
Pattern of TAAR5 Expression in the Human Brain Based on Transcriptome Datasets Analysis

 ,
,  , , , and
, , , and
Abstract
:1. Introduction
2. Results
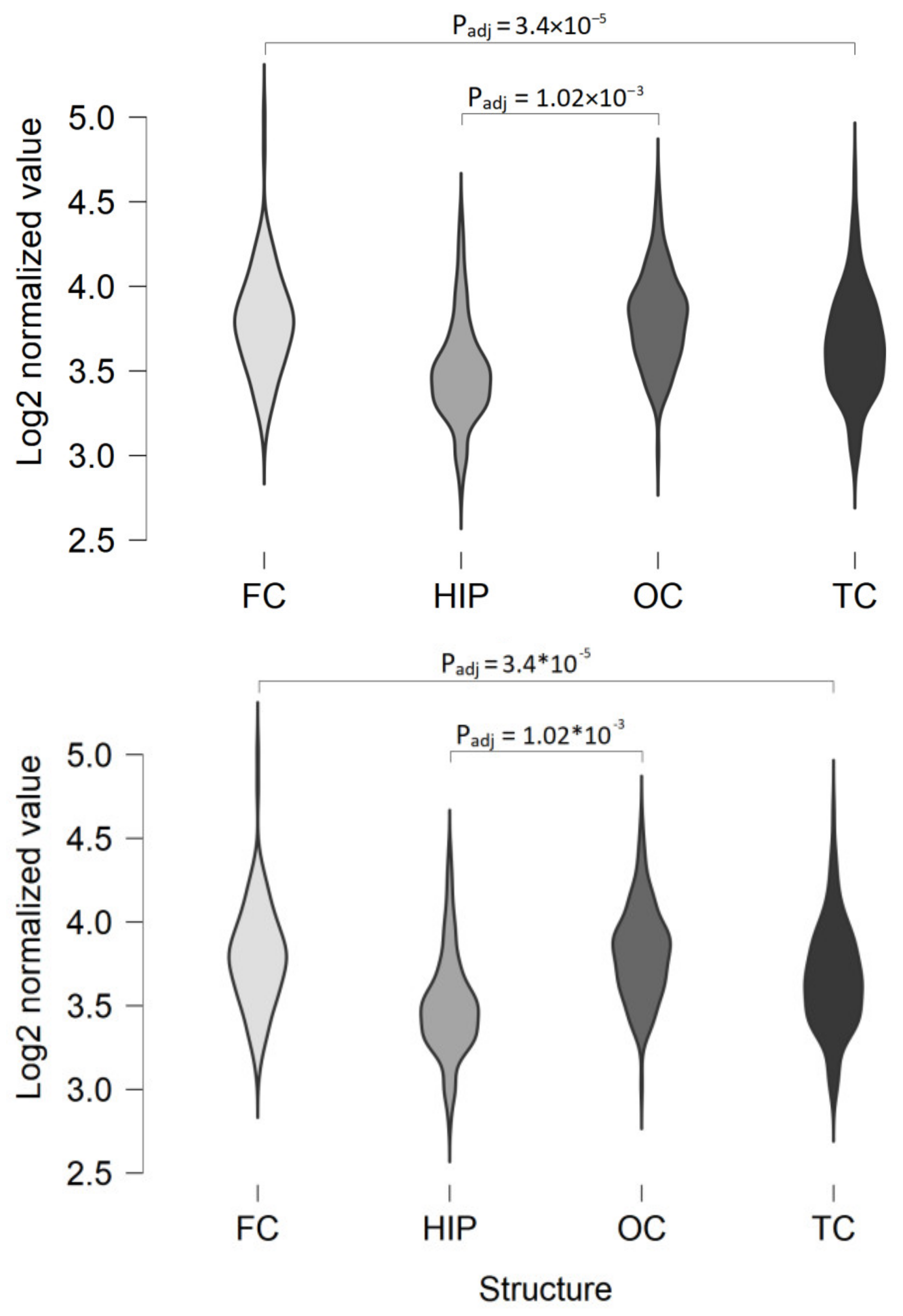
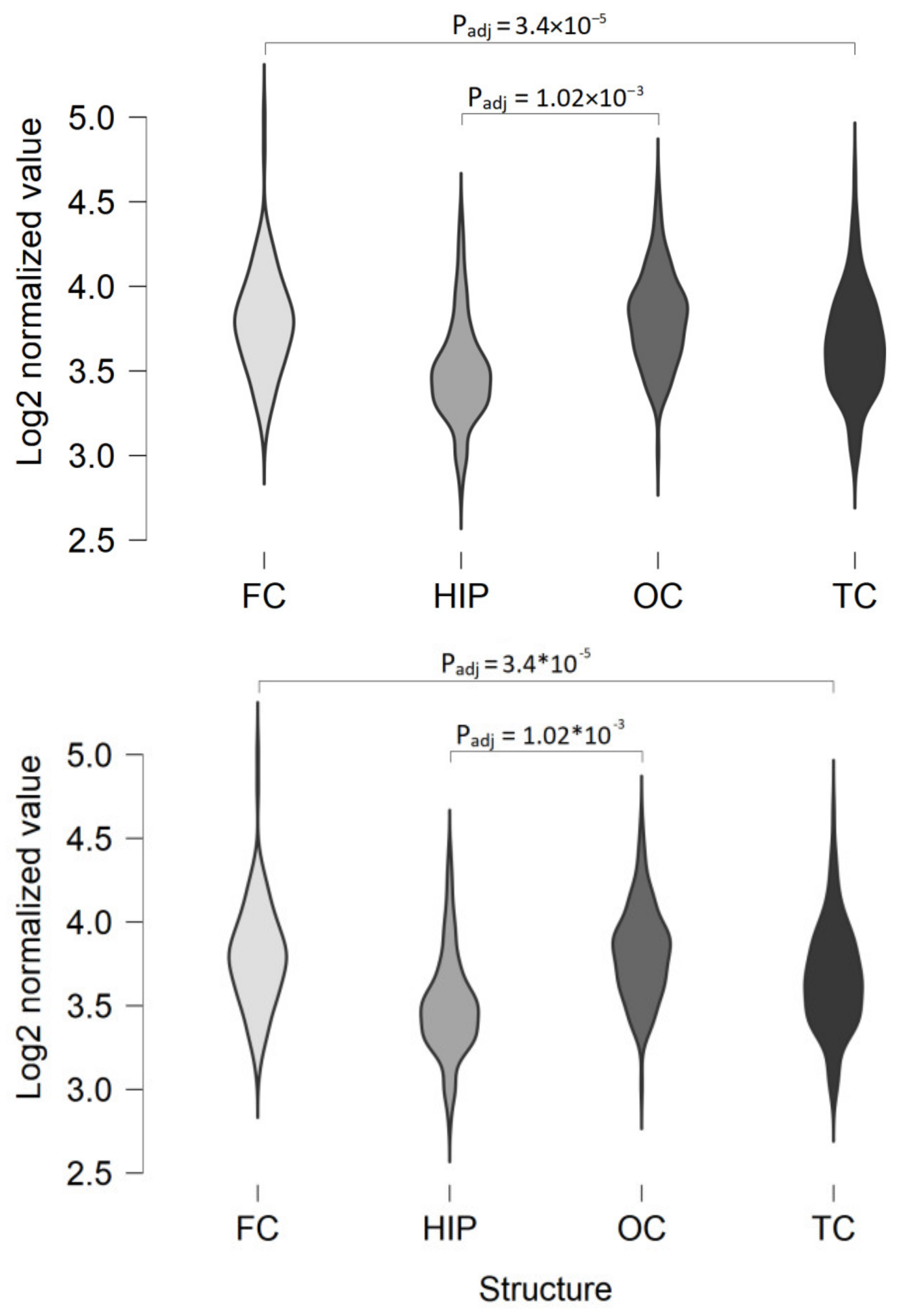
2.1. TAAR5 Expression in the Brain Cortical Areas and Hippocampus
2.2. TAAR5 Expression in the Amygdala and Basal Ganglia
2.3. TAAR5 Expression in the Diencephalon Structures
2.4. TAAR5 Expression in the Brain Stem and Cerebellum
2.5. TAAR5 Expression in the White Matter
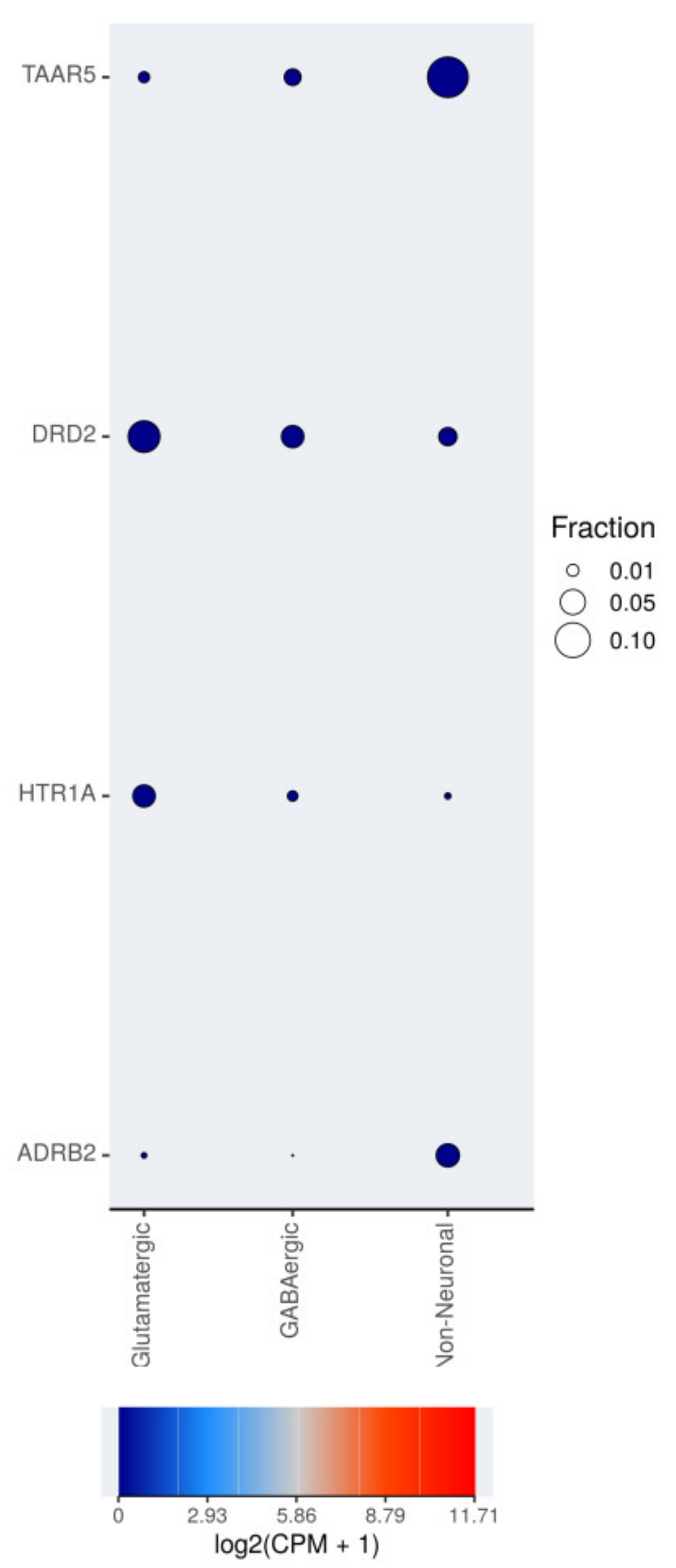
2.6. TAAR5 Expression at the Cell Level and Possible Sources of Bias
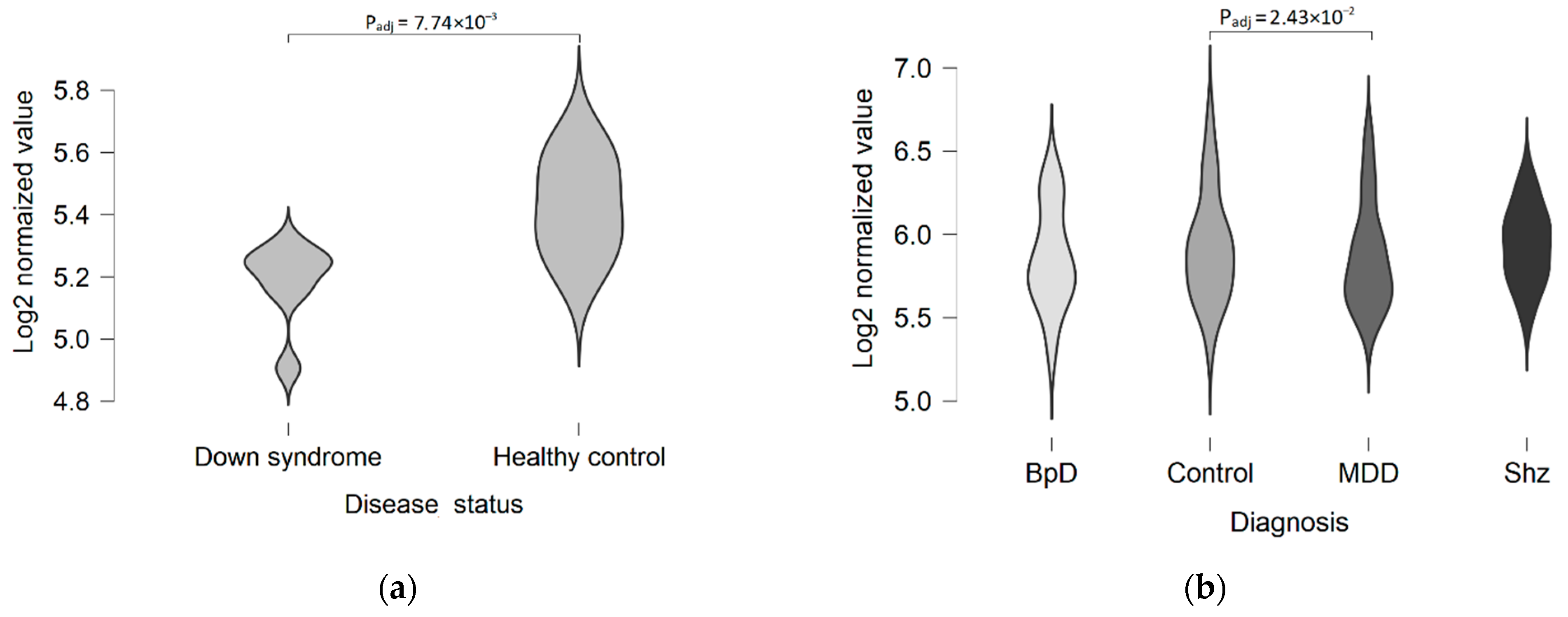
3. Discussion
4. Materials and Methods
4.1. Data Collection and Inclusion Criteria for Datasets
- -
- Taking into account low TAARs expression, datasets with small sample size were excluded to prevent sampling bias. The inclusion criteria were at least 5 samples from healthy adult subjects per structure in the dataset.
- -
- To escape the incorrect data comparison, the exact structure definition was applied as the inclusion criteria. Datasets with samples descriptions such as “brain sample” (with no any explanations of the studied brain part) or “cortex” (without clarifications such as “frontal cortex” or “insula”, etc.) were excluded.
- -
- Availability of log2 or non-logged expression values, including TAAR5 expression values.
- -
- As the majority of microarray-generated datasets were acquired on Affymetrix platforms, we excluded datasets generated on other kinds of microarray to include only comparable data in the review.
- -
- At least 5 samples from healthy adult subjects per structure in the dataset.
- -
- The exact structure definition (same as for microarray-generated datasets).
- -
- Because of the low TAARs mRNA transcription, its expression patterns may be estimated by RNAseq only if appropriate sequencing depth is applied. To prevent overload with false-negative results, at least 20 million reads in SRA file for each run, or mean reads number in SRA files in the dataset 40 million or higher, were applied as the threshold for dataset selection.
- -
- Availability of any quantitative data for TAAR5 expression in Supplementary Materials (count table or other attached data tables).
4.2. Microarray-Generated Datasets Analysis
4.3. RNAseq-Generated Datasets Analysis
4.4. Allen Brain Atlas Datasets
4.5. Human Protein Atlas
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Liberles, S.D.; Buck, L.B. A Second Class of Chemosensory Receptors in the Olfactory Epithelium. Nature 2006, 442, 645–650. [Google Scholar] [CrossRef] [PubMed]
- Berry, M.D.; Gainetdinov, R.R.; Hoener, M.C.; Shahid, M. Pharmacology of Human Trace Amine-Associated Receptors: Therapeutic Opportunities and Challenges. Pharmacol. Ther. 2017, 180, 161–180. [Google Scholar] [CrossRef]
- Gainetdinov, R.R.; Hoener, M.C.; Berry, M.D. Trace Amines and Their Receptors. Pharmacol. Rev. 2018, 70, 549–620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dewan, A. Olfactory Signaling via Trace Amine-Associated Receptors. Cell Tissue Res. 2021, 383, 395–407. [Google Scholar] [CrossRef] [PubMed]
- Espinoza, S.; Sukhanov, I.; Efimova, E.V.; Kozlova, A.; Antonova, K.A.; Illiano, P.; Leo, D.; Merkulyeva, N.; Kalinina, D.; Musienko, P.; et al. Trace Amine-Associated Receptor 5 Provides Olfactory Input Into Limbic Brain Areas and Modulates Emotional Behaviors and Serotonin Transmission. Front. Mol. Neurosci. 2020, 13, 18. [Google Scholar] [CrossRef] [PubMed]
- Efimova, E.V.; Kozlova, A.A.; Razenkova, V.; Katolikova, N.V.; Antonova, K.A.; Sotnikova, T.D.; Merkulyeva, N.S.; Veshchitskii, A.S.; Kalinina, D.S.; Korzhevskii, D.E.; et al. Increased Dopamine Transmission and Adult Neurogenesis in Trace Amine-Associated Receptor 5 (TAAR5) Knockout Mice. Neuropharmacology 2021, 182, 108373. [Google Scholar] [CrossRef]
- Zeng, Z.; Fan, P.; Rand, E.; Kyaw, H.; Su, K.; Madike, V.; Carter, K.C.; Li, Y. Cloning of a Putative Human Neurotransmitter Receptor Expressed in Skeletal Muscle and Brain. Biochem. Biophys. Res. Commun. 1998, 242, 575–578. [Google Scholar] [CrossRef]
- Carnicelli, V.; Santoro, A.; Sellari-Franceschini, S.; Berrettini, S.; Zucchi, R. Expression of Trace Amine-Associated Receptors in Human Nasal Mucosa. Chem. Percept. 2010, 3, 99–107. [Google Scholar] [CrossRef]
- Horowitz, L.F.; Saraiva, L.R.; Kuang, D.; Yoon, K.; Buck, L.B. Olfactory Receptor Patterning in a Higher Primate. J. Neurosci. 2014, 34, 12241–12252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gisladottir, R.S.; Ivarsdottir, E.V.; Helgason, A.; Jonsson, L.; Hannesdottir, N.K.; Rutsdottir, G.; Arnadottir, G.A.; Skuladottir, A.; Jonsson, B.A.; Norddahl, G.L.; et al. Sequence Variants in TAAR5 and Other Loci Affect Human Odor Perception and Naming. Curr. Biol. 2020, 30, 4643–4653.e3. [Google Scholar] [CrossRef] [PubMed]
- Scanlan, T.S.; Suchland, K.L.; Hart, M.E.; Chiellini, G.; Huang, Y.; Kruzich, P.J.; Frascarelli, S.; Crossley, D.A.; Bunzow, J.R.; Ronca-Testoni, S.; et al. 3-Iodothyronamine Is an Endogenous and Rapid-Acting Derivative of Thyroid Hormone. Nat. Med. 2004, 10, 638–642. [Google Scholar] [CrossRef]
- Zucchi, R.; Accorroni, A.; Chiellini, G. Update on 3-Iodothyronamine and Its Neurological and Metabolic Actions. Front. Physiol. 2014, 5, 402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, M.Z.; Nawaz, W. The Emerging Roles of Human Trace Amines and Human Trace Amine-Associated Receptors (HTAARs) in Central Nervous System. Biomed. Pharmacother. 2016, 83, 439–449. [Google Scholar] [CrossRef] [PubMed]
- Eyun, S. Accelerated Pseudogenization of Trace Amine-associated Receptor Genes in Primates. Genes Brain Behav. 2019, 18, e12543. [Google Scholar] [CrossRef] [PubMed]
- Staubert, C.; Böselt, I.; Bohnekamp, J.; Römpler, H.; Enard, W.; Schöneberg, T. Structural and Functional Evolution of the Trace Amine-Associated Receptors TAAR3, TAAR4 and TAAR5 in Primates. PLoS ONE 2010, 5, e11133. [Google Scholar] [CrossRef] [Green Version]
- Dinter, J.; Mühlhaus, J.; Wienchol, C.L.; Yi, C.-X.; Nürnberg, D.; Morin, S.; Grüters, A.; Köhrle, J.; Schöneberg, T.; Tschöp, M.; et al. Inverse Agonistic Action of 3-Iodothyronamine at the Human Trace Amine-Associated Receptor 5. PLoS ONE 2015, 10, e0117774. [Google Scholar] [CrossRef] [Green Version]
- Wallrabenstein, I.; Kuklan, J.; Weber, L.; Zborala, S.; Werner, M.; Altmüller, J.; Becker, C.; Schmidt, A.; Hatt, H.; Hummel, T.; et al. Human Trace Amine-Associated Receptor TAAR5 Can Be Activated by Trimethylamine. PLoS ONE 2013, 8, e54950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Pacifico, R.; Cawley, D.; Feinstein, P.; Bozza, T. Ultrasensitive Detection of Amines by a Trace Amine-Associated Receptor. J. Neurosci. 2013, 33, 3228–3239. [Google Scholar] [CrossRef]
- Aleksandrov, A.A.; Knyazeva, V.M.; Volnova, A.B.; Dmitrieva, E.S.; Korenkova, O.; Espinoza, S.; Gerasimov, A.; Gainetdinov, R.R. Identification of TAAR5 Agonist Activity of Alpha-NETA and Its Effect on Mismatch Negativity Amplitude in Awake Rats. Neurotox Res. 2018, 34, 442–451. [Google Scholar] [CrossRef]
- Aleksandrov, A.A.; Polyakova, N.V.; Vinogradova, E.P.; Gainetdinov, R.R.; Knyazeva, V.M. The TAAR5 Agonist α-NETA Causes Dyskinesia in Mice. Neurosci. Lett. 2019, 704, 208–211. [Google Scholar] [CrossRef] [PubMed]
- Aleksandrov, A.A.; Knyazeva, V.M.; Volnova, A.B.; Dmitrieva, E.S.; Polyakova, N.V. Putative TAAR5 Agonist Alpha-NETA Affects Event-Related Potentials in Oddball Paradigm in Awake Mice. Brain Res. Bull. 2020, 158, 116–121. [Google Scholar] [CrossRef]
- Belov, D.R.; Efimova, E.V.; Fesenko, Z.S.; Antonova, K.A.; Kolodyazhny, S.F.; Lakstygal, A.M.; Gainetdinov, R.R. Putative Trace-Amine Associated Receptor 5 (TAAR5) Agonist α-NETA Increases Electrocorticogram Gamma-Rhythm in Freely Moving Rats. Cell Mol. Neurobiol. 2020, 40, 203–213. [Google Scholar] [CrossRef] [PubMed]
- Rutigliano, G.; Zucchi, R. Molecular Variants in Human Trace Amine-Associated Receptors and Their Implications in Mental and Metabolic Disorders. Cell Mol. Neurobiol. 2020, 40, 239–255. [Google Scholar] [CrossRef] [Green Version]
- Uhlen, M.; Fagerberg, L.; Hallstrom, B.M.; Lindskog, C.; Oksvold, P.; Mardinoglu, A.; Sivertsson, A.; Kampf, C.; Sjostedt, E.; Asplund, A.; et al. Tissue-Based Map of the Human Proteome. Science 2015, 347, 1260419. [Google Scholar] [CrossRef]
- Hawrylycz, M.J.; Lein, E.S.; Guillozet-Bongaarts, A.L.; Shen, E.H.; Ng, L.; Miller, J.A.; van de Lagemaat, L.N.; Smith, K.A.; Ebbert, A.; Riley, Z.L.; et al. An Anatomically Comprehensive Atlas of the Adult Human Brain Transcriptome. Nature 2012, 489, 391–399. [Google Scholar] [CrossRef]
- Miller, J.A.; Ding, S.-L.; Sunkin, S.M.; Smith, K.A.; Ng, L.; Szafer, A.; Ebbert, A.; Riley, Z.L.; Royall, J.J.; Aiona, K.; et al. Transcriptional Landscape of the Prenatal Human Brain. Nature 2014, 508, 199–206. [Google Scholar] [CrossRef] [PubMed]
- Rinn, J.L.; Rozowsky, J.S.; Laurenzi, I.J.; Petersen, P.H.; Zou, K.; Zhong, W.; Gerstein, M.; Snyder, M. Major Molecular Differences between Mammalian Sexes Are Involved in Drug Metabolism and Renal Function. Dev. Cell 2004, 6, 791–800. [Google Scholar] [CrossRef] [Green Version]
- Allen Cell Types Database (2015), RNAseq Data Navigator. Available online: http://www.celltypes.brain-map.org/rnaseq/human (accessed on 10 March 2021).
- Zaghlool, A.; Niazi, A.; Björklund, Å.K.; Westholm, J.O.; Ameur, A.; Feuk, L. Characterization of the Nuclear and Cytosolic Transcriptomes in Human Brain Tissue Reveals New Insights into the Subcellular Distribution of RNA Transcripts. Sci. Rep. 2021, 11, 4076. [Google Scholar] [CrossRef]
- Gaudel, F.; Guiraudie-Capraz, G.; Féron, F. Limbic Expression of MRNA Coding for Chemoreceptors in Human Brain—Lessons from Brain Atlases. Int. J. Mol. Sci. 2021, 22, 6858. [Google Scholar] [CrossRef] [PubMed]
- Pacifico, R.; Dewan, A.; Cawley, D.; Guo, C.; Bozza, T. An Olfactory Subsystem That Mediates High-Sensitivity Detection of Volatile Amines. Cell Rep. 2012, 2, 76–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuvarzin, S.; Efimova, E.; Mor, M.; Katolikova, N.; Smirnova, D.; Kozlova, A.; Gainetdinov, R. Trace Amine-Associated Receptor 2: Brain Expression and It’s Role in Neurochemistry and Behaviour. Neurochemistry 2020, 40, S41–S42. [Google Scholar] [CrossRef]
- Gurevich, E.V.; Joyce, J.N. Distribution of Dopamine D3 Receptor Expressing Neurons in the Human Forebrain: Comparison with D2 Receptor Expressing Neurons. Neuropsychopharmacology 1999, 20, 60–80. [Google Scholar] [CrossRef] [Green Version]
- Woodward, N.D.; Zald, D.H.; Ding, Z.; Riccardi, P.; Ansari, M.S.; Baldwin, R.M.; Cowan, R.L.; Li, R.; Kessler, R.M. Cerebral Morphology and Dopamine D2/D3 Receptor Distribution in Humans: A Combined [18F]Fallypride and Voxel-Based Morphometry Study. Neuroimage 2009, 46, 31–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Birioukova, L.M.; Midzyanovskaya, I.S.; Lensu, S.; Tuomisto, L.; van Luijtelaar, G. Distribution of D1-like and D2-like Dopamine Receptors in the Brain of Genetic Epileptic WAG/Rij Rats. Epilepsy Res. 2005, 63, 89–96. [Google Scholar] [CrossRef]
- Wei, X.; Ma, T.; Cheng, Y.; Huang, C.C.Y.; Wang, X.; Lu, J.; Wang, J. Dopamine D1 or D2 Receptor-Expressing Neurons in the Central Nervous System. Addict. Biol. 2018, 23, 569–584. [Google Scholar] [CrossRef] [PubMed]
- Sung Choi, W.; Machida, C.A.; Ronnekleiv, O.K. Distribution of Dopamine D1, D2, and D5 Receptor MRNAs in the Monkey Brain: Ribonuclease Protection Assay Analysis. Mol. Brain Res. 1995, 31, 86–94. [Google Scholar] [CrossRef]
- Van Waarde, A.; Vaalburg, W.; Doze, P.; Bosker, F.J.; Elsinga, P.H. PET Imaging of Beta-Adrenoceptors in Human Brain: A Realistic Goal or a Mirage? Curr. Pharm. Des. 2004, 10, 1519–1536. [Google Scholar] [CrossRef] [Green Version]
- Burnet, P.W.J.; Eastwood, S.L.; Lacey, K.; Harrison, P.J. The Distribution of 5-HT1A and 5-HT2A Receptor MRNA in Human Brain. Brain Res. 1995, 676, 157–168. [Google Scholar] [CrossRef]
- Hall, H.; Lundkvist, C.; Halldin, C.; Farde, L.; Pike, V.W.; McCarron, J.A.; Fletcher, A.; Cliffe, I.A.; Barf, T.; Wikström, H.; et al. Autoradiographic Localization of 5-HT1A Receptors in the Post-Mortem Human Brain Using [3H]WAY-100635 and [11C]Way-100635. Brain Res. 1997, 745, 96–108. [Google Scholar] [CrossRef]
- Knowles, R.B.; Sabry, J.H.; Martone, M.E.; Deerinck, T.J.; Ellisman, M.H.; Bassell, G.J.; Kosik, K.S. Translocation of RNA Granules in Living Neurons. J. Neurosci. 1996, 16, 7812–7820. [Google Scholar] [CrossRef] [Green Version]
- Giuditta, A.; Kaplan, B.B.; Minnen, J.v.; Alvarez, J.; Koenig, E. Axonal and Presynaptic Protein Synthesis: New Insights into the Biology of the Neuron. Trends Neurosci. 2002, 25, 400–404. [Google Scholar] [CrossRef]
- Aleksandrov, A.A.; Dmitrieva, E.S.; Volnova, A.B.; Knyazeva, V.M.; Polyakova, N.V.; Ptukha, M.A.; Gainetdinov, R.R. Effect of Alpha-NETA on Auditory Event Related Potentials in Sensory Gating Study Paradigm in Mice. Neurosci. Lett. 2019, 712, 134470. [Google Scholar] [CrossRef]
- Harmer, C.J.; Duman, R.S.; Cowen, P.J. How Do Antidepressants Work? New Perspectives for Refining Future Treatment Approaches. Lancet Psychiatry 2017, 4, 409–418. [Google Scholar] [CrossRef] [Green Version]
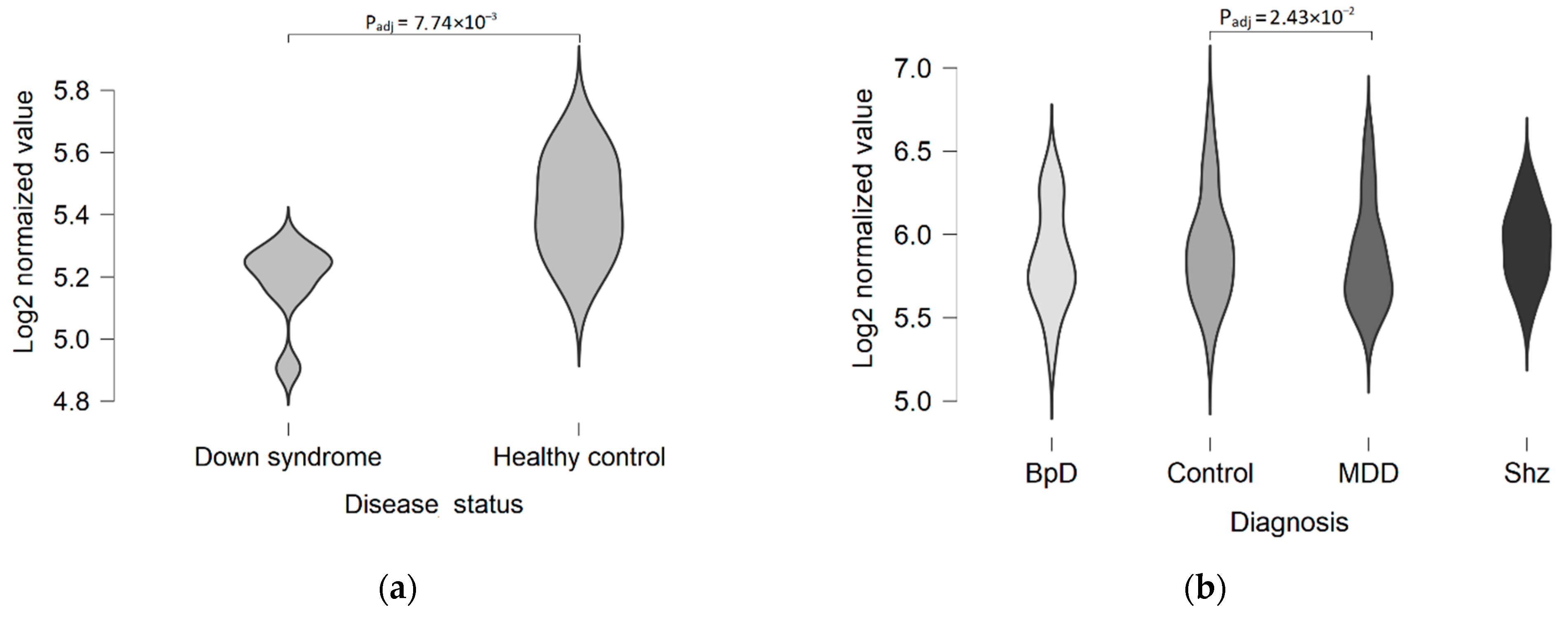
- Mann, D.M.; Yates, P.O.; Marcyniuk, B. Dopaminergic Neurotransmitter Systems in Alzheimer’s Disease and in Down’s Syndrome at Middle Age. J. Neurol. Neurosurg. Psychiatry 1987, 50, 341–344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Capone, G.T. Down Syndrome: Advances in Molecular Biology and the Neurosciences. J. Dev. Behav. Pediatr. 2001, 22, 40–59. [Google Scholar] [CrossRef] [Green Version]
- Ponnusamy, R.; McNerney, M.W.; Moghadam, S.; Salehi, A. Assessing Disease-Modifying Effects of Norepinephrine in Down Syndrome and Alzheimer’s Disease. Brain Res. 2019, 1702, 3–11. [Google Scholar] [CrossRef]
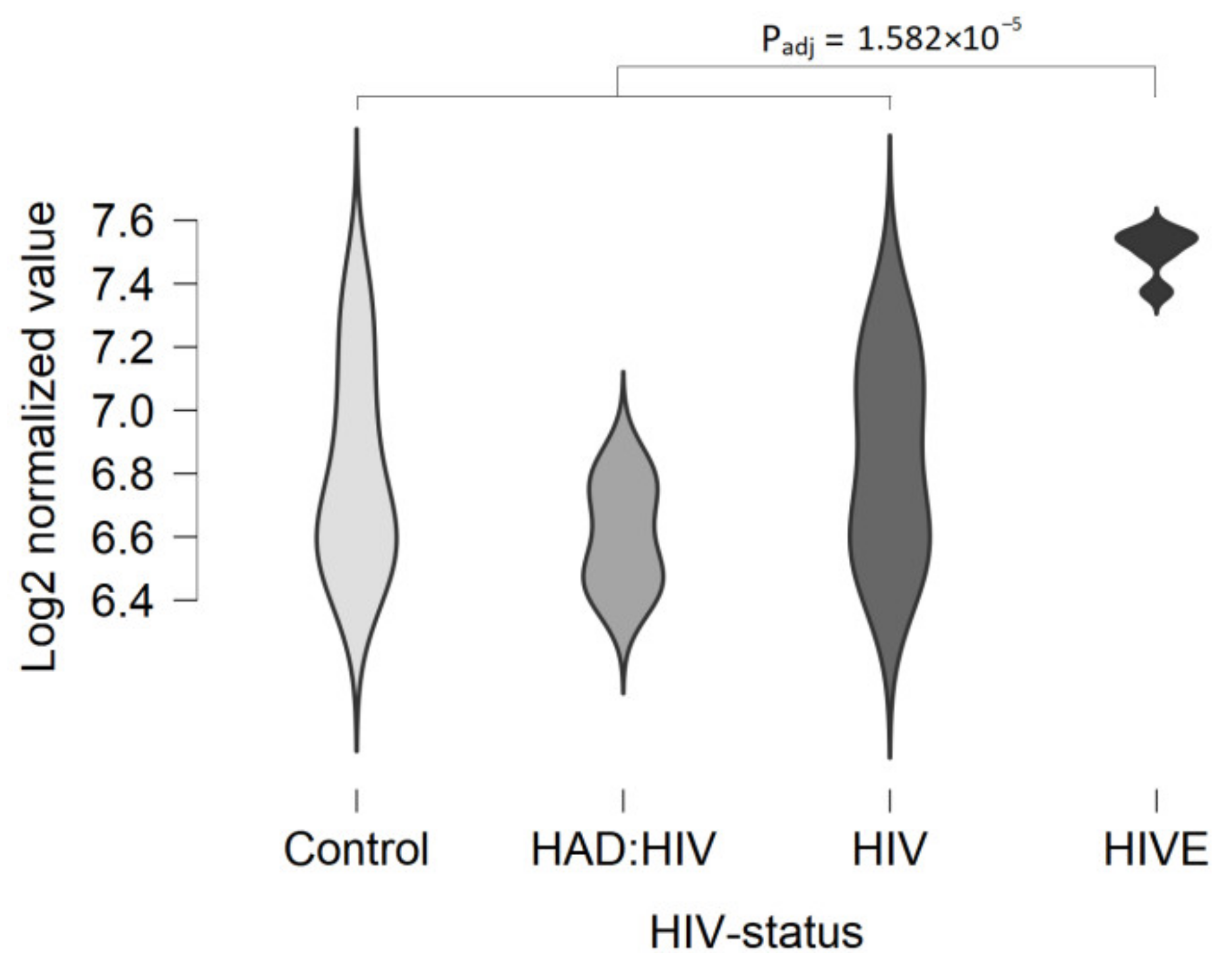
- Gelman, B.B.; Chen, T.; Lisinicchia, J.G.; Soukup, V.M.; Carmical, J.R.; Starkey, J.M.; Masliah, E.; Commins, D.L.; Brandt, D.; Grant, I.; et al. The National NeuroAIDS Tissue Consortium Brain Gene Array: Two Types of HIV-Associated Neurocognitive Impairment. PLoS ONE 2012, 7, e46178. [Google Scholar] [CrossRef]
- Siangphoe, U.; Archer, K.J. Gene Expression in HIV-Associated Neurocognitive Disorders: A Meta-Analysis. JAIDS J. Acquir. Immune Defic. Syndr. 2015, 70, 479–488. [Google Scholar] [CrossRef] [PubMed]
- Nelson, D.A.; Tolbert, M.D.; Singh, S.J.; Bost, K.L. Expression of Neuronal Trace Amine-Associated Receptor (Taar) MRNAs in Leukocytes. J. Neuroimmunol. 2007, 192, 21–30. [Google Scholar] [CrossRef] [Green Version]
- Barrett, T.; Wilhite, S.E.; Ledoux, P.; Evangelista, C.; Kim, I.F.; Tomashevsky, M.; Marshall, K.A.; Phillippy, K.H.; Sherman, P.M.; Holko, M.; et al. NCBI GEO: Archive for Functional Genomics Data Sets—Update. Nucleic Acids Res. 2013, 41, D991–D995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- GEO Browser. Available online: http://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/geo/browse/ (accessed on 17 March 2021).
- Phantasus. Available online: http://genome.ifmo.ru/phantasus-dev/ (accessed on 20 March 2021).
- Ritchie, M.E.; Phipson, B.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. Limma Powers Differential Expression Analyses for RNA-Sequencing and Microarray Studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef]
- GEO2R. Available online: http://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/geo/geo2r/ (accessed on 20 March 2021).
- Huang, R.S.; Duan, S.; Shukla, S.J.; Kistner, E.O.; Clark, T.A.; Chen, T.X.; Schweitzer, A.C.; Blume, J.E.; Dolan, M.E. Identification of Genetic Variants Contributing to Cisplatin-Induced Cytotoxicity by Use of a Genomewide Approach. Am. J. Hum. Genet. 2007, 81, 427–437. [Google Scholar] [CrossRef] [Green Version]
- Kernagis, D.N.; Hall, A.H.S.; Datto, M.B. Genes with Bimodal Expression Are Robust Diagnostic Targets That Define Distinct Subtypes of Epithelial Ovarian Cancer with Different Overall Survival. J. Mol. Diagn. 2012, 14, 214–222. [Google Scholar] [CrossRef] [PubMed]
- Mulla, W.; Hajaj, B.; Elyagon, S.; Mor, M.; Gillis, R.; Murninkas, M.; Klapper-Goldstein, H.; Plaschkes, I.; Chalifa-Caspi, V.; Etzion, S.; et al. Rapid Atrial Pacing Promotes Atrial Fibrillation Substrate in Unanesthetized Instrumented Rats. Front. Physiol. 2019, 10, 1218. [Google Scholar] [CrossRef] [PubMed]
- JASP Team. JASP [Computer Software]; JASP: Amsterdam, The Netherlands, 2020. [Google Scholar]
- Mahi, N.A.; Najafabadi, M.F.; Pilarczyk, M.; Kouril, M.; Medvedovic, M. GREIN: An Interactive Web Platform for Re-Analyzing GEO RNA-Seq Data. Sci. Rep. 2019, 9, 7580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- GREIN. Available online: http://www.ilincs.org/apps/grein/?gse= (accessed on 23 March 2021).
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. EdgeR: A Bioconductor Package for Differential Expression Analysis of Digital Gene Expression Data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papatheodorou, I.; Moreno, P.; Manning, J.; Fuentes, A.M.-P.; George, N.; Fexova, S.; Fonseca, N.A.; Füllgrabe, A.; Green, M.; Huang, N.; et al. Expression Atlas Update: From Tissues to Single Cells. Nucleic Acids Res. 2020, 48, D77–D83. [Google Scholar] [CrossRef] [Green Version]
- Grote, S.; Prüfer, K.; Kelso, J.; Dannemann, M. ABAEnrichment: An R Package to Test for Gene Set Expression Enrichment in the Adult and Developing Human Brain. Bioinformatics 2016, 32, 3201–3203. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
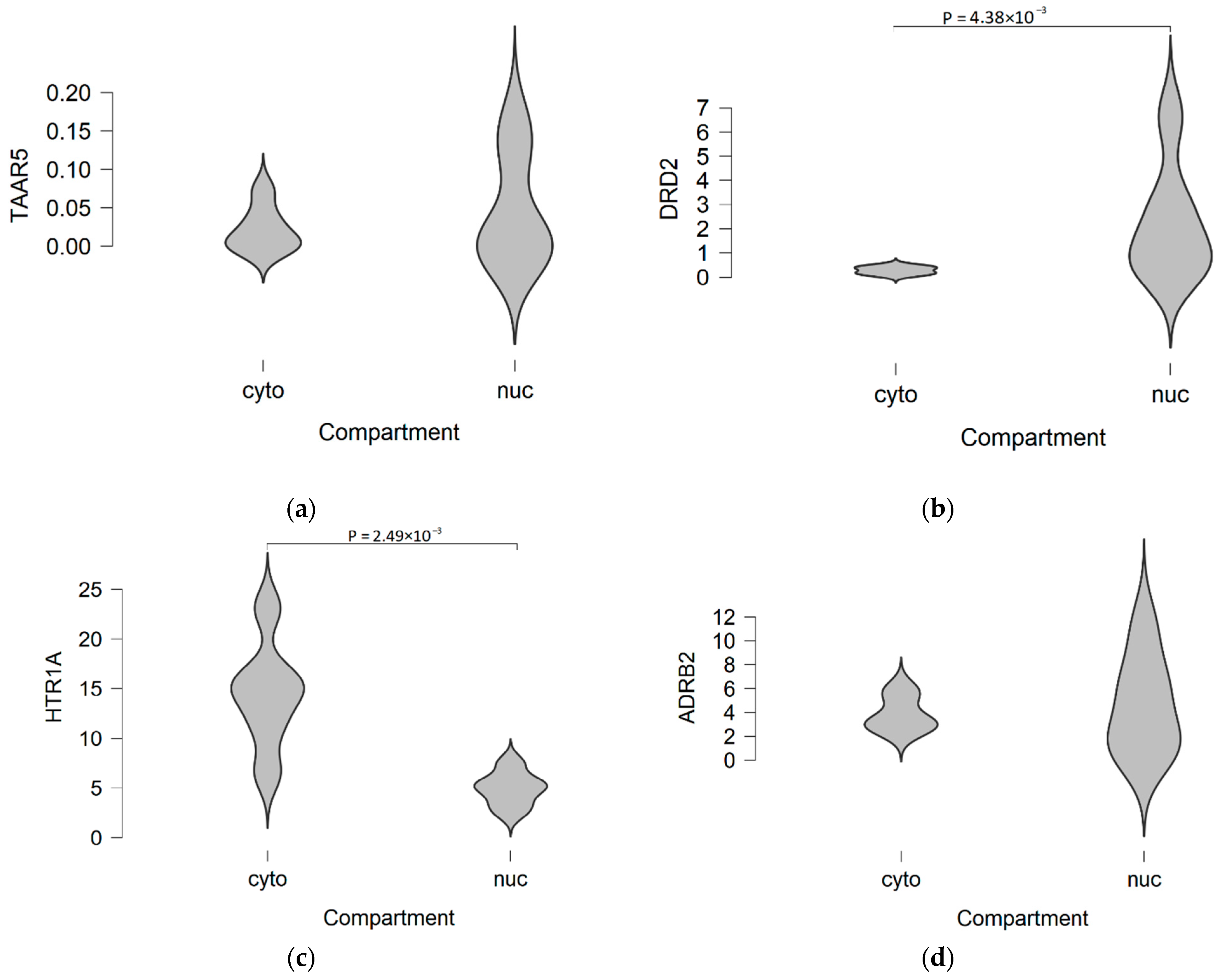{kind=link}
| Structure | Microarray | RNAseq | Public Databases |
|---|---|---|---|
| Cortical areas: frontal cortex, parietal cortex, temporal cortex, occipital cortex, cingulate cortex, and insula. | The expression level is over the cut-off value in some specimens (log2 values > 5.0). Moreover, some statistically significant variations between cortex areas or health conditions are detected. | In several datasets, some samples are true positive for TAAR5 expression (CPM > 0.5). | HPA data demonstrated accidental TAAR5 expression below the cut-off recommended for this database. |
| Hippocampus | The expression level is over the cut-off value in some specimens (log2 values > 5.0). | 30% (3 of 10) true positives for TAAR5 expression (CPM > 0.5) in GSE68559. | In the ABA, CA4 field samples may be considered positive. |
| Amygdala | In several datasets, some samples are true positives for TAAR5 expression (CPM > 0.5) | Positive TAAR5 expression is demonstrated in the BrainSpan Atlas of the Developing Human Brain and HPA (below cut-off). | |
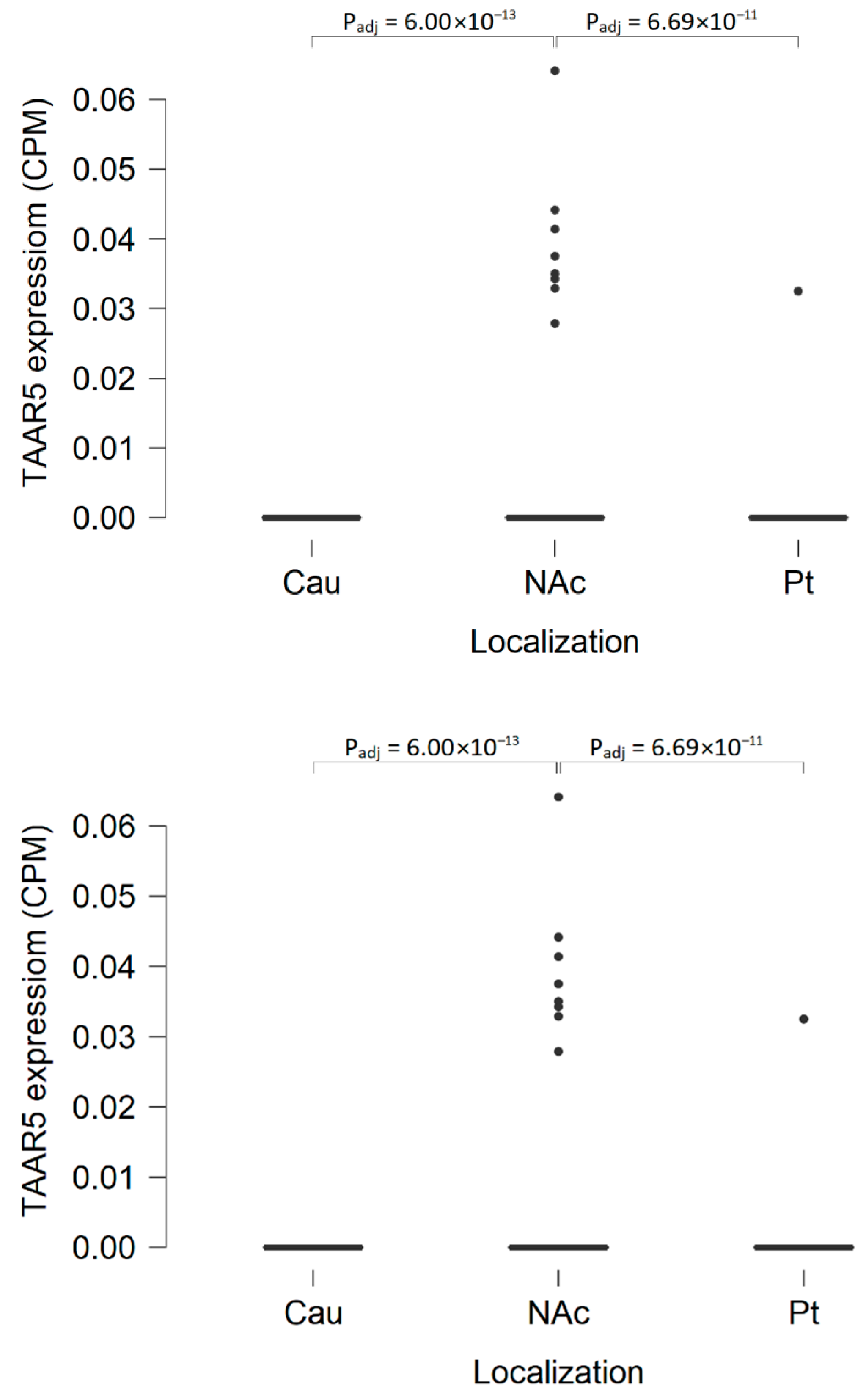
| Nucleus accumbens | More pronounced TAAR5 expression in the nucleus accumbens than in the caudate nucleus in GSE160521 (taking into account that TAAR5 is expressed at the levels below 0.5 CPM). | ||
| Thalamus | In the ABA, some thalamic structures may be considered positive. | ||
| Hypothalamus | The expression level is over the cut-off value in some specimens (log2 values > 5.0). | In the ABA, some hypothalamic structures may be considered positive. | |
| Striatum | The expression level is over the cut-off value in some specimens (log2 values > 5.0). | In several datasets, some samples are true positive for TAAR5 expression (CPM > 0.5). | |
| Cerebellum | The expression level is over the cut-off value in some specimens (log2 values > 5.0). | In several datasets, some samples are true positive for TAAR5 expression (CPM > 0.5). | HPA data demonstrated accidental TAAR5 expression below the cut-off recommended for this database. In the ABA, some cerebellar areas may be considered positive. |
| Substantia nigra | The expression level is over the cut-off value in some specimens (log2 values > 5.0). | Positive expression was detected in the ABA dataset. | |
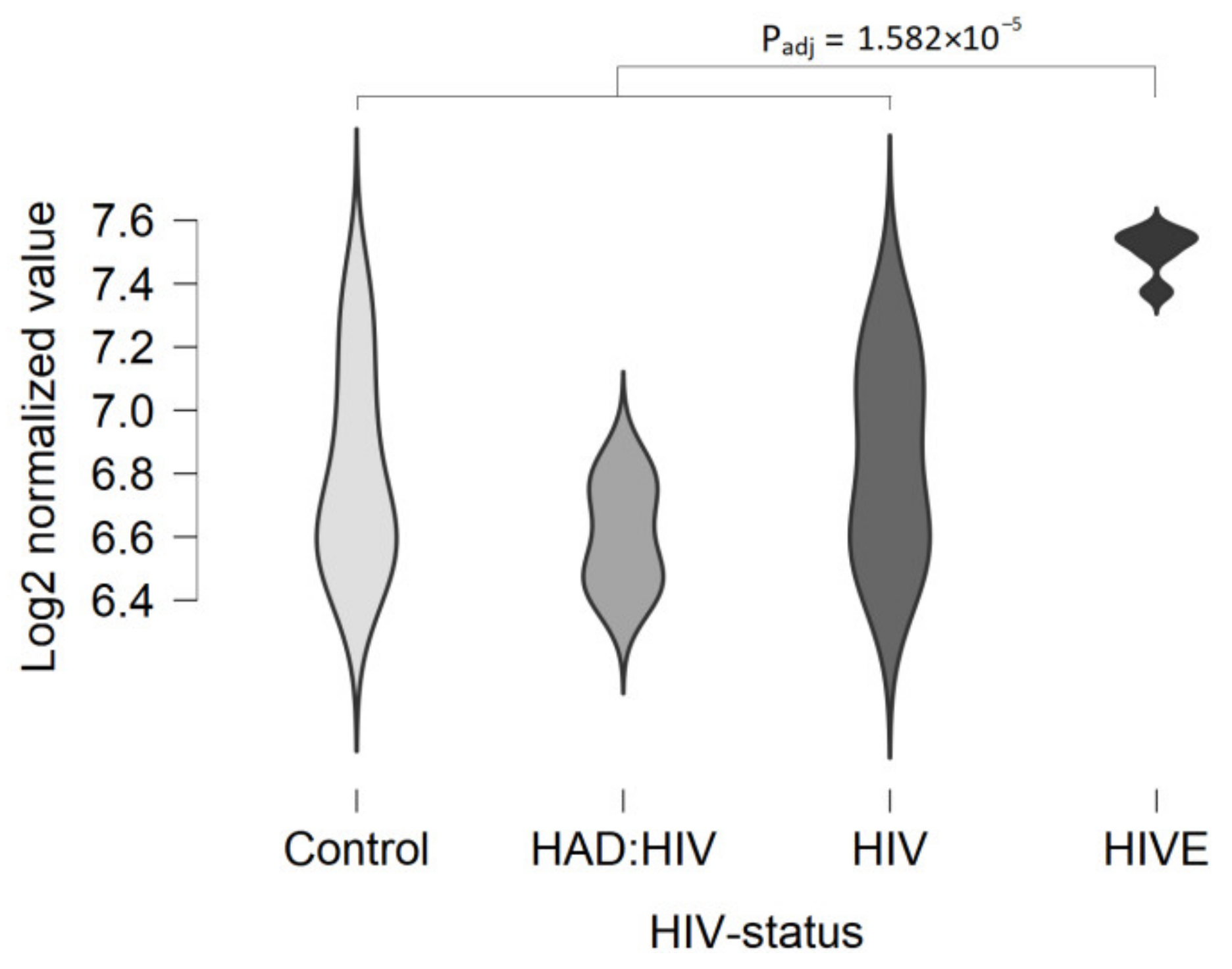
| White matter | The expression level is over the cut-off value in some specimens (log2 values > 5.0). The up-regulation of expression in HIV-associated encephalitis is statistically significant. | In several datasets, some samples are true positive for TAAR5 expression (CPM > 0.5). | |
| Source of bias | The method is not adopted to confirm the absence or presence of mRNA, is not the purpose is the differential gene expression examination. | The sensitivity of the study depends on its design. In some datasets, the number of reads in the SRA file varies from 20 to 100 million. Such datasets are highly heterogeneous. Samples that include only 20 million reads may be inadequate to estimate low-expressed genes. | Data were received by RNAseq or microarray and are not free from corresponding weaknesses. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vaganova, A.N.; Murtazina, R.Z.; Shemyakova, T.S.; Prjibelski, A.D.; Katolikova, N.V.; Gainetdinov, R.R. Pattern of TAAR5 Expression in the Human Brain Based on Transcriptome Datasets Analysis. Int. J. Mol. Sci. 2021, 22, 8802. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22168802
Vaganova AN, Murtazina RZ, Shemyakova TS, Prjibelski AD, Katolikova NV, Gainetdinov RR. Pattern of TAAR5 Expression in the Human Brain Based on Transcriptome Datasets Analysis. International Journal of Molecular Sciences. 2021; 22(16):8802. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22168802
Chicago/Turabian StyleVaganova, Anastasia N., Ramilya Z. Murtazina, Taisiia S. Shemyakova, Andrey D. Prjibelski, Nataliia V. Katolikova, and Raul R. Gainetdinov. 2021. "Pattern of TAAR5 Expression in the Human Brain Based on Transcriptome Datasets Analysis" International Journal of Molecular Sciences 22, no. 16: 8802. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22168802