
Modular and Molecular Optimization of a LOV (Light–Oxygen–Voltage)-Based Optogenetic Switch in Yeast

 and
and
Abstract
:1. Introduction
2. Results and Discussion
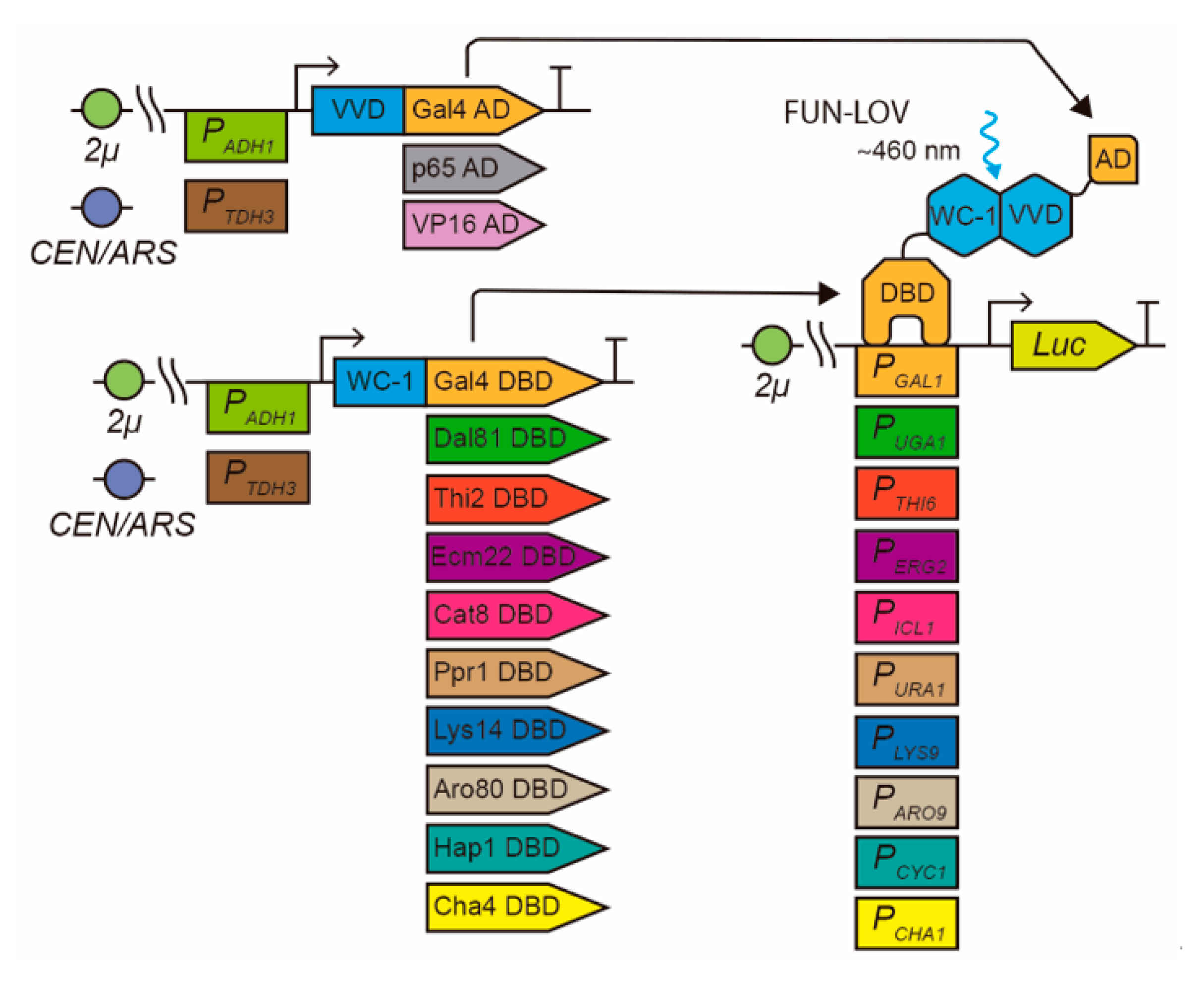
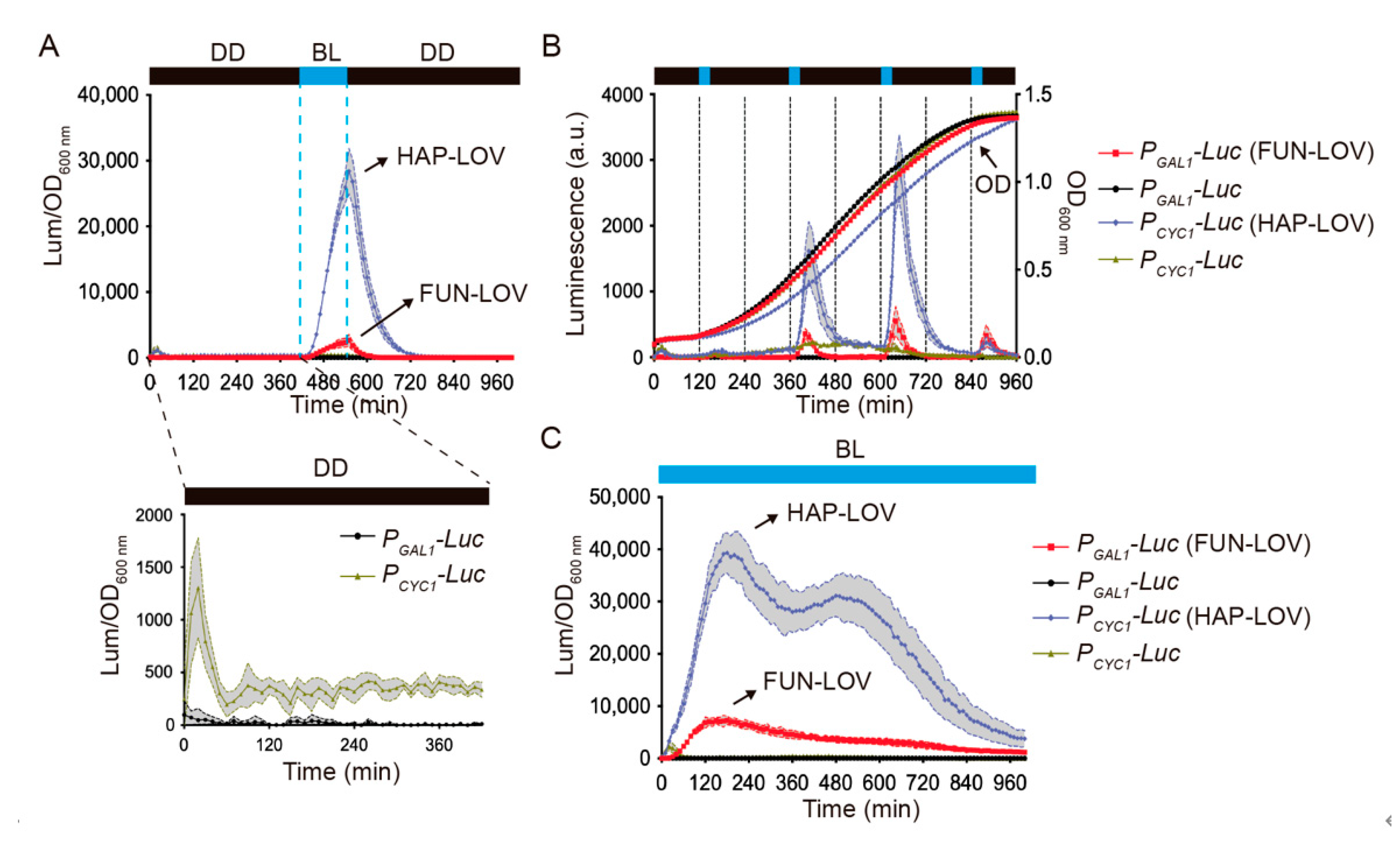
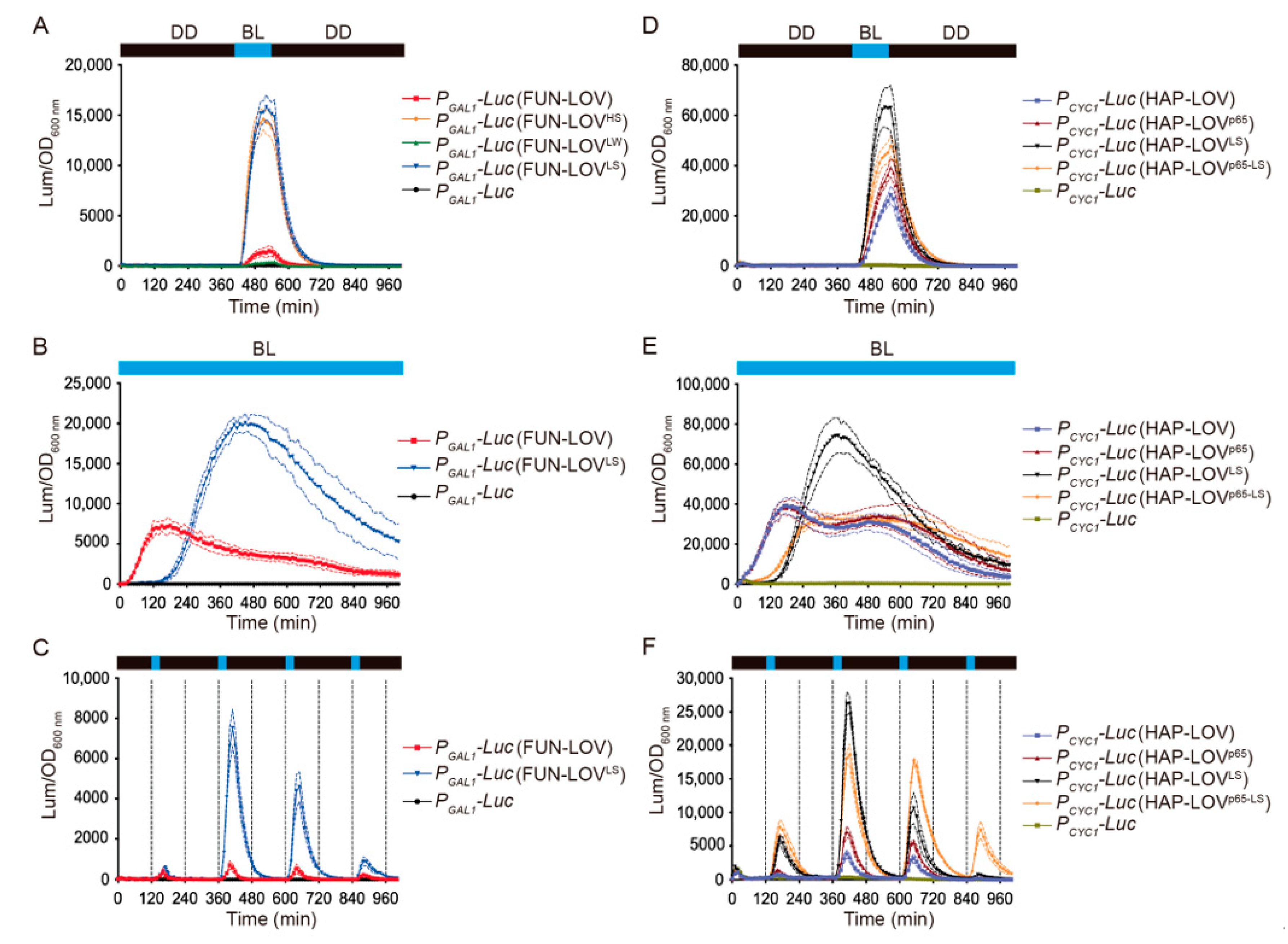
2.1. Modular Optimization of the FUN-LOV Optogenetic Switch
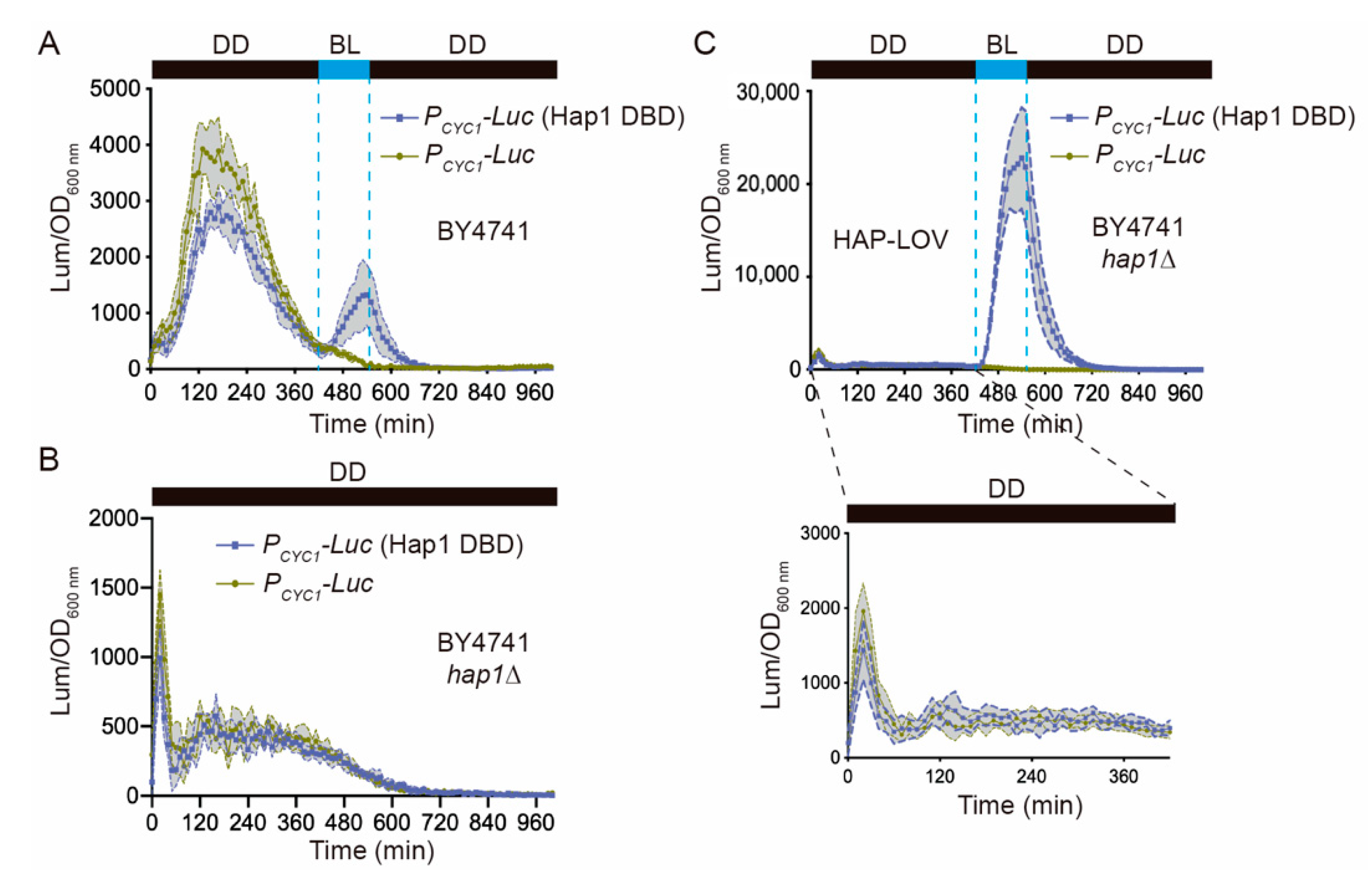
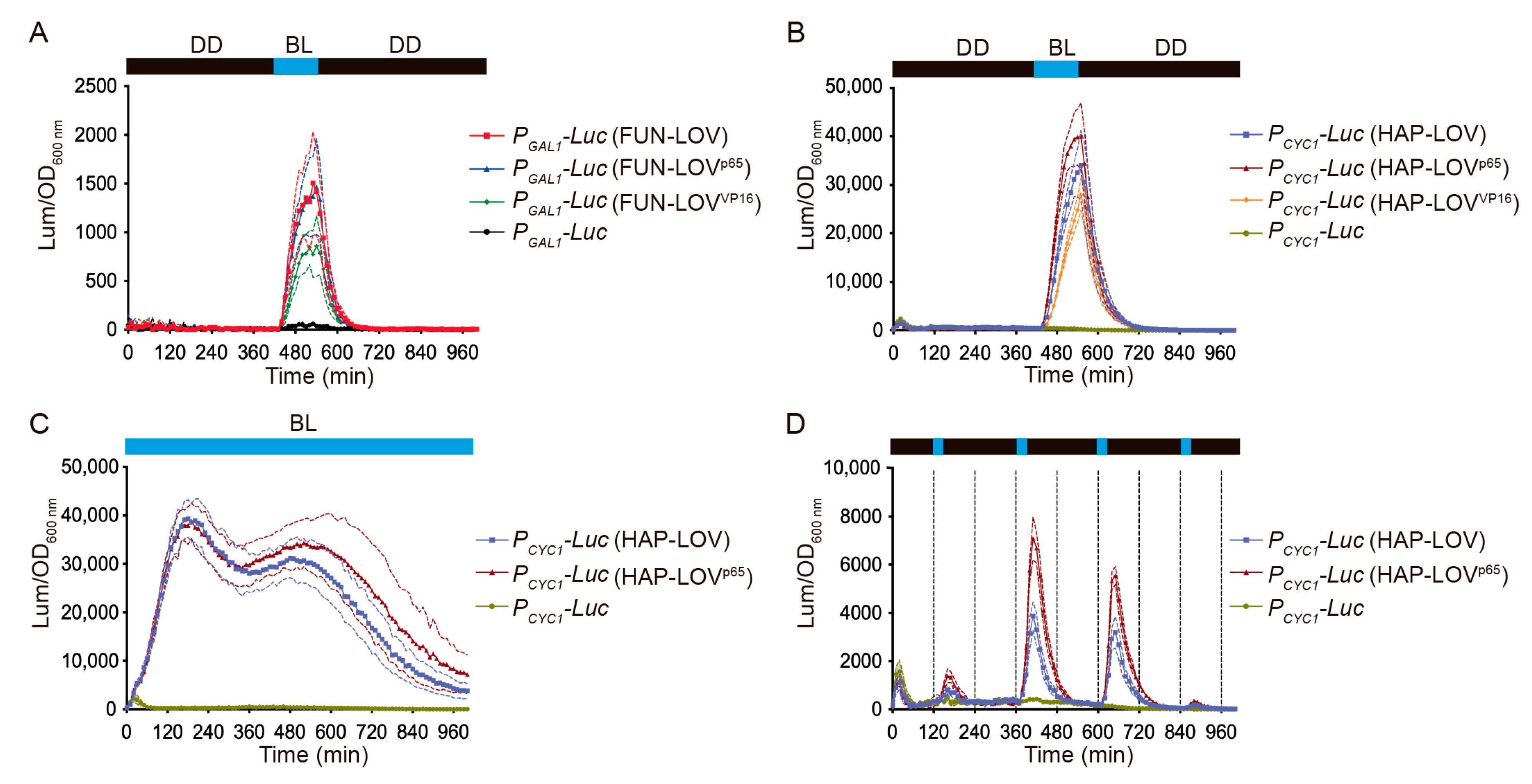
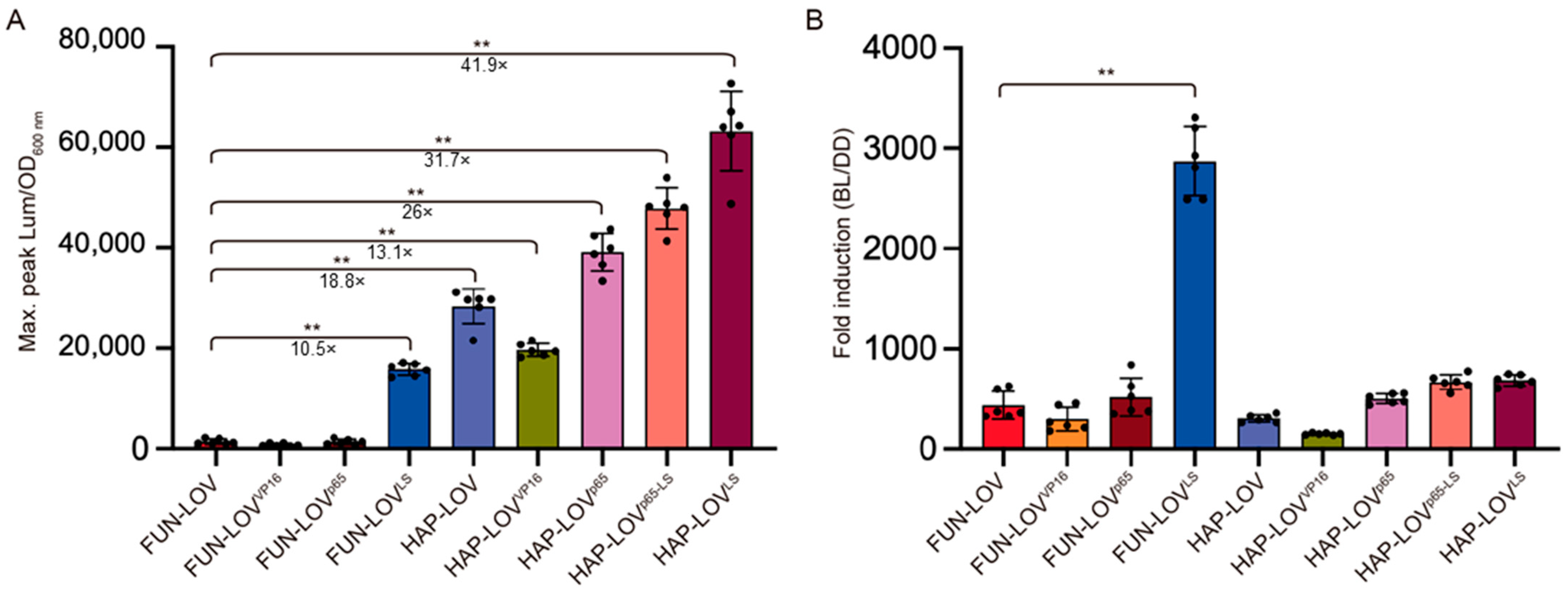
2.2. Molecular Optimization of the FUN-LOV and HAP-LOV Switches
2.3. Comparing Different Optogenetic Systems
3. Materials and Methods
3.1. Yeast Strains, Medium, and Culture Conditions
3.2. Illumination Conditions
3.3. Molecular Cloning and Strain Construction
3.4. In Vivo Luciferase Expression
3.5. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Boyden, E.S.; Zhang, F.; Bamberg, E.; Nagel, G.; Deisseroth, K. Millisecond-timescale, genetically targeted optical control of neural activity. Nat. Neurosci. 2005, 8, 1263–1268. [Google Scholar] [CrossRef] [PubMed]
- Carrasco-Lopez, C.; Garcia-Echauri, S.A.; Kichuk, T.; Avalos, J.L. Optogenetics and biosensors set the stage for metabolic cybergenetics. Curr. Opin. Biotechnol. 2020, 65, 296–309. [Google Scholar] [CrossRef] [PubMed]
- Spiltoir, J.I.; Tucker, C.L. Photodimerization systems for regulating protein–protein interactions with light. Curr. Opin. Struct. Biol. 2019, 57, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Figueroa, D.; Rojas, V.; Romero, A.; Larrondo, L.F.; Salinas, F. The rise and shine of yeast optogenetics. Yeast 2020. [Google Scholar] [CrossRef] [PubMed]
- Salinas, F.; Rojas, V.; Delgado, V.; Agosin, E.; Larrondo, L.F. Optogenetic switches for light-controlled gene expression in yeast. Appl. Microbiol. Biotechnol. 2017, 101, 2629–2640. [Google Scholar] [CrossRef]
- Renicke, C.; Schuster, D.; Usherenko, S.; Essen, L.-O.; Taxis, C. A LOV2 Domain-based optogenetic tool to control protein degradation and cellular function. Chem. Biol. 2013, 20, 619–626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delacour, Q.; Li, C.; Plamont, M.-A.; Billon-Denis, E.; Aujard, I.; Le Saux, T.; Jullien, L.; Gautier, A. Light-activated proteolysis for the spatiotemporal control of proteins. ACS Chem. Biol. 2015, 10, 1643–1647. [Google Scholar] [CrossRef]
- Salinas, F.; Rojas, V.; Delgado, V.; Lopez, J.; Agosin, E.; Larrondo, L.F. Fungal light-oxygen-voltage domains for optogenetic control of gene expression and flocculation in yeast. MBio 2018, 9. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.; Du, Z.; Liu, R.; Li, T.; Zhao, Y.; Chen, X.; Yang, Y. A single-component optogenetic system allows stringent switch of gene expression in yeast cells. ACS Synth. Biol. 2018, 7, 2045–2053. [Google Scholar] [CrossRef]
- Zhao, E.M.; Lalwani, M.A.; Lovelett, R.J.; García-Echauri, S.A.; Hoffman, S.M.; Gonzalez, C.L.; Toettcher, J.E.; Kevrekidis, I.G.; Avalos, J.L. Design and characterization of rapid optogenetic circuits for dynamic control in yeast metabolic engineering. ACS Synth. Biol. 2020, 9, 3254–3266. [Google Scholar] [CrossRef]
- Zhao, E.M.; Suek, N.; Wilson, M.Z.; Dine, E.; Pannucci, N.L.; Gitai, Z.; Avalos, J.L.; Toettcher, J.E. Light-based control of metabolic flux through assembly of synthetic organelles. Nat. Chem. Biol. 2019, 15, 589–597. [Google Scholar] [CrossRef]
- Zhao, E.M.; Zhang, Y.; Mehl, J.; Park, H.; Lalwani, M.A.; Toettcher, J.E.; Avalos, J.L. Optogenetic regulation of engineered cellular metabolism for microbial chemical production. Nature 2018, 555, 683–687. [Google Scholar] [CrossRef]
- Froehlich, A.C.; Liu, Y.; Loros, J.J.; Dunlap, J.C. White Collar-1, a circadian blue light photoreceptor, binding to the frequency promoter. Science 2002, 297, 815–819. [Google Scholar] [CrossRef]
- He, Q.; Cheng, P.; Yang, Y.; Wang, L.; Gardner, K.H.; Liu, Y. White collar-1, a DNA binding transcription factor and a light sensor. Science 2002, 297, 840–843. [Google Scholar] [CrossRef] [PubMed]
- Malzahn, E.; Ciprianidis, S.; Kaldi, K.; Schafmeier, T.; Brunner, M. Photoadaptation in Neurospora by competitive interaction of activating and inhibitory LOV domains. Cell 2010, 142, 762–772. [Google Scholar] [CrossRef] [Green Version]
- Fields, S.; Song, O.-k. A novel genetic system to detect protein–protein interactions. Nature 1989, 340, 245–246. [Google Scholar] [CrossRef] [PubMed]
- An-Adirekkun, J.M.; Stewart, C.J.; Geller, S.H.; Patel, M.T.; Melendez, J.; Oakes, B.L.; Noyes, M.B.; McClean, M.N. A yeast optogenetic toolkit (yOTK) for gene expression control in Saccharomyces cerevisiae. Biotechnol. Bioeng. 2020, 117, 886–893. [Google Scholar] [CrossRef]
- Brent, R.; Ptashne, M. A eukaryotic transcriptional activator bearing the DNA specificity of a prokaryotic repressor. Cell 1985, 43, 729–736. [Google Scholar] [CrossRef]
- Frankel, A.D.; Kim, P.S. Modular structure of transcription factors: Implications for gene regulation. Cell 1991, 65, 717–719. [Google Scholar] [CrossRef]
- Ma, J.; Ptashne, M. A new class of yeast transcriptional activators. Cell 1987, 51, 113–119. [Google Scholar] [CrossRef]
- Sadowski, I.; Ma, J.; Triezenberg, S.; Ptashne, M. GAL4-VP16 is an unusually potent transcriptional activator. Nature 1988, 335, 563–564. [Google Scholar] [CrossRef] [PubMed]
- Campbell, R.N.; Leverentz, M.K.; Ryan, L.A.; Reece, R.J. Metabolic control of transcription: Paradigms and lessons from Saccharomyces cerevisiae. Biochem. J. 2008, 414, 177–187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MacPherson, S.; Larochelle, M.; Turcotte, B. A fungal family of transcriptional regulators: The zinc cluster proteins. Microbiol. Mol. Biol. Rev. 2006, 70, 583–604. [Google Scholar] [CrossRef] [Green Version]
- Davison, J.M.; Akitake, C.M.; Goll, M.G.; Rhee, J.M.; Gosse, N.; Baier, H.; Halpern, M.E.; Leach, S.D.; Parsons, M.J. Transactivation from Gal4-VP16 transgenic insertions for tissue-specific cell labeling and ablation in zebrafish. Dev. Biol. 2007, 304, 811–824. [Google Scholar] [CrossRef] [Green Version]
- Fischer, J.A.; Giniger, E.; Maniatis, T.; Ptashne, M. GAL4 activates transcription in Drosophila. Nature 1988, 332, 853–856. [Google Scholar] [CrossRef]
- Hartley, K.O.; Nutt, S.L.; Amaya, E. Targeted gene expression in transgenic Xenopus using the binary Gal4-UAS system. Proc. Natl. Acad. Sci. USA 2002, 99, 1377–1382. [Google Scholar] [CrossRef] [Green Version]
- Kakidani, H.; Ptashne, M. GAL4 activates gene expression in mammalian cells. Cell 1988, 52, 161–167. [Google Scholar] [CrossRef]
- Mistry, J.; Chuguransky, S.; Williams, L.; Qureshi, M.; Salazar, G.A.; Sonnhammer, E.L.L.; Tosatto, S.C.E.; Paladin, L.; Raj, S.; Richardson, L.J.; et al. Pfam: The protein families database in 2021. Nucleic Acids Res. 2020, 49, D412–D419. [Google Scholar] [CrossRef]
- Guarente, L.; Lalonde, B.; Gifford, P.; Alani, E. Distinctly regulated tandem upstream activation sites mediate catabolite repression of the CYC1 gene of S. cerevisiae. Cell 1984, 36, 503–511. [Google Scholar] [CrossRef]
- Pfeifer, K.; Arcangioli, B.; Guarente, L. Yeast HAP1 activator competes with the factor RC2 for binding to the upstream activation site UAS1 of the CYC1 gene. Cell 1987, 49, 9–18. [Google Scholar] [CrossRef]
- Creusot, F.; Verdiere, J.; Gaisne, M.; Slonimski, P.P. CYP1 (HAP1) regulator of oxygen-dependent gene expression in yeast. I. Overall organization of the protein sequence displays several novel structural domains. J. Mol. Biol. 1988, 204, 263–276. [Google Scholar] [CrossRef]
- Bricmont, P.A.; Daugherty, J.R.; Cooper, T.G. The DAL81 gene product is required for induced expression of two differently regulated nitrogen catabolic genes in Saccharomyces cerevisiae. Mol. Cell. Biol. 1991, 11, 1161–1166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Talibi, D.; Grenson, M.; Andre, B. Cis- and trans-acting elements determining induction of the genes of the gamma-aminobutyrate (GABA) utilization pathway in Saccharomyces cerevisiae. Nucleic Acids Res. 1995, 23, 550–557. [Google Scholar] [CrossRef] [PubMed]
- Nosaka, K.; Nishimura, H.; Kawasaki, Y.; Tsujihara, T.; Iwashima, A. Isolation and characterization of the THI6 gene encoding a bifunctional thiamin-phosphate pyrophosphorylase/hydroxyethylthiazole kinase from Saccharomyces cerevisiae. J. Biol. Chem. 1994, 269, 30510–30516. [Google Scholar] [CrossRef]
- Davies, B.S.; Wang, H.S.; Rine, J. Dual activators of the sterol biosynthetic pathway of Saccharomyces cerevisiae: Similar activation/regulatory domains but different response mechanisms. Mol. Cell. Biol. 2005, 25, 7375–7385. [Google Scholar] [CrossRef] [Green Version]
- Vik, A.; Rine, J. Upc2p and Ecm22p, dual regulators of sterol biosynthesis in Saccharomyces cerevisiae. Mol. Cell. Biol. 2001, 21, 6395–6405. [Google Scholar] [CrossRef] [Green Version]
- Hedges, D.; Proft, M.; Entian, K.D. CAT8, a new zinc cluster-encoding gene necessary for derepression of gluconeogenic enzymes in the yeast Saccharomyces cerevisiae. Mol. Cell. Biol. 1995, 15, 1915–1922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scholer, A.; Schuller, H.J. A carbon source-responsive promoter element necessary for activation of the isocitrate lyase gene ICL1 is common to genes of the gluconeogenic pathway in the yeast Saccharomyces cerevisiae. Mol. Cell. Biol. 1994, 14, 3613–3622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kammerer, B.; Guyonvarch, A.; Hubert, J.C. Yeast regulatory gene PPR1. I. Nucleotide sequence, restriction map and codon usage. J. Mol. Biol. 1984, 180, 239–250. [Google Scholar] [CrossRef]
- Marmorstein, R.; Harrison, S.C. Crystal structure of a PPR1-DNA complex: DNA recognition by proteins containing a Zn2Cys6 binuclear cluster. Genes Dev. 1994, 8, 2504–2512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roy, A.; Exinger, F.; Losson, R. cis- and trans-acting regulatory elements of the yeast URA3 promoter. Mol. Cell. Biol. 1990, 10, 5257–5270. [Google Scholar] [CrossRef] [Green Version]
- Feller, A.; Dubois, E.; Ramos, F.; Pierard, A. Repression of the genes for lysine biosynthesis in Saccharomyces cerevisiae is caused by limitation of Lys14-dependent transcriptional activation. Mol. Cell. Biol. 1994, 14, 6411–6418. [Google Scholar] [CrossRef] [Green Version]
- Becker, B.; Feller, A.; el Alami, M.; Dubois, E.; Pierard, A. A nonameric core sequence is required upstream of the LYS genes of Saccharomyces cerevisiae for Lys14p-mediated activation and apparent repression by lysine. Mol. Microbiol. 1998, 29, 151–163. [Google Scholar] [CrossRef] [Green Version]
- Iraqui, I.; Vissers, S.; Andre, B.; Urrestarazu, A. Transcriptional induction by aromatic amino acids in Saccharomyces cerevisiae. Mol. Cell. Biol. 1999, 19, 3360–3371. [Google Scholar] [CrossRef] [Green Version]
- Holmberg, S.; Schjerling, P. Cha4p of Saccharomyces cerevisiae activates transcription via serine/threonine response elements. Genetics 1996, 144, 467–478. [Google Scholar] [CrossRef]
- Bornaes, C.; Ignjatovic, M.W.; Schjerling, P.; Kielland-Brandt, M.C.; Holmberg, S. A regulatory element in the CHA1 promoter which confers inducibility by serine and threonine on Saccharomyces cerevisiae genes. Mol. Cell. Biol. 1993, 13, 7604–7611. [Google Scholar] [CrossRef] [Green Version]
- Christianson, T.W.; Sikorski, R.S.; Dante, M.; Shero, J.H.; Hieter, P. Multifunctional yeast high-copy-number shuttle vectors. Gene 1992, 110, 119–122. [Google Scholar] [CrossRef]
- Partow, S.; Siewers, V.; Bjorn, S.; Nielsen, J.; Maury, J. Characterization of different promoters for designing a new expression vector in Saccharomyces cerevisiae. Yeast 2010, 27, 955–964. [Google Scholar] [CrossRef]
- Sikorski, R.S.; Hieter, P. A system of shuttle vectors and yeast host strains designed for efficient manipulation of DNA in Saccharomyces cerevisiae. Genetics 1989, 122, 19–27. [Google Scholar] [CrossRef]
- Karim, A.S.; Curran, K.A.; Alper, H.S. Characterization of plasmid burden and copy number in Saccharomyces cerevisiae for optimization of metabolic engineering applications. FEMS Yeast Res. 2013, 13, 107–116. [Google Scholar] [CrossRef] [Green Version]
- Treitel, M.A.; Carlson, M. Repression by SSN6-TUP1 is directed by MIG1, a repressor/activator protein. Proc. Natl. Acad. Sci. USA 1995, 92, 3132–3136. [Google Scholar] [CrossRef] [Green Version]
- Olesen, J.; Hahn, S.; Guarente, L. Yeast HAP2 and HAP3 activators both bind to the CYC1 upstream activation site, UAS2, in an interdependent manner. Cell 1987, 51, 953–961. [Google Scholar] [CrossRef]
- Chen, J.; Ding, M.; Pederson, D.S. Binding of TFIID to the CYC1 TATA boxes in yeast occurs independently of upstream activating sequences. Proc. Natl. Acad. Sci. USA 1994, 91, 11909–11913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martens, C.; Krett, B.; Laybourn, P.J. RNA polymerase II and TBP occupy the repressed CYC1 promoter. Mol. Microbiol. 2001, 40, 1009–1019. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Chen, X.; Yang, Y. Spatiotemporal control of gene expression by a light-switchable transgene system. Nat. Methods 2012, 9, 266–269. [Google Scholar] [CrossRef] [PubMed]
- Oldenburg, K.R.; Vo, K.T.; Michaelis, S.; Paddon, C. Recombination-mediated PCR-directed plasmid construction in vivo in yeast. Nucleic Acids Res. 1997, 25, 451–452. [Google Scholar] [CrossRef] [Green Version]
- Hughes, R.M.; Bolger, S.; Tapadia, H.; Tucker, C.L. Light-mediated control of DNA transcription in yeast. Methods 2012, 58, 385–391. [Google Scholar] [CrossRef] [Green Version]
- Rienzo, A.; Pascual-Ahuir, A.; Proft, M. The use of a real-time luciferase assay to quantify gene expression dynamics in the living yeast cell. Yeast 2012, 29, 219–231. [Google Scholar] [CrossRef] [Green Version]
- Gietz, R.D.; Schiestl, R.H. High-efficiency yeast transformation using the LiAc/SS carrier DNA/PEG method. Nat. Protoc. 2007, 2, 31–34. [Google Scholar] [CrossRef]
- Longtine, M.S.; McKenzie, A., 3rd; Demarini, D.J.; Shah, N.G.; Wach, A.; Brachat, A.; Philippsen, P.; Pringle, J.R. Additional modules for versatile and economical PCR-based gene deletion and modification in Saccharomyces cerevisiae. Yeast 1998, 14, 953–961. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Transcription Factor | DBD Region (aa) | Target Promoter | Promoter Region Used (bp) |
|---|---|---|---|
| Hap1p [31] | 38–187 | CYC1 [29] | −953 to −1 |
| Dal81p [32] | 124–273 | UGA1 [33] | −534 to −1 |
| Thi2p | 1–150 | THI6 [34] | −422 to −1 |
| Ecm22p [35] | 17–166 | ERG2 [36] | −492 to −1 |
| Cat8p [37] | 43–192 | ICL1 [38] | −830 to −1 |
| Ppr1 [39,40] | 1–150 | URA1 [41] | −1000 to −1 |
| Lys14p [42] | 133–282 | LYS9 [43] | −731 to −1 |
| Aro80p [44] | 1–150 | ARO9 [44] | −608 to −1 |
| Cha4p [45] | 18–167 | CHA1 [46] | −409 to −1 |
| Gal4p (FUN-LOV) [8] | 1–149 | GAL1 (FUN-LOV) [8] | −515 to −1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Romero, A.; Rojas, V.; Delgado, V.; Salinas, F.; Larrondo, L.F. Modular and Molecular Optimization of a LOV (Light–Oxygen–Voltage)-Based Optogenetic Switch in Yeast. Int. J. Mol. Sci. 2021, 22, 8538. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22168538
Romero A, Rojas V, Delgado V, Salinas F, Larrondo LF. Modular and Molecular Optimization of a LOV (Light–Oxygen–Voltage)-Based Optogenetic Switch in Yeast. International Journal of Molecular Sciences. 2021; 22(16):8538. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22168538
Chicago/Turabian StyleRomero, Andrés, Vicente Rojas, Verónica Delgado, Francisco Salinas, and Luis F. Larrondo. 2021. "Modular and Molecular Optimization of a LOV (Light–Oxygen–Voltage)-Based Optogenetic Switch in Yeast" International Journal of Molecular Sciences 22, no. 16: 8538. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22168538