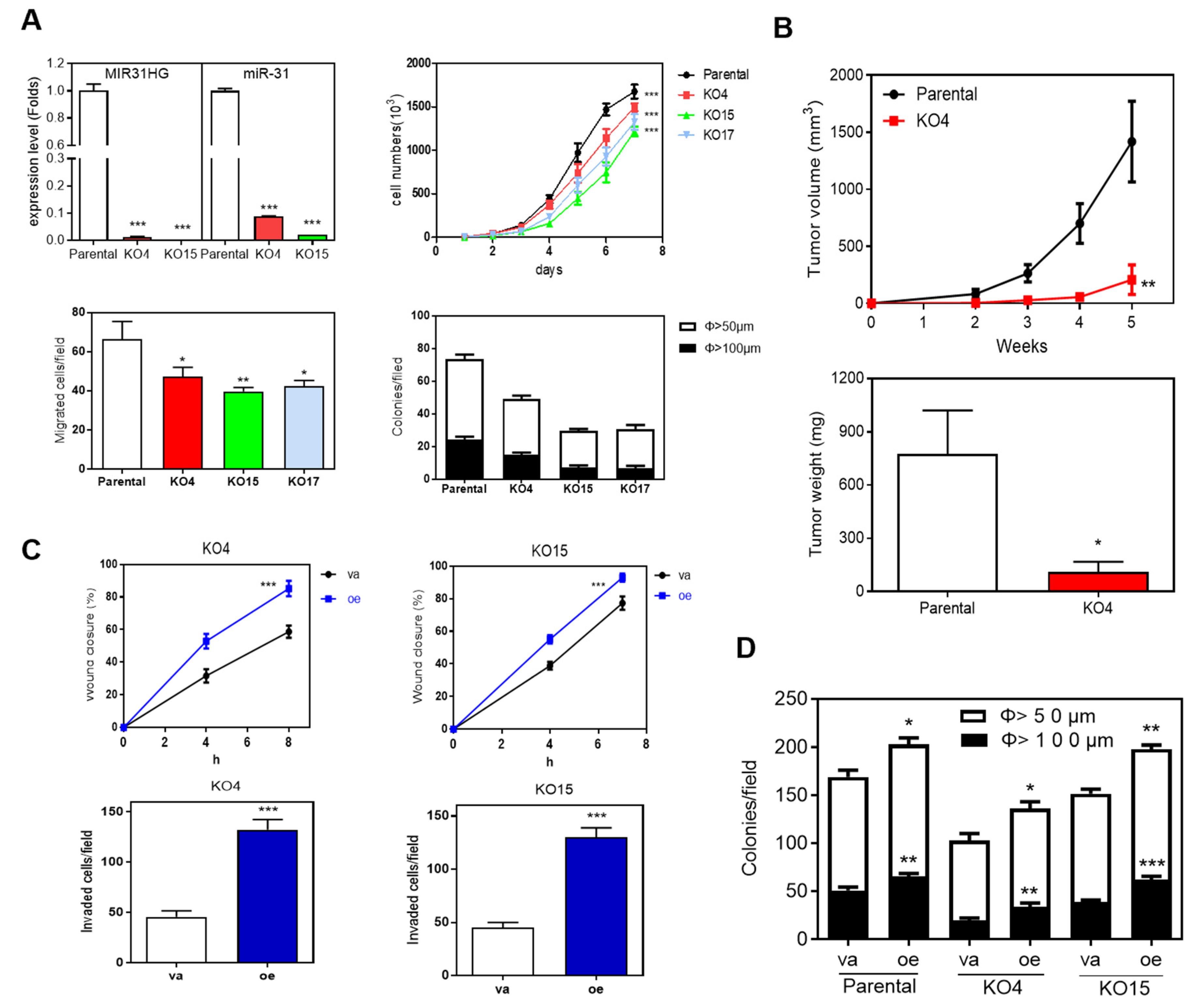
2.4. Deletion of MIR31HG Reduces Oncogenic Phenotypes
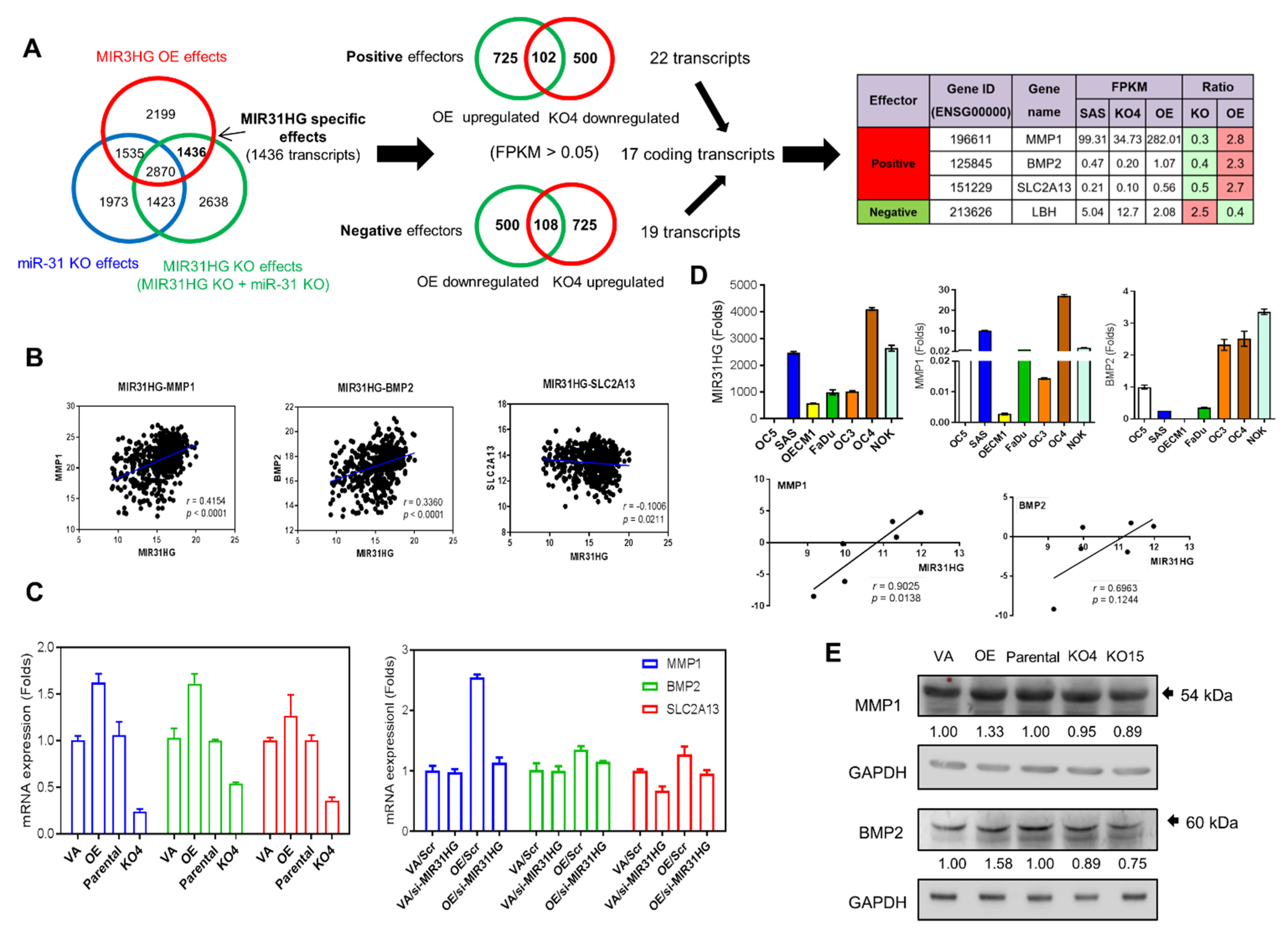
2.5. Identification of Downstream Effectors of MIR31HG
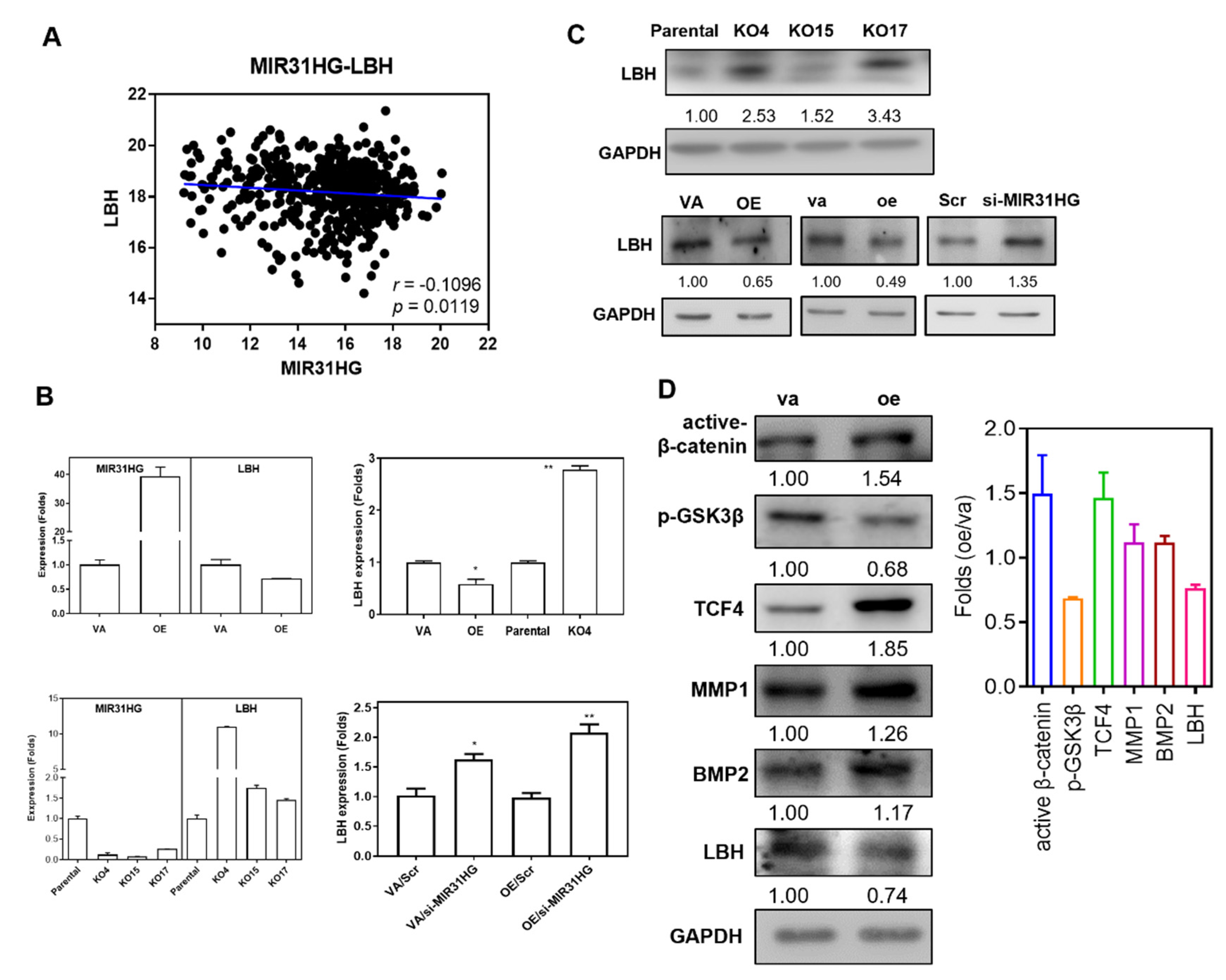
2.6. LBH Is a Negative Effector of MIR31HG
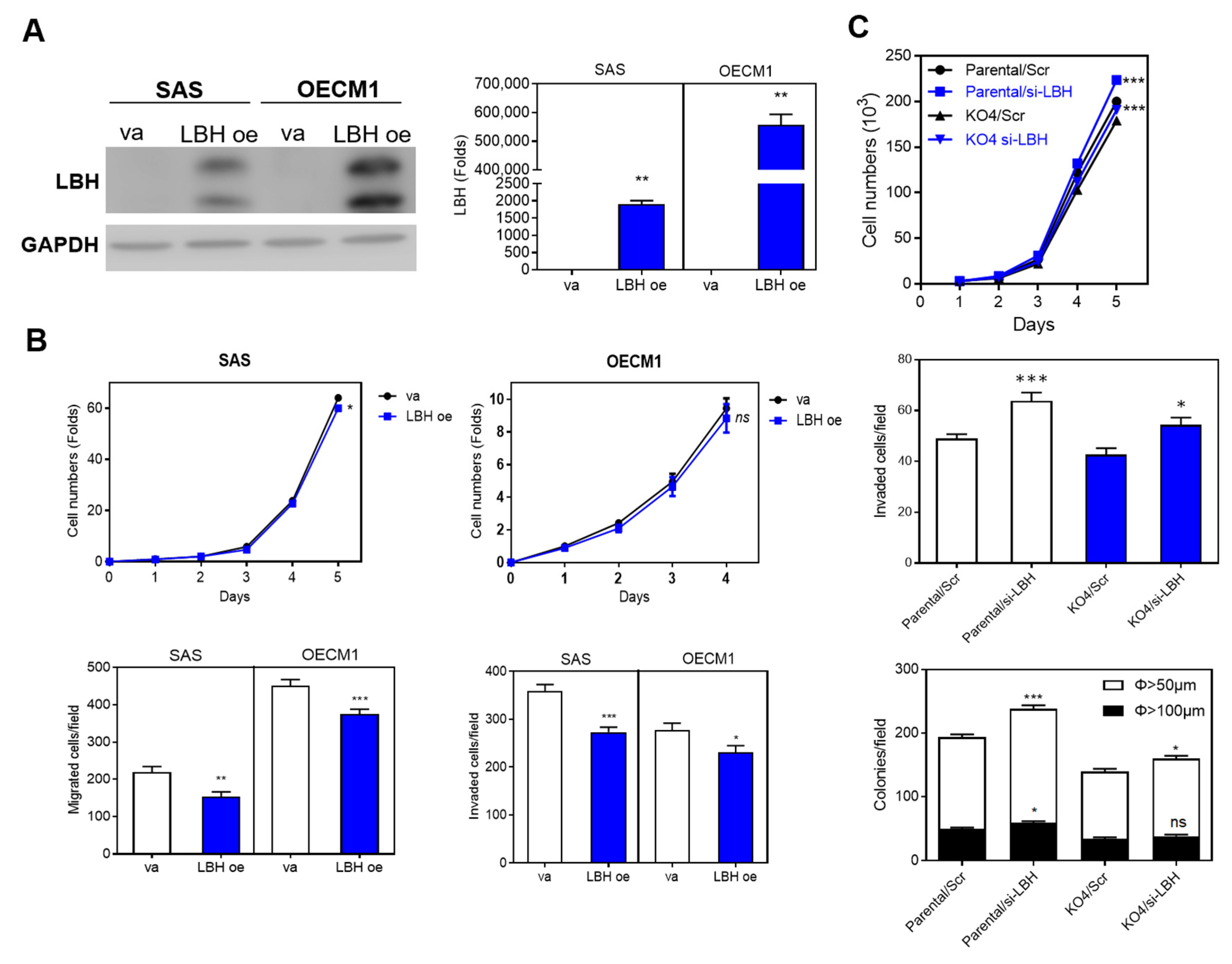
2.7. The MIR31HG Associated Phenotypes Are Attenuated by LBH
3. Discussion
4. Materials and Methods
4.1. Subjects
4.2. Cell Lines
4.3. qRT-PCR Analysis
4.4. Western Blotting
4.5. Plasmid Construction and Overexpression
4.6. The Implement of Clustered Regularly Interspaced Short Palindromic Repeat (Crispr)/Cas9 Approach for MIR31HG Deletion
4.7. Phenotypic and Tumorigenic Assays
4.8. RNASeq
4.9. Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| CRISPR | Clustered: regularly interspaced, short palindromic repeats |
| HNSCC | Head and neck squamous cell carcinoma |
| LBH | Limb-Bud and Heart development |
| LncRNA | Long non-coding RNA |
| NSCLC | Non-small cell lung cancer |
| OPMD | Oral potentially malignant disorder |
| OSCC | Oral squamous cell carcinoma |
| TCGA | The Cancer Genome Atlas |
References
- Liu, C.J.; Tsai, M.M.; Hung, P.S.; Kao, S.Y.; Liu, T.Y.; Wu, K.J.; Chiou, S.H.; Lin, S.C.; Chang, K.W. miR-31 ablates expression of the HIF regulatory factor FIH to activate the HIF pathway in head and neck carcinoma. Cancer Res. 2010, 70, 1635–1644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, S.Y.; Tu, H.F.; Yang, C.C.; Wu, C.H.; Liu, C.J.; Chang, K.W.; Lin, S.C. miR-134 targets PDCD7 to reduce E-cadherin expression and enhance oral cancer progression. Int. J. Cancer 2018, 143, 2892–2904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inchingolo, F.; Santacroce, L.; Ballini, A.; Topi, S.; Dipalma, G.; Haxhirexha, K.; Bottalico, L.; Charitos, I.A. Oral cancer: A historical review. Int. J. Environ. Res. Public Health 2020, 17, 3168. [Google Scholar] [CrossRef]
- Sasahira, T.; Kirita, T. Hallmarks of cancer-related newly prognostic factors of oral squamous cell carcinoma. Int. J. Mol. Sci. 2018, 19, 2413. [Google Scholar] [CrossRef] [Green Version]
- Siriwardena, S.; Tsunematsu, T.; Qi, G.; Ishimaru, N.; Kudo, Y. Invasion-related factors as potential diagnostic and therapeutic targets in oral squamous cell carcinoma-a review. Int. J. Mol. Sci. 2018, 19, 1462. [Google Scholar] [CrossRef] [Green Version]
- Dionne, K.R.; Warnakulasuriya, S.; Zain, R.B.; Cheong, S.C. Potentially malignant disorders of the oral cavity: Current practice and future directions in the clinic and laboratory. Int. J. Cancer 2015, 136, 503–515. [Google Scholar] [CrossRef]
- Hanna, R.; Dalvi, S.; Benedicenti, S.; Amaroli, A.; Salagean, T.; Pop, I.D.; Todea, D.; Bordea, I.R. Photobiomodulation therapy in oral mucositis and potentially malignant oral lesions: A therapy towards the future. Cancers 2020, 12, 1949. [Google Scholar] [CrossRef]
- Makunin, I.V.; Mattick, J.S. Non-coding RNA. Hum. Mol. Genet. 2006, 15, R17–R29. [Google Scholar]
- Kao, Y.Y.; Chou, C.H.; Yeh, L.Y.; Chen, Y.F.; Chang, K.W.; Liu, C.J.; Fan Chiang, C.Y.; Lin, S.C. MicroRNA miR-31 targets SIRT3 to disrupt mitochondrial activity and increase oxidative stress in oral carcinoma. Cancer Lett. 2019, 456, 40–48. [Google Scholar] [CrossRef] [PubMed]
- Lu, W.C.; Liu, C.J.; Tu, H.F.; Chung, Y.T.; Yang, C.C.; Kao, S.Y.; Chang, K.W.; Lin, S.C. miR-31 targets ARID1A and enhances the oncogenicity and stemness of head and neck squamous cell carcinoma. Oncotarget 2016, 7, 57254–57267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tseng, S.H.; Yang, C.C.; Yu, E.H.; Chang, C.; Lee, Y.S.; Liu, C.J.; Chang, K.W.; Lin, S.C. K14-EGFP-miR-31 transgenic mice have high susceptibility to chemical-induced squamous cell tumorigenesis that is associating with Ku80 repression. Int. J. Cancer 2015, 136, 1263–1275. [Google Scholar] [CrossRef] [PubMed]
- Hung, K.F.; Liu, C.J.; Chiu, P.C.; Lin, J.S.; Chang, K.W.; Shih, W.Y.; Kao, S.Y.; Tu, H.F. MicroRNA-31 upregulation predicts increased risk of progression of oral potentially malignant disorder. Oral Oncol. 2016, 53, 42–47. [Google Scholar] [CrossRef]
- Hung, P.S.; Tu, H.F.; Kao, S.Y.; Yang, C.C.; Liu, C.J.; Huang, T.Y.; Chang, K.W.; Lin, S.C. miR-31 is upregulated in oral premalignant epithelium and contributes to the immortalization of normal oral keratinocytes. Carcinogenesis 2014, 35, 1162–1171. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.J.; Kao, S.Y.; Tu, H.F.; Tsai, M.M.; Chang, K.W.; Lin, S.C. Increase of microRNA miR-31 level in plasma could be a potential marker of oral cancer. Oral Dis. 2010, 16, 360–364. [Google Scholar] [CrossRef]
- Liu, C.J.; Lin, S.C.; Yang, C.C.; Cheng, H.W.; Chang, K.W. Exploiting salivary miR-31 as a clinical biomarker of oral squamous cell carcinoma. Head Neck 2012, 34, 219–224. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.A.; Shah, N.; Wang, K.C.; Kim, J.; Horlings, H.M.; Wong, D.J.; Tsai, M.C.; Hung, T.; Argani, P.; Rinn, J.L.; et al. Long non-coding RNA HOTAIR reprograms chromatin state to promote cancer metastasis. Nature 2010, 464, 1071–1076. [Google Scholar] [CrossRef]
- Montes, M.; Lubas, M.; Arendrup, F.S.; Mentz, B.; Rohatgi, N.; Tumas, S.; Harder, L.M.; Skanderup, A.J.; Andersen, J.S.; Lund, A.H. The long non-coding RNA MIR31HG regulates the senescence associated secretory phenotype. Nat. Commun. 2021, 12, 2459. [Google Scholar] [CrossRef] [PubMed]
- Montes, M.; Nielsen, M.M.; Maglieri, G.; Jacobsen, A.; Hojfeldt, J.; Agrawal-Singh, S.; Hansen, K.; Helin, K.; van de Werken, H.J.G.; Pedersen, J.S.; et al. The lncRNA MIR31HG regulates p16(INK4A) expression to modulate senescence. Nat. Commun. 2015, 6, 6967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shih, J.W.; Chiang, W.F.; Wu, A.T.H.; Wu, M.H.; Wang, L.Y.; Yu, Y.L.; Hung, Y.W.; Wang, W.C.; Chu, C.Y.; Hung, C.L.; et al. Long noncoding RNA LncHIFCAR/MIR31HG is a HIF-1alpha co-activator driving oral cancer progression. Nat. Commun. 2017, 8, 15874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rybicki, B.A.; Savera, A.T.; Gomez, J.A.; Patel, S.C.; Ballard, N.E.; Benninger, M.S.; Zarbo, R.J.; Van Dyke, D.L. Allelic loss and tumor pathology in head and neck squamous cell carcinoma. Mod. Pathol. 2003, 16, 970–979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, P.; Li, H.; Huo, W.; Zhu, H.; Xu, C.; Zang, R.; Lv, W.; Xia, Y.; Tang, W. Aberrant expression of LncRNA-MIR31HG regulates cell migration and proliferation by affecting miR-31 and miR-31* in Hirschsprung’s disease. J. Cell Biochem. 2018, 119, 8195–8203. [Google Scholar] [CrossRef] [PubMed]
- Chu, J.; Jia, J.; Yang, L.; Qu, Y.; Yin, H.; Wan, J.; He, F. LncRNA MIR31HG functions as a ceRNA to regulate c-Met function by sponging miR-34a in esophageal squamous cell carcinoma. Biomed. Pharm. 2020, 128, 110313. [Google Scholar] [CrossRef] [PubMed]
- Dandan, W.; Jianliang, C.; Haiyan, H.; Hang, M.; Xuedong, L. Long noncoding RNA MIR31HG is activated by SP1 and promotes cell migration and invasion by sponging miR-214 in NSCLC. Gene 2019, 692, 223–230. [Google Scholar] [CrossRef] [PubMed]
- Eide, P.W.; Eilertsen, I.A.; Sveen, A.; Lothe, R.A. Long noncoding RNA MIR31HG is a bona fide prognostic marker with colorectal cancer cell-intrinsic properties. Int. J. Cancer 2019, 144, 2843–2853. [Google Scholar] [CrossRef] [Green Version]
- He, A.; Chen, Z.; Mei, H.; Liu, Y. Decreased expression of LncRNA MIR31HG in human bladder cancer. Cancer Biomark. 2016, 17, 231–236. [Google Scholar] [CrossRef]
- Li, Y. MIR31HG exhibits oncogenic property and acts as a sponge for miR-361-3p in cervical carcinoma. Biochem. Biophys Res. Commun. 2020, 529, 890–897. [Google Scholar] [CrossRef]
- Lin, Y.; Zhang, C.S.; Li, S.J.; Li, Z.; Sun, F.B. LncRNA LOC554202 promotes proliferation and migration of gastric cancer cells through regulating p21 and E-cadherin. Eur. Rev. Med. Pharm. Sci. 2018, 22, 8690–8697. [Google Scholar]
- Nie, F.Q.; Ma, S.; Xie, M.; Liu, Y.W.; De, W.; Liu, X.H. Decreased long noncoding RNA MIR31HG is correlated with poor prognosis and contributes to cell proliferation in gastric cancer. Tumour. Biol. 2016, 37, 7693–7701. [Google Scholar] [CrossRef]
- Qin, J.; Ning, H.; Zhou, Y.; Hu, Y.; Yang, L.; Huang, R. LncRNA MIR31HG overexpression serves as poor prognostic biomarker and promotes cells proliferation in lung adenocarcinoma. Biomed. Pharm. 2018, 99, 363–368. [Google Scholar] [CrossRef]
- Rajbhandari, R.; McFarland, B.C.; Patel, A.; Gerigk, M.; Gray, G.K.; Fehling, S.C.; Bredel, M.; Berbari, N.F.; Kim, H.; Marks, M.P.; et al. Loss of tumor suppressive microRNA-31 enhances TRADD/NF-kappaB signaling in glioblastoma. Oncotarget 2015, 6, 17805–17816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, K.; Zhao, X.; Wan, J.; Yang, L.; Chu, J.; Dong, S.; Yin, H.; Ming, L.; He, F. The diagnostic value of long non-coding RNA MIR31HG and its role in esophageal squamous cell carcinoma. Life Sci. 2018, 202, 124–130. [Google Scholar] [CrossRef] [PubMed]
- Yan, S.; Tang, Z.; Chen, K.; Liu, Y.; Yu, G.; Chen, Q.; Dang, H.; Chen, F.; Ling, J.; Zhu, L.; et al. Long noncoding RNA MIR31HG inhibits hepatocellular carcinoma proliferation and metastasis by sponging microRNA-575 to modulate ST7L expression. J. Exp. Clin. Cancer Res. 2018, 37, 214. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Liu, P.; Zhang, J.; Peng, X.; Lu, Z.; Yu, S.; Meng, Y.; Tong, W.M.; Chen, J. Long noncoding RNA MIR31HG exhibits oncogenic property in pancreatic ductal adenocarcinoma and is negatively regulated by miR-193b. Oncogene 2015, 35, 3647. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Jiang, H.; Wang, L.; Chen, X.; Wu, K.; Zhang, S.; Ma, S.; Xia, B. Increased MIR31HG lncRNA expression increases gefitinib resistance in non-small cell lung cancer cell lines through the EGFR/PI3K/AKT signaling pathway. Oncol. Lett. 2017, 13, 3494–3500. [Google Scholar] [CrossRef] [Green Version]
- Zheng, S.; Zhang, X.; Wang, X.; Li, J. MIR31HG promotes cell proliferation and invasion by activating the Wnt/β-catenin signaling pathway in non-small cell lung cancer. Oncol. Lett. 2019, 17, 221–229. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.; Ma, Z.; Feng, L.; Yang, Y.; Tan, C.; Shi, Q.; Lian, M.; He, S.; Ma, H.; Fang, J. LncRNA MIR31HG targets HIF1A and P21 to facilitate head and neck cancer cell proliferation and tumorigenesis by promoting cell-cycle progression. Mol. Cancer 2018, 17, 162. [Google Scholar] [CrossRef]
- Augoff, K.; McCue, B.; Plow, E.F.; Sossey-Alaoui, K. miR-31 and its host gene lncRNA LOC554202 are regulated by promoter hypermethylation in triple-negative breast cancer. Mol. Cancer 2012, 11, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Briegel, K.J.; Joyner, A.L. Identification and characterization of Lbh, a novel conserved nuclear protein expressed during early limb and heart development. Dev. Biol. 2001, 233, 291–304. [Google Scholar] [CrossRef] [PubMed]
- Lindley, L.E.; Curtis, K.M.; Sanchez-Mejias, A.; Rieger, M.E.; Robbins, D.J.; Briegel, K.J. The WNT-controlled transcriptional regulator LBH is required for mammary stem cell expansion and maintenance of the basal lineage. Development 2015, 142, 893–904. [Google Scholar] [CrossRef] [Green Version]
- Rieger, M.E.; Sims, A.H.; Coats, E.R.; Clarke, R.B.; Briegel, K.J. The embryonic transcription cofactor LBH is a direct target of the Wnt signaling pathway in epithelial development and in aggressive basal subtype breast cancers. Mol. Cell Biol 2010, 30, 4267–4279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashad-Bishop, K.; Garikapati, K.; Lindley, L.E.; Jorda, M.; Briegel, K.J. Loss of Limb-Bud-and-Heart (LBH) attenuates mammary hyperplasia and tumor development in MMTV-Wnt1 transgenic mice. Biochem. Biophys. Res. Commun. 2019, 508, 536–542. [Google Scholar] [CrossRef]
- Deng, M.; Yu, R.; Wang, S.; Zhang, Y.; Li, Z.; Song, H.; Liu, B.; Xu, L.; Wang, X.; Zhang, Z.; et al. Limb-Bud and Heart attenuates growth and invasion of human lung adenocarcinoma cells and predicts survival outcome. Cell Physiol. Biochem. 2018, 47, 223–234. [Google Scholar] [CrossRef]
- Liu, Q.; Guan, X.; Lv, J.; Li, X.; Wang, Y.; Li, L. Limb-bud and Heart (LBH) functions as a tumor suppressor of nasopharyngeal carcinoma by inducing G1/S cell cycle arrest. Sci. Rep. 2015, 5, 7626. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Li, E.; Huang, L.; Cheng, M.; Li, L. Limb-bud and Heart overexpression inhibits the proliferation and migration of PC3M cells. J. Cancer 2018, 9, 424–432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, A.; Zhang, L.; Luo, N.; Zhang, L.; Li, L.; Liu, Q. Limb-bud and heart (LBH) inhibits cellular migration, invasion and epithelial-mesenchymal transition in nasopharyngeal carcinoma via downregulating alphaB-crystallin expression. Cell Signal. 2021, 85, 110045. [Google Scholar] [CrossRef] [PubMed]
- Reis, P.P.; Tokar, T.; Goswami, R.S.; Xuan, Y.; Sukhai, M.; Seneda, A.L.; Moz, L.E.S.; Perez-Ordonez, B.; Simpson, C.; Goldstein, D.; et al. A 4-gene signature from histologically normal surgical margins predicts local recurrence in patients with oral carcinoma: Clinical validation. Sci. Rep. 2020, 10, 1713. [Google Scholar] [CrossRef] [PubMed]
- Sand, J.P.; Kokorina, N.A.; Zakharkin, S.O.; Lewis, J.S., Jr.; Nussenbaum, B. BMP-2 expression correlates with local failure in head and neck squamous cell carcinoma. Otolaryngol. Head Neck Surg. 2014, 150, 245–250. [Google Scholar] [CrossRef]
- Cloitre, A.; Rosa, R.W.; Arrive, E.; Fricain, J.C. Outcome of CO2 laser vaporization for oral potentially malignant disorders treatment. Med. Oral Patol. Oral Cir. Bucal. 2018, 23, e237–e247. [Google Scholar] [CrossRef]
- Tang, H.; Wu, Z.; Zhang, J.; Su, B. Salivary lncRNA as a potential marker for oral squamous cell carcinoma diagnosis. Mol. Med. Rep. 2013, 7, 761–766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, W.C.; Kao, S.Y.; Yang, C.C.; Tu, H.F.; Wu, C.H.; Chang, K.W.; Lin, S.C. EGF up-regulates miR-31 through the C/EBPbeta signal cascade in oral carcinoma. PLoS ONE 2014, 9, e108049. [Google Scholar]
- Wu, S.; Nitschke, K.; Worst, T.S.; Fierek, A.; Weis, C.A.; Eckstein, M.; Porubsky, S.; Kriegmair, M.; Erben, P. Long noncoding RNA MIR31HG and its splice variants regulate proliferation and migration: Prognostic implications for muscle invasive bladder cancer. J. Exp. Clin. Cancer Res. 2020, 39, 288. [Google Scholar] [CrossRef]
- Wang, Q.; Gong, Y.; He, Y.; Xin, Y.; Lv, N.; Du, X.; Li, Y.; Jeong, B.R.; Xu, J. Genome engineering of Nannochloropsis with hundred-kilobase fragment deletions by Cas9 cleavages. Plant J. 2021, 106, 1148–1162. [Google Scholar] [CrossRef] [PubMed]
- Harismendy, O.; Notani, D.; Song, X.; Rahim, N.G.; Tanasa, B.; Heintzman, N.; Ren, B.; Fu, X.D.; Topol, E.J.; Rosenfeld, M.G.; et al. 9p21 DNA variants associated with coronary artery disease impair interferon-gamma signalling response. Nature 2011, 470, 264–268. [Google Scholar] [CrossRef] [Green Version]
- Khattab, H.M.; Kubota, S.; Takigawa, M.; Kuboki, T.; Sebald, W. The BMP-2 mutant L51P: A BMP receptor IA binding-deficient inhibitor of noggin. J. Bone Min. Metab. 2019, 37, 199–205. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.C.; Liu, C.J.; Ko, S.Y.; Chang, H.C.; Liu, T.Y.; Chang, K.W. Copy number amplification of 3q26–27 oncogenes in microdissected oral squamous cell carcinoma and oral brushed samples from areca chewers. J. Pathol. 2005, 206, 417–422. [Google Scholar] [CrossRef]
- Trapnell, C.; Roberts, A.; Goff, L.; Pertea, G.; Kim, D.; Kelley, D.R.; Pimentel, H.; Salzberg, S.L.; Rinn, J.L.; Pachter, L. Differential gene and transcript expression analysis of RNA-seq experiments with TopHat and Cufflinks. Nat. Protoc. 2012, 7, 562–578. [Google Scholar] [CrossRef] [PubMed] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chang, K.-W.; Hung, W.-W.; Chou, C.-H.; Tu, H.-F.; Chang, S.-R.; Liu, Y.-C.; Liu, C.-J.; Lin, S.-C. LncRNA MIR31HG Drives Oncogenicity by Inhibiting the Limb-Bud and Heart Development Gene (LBH) during Oral Carcinoma. Int. J. Mol. Sci. 2021, 22, 8383. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22168383
Chang K-W, Hung W-W, Chou C-H, Tu H-F, Chang S-R, Liu Y-C, Liu C-J, Lin S-C. LncRNA MIR31HG Drives Oncogenicity by Inhibiting the Limb-Bud and Heart Development Gene (LBH) during Oral Carcinoma. International Journal of Molecular Sciences. 2021; 22(16):8383. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22168383
Chicago/Turabian StyleChang, Kuo-Wei, Wan-Wen Hung, Chung-Hsien Chou, Hsi-Feng Tu, Shi-Rou Chang, Ying-Chieh Liu, Chung-Ji Liu, and Shu-Chun Lin. 2021. "LncRNA MIR31HG Drives Oncogenicity by Inhibiting the Limb-Bud and Heart Development Gene (LBH) during Oral Carcinoma" International Journal of Molecular Sciences 22, no. 16: 8383. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22168383