
The Impact of Physical Exercise on the Circulating Levels of BDNF and NT 4/5: A Review

 ,
,  , ,
, ,  and
and
Abstract
:1. Introduction
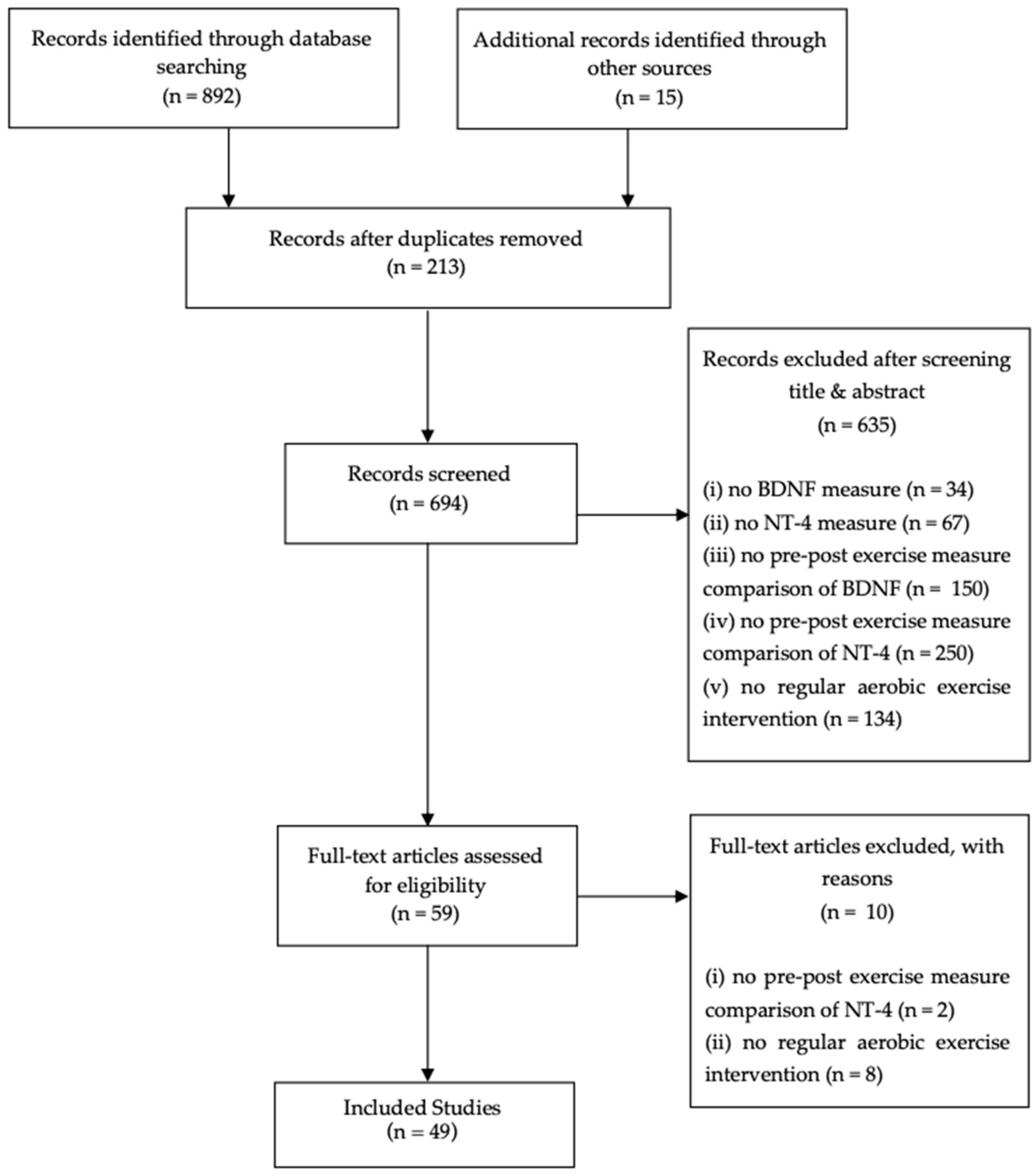
2. Methods
Data Collection, Extraction, and Analysis
3. Results
3.1. Relationship between a Chronic Aerobic Exercise Intervention and the Variation of BDNF Blood Concentration Levels in Human Models
3.2. Relationship between a Chronic Aerobic Exercise Intervention and the Variation of BDNF Blood Concentration Levels in Animal Models
3.3. Relationship between a Chronic Aerobic Exercise Intervention and the Variation of NT-4/5 Blood Concentration Levels in Animal Models
4. Discussion
4.1. BDNF
4.2. NT-4/5
4.3. Physical Exercise & Neurotrophic Factors
4.4. BDNF & Aerobic Exercise
4.5. NT-4/5 and Aerobic Exercise
4.6. The Impact of Aerobic Exercise on Circulating Neurotrophin Levels (BDNF and NT-4/5) from Cerebral and Peripheral Circulation: Which Organs Are the Main Contributors?
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dishman, R.K.; Berthoud, H.R.; Booth, F.W.; Cotman, C.W.; Edgerton, V.R.; Fleshner, M.R.; Gandevia, S.C.; Gomez-Pinilla, F.; Greenwood, B.N.; Hillman, C.H.; et al. Neurobiology of exercise. Obesity (Silver Spring) 2006, 14, 345–356. [Google Scholar] [CrossRef]
- Zoladz, J.A.; Pilc, A. The effect of physical activity on the brain derived neurotrophic factor: From animal to human studies. J. Physiol. Pharmacol. Off. J. Pol. Physiol. Soc. 2010, 61, 533–541. [Google Scholar]
- Zembron-Lacny, A.; Dziubek, W.; Rynkiewicz, M.; Morawin, B.; Woźniewski, M. Peripheral brain-derived neurotrophic factor is related to cardiovascular risk factors in active and inactive elderly men. Braz. J. Med. Biol. Res. 2016, 49. [Google Scholar] [CrossRef] [Green Version]
- Knaepen, K.; Goekint, M.; Heyman, E.M.; Meeusen, R. Neuroplasticity–exercise-induced response of peripheral brain-derived neurotrophic factor: A systematic review of experimental studies in human subjects. Sports Med. 2010, 40, 765–801. [Google Scholar] [CrossRef] [PubMed]
- Cotman, C.W.; Berchtold, N.C.; Christie, L.-A. Exercise builds brain health: Key roles of growth factor cascades and inflammation. Trends Neurosci. 2007, 30, 464–472. [Google Scholar] [CrossRef] [PubMed]
- Sleiman, S.F.; Chao, M.V. Downstream Consequences of Exercise Through the Action of BDNF. Brain Plast. 2015, 1, 143–148. [Google Scholar] [CrossRef] [Green Version]
- Friedman, W. Growth Factors; Brady, S., Siegel, G., Albers, W.R., Price, D., Eds.; Elsevier: Amsterdam, The Netherlands, 2012; pp. 546–557. [Google Scholar]
- Al-Qudah, M.A.; Al-Dwairi, A. Mechanisms and regulation of neurotrophin synthesis and secretion. Neurosciences 2016, 21, 306–313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lippi, G.; Mattiuzzi, C.; Sanchis-Gomar, F. Updated overview on interplay between physical exercise, neurotrophins, and cognitive function in humans. J. Sport Health Sci. 2019, 9, 74–81. [Google Scholar] [CrossRef]
- Kashyap, M.P.; Roberts, C.; Waseem, M.; Tyagi, P. Drug Targets in Neurotrophin Signaling in the Central and Peripheral Nervous System. Mol. Neurobiol. 2018, 55, 6939–6955. [Google Scholar] [CrossRef]
- Chung, J.Y.; Kim, M.W.; Bang, M.S.; Kim, M. Increased expression of neurotrophin 4 following focal cerebral ischemia in adult rat brain with treadmill exercise. PLoS ONE 2013, 8, e52461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hölscher, C. Growth Factors: Neuronal Atrophy. In Reference Module in Neuroscience and Biobehavioral Psychology; Laviola, G., Ed.; Elsevier: Amsterdam, The Netherlands, 2017; pp. 1–7. [Google Scholar]
- Sakuma, K.; Yamaguchi, A. The recent understanding of the neurotrophin’s role in skeletal muscle adaptation. J. Biomed. Biotechnol. 2011, 2011, 201696. [Google Scholar] [CrossRef] [Green Version]
- Sandrini, L.; Di Minno, A.; Amadio, P.; Ieraci, A.; Tremoli, E.; Barbieri, S.S. Association between Obesity and Circulating Brain-Derived Neurotrophic Factor (BDNF) Levels: Systematic Review of Literature and Meta-Analysis. Int. J. Mol. Sci. 2018, 19, 2281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sariola, H. The neurotrophic factors in non-neuronal tissues. Cell. Mol. Life Sci. 2001, 58, 1061–1066. [Google Scholar] [CrossRef]
- Jeon, Y.K.; Ha, C.H. The effect of exercise intensity on brain derived neurotrophic factor and memory in adolescents. Environ. Health Prev. Med. 2017, 22, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mackay, C.P.; Kuys, S.S.; Brauer, S.G. The Effect of Aerobic Exercise on Brain-Derived Neurotrophic Factor in People with Neurological Disorders: A Systematic Review and Meta-Analysis. Neural Plast. 2017, 2017, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maejima, H.; Kanemura, N.; Kokubun, T.; Murata, K.; Takayanagi, K. Exercise enhances cognitive function and neurotrophin expression in the hippocampus accompanied by changes in epigenetic programming in senescence-accelerated mice. Neurosci. Lett. 2018, 665, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Domínguez-Sanchéz, M.A.; Cruz, H.B.; Velasco-Orjuela, G.P.; Quintero, A.P.; Tordecilla-Sanders, A.; Correa-Bautista, J.E.; Triana-Reina, H.R.; García-Hermoso, A.; González-Ruíz, K.; Peña-Guzmán, C.A.; et al. Acute Effects of High Intensity, Resistance, or Combined Protocol on the Increase of Level of Neurotrophic Factors in Physically Inactive Overweight Adults: The BrainFit Study. Front. Physiol. 2018, 9, 741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pareja-Galeano, H.; Alis, R.; Sanchis-Gomar, F.; Cabo, H.; Cortell-Ballester, J.; Gomez-Cabrera, M.C.; Lucia, A.; Viña, J. Methodological considerations to determine the effect of exercise on brain-derived neurotrophic factor levels. Clin. Biochem. 2014, 48, 162–166. [Google Scholar] [CrossRef]
- Ogborn, D.I.; Gardiner, P.F. Effects of exercise and muscle type on BDNF, NT-4/5, and TrKB expression in skeletal muscle. Muscle Nerve 2009, 41, 385–391. [Google Scholar] [CrossRef]
- Pan, W.; Banks, W.A.; Kastin, A.J. Permeability of the blood–brain barrier to neurotrophins. Brain Res. 1998, 788, 87–94. [Google Scholar] [CrossRef]
- Pedersen, B.K. Physical activity and muscle–brain crosstalk. Nat. Rev. Endocrinol. 2019, 15, 383–392. [Google Scholar] [CrossRef] [PubMed]
- Moher, D.; Liberati, A.; Tetzlaff, J.; Altman, D.G.; Group, P. Preferred reporting items for systematic reviews and meta-analyses: The PRISMA Statement. Open Med. 2009, 3, e123–e130. [Google Scholar] [PubMed]
- Griffin, E.W.; Mullally, S.; Foley, C.; Warmington, S.A.; O’Mara, S.M.; Kelly, A.M. Aerobic exercise improves hippocampal function and increases BDNF in the serum of young adult males. Physiol. Behav. 2011, 104, 934–941. [Google Scholar] [CrossRef]
- Salehi, I.; Hosseini, S.M.; Haghighi, M.; Jahangard, L.; Bajoghli, H.; Gerber, M.; Pühse, U.; Holsboer-Trachsler, E.; Brand, S. Electroconvulsive therapy (ECT) and aerobic exercise training (AET) increased plasma BDNF and ameliorated depressive symptoms in patients suffering from major depressive disorder. J. Psychiatr. Res. 2016, 76, 1–8. [Google Scholar] [CrossRef]
- Zoladz, J.A.; Pilc, A.; Majerczak, J.; Grandys, M.; Zapart-Bukowska, J.; Duda, K. Endurance training increases plasma brain-derived neurotrophic factor concentration in young healthy men. J. Physiol. Pharmacol. Off. J. Pol. Physiol. Soc. 2008, 59 (Suppl. 7), 119–132. [Google Scholar]
- Kerling, A.; Kück, M.; Tegtbur, U.; Grams, L.; Weber-Spickschen, S.; Hanke, A.; Stubbs, B.; Kahl, K. Exercise increases serum brain-derived neurotrophic factor in patients with major depressive disorder. J. Affect. Disord. 2017, 215, 152–155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Babaei, P.; Alamdari, K.A.; Tehrani, B.S.; Damirchi, A. Effect of six weeks of endurance exercise and following detraining on serum brain derived neurotrophic factor and memory performance in middle aged males with metabolic syndrome. J. Sports Med. Phys. Fit. 2013, 53, 437–443. [Google Scholar]
- Damirchi, A.; Tehrani, B.S.; Alamdari, K.A.; Babaei, P. Influence of aerobic training and detraining on serum BDNF, insulin resistance, and metabolic risk factors in middle-aged men diagnosed with metabolic syndrome. Clin. J. Sport Med. Off. J. Can. Acad. Sport Med. 2014, 24, 513–518. [Google Scholar] [CrossRef] [PubMed]
- Wagner, G.; Herbsleb, M.; de la Cruz, F.; Schumann, A.; Köhler, S.; Puta, C.; Gabriel, H.W.; Reichenbach, J.R.; Bär, K.-J. Changes in fMRI activation in anterior hippocampus and motor cortex during memory retrieval after an intense exercise intervention. Biol. Psychol. 2017, 124, 65–78. [Google Scholar] [CrossRef]
- Castellano, V.; White, L.J. Serum brain-derived neurotrophic factor response to aerobic exercise in multiple sclerosis. J. Neurol. Sci. 2008, 269, 85–91. [Google Scholar] [CrossRef] [PubMed]
- Cho, S.Y.; Roh, H.T. Effects of aerobic exercise training on peripheral brain-derived neurotrophic factor and eotaxin-1 levels in obese young men. J. Phys. Ther. Sci. 2016, 28, 1355–1358. [Google Scholar] [CrossRef] [Green Version]
- El-Tamawy, M.S.; Abd-Allah, F.; Ahmed, S.M.; Darwish, M.H.; Khalifa, H.A. Aerobic exercises enhance cognitive functions and brain derived neurotrophic factor in ischemic stroke patients. NeuroRehabilitation 2014, 34, 209–213. [Google Scholar] [CrossRef] [PubMed]
- Goekint, M.; Roelands, B.; De Pauw, K.; Knaepen, K.; Bos, I.; Meeusen, R. Does a period of detraining cause a decrease in serum brain-derived neurotrophic factor? Neurosci. Lett. 2010, 486, 146–149. [Google Scholar] [CrossRef]
- Marusiak, J.; Zeligowska, E.; Mencel, J.; Kisiel-Sajewicz, K.; Majerczak, J.; Zoladz, J.A.; Jaskolski, A.; Jaskolska, A. Interval training-induced alleviation of rigidity and hypertonia in patients with Parkinson’s disease is accompanied by increased basal serum brain-derived neurotrophic factor. J. Rehabil. Med. 2015, 47, 372–375. [Google Scholar] [CrossRef] [Green Version]
- Roh, H.T.; So, W.Y. The effects of aerobic exercise training on oxidant-antioxidant balance, neurotrophic factor levels, and blood-brain barrier function in obese and non-obese men. J. Sport Health Sci. 2017, 6, 447–453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schulz, K.-H.; Gold, S.; Witte, J.; Bartsch, K.; Lang, U.E.; Hellweg, R.; Reer, R.; Braumann, K.-M.; Heesen, C. Impact of aerobic training on immune-endocrine parameters, neurotrophic factors, quality of life and coordinative function in multiple sclerosis. J. Neurol. Sci. 2004, 225, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Zoladz, J.A.; Majerczak, J.; Zeligowska, E.; Mencel, J.; Jaskolski, A.; Jaskolska, A.; Marusiak, J. Moderate-intensity interval training increases serum brain-derived neurotrophic factor level and decreases inflammation in Parkinson’s disease patients. J. Physiol. Pharmacol. Off. J. Pol. Physiol. Soc. 2014, 65, 441–448. [Google Scholar]
- Enette, L.; Vogel, T.; Merle, S.; Valard-Guiguet, A.G.; Ozier-Lafontaine, N.; Neviere, R.; Leuly-Joncart, C.; Fanon, J.L.; Lang, P.O. Effect of 9 weeks continuous vs. interval aerobic training on plasma BDNF levels, aerobic fitness, cognitive capacity and quality of life among seniors with mild to moderate Alzheimer’s disease: A randomized controlled trial. Eur. Rev. Aging Phys. Act. Off. J. Eur. Group Res. Elder. Phys. Act. 2020, 17, 2. [Google Scholar] [CrossRef]
- Briken, S.; Rosenkranz, S.C.; Keminer, O.; Patra, S.; Ketels, G.; Heesen, C.; Hellweg, R.; Pless, O.; Schulz, K.-H.; Gold, S. Effects of exercise on Irisin, BDNF and IL-6 serum levels in patients with progressive multiple sclerosis. J. Neuroimmunol. 2016, 299, 53–58. [Google Scholar] [CrossRef] [PubMed]
- Araya, A.V.; Orellana, X.; Godoy, D.; Soto, L.; Fiedler, J. Effect of Exercise on Circulating Levels of Brain-derived Neurotrophic Factor (BDNF) in Overweight and Obese Subjects. Horm. Metab. Res. 2013, 45, 541–544. [Google Scholar] [CrossRef]
- Kimhy, D.; Vakhrusheva, J.; Bartels, M.N.; Armstrong, H.F.; Ballon, J.S.; Khan, S.; Chang, R.W.; Hansen, M.C.; Ayanruoh, L.; Lister, A.; et al. The Impact of Aerobic Exercise on Brain-Derived Neurotrophic Factor and Neurocognition in Individuals With Schizophrenia: A Single-Blind, Randomized Clinical Trial. Schizophr. Bull. 2015, 41, 859–868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krogh, J.; Rostrup, E.; Thomsen, C.; Elfving, B.; Videbech, P.; Nordentoft, M. The effect of exercise on hippocampal volume and neurotrophines in patients with major depression–A randomized clinical trial. J. Affect. Disord. 2014, 165, 24–30. [Google Scholar] [CrossRef]
- Maass, A.; Düzel, S.; Brigadski, T.; Goerke, M.; Becke, A.; Sobieray, U.; Neumann, K.; Lövdén, M.; Lindenberger, U.; Bäckman, L.; et al. Relationships of peripheral IGF-1, VEGF and BDNF levels to exercise-related changes in memory, hippocampal perfusion and volumes in older adults. NeuroImage 2016, 131, 142–154. [Google Scholar] [CrossRef] [Green Version]
- Matura, S.; Fleckenstein, J.; Deichmann, R.; Engeroff, T.; Füzéki, E.; Hattingen, E.; Hellweg, R.; Lienerth, B.; Pilatus, U.; Schwarz, S.; et al. Effects of aerobic exercise on brain metabolism and grey matter volume in older adults: Results of the randomised controlled SMART trial. Transl. Psychiatry 2017, 7, e1172. [Google Scholar] [CrossRef] [Green Version]
- Seifert, T.; Brassard, P.; Wissenberg, M.; Rasmussen, P.; Nordby, P.; Stallknecht, B.; Adser, H.; Jakobsen, A.H.; Pilegaard, H.; Nielsen, H.B.; et al. Endurance training enhances BDNF release from the human brain. Am. J. Physiol. Integr. Comp. Physiol. 2010, 298, R372–R377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schiffer, T.; Schulte, S.; Hollmann, W.; Bloch, W.; Strüder, H.K. Effects of Strength and Endurance Training on Brain-derived Neurotrophic Factor and Insulin-like Growth Factor 1 in Humans. Horm. Metab. Res. 2008, 41, 250–254. [Google Scholar] [CrossRef]
- Williams, J.S.; Ferris, L.T. Effects of Endurance Exercise Training on Brain- Derived Neurotrophic Factor. J. Exerc. Physiol. Online 2012, 15, 11–17. [Google Scholar]
- Baker, L.D.; Frank, L.L.; Foster-Schubert, K.; Green, P.S.; Wilkinson, C.W.; McTiernan, A.; Plymate, S.R.; Fishel, M.A.; Watson, G.S.; Cholerton, B.A.; et al. Effects of aerobic exercise on mild cognitive impairment: A controlled trial. Arch. Neurol. 2010, 67, 71–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, H.C.; Kim, J.K.; Lee, N.J.; Kim, S.Y.; Yoon, N.K. Effects of combined exercise on cardiovascular risk factors and serum BDNF level in mid-aged women. J. Exerc. Nutr. Biochem. 2014, 18, 61–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruscheweyh, R.; Willemer, C.; Krüger, K.; Duning, T.; Warnecke, T.; Sommer, J.; Völker, K.; Ho, H.; Mooren, F.; Knecht, S.; et al. Physical activity and memory functions: An interventional study. Neurobiol. Aging 2011, 32, 1304–1319. [Google Scholar] [CrossRef] [PubMed]
- Swift, D.L.; Johannsen, N.M.; Myers, V.H.; Earnest, C.P.; Smits, J.A.J.; Blair, S.N.; Church, T.S. The Effect of Exercise Training Modality on Serum Brain Derived Neurotrophic Factor Levels in Individuals with Type 2 Diabetes. PLoS ONE 2012, 7, e42785. [Google Scholar] [CrossRef]
- Erickson, K.I.; Voss, M.W.; Prakash, R.S.; Basak, C.; Szabo, A.; Chaddock, L.; Kim, J.S.; Heo, S.; Alves, H.; White, S.M.; et al. Exercise training increases size of hippocampus and improves memory. Proc. Natl. Acad. Sci. USA 2011, 108, 3017–3022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahn, J.H.; Choi, J.H.; Park, J.H.; Kim, I.H.; Cho, J.-H.; Lee, J.-C.; Koo, H.-M.; Hwangbo, G.; Yoo, K.-Y.; Lee, C.H.; et al. Long-Term Exercise Improves Memory Deficits via Restoration of Myelin and Microvessel Damage, and Enhancement of Neurogenesis in the Aged Gerbil Hippocampus After Ischemic Stroke. Neurorehabilit. Neural Repair 2016, 30, 894–905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.-F.; Chen, H.-I.; Yu, L.; Kuo, Y.-M.; Wu, F.-S.; Chuang, J.-I.; Liao, P.-C.; Jen, C.J. Upregulation of hippocampal TrkB and synaptotagmin is involved in treadmill exercise-enhanced aversive memory in mice. Neurobiol. Learn. Mem. 2008, 90, 81–89. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.-F.; Chen, H.-I.; Wu, C.-L.; Kuo, Y.-M.; Yu, L.; Huang, A.-M.; Wu, F.-S.; Chuang, J.-I.; Jen, C.J. Differential effects of treadmill running and wheel running on spatial or aversive learning and memory: Roles of amygdalar brain-derived neurotrophic factor and synaptotagmin I. J. Physiol. 2009, 587, 3221–3231. [Google Scholar] [CrossRef] [PubMed]
- Skup, M.; Dwornik, A.; Macias, M.; Sulejczak, D.; Wiater, M.; Czarkowska-Bauch, J. Long-Term Locomotor Training Up-Regulates TrkBFL Receptor-like Proteins, Brain-Derived Neurotrophic Factor, and Neurotrophin 4 with Different Topographies of Expression in Oligodendroglia and Neurons in the Spinal Cord. Exp. Neurol. 2002, 176, 289–307. [Google Scholar] [CrossRef] [PubMed]
- Fahimi, A.; Baktir, M.A.; Moghadam, S.; Mojabi, F.S.; Sumanth, K.; McNerney, M.W.; Ponnusamy, R.; Salehi, A. Physical exercise induces structural alterations in the hippocampal astrocytes: Exploring the role of BDNF-TrkB signaling. Brain Struct. Funct. 2017, 222, 1797–1808. [Google Scholar] [CrossRef]
- da Silva, S.G.; Unsain, N.; Mascó, D.H.; Toscano-Silva, M.; de Amorim, H.A.; Araujo, B.; Simões, P.S.R.; Mazzacoratti, M.D.G.N.; Mortara, R.; Scorza, F.; et al. Early exercise promotes positive hippocampal plasticity and improves spatial memory in the adult life of rats. Hippocampus 2010, 22, 347–358. [Google Scholar] [CrossRef]
- Alomari, M.A.; Khabour, O.F.; Alzoubi, K.H.; Alzubi, M.A. Forced and voluntary exercises equally improve spatial learning and memory and hippocampal BDNF levels. Behav. Brain Res. 2013, 247, 34–39. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.-J.; Ko, I.-G.; Kim, S.-E.; Hwang, L.; Lee, M.-G.; Kim, D.-Y.; Jung, S.-Y. Age-dependent differences of treadmill exercise on spatial learning ability between young- and adult-age rats. J. Exerc. Rehabil. 2017, 13, 381–386. [Google Scholar] [CrossRef] [Green Version]
- So, J.H.; Huang, C.; Ge, M.; Cai, G.; Zhang, L.; Lu, Y.; Mu, Y. Intense Exercise Promotes Adult Hippocampal Neurogenesis But Not Spatial Discrimination. Front. Cell. Neurosci. 2017, 11, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uysal, N.; Kiray, M.; Sisman, A.R.; Camsari, U.M.; Gencoglu, C.; Baykara, B.; Cetinkaya, C.; Aksu, I. Effects of voluntary and involuntary exercise on cognitive functions, and VEGF and BDNF levels in adolescent rats. Biotech. Histochem. Off. Publ. Biol. Stain. Comm. 2015, 90, 55–68. [Google Scholar] [CrossRef] [PubMed]
- Cassilhas, R.; Lee, K.; Fernandes, J.; Oliveira, M.; Tufik, S.; Meeusen, R.; de Mello, M.T. Spatial memory is improved by aerobic and resistance exercise through divergent molecular mechanisms. Neuroscience 2012, 202, 309–317. [Google Scholar] [CrossRef] [PubMed]
- Costa, M.; Ardais, A.; Fioreze, G.; Mioranzza, S.; Botton, P.; Souza, D.; Rocha, J.; Porciúncula, L. The impact of the frequency of moderate exercise on memory and brain-derived neurotrophic factor signaling in young adult and middle-aged rats. Neuroscience 2012, 222, 100–109. [Google Scholar] [CrossRef] [PubMed]
- Etemad, A.; Sheikhzadeh, F.; Ahmadiasl, N. Evaluation of brain-derived neurotrophic factor in diabetic rats. Neurol. Res. 2014, 37, 217–222. [Google Scholar] [CrossRef] [PubMed]
- Jiménez-Maldonado, A.; De Álvarez-Buylla, E.R.; Montero, S.; Melnikov, V.; Castro-Rodríguez, E.; Gamboa-Domínguez, A.; Rodríguez-Hernández, A.; Lemus, M.; Murguía, J.M. Chronic Exercise Increases Plasma Brain-Derived Neurotrophic Factor Levels, Pancreatic Islet Size, and Insulin Tolerance in a TrkB-Dependent Manner. PLoS ONE 2014, 9, e115177. [Google Scholar] [CrossRef]
- Radak, Z.; Toldy, A.; Szabo, Z.; Siamilis, S.; Nyakas, C.; Silye, G.; Jakus, J.; Goto, S. The effects of training and detraining on memory, neurotrophins and oxidative stress markers in rat brain. Neurochem. Int. 2006, 49, 387–392. [Google Scholar] [CrossRef]
- Sheikhzadeh, F.; Etemad, A.; Khoshghadam, S.; Asl, N.A.; Zare, P. Hippocampal BDNF content in response to short- and long-term exercise. Neurol. Sci. 2015, 36, 1163–1166. [Google Scholar] [CrossRef]
- Vilela, T.C.; Muller, A.P.; Damiani, A.P.; Macan, T.P.; Da Silva, S.; Canteiro, P.B.; Casagrande, A.D.S.; Pedroso, G.D.S.; Nesi, R.T.; De Andrade, V.M.; et al. Strength and Aerobic Exercises Improve Spatial Memory in Aging Rats Through Stimulating Distinct Neuroplasticity Mechanisms. Mol. Neurobiol. 2016, 54, 7928–7937. [Google Scholar] [CrossRef]
- Pietrelli, A.; Matković, L.; Vacotto, M.; Lopez-Costa, J.; Basso, N.; Brusco, A. Aerobic exercise upregulates the BDNF-Serotonin systems and improves the cognitive function in rats. Neurobiol. Learn. Mem. 2018, 155, 528–542. [Google Scholar] [CrossRef] [PubMed]
- Huang, E.J.; Reichardt, L.F. Neurotrophins: Roles in Neuronal Development and Function. Annu. Rev. Neurosci. 2001, 24, 677–736. [Google Scholar] [CrossRef] [Green Version]
- Bekinschtein, P.; Halbach, O.V.B.U. Editorial: Cellular and Molecular Mechanisms of Neurotrophin Function in the Nervous System. Front. Cell. Neurosci. 2020, 14, 101. [Google Scholar] [CrossRef] [PubMed]
- Ivanisevic, L.; Saragovi, H.U. Neurotrophins. In Handbook of Biologically Active Peptides; Kastin, A., Ed.; Academic Press: Cambridge, MA, USA, 2013; pp. 1639–1646. [Google Scholar]
- Platholi, J.; Lee, F.S. Neurotrophic Factors. In Handbook of Developmental Neurotoxicology, 2nd ed.; Slikker, W.J., Paule, M.G., Wang, C., Eds.; Academic Press: Cambridge, MA, USA, 2018; pp. 55–64. [Google Scholar]
- Sánchez-Sánchez, J.; Arévalo, J.C. A Review on Ubiquitination of Neurotrophin Receptors: Facts and Perspectives. Int. J. Mol. Sci. 2017, 18, 630. [Google Scholar] [CrossRef] [Green Version]
- Phillips, C. Brain-Derived Neurotrophic Factor, Depression, and Physical Activity: Making the Neuroplastic Connection. Neural Plast. 2017, 2017, 1–17. [Google Scholar] [CrossRef]
- Teng, H.K.; Teng, K.; Lee, R.; Wright, S.; Tevar, S.; Almeida, R.; Kermani, P.; Torkin, R.; Chen, Z.-Y.; Lee, F.S.; et al. ProBDNF Induces Neuronal Apoptosis via Activation of a Receptor Complex of p75NTR and Sortilin. J. Neurosci. 2005, 25, 5455–5463. [Google Scholar] [CrossRef] [PubMed]
- Zagrebelsky, M.; Holz, A.; DeChant, G.; Barde, Y.; Bonhoeffer, T.; Korte, M. The p75 Neurotrophin Receptor Negatively Modulates Dendrite Complexity and Spine Density in Hippocampal Neurons. J. Neurosci. 2005, 25, 9989–9999. [Google Scholar] [CrossRef] [Green Version]
- Woo, N.H.; Teng, H.K.; Siao, C.-J.; Chiaruttini, C.; Pang, P.T.; Milner, T.A.; Hempstead, B.L.; Lu, B. Activation of p75NTR by proBDNF facilitates hippocampal long-term depression. Nat. Neurosci. 2005, 8, 1069–1077. [Google Scholar] [CrossRef] [PubMed]
- Villanueva, R. Neurobiology of Major Depressive Disorder. Neural Plast. 2013, 2013, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.; Christian, K.; Lu, B. BDNF: A key regulator for protein synthesis-dependent LTP and long-term memory? Neurobiol. Learn. Mem. 2008, 89, 312–323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Messaoudi, E.; Ying, S.-W.; Kanhema, T.; Croll, S.; Bramham, C. Brain-Derived Neurotrophic Factor Triggers Transcription-Dependent, Late Phase Long-Term Potentiation In Vivo. J. Neurosci. 2002, 22, 7453–7461. [Google Scholar] [CrossRef] [Green Version]
- Lee, R.; Kermani, P.; Teng, K.K.; Hempstead, B.L. Regulation of Cell Survival by Secreted Proneurotrophins. Science 2001, 294, 1945–1948. [Google Scholar] [CrossRef] [Green Version]
- Kojima, M.; Mizui, T. BDNF Propeptide: A Novel Modulator of Synaptic Plasticity. In Vitamins and Hormones; Litwack, G., Ed.; Elsevier: Amsterdam, The Netherlands, 2017; pp. 19–28. [Google Scholar]
- Reichardt, L.F. Neurotrophin-regulated signalling pathways. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 2006, 361, 1545–1564. [Google Scholar] [CrossRef] [Green Version]
- Prakash, Y.; Thompson, M.A.; Meuchel, L.; Pabelick, C.M.; Mantilla, C.; Zaidi, S.; Martin, R.J. Neurotrophins in lung health and disease. Expert Rev. Respir. Med. 2010, 4, 395–411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bathina, S.; Das, U.N. Brain-derived neurotrophic factor and its clinical implications. Arch. Med. Sci. 2015, 6, 1164–1178. [Google Scholar] [CrossRef] [PubMed]
- Barde, Y.A.; Edgar, D.; Thoenen, H. Purification of a New Neurotrophic Factor from Mammalian Brain. EMBO J. 1982, 1, 549–553. [Google Scholar] [CrossRef] [PubMed]
- Suliman, S.; Hemmings, S.M.J.; Seedat, S. Brain-Derived Neurotrophic Factor (BDNF) protein levels in anxiety disorders: Systematic review and meta-regression analysis. Front. Integr. Neurosci. 2013, 7, 55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miranda, M.; Morici, J.F.; Zanoni, M.B.; Bekinschtein, P. Brain-Derived Neurotrophic Factor: A Key Molecule for Memory in the Healthy and the Pathological Brain. Front. Cell. Neurosci. 2019, 13, 363. [Google Scholar] [CrossRef]
- Caporali, A.; Emanueli, C. Cardiovascular Actions of Neurotrophins. Physiol. Rev. 2009, 89, 279–308. [Google Scholar] [CrossRef] [Green Version]
- Nakagawa, T.; Tsuchida, A.; Itakura, Y.; Nonomura, T.; Ono, M.; Hirota, F.; Inoue, T.; Nakayama, C.; Taiji, M.; Noguchi, H. Brain-derived neurotrophic factor regulates glucose metabolism by modulating energy balance in diabetic mice. Diabetes 2000, 49, 436–444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsuchida, A.; Nonomura, T.; Nakagawa, T.; Itakura, Y.; Ono-Kishino, M.; Yamanaka, M.; Sugaru, E.; Taiji, M.; Noguchi, H. Brain-derived neurotrophic factor ameliorates lipid metabolism in diabetic mice. Diabetes Obes. Metab. 2002, 4, 262–269. [Google Scholar] [CrossRef]
- Krabbe, K.S.; Nielsen, A.R.; Krogh-Madsen, R.; Plomgaard, P.; Rasmussen, P.; Erikstrup, C.; Fischer, C.P.; Lindegaard, B.; Petersen, A.M.; Taudorf, S.; et al. Brain-derived neurotrophic factor (BDNF) and type 2 diabetes. Diabetologia 2007, 50, 431–438. [Google Scholar] [CrossRef]
- Lessmann, V.; Brigadski, T. Mechanisms, locations, and kinetics of synaptic BDNF secretion: An update. Neurosci. Res. 2009, 65, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Kowiański, P.; Lietzau, G.; Czuba, E.; Waśkow, M.; Steliga, A.; Moryś, J. BDNF: A Key Factor with Multipotent Impact on Brain Signaling and Synaptic Plasticity. Cell. Mol. Neurobiol. 2017, 38, 579–593. [Google Scholar] [CrossRef] [PubMed]
- Foltran, R.B.; Diaz, S.L. BDNF isoforms: A round trip ticket between neurogenesis and serotonin? J. Neurochem. 2016, 138, 204–221. [Google Scholar] [CrossRef]
- Hashimoto, K. Regulation of brain-derived neurotrophic factor (BDNF) and its precursor proBDNF in the brain by serotonin. Eur. Arch. Psychiatry Clin. Neurosci. 2016, 266, 195–197. [Google Scholar] [CrossRef] [PubMed]
- Maisonpierre, P.C.; Le Beau, M.M.; Espinosa, R., 3rd; Ip, N.Y.; Belluscio, L.; de la Monte, S.M.; Squinto, S.; Furth, M.E.; Yancopoulos, G.D. Human and rat brain-derived neurotrophic factor and neurotrophin-3: Gene structures, distributions, and chromosomal localizations. Genomics 1991, 10, 558–568. [Google Scholar] [CrossRef]
- Chao, M.V.; Hempstead, B.L. p75 and Trk: A two-receptor system. Trends Neurosci. 1995, 18, 321–326. [Google Scholar] [CrossRef]
- Feter, N.; Alt, R.; Dias, M.; Rombaldi, A. How do different physical exercise parameters modulate brain-derived neurotrophic factor in healthy and non-healthy adults? A systematic review, meta-analysis and meta-regression. Sci. Sports 2019, 34, 293–304. [Google Scholar] [CrossRef]
- Szuhany, K.L.; Bugatti, M.; Otto, M. A meta-analytic review of the effects of exercise on brain-derived neurotrophic factor. J. Psychiatr. Res. 2014, 60, 56–64. [Google Scholar] [CrossRef] [Green Version]
- Walsh, J.J.; Tschakovsky, M.E. Exercise and circulating BDNF: Mechanisms of release and implications for the design of exercise interventions. Appl. Physiol. Nutr. Metab. 2018, 43, 1095–1104. [Google Scholar] [CrossRef]
- Brunelli, A.; Dimauro, I.; Sgro’, P.G.M.; Emerenziani, G.P.; Magi, F.; Baldari, C.; Guidetti, L.; DI Luigi, L.; Parisi, P.; Caporossi, D. Acute Exercise Modulates BDNF and pro-BDNF Protein Content in Immune Cells. Med. Sci. Sports Exerc. 2012, 44, 1871–1880. [Google Scholar] [CrossRef] [PubMed]
- Fujimura, H.; Chen, R.; Nakamura, T.; Nakahashi, T.; Kambayashi, J.-I.; Sun, B.; Altar, C.A.; Tandon, N.N. Brain-derived Neurotrophic Factor Is Stored in Human Platelets and Released by Agonist Stimulation. Thromb. Haemost. 2002, 87, 728–734. [Google Scholar] [CrossRef] [Green Version]
- Lommatzsch, M.; Braun, A.; Mannsfeldt, A.; Botchkarev, V.; Botchkareva, N.V.; Paus, R.; Fischer, A.; Lewin, G.R.; Renz, H. Abundant Production of Brain-Derived Neurotrophic Factor by Adult Visceral Epithelia: Implications for Paracrine and Target-Derived Neurotrophic Functions. Am. J. Pathol. 1999, 155, 1183–1193. [Google Scholar] [CrossRef]
- Marosi, K.; Mattson, M.P. BDNF mediates adaptive brain and body responses to energetic challenges. Trends Endocrinol. Metab. 2013, 25, 89–98. [Google Scholar] [CrossRef] [Green Version]
- Matthews, V.B.; Astrom, M.B.; Chan, M.H.; Bruce, C.R.; Krabbe, K.S.; Prelovsek, O.; Akerstrom, T.; Yfanti, C.; Broholm, C.; Mortensen, O.H.; et al. Brain-derived neurotrophic factor is produced by skeletal muscle cells in response to contraction and enhances fat oxidation via activation of AMP-activated protein kinase. Diabetologia 2009, 52, 1409–1418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakagomi, A.; Okada, S.; Yokoyama, M.; Yoshida, Y.; Shimizu, I.; Miki, T.; Kobayashi, Y.; Minamino, T. Role of the central nervous system and adipose tissue BDNF/TrkB axes in metabolic regulation. npj Aging Mech. Dis. 2015, 1, 15009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prigent-Tessier, A.; Quirié, A.; Maguin-Gaté, K.; Szostak, J.; Mossiat, C.; Nappey, M.; Devaux, S.; Marie, C.; Demougeot, C. Physical training and hypertension have opposite effects on endothelial brain-derived neurotrophic factor expression. Cardiovasc. Res. 2013, 100, 374–382. [Google Scholar] [CrossRef] [Green Version]
- Quirie, A.; Hervieu, M.; Garnier, P.; Demougeot, C.; Mossiat, C.; Bertrand, N.; Martin, A.; Marie, C.; Prigent-Tessier, A. Comparative Effect of Treadmill Exercise on Mature BDNF Production in Control versus Stroke Rats. PLoS ONE 2012, 7, e44218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walsh, J.J.; Edgett, B.; Tschakovsky, M.E.; Gurd, B.J. Fasting and exercise differentially regulate BDNF mRNA expression in human skeletal muscle. Appl. Physiol. Nutr. Metab. 2015, 40, 96–98. [Google Scholar] [CrossRef]
- Naegelin, Y.; Dingsdale, H.; Säuberli, K.; Schädelin, S.; Kappos, L.; Barde, Y.-A. Measuring and Validating the Levels of Brain-Derived Neurotrophic Factor in Human Serum. Eneuro 2018, 5. [Google Scholar] [CrossRef] [Green Version]
- Fernandez, P.J.C.; Säuberli, K.; Colzani, M.; Moreau, T.; Ghevaert, C.; Barde, Y.-A. Brain-derived Neurotrophic Factor in Megakaryocytes. J. Biol. Chem. 2016, 291, 9872–9881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.; Zettler, C.; Cupler, E.J.; Hurtado, P.; Wong, K.; Rush, R.A. Neurotrophin 4/5 immunoassay: Identification of sources of errors for the quantification of neurotrophins. J. Neurosci. Methods 2000, 99, 119–127. [Google Scholar] [CrossRef]
- László, A.; Lénárt, L.; Illésy, L.; Fekete, A.; Nemcsik, J. The role of neurotrophins in psychopathology and cardiovascular diseases: Psychosomatic connections. J. Neural. Transm. 2019, 126, 265–278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fitzgerald, K.A.; O’Neill, L.A.; Gearing, A.J.; Callard, R.E. NT-4. In The Cytokine FactsBook and Webfacts, 2nd ed.; Academic Press: Cambridge, MA, USA, 2001; pp. 412–414. [Google Scholar]
- Ibanez, C.F. Neurotrophin-4: The odd one out in the neurotrophin family. Neurochem. Res. 1996, 21, 787–793. [Google Scholar] [CrossRef] [PubMed]
- Timmusk, T.; Belluardo, N.; Metsis, M.; Persson, H. Widespread and developmentally regulated expression of neurotrophin-4 mRNA in rat brain and peripheral tissues. Eur. J. Neurosci. 1993, 5, 605–613. [Google Scholar] [CrossRef]
- Ip, N.Y.; Ibanez, C.F.; Nye, S.H.; McClain, J.; Jones, P.F.; Gies, D.R.; Belluscio, L.; Le Beau, M.M.; Espinosa, R., 3rd; Squinto, S.P.; et al. Mammalian neurotrophin-4: Structure, chromosomal localization, tissue distribution, and receptor specificity. Proc. Natl. Acad. Sci. USA 1992, 89, 3060–3064. [Google Scholar] [CrossRef] [Green Version]
- Dąbkowska, M.; Adamczak, M.; Barbasz, J.; Cieśla, M.; Machalinski, B. Adsorption/Desorption Transition of Recombinant Human Neurotrophin 4: Physicochemical Characterization. Langmuir 2017, 33, 9548–9557. [Google Scholar] [CrossRef]
- Chao, M.V. Neurotrophins and their receptors: A convergence point for many signalling pathways. Nat. Rev. Neurosci. 2003, 4, 299–309. [Google Scholar] [CrossRef]
- Sarnat, H.B. Development of Olfaction and Taste in the Human Fetus and Neonate. In Fetal and Neonatal Physiology, 5th ed.; Polin, A.R., Abman, S.H., Rowitch, D.H., Benitz, W.E., Fox, W.W., Eds.; Elsevier: Amsterdam, The Netherlands, 2017; pp. 1411–1420. [Google Scholar]
- Berkemeier, L.R.; Winslow, J.W.; Kaplan, D.R.; Nikolics, K.; Goeddel, D.V.; Rosenthal, A. Neurotrophin-5: A novel neurotrophic factor that activates trk and trkB. Neuron 1991, 7, 857–866. [Google Scholar] [CrossRef]
- Sorour, N.E.; Elesawy, F.M.; Tabl, H.A.; Ibrahim, M.E.; Akl, E.M. Evaluation of serum levels of neurotrophin 4 and brain-derived nerve growth factor in uremic pruritus patients. Clin. Cosmet. Investig. Dermatol. 2019, 12, 109–114. [Google Scholar] [CrossRef] [Green Version]
- Mrówczyński, W. Health Benefits of Endurance Training: Implications of the Brain-Derived Neurotrophic Factor-A Systematic Review. Neural Plast. 2019, 2019, 5413067. [Google Scholar] [CrossRef] [PubMed]
- Warburton, D.E.R.; Bredin, S.S.D. Health benefits of physical activity: A systematic review of current systematic reviews. Curr. Opin. Cardiol. 2017, 32, 541–556. [Google Scholar] [CrossRef] [PubMed]
- Coelho, F.G.D.M.; Gobbi, S.; Andreatto, C.A.D.A.; Corazza, D.I.; Pedroso, R.V.; Santos-Galduróz, R.F. Physical exercise modulates peripheral levels of brain-derived neurotrophic factor (BDNF): A systematic review of experimental studies in the elderly. Arch. Gerontol. Geriatr. 2013, 56, 10–15. [Google Scholar] [CrossRef] [PubMed]
- Barré-Sinoussi, F.; Montagutelli, X. Animal models are essential to biological research: Issues and perspectives. Futur. Sci. OA 2015, 1, FSO63. [Google Scholar] [CrossRef] [Green Version]
- Kapp, M.B. Ethical and legal issues in research involving human subjects: Do you want a piece of me? J. Clin. Pathol. 2006, 59, 335–339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, W.; Kastin, A.J. Penetration of neurotrophins and cytokines across the blood–brain/blood–spinal cord barrier. Adv. Drug Deliv. Rev. 1999, 36, 291–298. [Google Scholar] [CrossRef]
- Rasmussen, P.; Brassard, P.; Adser, H.; Pedersen, M.V.; Leick, L.; Hart, E.; Secher, N.H.; Pedersen, B.K.; Pilegaard, H. Evidence for a release of brain-derived neurotrophic factor from the brain during exercise. Exp. Physiol. 2009, 94, 1062–1069. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Author(s) | Population Studied (Exercise Group n vs. Control Group n ) | n | BDNF Measure (Analysis Technique) | Time of Measure | Duration of Exercise | Aerobic Exercise (Days p/Week of Training) | Intensity of Exercise for Exercise Group (Physiological Parameter) | Main Outcomes |
|---|---|---|---|---|---|---|---|---|
| <12 weeks | ||||||||
| Griffin, et al. (2011) [25] | Young Sedentary Healthy Adult Males (3-week Aerobic Exercise Group-C-Ex3 n = 9 vs. 5-week Aerobic Exercise Group C-Ex5 n = 9 vs. Sedentary Non-Exercise Control Group-CON n = 15) | 33 | Serum (ELISA) | pre-exercise and post-exercise at weeks 3/5 | 3 weeks & 5 weeks | stationary cycle ergometer (3 days/week) | 60% VO2max | 3 weeks of cycling training presented no effect on serum BDNF concentration; however, an increase of serum BDNF concentration was observed at week 5. |
| Salehi, et al. (2016) [26] | Major Depressive Disorder Patients (Aerobic Exercise Group n = 20) | 20 | Plasma (ELISA) | pre- and post intervention | 4 weeks | cycling on a treadmill (3 days/week) | 60–75% of VO2max | Pre- to post-assessment plasma BDNF levels were increased in the Aerobic Exercise Training group. |
| Zoladz, et al. (2008) [27] | Healthy Physically Active Young Men (Exercise Group n = 13) | 13 | Plasma (ELISA) | pre- and post intervention | 5 weeks | cycle ergometer (4 days/week) | 90% of VO2 at lactacte threshold | A 5-week exercise program of moderate intensity endurance training resulted in a significant “chronic” increase in the basal, as well as in the exercise induced “acute” increase in plasma BDNF levels. |
| Kerling, et al. (2017) [28] | Depressed Inpatients (Aerobic Exercise Group + Treatment as Usual n = 22 vs. Treament as Usual Control Group n = 20) | 42 | Serum (Quantikine Sandwich Enzyme Immunoassay) | pre- and post intervention | 6 weeks | bicycle ergometer, treadmill, cross-trainer or rowing (n.d.) | n.d. | An increase in serum BDNF in the exercise group was detected. Alterations over time reached no statistical significance in both groups. The Treatment as Usual Control Group serum BDNF levels decreased slightly. Exercise appears to have additional effects on BDNF serum concentrations in people with major depressive disorder. |
| Babaei, et al. (2013) [29] | Metabolic Syndrome (MetS) and Healthy Middle-Aged Males (MetS-Exercise n = 11 vs. MetS-Control n = 10 vs. Healthy-Exercise n = 11 vs. Healthy-Control n = 10) | 40 | Serum (ELISA) | pre- and post intervention | 6 weeks | walking &/or running (3 days/week) | between 50–60% of VO2peak | After aerobic exercise, BDNF levels significantly increased in the Healthy Exercise Group, but decreased in the Metabolic Syndrome Exercise Group. |
| Damirchi, et al. (2014) [30] | Middle-Aged Men Diagnosed With Metabolic Syndrome (Aerobic Training Exercise Group n = 11 vs. Non-exercise Control Group n = 10) | 21 | Serum (ELISA) | pre- and post intervention | 6 weeks | walking or running (3 days/week) | 50–60% of VO2 peak | Serum BDNF significantly decreased after chronic aerobic training in the Metabolic Syndrome exercise group subjects (Blood samples were collected at the end of the 6-week training). |
| Wagner, et al. (2017) [31] | Male Students (Aerobic Exercise Group n = 17 vs. Control Group n = 17) | 34 | Serum (ELISA) | pre- and post intervention | 6 weeks | bicycle ergometer (3 days/week) | 77± 9% of pre-training VO2max (ranging from 60% to 88%) | The results indicate a decrease in the exercise-induced BDNF concentration after the intervention in the exercise group and an increase in the control group. Unchanged baseline BDNF serum concentrations and decreased exercise-induced BDNF levels were reported in the Exercise Group. |
| Castellano & White, (2008) [32] | Multiple Sclerosis (MS) (MS Exercise Group n = 11 vs. Healthy Exercise Control Group n = 11) | 22 | Serum (Quantikine Sandwich Enzyme Immunoassay) | pre- and post intervention | 8 weeks | cycle ergometer (3 days/week) | 60% VO2peak | Resting serum BDNF was significantly lower in MS compared to control subjects at week 0, but was not significantly different at week 8. In the MS exercise group, an elevated concentration of serum BDNF was observed following 4 weeks of training with a return to baseline at week 8. Non-significant difference in the MS group at 8 weeks. In contrast, resting BDNF concentrations remained unchanged at 4 and 8 weeks of training in controls. |
| Cho & Roh (2016) [33] | Young Obese Men (Aerobic Exercise Group n = 8 vs. Non-Exercise Control Group n = 8) | 16 | Serum (ELISA) | pre- and post intervention | 8 weeks | treadmill running (3 days/week) | 70% of the heart rate reserve (HRR) | Following the intervention (regular aerobic exercise training), serum BDNF levels were significantly higher than those prior to the intervention in the exercise group of obese individuals. |
| El-Tamawy, et al. (2014) [34] | Ischemic Stroke Patients (G2 subjected to a physiotherapy program followed by aerobic exercise n = 15 vs. G1-control group-subject to a physiotherapy program without aerobic exercise n = 15) | 30 | Serum (ELISA) | pre- and post intervention | 8 weeks | bicycle ergometer (3 days/week) | n.d. | Aerobic exercise, following an acute ischemic stroke, is accompanied by an increase in the serum levels of BDNF. At the end of the treatment, compared with the pre-concentration levels, there was a significant increase in the serum BDNF levels within the exercise group, while the control group showed no significant increase in the serum BDNF levels. |
| Goekint, et al. (2010) [35] | Young Sedentary Students (Exercise Group n = 9 vs. Physically Inactive Control Group n = 7) | 16 | Serum (ELISA) | pre- and post intervention | 8 weeks | walking, running, cycling, synchro, wave (3 days/week) | n.d. | Peripheral serum BDNF concentration levels were not influenced by an 8 week aerobic training protocol. |
| Marusiak, et al. (2015) [36] | Mild-to-Moderate Parkinson Disease Patients (Parkinson Disease Patients Exercise Group n = 11 vs. Healthy Non-Trained Control Group n = 11) | 22 | Serum (Assay/Microplate Reader) | pre- and post intervention | 8 weeks | stationary cycloergometer (3 days/week) | 62–68% of HRmax | The BDNF serum levels increased in the Parkinson Disease Patients from the exercise group, whereas no change was observed in the Healthy Control Group. Training resulted in an increase in BDNF levels relative to the pre-training values. |
| Roh & So (2016) [37] | Obese and Non-Obese Men (Obese Exercise Group n = 10 vs. Healthy Non-Obese Exercise Group n = 10) | 20 | Serum (ELISA) | pre- and post intervention | 8 weeks | treadmill running (3 days/week) | 70% heart rate reserve | The obese group showed a significantly lower BDNF level at baseline compared with the non-obese group. The non-obese group showed no significant difference in blood neurotrophic factor levels before and after training, whereas the obese group showed a significantly higher BDNF level after training. |
| Schulz, et al. (2004) [38] | Multiple Sclerosis (MS) (Exercise Group n = 15 vs. Control-no intervention-Group n = 13) | 28 | Serum (ELISA) | pre- and post intervention | 8 weeks | bicycle ergometer (2 days/week) | 60% VO2max | Although not with statistical significance, an increase of BDNF levels was seen in the MS training group, while the levels in the MS control group decreased. |
| Zoladz, et al. (2014) [39] | Patients with Idiopatic Parkinson’s Disease (Exercise Group n = 12) | 12 | Serum (ELISA) | pre- and post intervention | 8 weeks | stationary cycle ergometer (3 days/week) | 60–75% of HRmax | Serum BDNF levels increased significantly after an 8-week moderate-intensity interval training intervention in the Parkinson Disease Exercise Patients. |
| Enette, et al. (2020) [40] | Seniors with Mild to Moderate Alzheimer’s Disease (Continuous Aerobic Exercise Group-CAT n = 14 vs. vs. Control Group n = 21) | 35 | Plasma (ELISA) | pre- and post intervention at week 10 | 9 weeks | cycle ergometer (2 days/week) | 70% of HRmax | 9 weeks of continuous aerobic training failed to induce significant plasma BDNF response compared to baseline levels. No significant change was measured in terms of plasma BDNF levels after the training program. |
| Briken, et al. (2016) [41] | Patients with Primary or Secondary Progressive Multiple Sclerosis (Exercise Group-arm ergometry/rowing or bicycle ergometry n = 28 vs. Control Group n = 9) | 37 | Serum (ELISA) | pre- and post intervention | 9 weeks | arm ergometry, rowing or bicycle ergometer (2–3 days/week) | n.d. | BDNF serum levels did not increase significantly after a training intervention of 22 sessions in comparison to the control group. Long-term effects of BDNF are less pronounced when compared with acute exercise. |
| Araya, et al. (2013) [42] | Overweight and Obese subjects (n = 15) | 15 | Serum, Plasma & Platelet (ELISA) | pre- and post intervention | 10 weeks | treadmill or bike + stretching (3 days/week) | ≥65% VO2max | In sedentary, nondepressed, overweight and obese subjects, serum and platelet BDNF circulating levels increased after 30 sessions of aerobic exercise. |
| 12–36 weeks | ||||||||
| Kimhy, et al. (2015) [43] | Schizophrenia or Related Disorder Individuals (Aerobic Exercise Group + Treatment As Usual n = 16 vs. Control Treatment As Usual Group n = 17) | 33 | Serum (ELISA) | pre- and post intervention | 12 weeks | treadmill, stationary bike or eliptical machine (3 days/week) | 60–75% of HRmax | At follow-up, BDNF increased in the Aerobic Exercise group. Group difference changes in BDNF did not reach significance, potentially due to the small sample size. |
| Krogh, et al. (2014) [44] | Major Depression Patients (Aerobic Exercise Group n = 41 vs. Control Group n = 38) | 79 | Serum (ELISA) | pre- and post intervention | 12 weeks | stationary bikes (3 days/week) | 80% of HRmax | A 3-month aerobic exercise training program did not increase BDNF serum levels. No differences were found in serum BDNF between the aerobic exercise group and the control group in the post-intervention samples. |
| Maass, et al. (2016) [45] | Healthy Sedentary Older Adults (Aerobic Exercise Group n = 21 vs. Control Group n = 19) | 40 | Plasma & Serum (ELISA) | pre- and post intervention | 12 weeks | stationary treadmill (3 days/week) | 65% of target heart rate +5% in steps for 4 weeks | BDNF did not significantly change between pre- and post exercise levels. The 3-month intervention did not alter BDNF levels in the blood. |
| Matura, et al. (2017) [46] | Healthy Older Adults (Aerobic Exercise Program Group n = 29 vs. Non-Exercise Control Group n = 24) | 53 | Serum (ELISA) | pre- and post intervention | 12 weeks | cycle ergometer (3 days/week) | 64 ± 9% VO2 max | No effect of aerobic exercise training was seen on serum BDNF concentrations compared with the control group. |
| Seifart, et al. (2010) [47] | Healthy Sedentary Males (Endurance Training n = 7 vs. Sedentary Controls n = 5) | 12 | Plasma (ELISA) | pre- and post intervention | 12 weeks | cycling, running, swimming or rowing (7 days/week) | 70% of maximal HR, equivalent to approximately 65% VO2max | 3 months of endurance training enhanced the resting release of BDNF, with no significant change in the control group. There was no training-induced increase in the release of BDNF during exercise. |
| Schiffer, et al. (2009) [48] | Healthy Sports Students (Moderate Endurance Training Group vs. Control Group) | 27 | Plasma (ELISA) | pre- and post intervention | 12 weeks | treadmill (3 days/week) | 80% of the HR at the aerobic-anaerobic threshold | There were no significant changes for BDNF. The authors suggest that exercise per se does not result in changes in plasma concentrations of BDNF. |
| Williams & Ferris, (2012) [49] | Physically Active & Healthy Young Subjects (n = 18) | 18 | Serum (ELISA) | pre- and post intervention | 12 weeks | jogging (3 days/week) | between 65–70% of HRmax | BDNF was unchanged in response to the moderate intensity training program. A 12 week aerobic training program of moderate intensity, completed by healthy subjects, did not result in an increase in BDNF concentrations. |
| Baker, et al. (2010) [50] | Adults with Amnestic Mild Cognitive Impairment (High-Intensity Aerobic Exercise Group n = 19 vs. Stretching Control Group n = 10) | 29 | Plasma (ELISA) | pre- and post intervention | 24 weeks | treadmill, stationary bicycle, or elliptical trainer (4 days/week) | 75% to 85% of HR reserve | Relative to controls, aerobic exercise increased circulating levels of BDNF in men patients and decreased levels of BDNF in women. |
| Cho, et al. (2014) [51] | Healthy Middle-Aged Women (Aerobic-Exercise Group n = 15 vs. vs. Non-exercise Control Group n = 7) | 22 | Serum (ELISA) | pre- and post intervention | 24 weeks | treadmill (4 days/week) | between 50–80% of VO2max | After 24 weeks, there were significant serum BDNF level changes in the aerobic exercise group, when compared to the control group. |
| Ruscheweyh, et al. (2011) [52] | Older Healthy Individuals (Medium-Intensity Aerobic Exercise Nordic Walking Group n = 20 vs. Exercise Gymnastics Low-Intensity Aerobic Group n = 21 vs. No Intervention Control Group n = 21) | 62 | Serum (ELISA) | pre- and post intervention | 24 weeks | nordic walking & gymnastics (3 days/week) | medium-intensity aerobic exercise: (50–60% of maximal exertion) and low-intensity aerobic exercise (30–40% of maximal exertion) | Change in physical activity levels with the intervention showed a trend for a positive association with the change in BDNF levels. The authors detected a trend of an increase in BDNF with increasing physical activity over six months. |
| >36 weeks | ||||||||
| Swift, et al. (2012) [53] | Type 2 Diabetes Individuals (Aerobic Training Only n = 40 vs. Resistance Training Only n = 44 vs. Combination of Resistance and Aerobic n = 43 vs. Non-exercise Control Group n = 23) | 150 | Serum (Chemkine Sandwich Enzyme Immunoassay) | pre- and post intervention | 36 weeks | treadmill (150 min/week) | between 50–80% of VO2max | Serum BDNF measures were not altered by 9 months of aerobic, resistance, or combination (aerobic + resistance) training when compared with the non-exercise control group. |
| Erickson, et al. (2011) [54] | Older Adults (Moderate-Intensity Aerobic Exercise Group n = 60 vs. Stretching Control Group n = 60) | 120 | Serum (Quantikine Sandwich Enzyme Immunoassay) | pre- and post intervention | 48 weeks | treadmill (3 days/week) | moderate-intensity 50–60% of HR reserve (1–7 weeks) 60–75% HR reserve (remainder of the program) | The aerobic exercise group did not demonstrate greater changes in circulating serum BDNF levels compared with the control group. However, the aerobic exercise group revealed higher post-intervention BDNF levels, when compared with the pre-intervention BDNF concentrations. |
| Author(s) | Population Studied (Exercise Group n vs. Control Group n) | n | BDNF Measure (Analysis Technique) | Time of Measure | Duration of Exercise | Aerobic Exercise (Days p/Week of Training) | Intensity of Exercise for Exercise Group (Physiological Parameter) | Main Outcomes |
|---|---|---|---|---|---|---|---|---|
| <12 weeks | ||||||||
| Ahn, et al. (2016) [55] | Aged Mongolian Gerbil Males (Sham-Operated Group n = 14 Ischemia-Operated 1-week Exercise Group n = 14 vs. Ischemia-Operated 4-week Exercise Group n = 14 vs. 4-week Sedentary Ischemia-Operated Control Group n = 14) | 56 | western blot | post-intervention | 1 and 4 weeks | treadmill running (5 days/week) | n.d. | In the Ischemia Operated 4-week Sedentary Group, BDNF protein levels were increased compared with those in the Sham-Operated Group; BDNF protein levels in the Ischemia-Operated 4-week Exercise Group were significantly increased compared with the Ischemia Operated 4-week Sedentary Group, and Ischemia-Operated 1-week Exercise Group. 4 weeks of exercise (long-term exercise) enhanced the BDNF expression. |
| Liu, et al. (2008) [56] | Male BALB/c Mice (Exercise Group n = n.d. vs. Control Group n = n.d.) | n.d. | ELISA | post-intervention | 4 weeks | treadmill exercise (5 days/week) | n.d. | The exercise effect of training on hippocampal BDNF level was acute rather than chronic; training transiently increased the hippocampal BDNF level at 1, 2, and 4 h post-exercise when compared to the control. |
| Liu, et al. (2009) [57] | Male BALB/c Mice (Forced Treadmill Exercise Group n = n.d. vs. Control Group n = n.d.) | n.d. | ELISA | post-intervention | 4 weeks | treadmill exercise (5 days/week) | n.d. | Training elevated BDNF levels. Forced treadmill running transiently elevated hippocampal BDNF levels and significantly increased BDNF levels in the amygdala compared to the control. |
| Maejima, et al. (2018) [18] | Senescence-Accelerated Mice (Control Group n = 6 vs. Resistant Exercise Group n = 6 vs. Prone Sedentary Control Group n = 8 vs. Prone Exercise Group n= 7) | 27 | ELISA | post-intervention | 4 weeks | treadmill running (5 days/week) | n.d. | The study revealed that aerobic exercise for 4 weeks incresed transcriptional and protein expression levels of BDNF in mice. Long-term exercise enhances the expression of BDNF in the hippocampus. |
| Skup, et al. (2002) [58] | Adult Male Wistar Rats (Exercise Group n = 6 vs. Sedentary Sedentary Control Group n = 6) | 12 | western blot | post-intervention | 4 weeks | treadmill exercise (5 days/week) | n.d. | Training enhanced expression of BDNF in the spinal gray matter fibers and, to a lesser extent, caused an increase of BDNF within neuronal perikarya. Immunocytochemical results show that long-lasting, moderate locomotor training, enhanced BDNF expression. |
| Fahimi, et al. (2017) [59] | Adult Male C57BL/6 Mice (Exercise Group n = 6 vs. Non-Exercise Control Group n = 6) | 12 | ELISA | post-intervention | 5 weeks | treadmill running (5–7 days/week) | n.d. | BDNF significantly increased in mRNA levels following long-term exercise. A significant increase in BDNF levels in the exercise group was observed compared with the non-exercise control group. |
| Seifart, et al. (2010) [47] | Mice (Exercise Training Group n = 8/Control Group n = 8) | 16 | fluorescence-based real-time PCR | post-intervention | 5 weeks | treadmill running (5 days/week) | n.d. | The BDNF mRNA levels in the hippocampus were higher in the trained mice than in the untrained mice from the control group. In the cerebral cortex, the BDNF mRNA levels were not significantly elevated by training but were comparable to those in the hippocampus of the untrained mice. |
| da Silva, et al. (2012) [60] | Male Wistar Rats (Exercise Group n = 27 vs. Control Group n = 27) | 54 | ELISA | post-intervention | ±5–6 weeks | treadmill (39 sessions in total) | n.d. | A significant increase in hippocampal BDNF expression was noted in the exercise group (aerobic exercise program) when compared with the control group. |
| Alomari, et al. (2013) [61] | Male Wistar Rats (Forced Exercise Group n = 14–15 vs. Control Group n = 14–15) | 28–30 | ELISA | post-intervention | 6 weeks | swimming (5 days/week) | n.d. | BDNF levels in the hippocampus were greater in the forced exercise training group when compared to the control group. BDNF was significantly higher in the forced exercise than in the control group. |
| Jin, et al. (2017) [62] | Adult Male Sprague-Dawley Rats (Young-Age Exercise Group n = 10 vs. Young-Age Control Group n = 10 vs. Adult-Age Exercise Group n = 10 vs. Adult-Age Control Group n = 10) | 40 | western blot | post-intervention | 6 weeks | treadmill running (7 days/week) | n.d. | Treadmill exercise increased BDNF expression in both young and adult-aged rats compared with the young and adult-aged control groups. |
| So, et al. (2017) [63] | C57BL/6 Female Mice (Moderate Exercise Group vs. Fatiguing Exercise Group vs. Sedentary Control Group) | n.d. | ELISA | post-intervention | 6 weeks | treadmill running (7 days/week) | n.d. | The moderate exercise group had significantly higher hippocampal BDNF expression levels when compared with the fatiguing exercise group and with the sedentary control group. No significant difference was found between the fatiguing and control groups. |
| Uysal, et al. (2015) [64] | Wistar Albino Rats (Involuntary Exercise Group n = 14 vs. Control Group n = 14) | 28 | ELISA | post-intervention | 6 weeks | treadmill running (5 days/week) | n.d. | Prefrontal Cortex BDNF levels were greater in the males of the forced exercise group than in the control group. The BDNF levels were greater in females of the forced exercise group than in controls. Hippocampal BDNF levels were increased in the forced involuntary exercise group compared with the sedentary control group. |
| Cassilhas, et al. (2012) [65] | Adult Male Wistar Rats (Aerobic Exercise Group n = 11 vs. Sham Group n = 11 vs. Control Group n = 11) | 33 | ELISA | post-intervention | 8 weeks | treadmill running (5 days/week) | n.d. | A significantly higher level of hippocampal BDNF was observed in the Aerobic Exercise Group compared with the Control Group. Aerobic Exercise modulates BDNF. |
| Costa, et al. (2012) [66] | Young Adult and Middle-Aged Male Wistar Rats (Young Adult Treadmill Running 1 days/week vs. Young Adult Treadmill Running 3 days/week vs. Young-Adult Treadmill Running 7 days/week) vs. Middle-Aged Treadmill Running 1 days/week vs. Middle-Aged Treadmill Running 3 days/week vs. Middle-Aged Treadmill Running 7 days/week vs. Young Adult Control Group vs. Middle-Aged Control Group) | n.d. | western blot | post-intervention | 8 weeks | treadmill running (1/3/5 days/week) | 60–75% VO2max | While a 1 day/week exercise program caused an increase in both proBDNF and BDNF in young rats, the other frequencies of exercise increased only BDNF levels when compared with the control group. The middle-aged exercise groups presented an increase in BDNF levels when compared with the control group. |
| Etemad, et al. (2014) [67] | Male Wistar Rats (2-Week-ShortTerm-Diabetic Exercise-Group n = 10 vs. 8-Week-LongTerm-Exercise-Diabetic Group n = 10 vs. Sedentary Diabetic Group n = 10 vs. Control Group n = 10) | 40 | ELISA | post-intervention | 2 and 8 weeks | treadmill running (7 days/week) | n.d. | No significant change on BDNF levels was observed following a short and long-term aerobic exercise program compared with control group. There was also no significant difference between groups with 2 and 8 weeks of treadmill running in BDNF levels. An exercise protocol did not change significantly BDNF levels in either group. |
| Jiménez-Maldonado, et al. (2015) [68] | Male Wistar Rats (Moderate-Intensity Exercise Group n = 6 vs. High-Intensity Exercise Group n = 4 vs. Moderate-Intensity Exercise Group + TrkB Inhibitor Injections n = 4 vs. High-Intensity Exercise Group + TrkB Inhibitor Injections n = 5 vs. Sedentary Control Group n = 6) | 25 | ELISA | post-intervention | 8 weeks | treadmill running (3 days/week) | moderate- or high-intensity (60 or 80% of VO2 max) | Chronic treadmill exercise significantly increased plasma BDNF. After training, plasma BDNF concentrations were significantly higher in the trained groups than those in the sedentary control. These results indicate that the circulating BDNF concentrations increased with chronic physical exercise. |
| Radak, et al. (2006) [69] | Wistar Rats (Exercise Trained n = 7 vs. Control n = 7) | 14 | ELISA | post-intervention at week 8 | 8 weeks | swimming training (5 days/week) | n.d. | Regular training significantly increases the production of BDNF. Exercise training significantly increased the protein content of BDNF, compared to control animal results after an 8-week aerobic exercise protocol. |
| Sheikhzaden, et al. (2015) [70] | Male Wistar Rats (2-week Exercise Group n = 10 vs. 8-week Exercise Group n = 10 vs. Control Group n = 10) | 30 | ELISA | post-intervention | 2 and 8 weeks | treadmill running (7 days/week) | n.d. | Exercise did not change BDNF content in the hippocampus of trained animals. Neither long-term (8 weeks) nor short-term (2 weeks) exercise made any significant change to BDNF levels in the hippocampus. |
| Vilela, et al. (2017) [71] | Aging Wistar Rats (Aerobic Training Group vs. Untrained Control Group) | 18 | western blot | post-intervention | 8 weeks | treadmill running (3–4 days/week) | n.d. | BDNF levels were increased after training for the aerobic exercise group compared to the rats in the untrained control group. The results show BDNF levels increased after aerobic training |
| >12 weeks | ||||||||
| Pietrelli, et al. (2018) [72] | Weaning Male Wistar Rats (Aerobically Exercised (AE) Group n = 30 vs. Sedentary Control Group n = 30) | 60 | ELISA | at 32-weeks and post-intervention | 64 weeks | treadmill running (3 days/week) | low-moderate intensity: 60–70% VO2max | AE increased the BDNF concentration in most of the brain regions studied, nonetheless this response was modulated by age (significant differences due to exercise in the striatum were only seen among the old rats). |
| Author(s) | Population Studied (Exercise Group n vs. Control Group n) | n | NT-4/5 Measure (Analysis Technique) | Time of Measure | Duration of Exercise | Aerobic Exercise (Days p/Week of Training) | Intensity of Exercise for Exercise Group (Physiological Parameter) | Main Outcomes |
|---|---|---|---|---|---|---|---|---|
| Maejima, et al. (2018) [18] | Senescence-Accelerated Mice (Resistant Sedentary Control Group n = 6 vs. Resistant Exercise Group n = 6 vs. Prone Sedentary Control Group n = 8 vs. Prone Exercise Group n= 7) | 27 | quantitative pcr based on real-time pcr | post-intervention | 4 weeks | treadmill (5 days/week) | n.d. | Although exercise upregulated transcriptional NT-4 only in SAMP, NT-4 protein did not increase in SAMP or SAMR1. Two-way ANOVA for NT-4 expression showed no significant factorial effect. |
| Skup, et al. (2002) [58] | Adult Male Wistar Rats (Exercise Group n = 6 vs. Sedentary Control Group n = 6) | 12 | western blot | post-intervention | 4 weeks | treadmill (5 days/week) | n.d. | Immunocytochemical results show that long-lasting, moderate training, enhanced NT-4/5 expression. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ribeiro, D.; Petrigna, L.; Pereira, F.C.; Muscella, A.; Bianco, A.; Tavares, P. The Impact of Physical Exercise on the Circulating Levels of BDNF and NT 4/5: A Review. Int. J. Mol. Sci. 2021, 22, 8814. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22168814
Ribeiro D, Petrigna L, Pereira FC, Muscella A, Bianco A, Tavares P. The Impact of Physical Exercise on the Circulating Levels of BDNF and NT 4/5: A Review. International Journal of Molecular Sciences. 2021; 22(16):8814. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22168814
Chicago/Turabian StyleRibeiro, Daniel, Luca Petrigna, Frederico C. Pereira, Antonella Muscella, Antonino Bianco, and Paula Tavares. 2021. "The Impact of Physical Exercise on the Circulating Levels of BDNF and NT 4/5: A Review" International Journal of Molecular Sciences 22, no. 16: 8814. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22168814