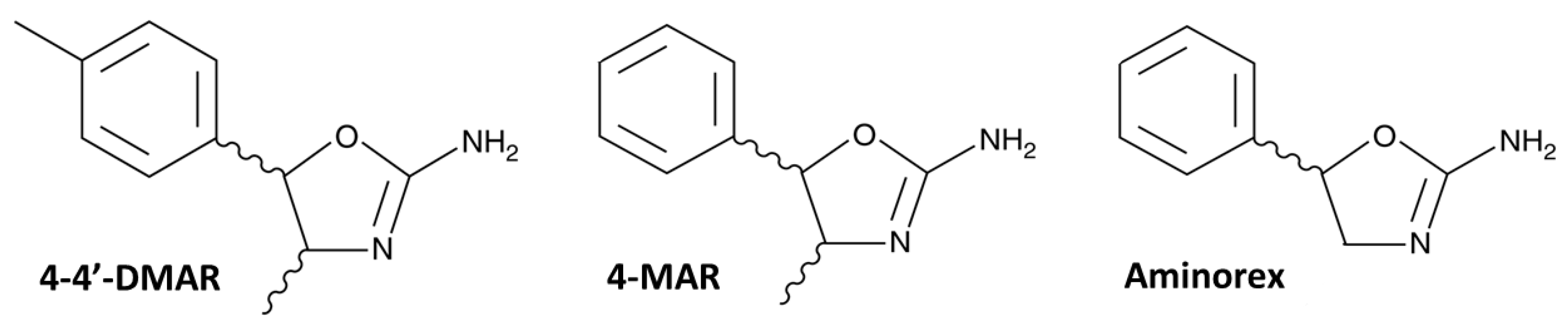
Figure 1.
Structures of 4,4’-DMAR (4-Methyl-5-(4-methylphenyl)-4,5-dihydrooxazol-2-amine), 4-MAR (4-Methyl-5-phenyl-4,5-dihydrooxazol-2-amine) and Aminorex (5-phenyl-4,5-dihydro-1,3-oxazol-2-amine) copied from the Cayman Chemical website (
https://www.caymanchem.com, accessed date 17 May 2021).
Figure 1.
Structures of 4,4’-DMAR (4-Methyl-5-(4-methylphenyl)-4,5-dihydrooxazol-2-amine), 4-MAR (4-Methyl-5-phenyl-4,5-dihydrooxazol-2-amine) and Aminorex (5-phenyl-4,5-dihydro-1,3-oxazol-2-amine) copied from the Cayman Chemical website (
https://www.caymanchem.com, accessed date 17 May 2021).
Figure 2.
The frontal cortex of the treated mice ((±)cis 30; (b) showed mild edema (i.e., vacuolization areas are indicated by yellow arrows) compared to controls (a).
Figure 2.
The frontal cortex of the treated mice ((±)cis 30; (b) showed mild edema (i.e., vacuolization areas are indicated by yellow arrows) compared to controls (a).
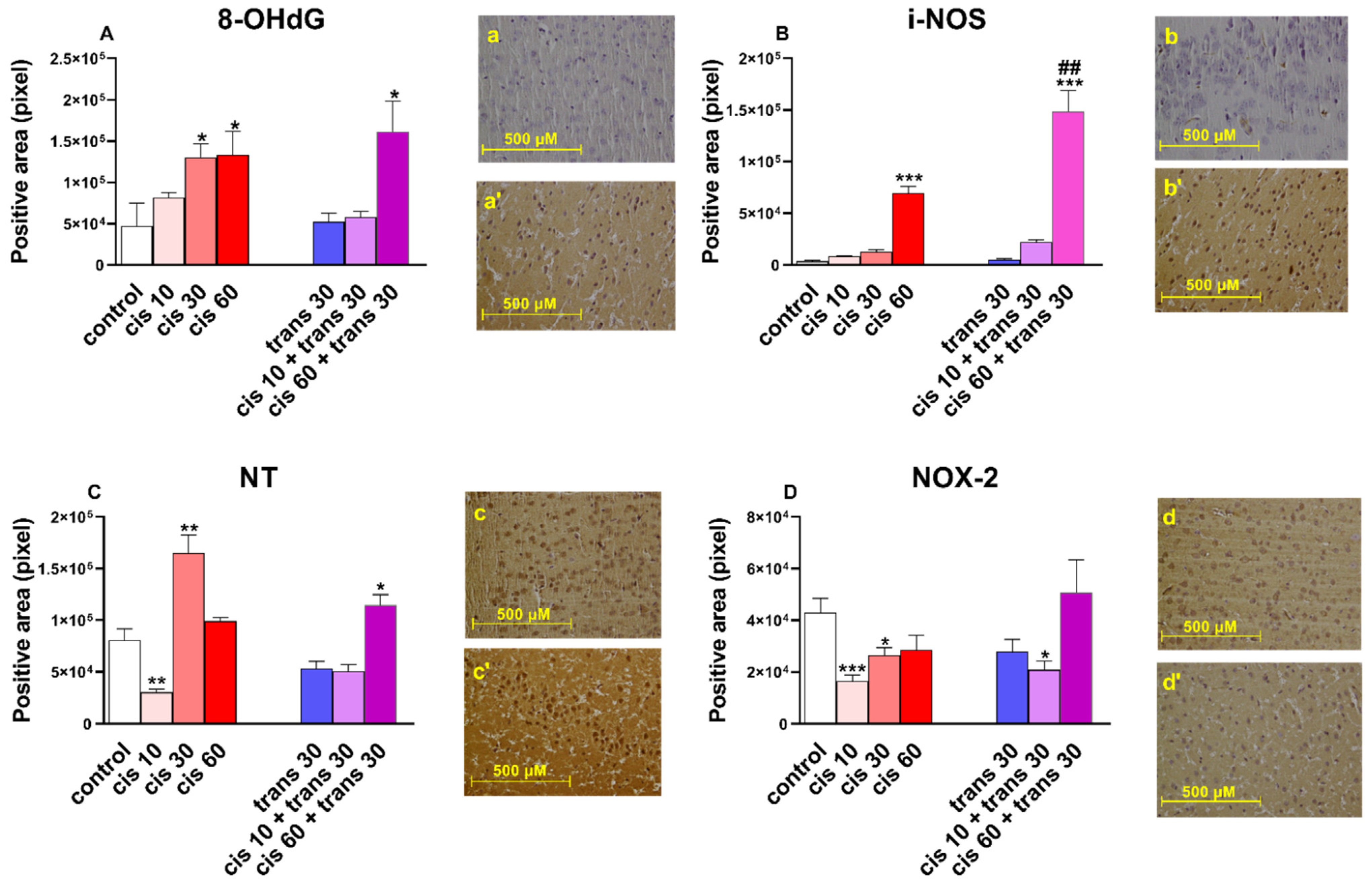
Figure 3.
(A) Quantification and comparison of 8-OHdG positive areas in controls versus 4-4′-DMAR-mice ((±)cis 10, (±)cis 30, (±)cis 60, (±)trans 30, (±)trans 30 + (±)cis 10, (±)trans 30 + (±)cis 60). Representative image (light microscopy, 40×) of 8-OHdG immunostaining (brown areas) in the cortex of mice receiving 4-4′-DMAR ((±)cis 30, a’) and controls (a). (B) Quantification and comparison of iNOS positive areas in controls versus 4-4′-DMAR-mice ((±)cis 10, (±)cis 30, (±)cis 60, (±)trans 30, (±)trans 30 + (±)cis 10, (±)trans 30 + (±)cis 60). Comparison of (±)cis 60 versus (±)trans 30 + (±)cis 60. Representative image (light microscopy, 40×) of iNOS immunostaining (brown areas) in the cortex of mice receiving 4-4′-DMAR ((±)cis 60, b’) and controls (b). (C) Quantification and comparison of NT (nitrotyrosine) positive areas in controls versus 4-4′-DMAR-mice ((±)cis 10, (±)cis 30, (±)cis 60, (±)trans 30, (±)trans 30 + (±)cis 10, (±)trans 30 + (±)cis 60). Representative image (light microscopy, 40x) of NT immunostaining (brown areas) in the cortex of mice receiving 4-4′-DMAR ((±)cis 30, c’) and controls (c). (D) Quantification and comparison of NOX2 positive areas in controls versus 4-4′-DMAR-mice ((±)cis 10, (±)cis 30, (±)cis 60, (±)trans 30, (±)trans 30 + (±)cis 10, (±)trans 30 + (±)cis 60). Representative image (light microscopy, 40×) of NOX2 immunostaining (brown areas) in the cortex of mice receiving 4-4′-DMAR ((±)cis 30, d’) and controls (d). * p < 0.05, ** p < 0.01 and *** p < 0.001 different from control; ## p < 0.001 different from (±)cis 60.
Figure 3.
(A) Quantification and comparison of 8-OHdG positive areas in controls versus 4-4′-DMAR-mice ((±)cis 10, (±)cis 30, (±)cis 60, (±)trans 30, (±)trans 30 + (±)cis 10, (±)trans 30 + (±)cis 60). Representative image (light microscopy, 40×) of 8-OHdG immunostaining (brown areas) in the cortex of mice receiving 4-4′-DMAR ((±)cis 30, a’) and controls (a). (B) Quantification and comparison of iNOS positive areas in controls versus 4-4′-DMAR-mice ((±)cis 10, (±)cis 30, (±)cis 60, (±)trans 30, (±)trans 30 + (±)cis 10, (±)trans 30 + (±)cis 60). Comparison of (±)cis 60 versus (±)trans 30 + (±)cis 60. Representative image (light microscopy, 40×) of iNOS immunostaining (brown areas) in the cortex of mice receiving 4-4′-DMAR ((±)cis 60, b’) and controls (b). (C) Quantification and comparison of NT (nitrotyrosine) positive areas in controls versus 4-4′-DMAR-mice ((±)cis 10, (±)cis 30, (±)cis 60, (±)trans 30, (±)trans 30 + (±)cis 10, (±)trans 30 + (±)cis 60). Representative image (light microscopy, 40x) of NT immunostaining (brown areas) in the cortex of mice receiving 4-4′-DMAR ((±)cis 30, c’) and controls (c). (D) Quantification and comparison of NOX2 positive areas in controls versus 4-4′-DMAR-mice ((±)cis 10, (±)cis 30, (±)cis 60, (±)trans 30, (±)trans 30 + (±)cis 10, (±)trans 30 + (±)cis 60). Representative image (light microscopy, 40×) of NOX2 immunostaining (brown areas) in the cortex of mice receiving 4-4′-DMAR ((±)cis 30, d’) and controls (d). * p < 0.05, ** p < 0.01 and *** p < 0.001 different from control; ## p < 0.001 different from (±)cis 60.
![Ijms 22 08771 g003]()
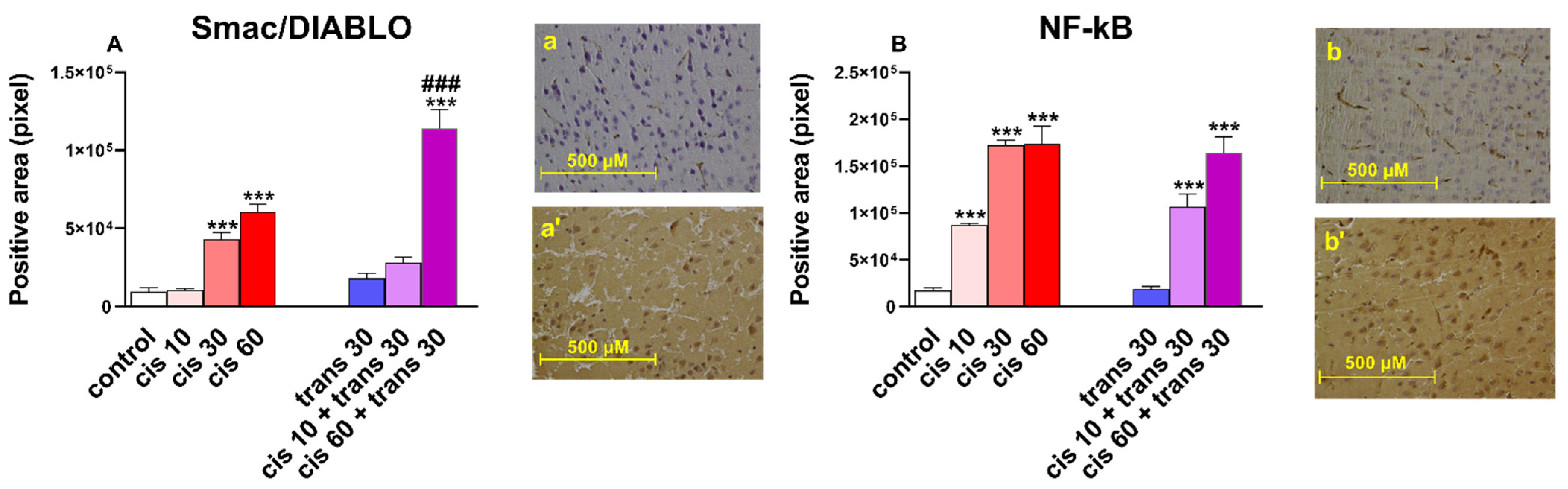
Figure 4.
(A) Quantification and comparison of Smac/DIABLO positive areas in controls versus 4-4′-DMAR-mice ((±)cis 10, (±)cis 30, (±)cis 60, (±)trans 30, (±)trans 30 + (±)cis 10, (±)trans 30 + (±)cis 60).). Comparison of (±)cis 60 versus (±)trans 30 + (±)cis 60. Representative image (light microscopy, 40×) of Smac/DIABLO immunostaining (brown areas) in the cortex of mice receiving 4-4′-DMAR ((±)trans 30+(±)cis 60, a’) and controls (a). (B) Quantification and comparison of NF-κB positive areas in controls versus 4-4′-DMAR-mice ((±)cis 10, (±)cis 30, (±)cis 60, (±)trans 30, (±)trans 30 + (±)cis 10, (±)trans 30 + (±)cis 60). Representative image (light microscopy, 40x) of NF-κB immunostaining (brown areas) in the cortex of mice receiving 4-4′-DMAR ((±)cis 10, b’) and controls (b). *** p < 0.001 different from control; ### p < 0.001 different from (±)cis 60.
Figure 4.
(A) Quantification and comparison of Smac/DIABLO positive areas in controls versus 4-4′-DMAR-mice ((±)cis 10, (±)cis 30, (±)cis 60, (±)trans 30, (±)trans 30 + (±)cis 10, (±)trans 30 + (±)cis 60).). Comparison of (±)cis 60 versus (±)trans 30 + (±)cis 60. Representative image (light microscopy, 40×) of Smac/DIABLO immunostaining (brown areas) in the cortex of mice receiving 4-4′-DMAR ((±)trans 30+(±)cis 60, a’) and controls (a). (B) Quantification and comparison of NF-κB positive areas in controls versus 4-4′-DMAR-mice ((±)cis 10, (±)cis 30, (±)cis 60, (±)trans 30, (±)trans 30 + (±)cis 10, (±)trans 30 + (±)cis 60). Representative image (light microscopy, 40x) of NF-κB immunostaining (brown areas) in the cortex of mice receiving 4-4′-DMAR ((±)cis 10, b’) and controls (b). *** p < 0.001 different from control; ### p < 0.001 different from (±)cis 60.
Figure 5.
(A) Quantification and comparison of HSP27 positive areas in controls versus 4-4′-DMAR-mice ((±)cis 10, (±)cis 30, (±)cis 60, (±)trans 30, (±)trans 30 + (±)cis 10, (±)trans 30 + (±)cis 60). ). Comparison of (±)cis 60 versus (±)trans 30 + (±)cis 60). Comparison of (±)cis 10 versus (±)trans 30 + (±)cis 10 [°]. Representative image (light microscopy, 40×) of HSP27 immunostaining (brown areas) in the cortex of mice receiving 4-4′-DMAR ((±)cis 60, a’) and controls (a). (B) Quantification and comparison of HSP70 positive areas in controls versus 4-4′-DMAR-mice ((±)cis 10, (±)cis 30, (±)cis 60, (±)trans 30, (±)trans 30 + (±)cis 10, (±)trans 30 + (±)cis 60). Comparison of (±)cis 10 versus (±)trans 30 + (±)cis 10 [°]. Representative image (light microscopy, 40x) of HSP70 immunostaining (brown areas) in the cortex of mice receiving 4-4′-DMAR ((±)cis 60, b’) and controls (b). (C) Quantification and comparison of HSP90 positive areas in controls versus 4-4′-DMAR-mice ((±)cis 10, (±)cis 30, (±)cis 60, (±)trans 30, (±)trans 30 + (±)cis 10, (±)trans 30 + (±)cis 60). Representative image (light microscopy, 40×) of HSP90 immunostaining (brown areas) in the cortex of mice receiving 4-4′-DMAR ((±)cis 30, c’) and controls (c). * p < 0.05, ** p < 0.01 and *** p < 0.001 different from control; ° p < 0.05 and °°° p< 0.001 different from (±)cis 30; ## p < 0.001 different from (±)cis 60.
Figure 5.
(A) Quantification and comparison of HSP27 positive areas in controls versus 4-4′-DMAR-mice ((±)cis 10, (±)cis 30, (±)cis 60, (±)trans 30, (±)trans 30 + (±)cis 10, (±)trans 30 + (±)cis 60). ). Comparison of (±)cis 60 versus (±)trans 30 + (±)cis 60). Comparison of (±)cis 10 versus (±)trans 30 + (±)cis 10 [°]. Representative image (light microscopy, 40×) of HSP27 immunostaining (brown areas) in the cortex of mice receiving 4-4′-DMAR ((±)cis 60, a’) and controls (a). (B) Quantification and comparison of HSP70 positive areas in controls versus 4-4′-DMAR-mice ((±)cis 10, (±)cis 30, (±)cis 60, (±)trans 30, (±)trans 30 + (±)cis 10, (±)trans 30 + (±)cis 60). Comparison of (±)cis 10 versus (±)trans 30 + (±)cis 10 [°]. Representative image (light microscopy, 40x) of HSP70 immunostaining (brown areas) in the cortex of mice receiving 4-4′-DMAR ((±)cis 60, b’) and controls (b). (C) Quantification and comparison of HSP90 positive areas in controls versus 4-4′-DMAR-mice ((±)cis 10, (±)cis 30, (±)cis 60, (±)trans 30, (±)trans 30 + (±)cis 10, (±)trans 30 + (±)cis 60). Representative image (light microscopy, 40×) of HSP90 immunostaining (brown areas) in the cortex of mice receiving 4-4′-DMAR ((±)cis 30, c’) and controls (c). * p < 0.05, ** p < 0.01 and *** p < 0.001 different from control; ° p < 0.05 and °°° p< 0.001 different from (±)cis 30; ## p < 0.001 different from (±)cis 60.
![Ijms 22 08771 g005]()
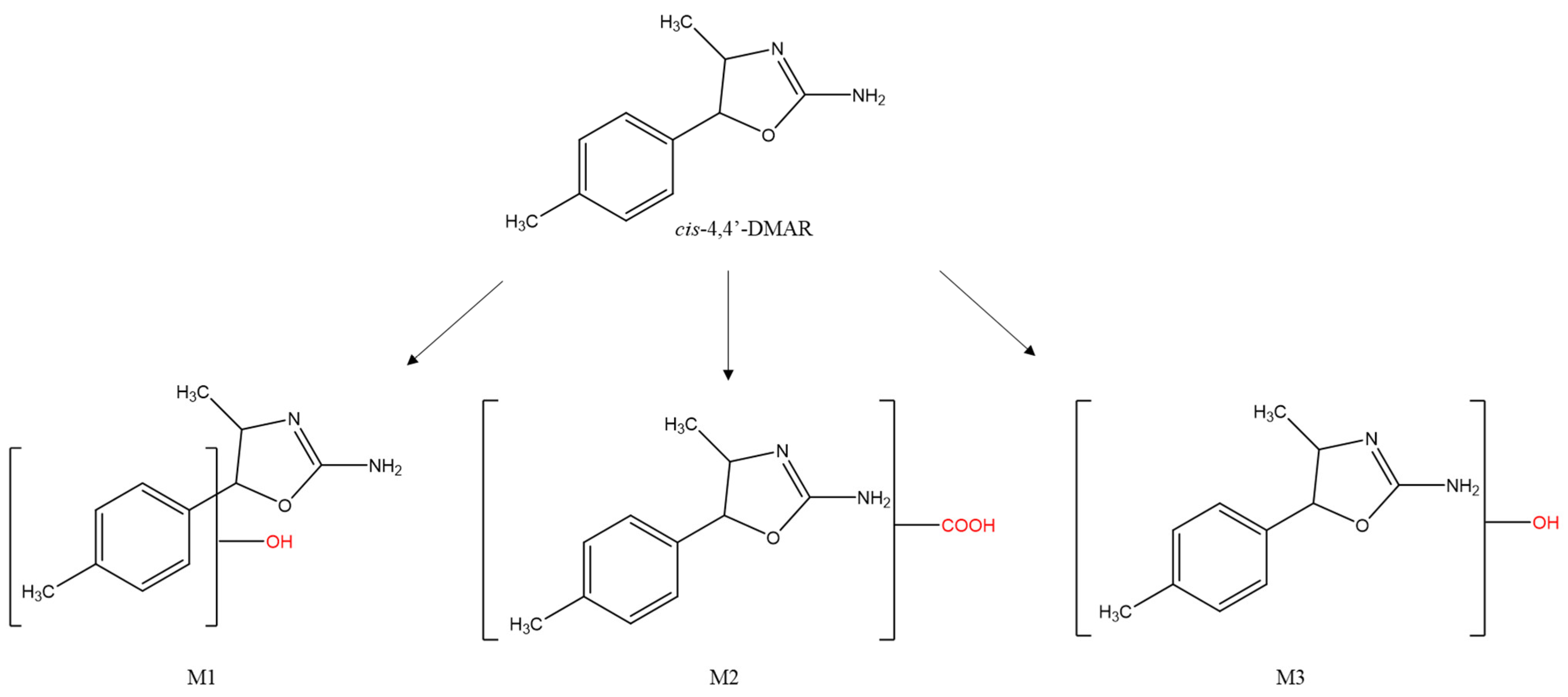
Figure 6.
Structures of cis-4,4’-DMAR and its principal markers: hydroxylated (M1, M3) and carboxylated (M2).
Figure 6.
Structures of cis-4,4’-DMAR and its principal markers: hydroxylated (M1, M3) and carboxylated (M2).
Figure 7.
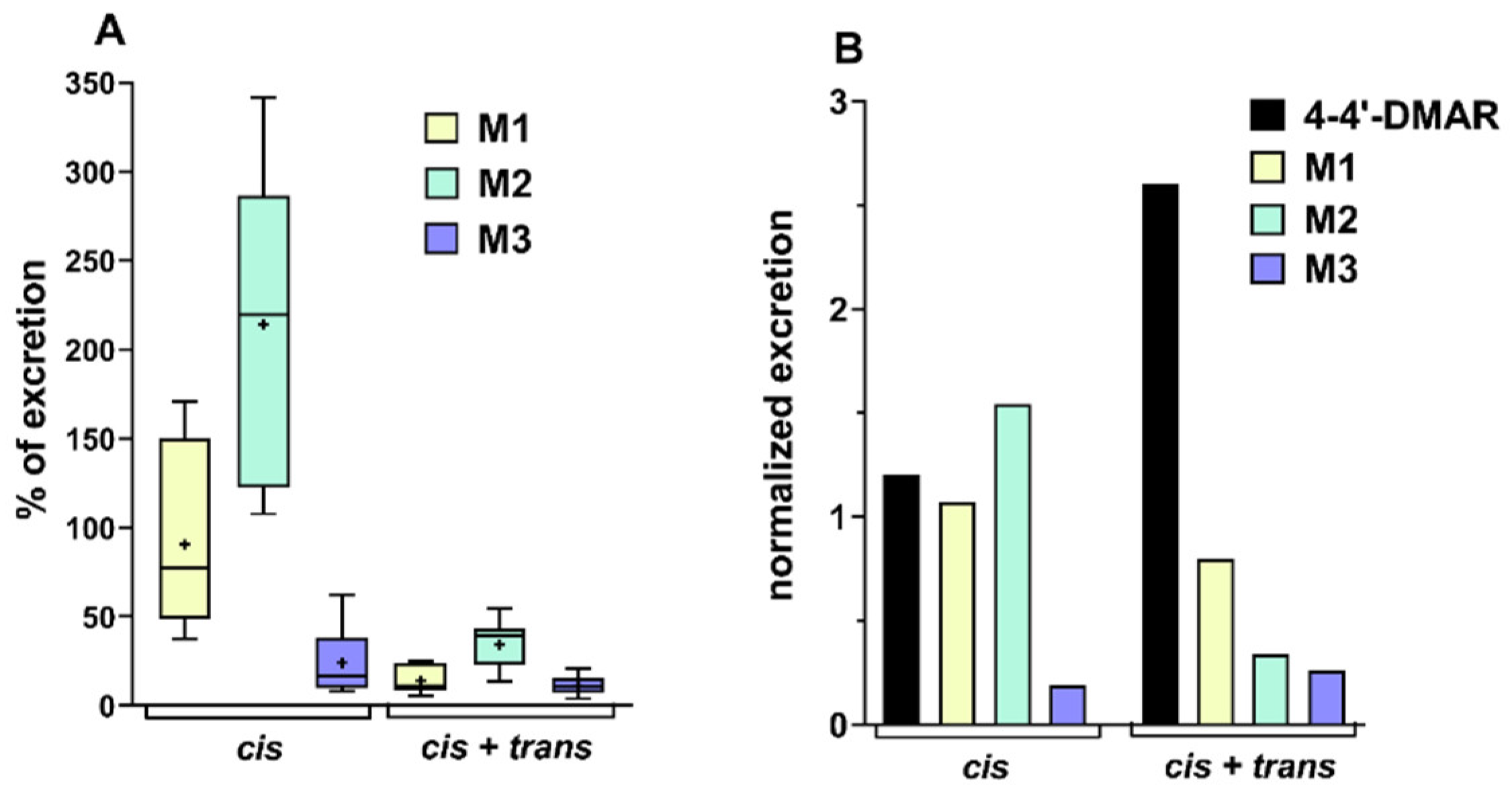
(A) Percentage ratio of excretion of M1, M2 and M3, the principal metabolites of cis-4,4′-DMAR for the administration of a dose of 10 mg/kg of 4,4′-DMAR (cis) and the same dose of both isomers (cis + trans). The box plots report maximum and minimum value, median and average (+). (B) Excretion data of 4,4′-cis-DMAR and its principal metabolites normalised to the sum of excretion of 4,4′-DMAR and its metabolites. Data normalised and reported, respectively, for the administration of cis or cis + trans isomers at a dose of 10 mg/kg.
Figure 7.
(A) Percentage ratio of excretion of M1, M2 and M3, the principal metabolites of cis-4,4′-DMAR for the administration of a dose of 10 mg/kg of 4,4′-DMAR (cis) and the same dose of both isomers (cis + trans). The box plots report maximum and minimum value, median and average (+). (B) Excretion data of 4,4′-cis-DMAR and its principal metabolites normalised to the sum of excretion of 4,4′-DMAR and its metabolites. Data normalised and reported, respectively, for the administration of cis or cis + trans isomers at a dose of 10 mg/kg.
Table 1.
Effect of (±)cis-4,4′-DMAR (0.1–60 mg/kg i.p.) on neuro-behavioural changes (psychomotor agitation, aggressiveness, convulsions), physiological alterations (sweating, salivation, hyperthermia) and lethality in mice. The data here shown (see material and methods) refers to the mean ± SEM values relating to six animals for each treatment. The statistical analysis of the effects of the (±)cis-4,4′-DMAR in different concentrations were performed using a one-way ANOVA, followed by a Bonferroni test for multiple comparisons. A Student’s t-test was used to determine statistical significance (p < 0.05) between the two groups. a p < 0.05, versus cis 3 mg/kg; b p < 0.05, versus cis 10 mg/kg; c p < 0.05, versus vehicle; d p < 0.05, versus cis 30 mg/kg.
Table 1.
Effect of (±)cis-4,4′-DMAR (0.1–60 mg/kg i.p.) on neuro-behavioural changes (psychomotor agitation, aggressiveness, convulsions), physiological alterations (sweating, salivation, hyperthermia) and lethality in mice. The data here shown (see material and methods) refers to the mean ± SEM values relating to six animals for each treatment. The statistical analysis of the effects of the (±)cis-4,4′-DMAR in different concentrations were performed using a one-way ANOVA, followed by a Bonferroni test for multiple comparisons. A Student’s t-test was used to determine statistical significance (p < 0.05) between the two groups. a p < 0.05, versus cis 3 mg/kg; b p < 0.05, versus cis 10 mg/kg; c p < 0.05, versus vehicle; d p < 0.05, versus cis 30 mg/kg.
| COMPOUND | | cis-4,4’-DMAR |
|---|
| Doses (mg/kg) | vehicle | 0.1 | 1 | 3 | 10 | 30 | 60 |
|---|
| Psychomotor agitation | Frequency % (n° of mice) | - | - | - | 100% (6) | 100% (6) | 100% (6) | 100% (6) |
| Duration (min) | - | - | - | 54.2 ± 10.30 | 121.0 ± 12.35 a | 235.1 ± 15.25 ab | 280.1 ± 12.35 ab |
| Latency (min) | - | - | - | 61.2 ± 6.30 | 28.2 ± 7.20 a | 3.2 ± 0.25 ab | 2.2 ± 0.15 ab |
| Sweating | Frequency % (n° of mice) | - | - | - | - | - | 100% (6) | 100% (6) |
| Salivation | Frequency % (n° of mice) | - | - | - | - | - | 100% (6) | 50% (3) |
| Hyperthermia | Variation (Δ°C) | −0.28 ± 0.12 | −0.18 ± 0.14 | −0.22 ± 0.17 | −0.34 ± 0.22 | −0.41 ± 0.13 | 1.52 ± 0.11 c | 2.22 ± 0.12 c |
| Latency (min) | - | - | - | - | - | 25 ± 0.25 | 7.12 ± 0.23 d |
| Aggressiveness | Spontaneus | Frequency % (n° of mice) | - | - | - | - | - | - | nd |
| Stimulated | Frequency % (n° of mice) | - | - | - | - | 100% (6) | 100% (6) | nd |
| Score (n° of bites) | - | - | - | - | 6 ± 0.23 | 10 ± 0.23 b | nd |
| Convulsion | Frequency % (n° of mice) | - | - | - | - | - | 50% (3) | 100% (6) |
| Episodes (n°) | - | - | - | - | - | 3.5 ± 0.5 | 2.0 ± 0.41 d |
| Latency of first episode (sec) | - | - | - | - | - | 10.5 ± 1.5 | 8.25 ± 1.1 |
| Duration of each episode (sec) | - | - | - | - | - | 4.86 ± 1.26 | 16.14 ± 3.2 d |
| Lethality | Frequency % (n° of mice) | - | - | - | - | - | 50% (3) | 100% (6) |
| Time of death (min) | - | - | - | - | - | 59.3 ± 2.3 | 31.9 ± 5.6 d |
| Surviving mice % (n° of mice) | - | - | - | - | - | 50% (3) | 0% (0) |
Table 2.
Effect of (±)trans-4,4′-DMAR (30 and 60 mg/kg i.p.) on neuro-behavioural changes (psychomotor agitation, aggressiveness, convulsion), physiological alterations (sweating, salivation, hyperthermia) and lethality in mice. Data expressed (see material and methods) represents the mean ± SEM of six animals for each treatment. The statistical analysis of the effects of the (±)trans-4,4′-DMAR in different concentrations were performed using a one-way ANOVA followed by a Bonferroni test for multiple comparisons. A Student’s t-test was used to determine statistical significance (p < 0.05) between the two groups.
Table 2.
Effect of (±)trans-4,4′-DMAR (30 and 60 mg/kg i.p.) on neuro-behavioural changes (psychomotor agitation, aggressiveness, convulsion), physiological alterations (sweating, salivation, hyperthermia) and lethality in mice. Data expressed (see material and methods) represents the mean ± SEM of six animals for each treatment. The statistical analysis of the effects of the (±)trans-4,4′-DMAR in different concentrations were performed using a one-way ANOVA followed by a Bonferroni test for multiple comparisons. A Student’s t-test was used to determine statistical significance (p < 0.05) between the two groups.
| COMPOUND | | trans-4,4’-DMAR |
|---|
| Doses (mg/kg) | vehicle | 30 | 60 |
|---|
| Psychomotor agitation | Frequency % (n° of mice) | - | - | - |
| Duration (min) | - | - | - |
| Latency (min) | - | - | - |
| Sweating | Frequency % (n° of mice) | - | - | - |
| Salivation | Frequency % (n° of mice) | - | - | - |
| Hyperthermia | Variation (Δ°C) | −0.28 ± 0.12 | −0.37 ± 0.13 | −0.35 ± 0.16 |
| Latency (min) | - | - | - |
| Aggressiveness | Spontaneus | Frequency % (n° of mice) | - | - | - |
| Stimulated | Frequency % (n° of mice) | - | - | - |
| Score (n° of bites) | - | - | - |
| Convulsion | Frequency % (n° of mice) | - | - | - |
| Episodes (n°) | - | - | - |
| Latency of first episode (sec) | - | - | - |
| Duration of each episode (sec) | - | - | - |
| Lethality | Frequency % (n° of mice) | - | - | - |
| Time of death (min) | - | - | - |
Table 3.
Effect of co-administration of (±)cis-4,4′-DMAR (1, 10 and 60 mg/kg i.p.) and (±)trans-4,4′-DMAR (30 mg/kg i.p.) on neuro-behavioural changes (psychomotor agitation, aggressiveness, convulsion), physiological alterations (sweating, salivation, hyperthermia) and death in mice. Data expressed (see material and methods) represents the mean ± SEM of six animals for each treatment. The statistical analysis of the effects of the interactions between (±)cis-4,4′-DMAR and (±)trans-4,4′-DMAR were performed using a one-way ANOVA followed by a Bonferroni test for multiple comparisons. A Student’s t-test was used to determine statistical significance (p < 0.05) between the two groups. e p < 0.05, versus cis 10 mg/kg; f p < 0.05, versus cis 60 mg/kg.
Table 3.
Effect of co-administration of (±)cis-4,4′-DMAR (1, 10 and 60 mg/kg i.p.) and (±)trans-4,4′-DMAR (30 mg/kg i.p.) on neuro-behavioural changes (psychomotor agitation, aggressiveness, convulsion), physiological alterations (sweating, salivation, hyperthermia) and death in mice. Data expressed (see material and methods) represents the mean ± SEM of six animals for each treatment. The statistical analysis of the effects of the interactions between (±)cis-4,4′-DMAR and (±)trans-4,4′-DMAR were performed using a one-way ANOVA followed by a Bonferroni test for multiple comparisons. A Student’s t-test was used to determine statistical significance (p < 0.05) between the two groups. e p < 0.05, versus cis 10 mg/kg; f p < 0.05, versus cis 60 mg/kg.
| COMPOUND | | cis-4,4’-DMAR | trans | (cis + trans)-4-4’DMAR |
|---|
| Doses (mg/kg) | vehicle | 1 | 10 | 60 | 30 | 1 + 30 | 10 + 30 | 60 + 30 |
|---|
Psychomotor
agitation | Frequency % (n° of mice) | - | - | 100% (6) | 100% (6) | - | 50% (3) | 100% (6) | 100% (6) |
| Duration (min) | - | - | 121.0 ± 12.35 | 220.1 ± 12.35 | - | 55.0 ± 5.0 | 185.5 ± 11.0 e | nd |
| Latency (min) | - | - | 28.2 ± 7.20 | 2.2 ± 0.15 | - | 45.2 ± 5.20 | 6.2 ± 2.20 e | 0.45 ± 0.10 f |
| Sweating | Frequency % (n° of mice) | - | - | - | 100% (6) | - | - | 33% (2) | 100% (6) |
| Salivation | Frequency % (n° of mice) | - | - | - | 50% (3) | - | - | 33% (2) | 50% (3) |
| Hyperthermia | Variation (Δ°C) | −0.28 ± 0.12 | −0.22 ± 0.17 | −0.41 ± 0.13 | 2.22 ± 0.12 | −0.37 ± 0.13 | −0.70 ± 0.15 | 1.6 ± 0.11 e | 2.25 ± 0.09 |
| Latency (min) | - | - | - | 7.12 ± 0.23 | - | - | 28.0 ± 0.21 | 5.0 ± 0.23 f |
| Aggressiveness | Spontaneus | Frequency % (n° of mice) | - | - | - | nd | - | - | - | nd |
| Stimulated | Frequency % (n° of mice) | - | - | 100% (6) | nd | - | - | 100% (6) | nd |
| Score (n° of bites) | - | - | 6 ± 0.23 | nd | - | - | 10 ± 0.05 e | nd |
| Convulsion | Frequency % (n° of mice) | - | - | - | 100% (6) | - | - | - | 100% (6) |
| Episodes (n°) | - | - | - | 2.0 ± 0.41 | - | - | - | 1.0 ± 0.0 f |
| Latency of first episode (sec) | - | - | - | 8.25 ± 1.1 | - | - | - | 6.67 ± 2.73 |
| Duration of each episode (sec) | - | - | - | 16.14 ± 6.2 | - | - | - | 40.0 ± 5.0 f |
| Lethality | Frequency % (n° of mice) | - | - | - | 100% (6) | - | - | - | 100% (6) |
| Time of death (min) | - | - | - | 31.9 ± 5.6 | - | - | - | 11.0 ± 3.9 f |
| Surviving mice % (n° of mice) | - | - | - | 0% (0) | - | - | - | 0% (0) |
Table 4.
Effect of co-administration of (±)cis-4,4′-DMAR (10 mg/kg i.p.) and (±)trans-4,4′-DMAR (10 mg/kg i.p.) on neuro-behavioural changes (psychomotor agitation, aggressiveness, convulsion), physiological alterations (sweating, salivation, hyperthermia) and death in mice. Data expressed (see material and methods) represents the mean ± SEM of four animals for each treatment. The statistical analysis of the effects of the interactions between (±)cis-4,4′-DMAR and (±)trans-4,4′-DMAR were performed using a one-way ANOVA followed by a Bonferroni test for multiple comparisons. A Student’s t-test was used to determine statistical significance (p < 0.05) between the two groups. e p < 0.05, versus cis 10 mg/kg.
Table 4.
Effect of co-administration of (±)cis-4,4′-DMAR (10 mg/kg i.p.) and (±)trans-4,4′-DMAR (10 mg/kg i.p.) on neuro-behavioural changes (psychomotor agitation, aggressiveness, convulsion), physiological alterations (sweating, salivation, hyperthermia) and death in mice. Data expressed (see material and methods) represents the mean ± SEM of four animals for each treatment. The statistical analysis of the effects of the interactions between (±)cis-4,4′-DMAR and (±)trans-4,4′-DMAR were performed using a one-way ANOVA followed by a Bonferroni test for multiple comparisons. A Student’s t-test was used to determine statistical significance (p < 0.05) between the two groups. e p < 0.05, versus cis 10 mg/kg.
| COMPOUND | cis-4,4’-DMAR | trans-4,4’-DMAR | (cis + trans)-4-4’DMAR |
|---|
| Doses (mg/kg) | 10 | 10 | 10+10 |
|---|
| Psychomotoragitation | Frequency % (n° of mice) | 100% (4) | - | 100% (4) |
| Duration (min) | 125.0 ± 10.12 | - | 171.0 ± 9.25 e |
| Latency (min) | 26.4 ± 6.25 | - | 8.0 ± 4.25 e |
| Sweating | Frequency % (n° of mice) | - | - | - |
| Salivation | Frequency % (n° of mice) | - | - | - |
| Hyperthermia | Variation (Δ°C) | −0.21 ± 0.15 | −0.32 ± 0.1 | 1.58 ± 0.23 e |
| Latency (min) | - | - | 32.0 ± 2.45 |
| Aggressiveness | Spontaneus | Frequency % (n° of mice) | - | - | - |
| Stimulated | Frequency % (n° of mice) | 100% (4) | - | 100% (4) |
| Score (n° of bites) | 5 ± 0.23 | - | 8 ± 0.13 e |
| Convulsion | Frequency % (n° of mice) | - | - | - |
| Episodes (n°) | - | - | - |
| Latency of first episode (sec) | - | - | - |
| Duration of each episode (sec) | - | - | - |
| Lethality | Frequency % (n° of mice) | - | - | - |
| Time of death (min) | - | - | - |
Table 5.
Correlation between mouse doses (mg/kg) and human equivalent doses (HED, mg/kg). The table also reported the correlation between doses and effects in human.
Table 5.
Correlation between mouse doses (mg/kg) and human equivalent doses (HED, mg/kg). The table also reported the correlation between doses and effects in human.
| Mouse Dose (mg/kg) | HED | Human Dose | Human Dosage | Effects |
|---|
| (mg/kg) | (mg) |
|---|
| 0.1 | 0.0081 | 0.486 | Low | high state of vigilance, euphoria, decreased appetite, increased frequency of heartbeat and motor activity |
| 1 | 0.081 | 4.86 |
| 3 | 0.243 | 14.58 |
| 10 | 0.81 | 48.6 | Intermediate | restlessness, agitation and insomnia |
| 30 | 2.43 | 145.8 | high | involve severe anorexia, mild paranoia (sometimes hallucinations), hyperthermia, bruxism, facial spasms, an increase in aggression and desire for violence, seizures, an increased heart rate that will be involved in a cardiac arrest |
| 60 | 4.86 | 291.6 |
Table 6.
Antibodies used for immunohistochemical analyses with the relative dilutions and antigenic retrieval methods.
Table 6.
Antibodies used for immunohistochemical analyses with the relative dilutions and antigenic retrieval methods.
| Marker | | Dilution | Retrieval |
|---|
| HSP27 | Santa Cruz Biotechnology, Inc. | 1:50 | HIER (0.25 mM EDTA buffer ) |
| HSP70 | Santa Cruz Biotechnology, Inc. | 1:50 | HIER (0.25 mM EDTA buffer ) |
| HSP90 | Santa Cruz Biotechnology, Inc. | 1:50 | HIER (0.25 mM EDTA buffer ) |
| SMAC | Santa Cruz Biotechnology, Inc. | 1:100 | HIER (0.01 M citrate buffer) |
| NF-kB | Santa Cruz Biotechnology, Inc. | 1:50 | HIER (0.25 mM EDTA buffer ) |
| iNOS | Santa Cruz Biotechnology, Inc. | 1:100 | HIER (0.01 M citrate buffer) |
| NOX-2 | Proteintech | 1:100 | HIER (0.01 M citrate buffer) |
| NT | Santa Cruz Biotechnology, Inc. | 1:600 | HIER (0.01 M citrate buffer) |
| 8OHDG | Santa Cruz Biotechnology, Inc. | 1:500 | HIER (0.01 M citrate buffer) |


 ,
, 









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}