
The CB2 Receptor as a Novel Therapeutic Target for Epilepsy Treatment


1
Brain Function and Disease Laboratory, Shantou University Medical College, Xin-Ling Road #22, Shantou 515041, China
2
Medical Education Assessment and Research Center, Shantou University Medical College, Xin-Ling Road #22, Shantou 515041, China
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2021, 22(16), 8961; https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22168961
Submission received: 5 July 2021
/
Revised: 2 August 2021
/
Accepted: 10 August 2021
/
Published: 20 August 2021
(This article belongs to the Special Issue Endocannabinoid System in Health and Disease: Current Situation and Future Perspectives 3.0)
Abstract
:Epilepsy is characterized by repeated spontaneous bursts of neuronal hyperactivity and high synchronization in the central nervous system. It seriously affects the quality of life of epileptic patients, and nearly 30% of individuals are refractory to treatment of antiseizure drugs. Therefore, there is an urgent need to develop new drugs to manage and control refractory epilepsy. Cannabinoid ligands, including selective cannabinoid receptor subtype (CB1 or CB2 receptor) ligands and non-selective cannabinoid (synthetic and endogenous) ligands, may serve as novel candidates for this need. Cannabinoid appears to regulate seizure activity in the brain through the activation of CB1 and CB2 cannabinoid receptors (CB1R and CB2R). An abundant series of cannabinoid analogues have been tested in various animal models, including the rat pilocarpine model of acquired epilepsy, a pentylenetetrazol model of myoclonic seizures in mice, and a penicillin-induced model of epileptiform activity in the rats. The accumulating lines of evidence show that cannabinoid ligands exhibit significant benefits to control seizure activity in different epileptic models. In this review, we summarize the relationship between brain CB2 receptors and seizures and emphasize the potential mechanisms of their therapeutic effects involving the influences of neurons, astrocytes, and microglia cells. The unique features of CB2Rs, such as lower expression levels under physiological conditions and high inducibility under epileptic conditions, make it an important target for future research on drug-resistant epilepsy.
1. Introduction
Epilepsy is the third most common chronic neurological disorder that affects over 70 million people worldwide [1]. Occurrence of epileptic seizure in different brain areas may lead to loss of consciousness, motor or sensory disorders, and emotional or cognitive dysfunction [2]. Despite much progress in medical treatment using antiseizure drugs to control epileptic seizures, it still remains at 30% of patients that fail to be controlled by or respond to antiseizure drugs [3]. Thus, there is an urgent need to develop new medicines to control refractory epilepsy.
More understanding of underlying mechanisms in epileptogenesis has identified cellular and molecular targets for new therapies, for example, anti-inflammatory drugs that can overcome the limitations of current drugs and provide symptomatic control of epileptic seizures [4]. Accumulating data have demonstrated that cannabinoid systems, including endocannabinoids, anandamide, and 2-arachidonoyl glycerol, and their targets, the cannabinoid receptor subtype 1 (CB1R) and subtype 2 (CB2R), appear to regulate seizure activity [5,6,7,8,9,10,11,12,13]. The rationale for the antiseizure effects of the cannabinoid system is that the CB1Rs (possibly also CB2Rs) are linked to an inhibitory G protein (Gi/o) signaling, which reduces neuronal excitability and/or neural synchronization. For example, the activation of brain CB1R modulates A-type K+ channels and N- and P/Q-type voltage-gated Ca2+ currents and stabilizes the membrane potentials [14,15], and it modulates presynaptic neurotransmitter release [16,17,18]. Furthermore, cannabidiol has been shown to not only reduce the frequency of seizures in animal models of epilepsy but also greatly decrease the frequency of drop seizures among children and adults with Lennox–Gastaut syndrome [19]. Based on these concepts, numerous cannabinoid analogues have been examined in a variety of animal models [5,8,11,20,21,22,23]. However, although cannabinoid ligands and CB1R agonists possess some antiseizure effects, non-specific modulations of cannabinoid systems will limit their therapeutic use for treatment of human epilepsy because of their severe adverse effects, for example, THC (50 mg or more) has been shown to lead to anxiety, psychosis, heart attack, and irregular heart rhythm. Regularly taking large amounts of cannabis over a long period of time might cause a cannabinoid hyperemesis syndrome. Therefore, significant attention is currently being directed toward the possibility of developing medicines from compounds that can selectively activate CB2Rs and have important potential therapeutic applications at doses that induce little or no CB1R-mediated effects.
In this review, we summarize the current state of knowledge on CB2R expression and function, which could serve as an important tool for intervention and control of seizure activity by modulating neuronal excitability and neuroinflammation.
2. CB2R Expression and Inducible Feature
Cannabinoid receptor type 2 (CB2R) is a plasma membrane G protein-coupled receptor that was characterized from spleen by Munro [24]. The expression and function of CB2 in the brain have been debated due to early studies implying that CB2Rs were missing in the central nervous system (CNS), since CB2R mRNA expression was not measured in rat brains by using in situ hybridization [24]. In accordance with this result, Northern blot analysis also failed to detect CB2R mRNA in rat, mouse, and human brains [25,26,27]. RT-PCR experiments demonstrated abundant CB2R expression among peripheral immune tissues, such as on spleen T cells and on macrophages, but barely measurable levels in rodent brains [25,26,28,29]. Little is revealed about CB2Rs receptor expression in microglia, astrocytes, and astrocytoma, and the activation of these receptors affecting cellular function and activity [28]. Based on the above research, CB2Rs have been classically considered as peripheral cannabinoid receptors [24,30,31]. Recently, this concept of CB2 deficiency within the brain has been challenged along with the identification of CB2Rs widespread in the CNS, though they are expressed at lower densities than CB1. Emerging evidence shows that significant CB2R mRNA can be detected by ISH in cultured granule cells among the granule layer and Purkinje cell layer of the mouse cerebellum [29], in the mouse retina [32], and in the globus pallidus of non-human primates [30]. RT-PCR analysis has also been applied to distinguish CB2 mRNA expression in multiple brain regions, including the retina [32], cortex [30,31,33,34], striatum [26,34], hippocampus [30], amygdala [33,34], brainstem [35], and cerebellum [36]. Furthermore, two CB2 isoforms, CB2A and CB2B, have been characterized in the rodent and human brain [31] along with a new CB2 transcript that has been found in mouse and monkey B lymphocytes [37]. This suggests the possibility that CB2R expression not only exists in peripheral tissues, but also in the brain. It has been reported that CB2Rs modulate a variety of important processes in dopamine (DA)-related behaviors [38], including food intake [39,40,41,42], anxiety [33,43], depression [44], and schizophrenia-like behavior [34,45]. Recent evidences emerging from several laboratories, including ours, have indicated that brain CB2Rs play a pivotal role in the reduction of cocaine, alcohol, and nicotine addiction [46,47,48]. Collectively, these lines of evidence strongly suggest an important role of CB2R in the mesocorticolimbic system, as well as in various brain functions involving psychiatric, cognitive, and neurobiological activity. Compared to CB1Rs, central CB2Rs display the following unique features: (1) low expression grades, suggesting that they may not modulate neural functions under physiological conditions, (2) high pathological expression, meaning that under some pathological conditions (for example, addiction, stroke, stress, schizophrenia, inflammation, anxiety), CB2R expression increased in the brain [49], suggesting that the change of CB2R expression/function is closely related to various mental and neurological diseases, and (3) post-synaptic localization, where CB2R is mainly expressed in the neuronal somatodendritic area [50], whereas CB1Rs are chiefly expressed on neuronal terminals, especially on GABAergic terminals (presynaptic), which leads to some opposing effects after activation by these two receptor subtypes [51]. In consideration of these features, CB2R is likely an important target for neuroprotection [52], and targeting CB2Rs likely provide a novel therapeutic strategy for treating neuropsychiatric and neurological diseases without typical CB1-mediated side-effects. However, to fulfill this possibility, an understanding of the functional effects of CB2Rs in the brain is required. Unfortunately, the function of CB2Rs in the CNS has not been well-established, and studies of the functional effects of CB2Rs in neurons have ignited debate and controversy. A consensus has yet to emerge regarding the expression and function of CB2Rs in midbrain ventral tegmental area (VTA) neurons, which are the source of mesocorticolimbic dopamine (DA) signaling. In our recent study, we found that functional CB2Rs are expressed in VTA DA neurons, and the activation of these CB2Rs reduced the excitability of DA neurons through both intrinsic and synaptic mechanisms [53].
3. Drug Resistance in Epilepsy
The occurrence of epilepsy is usually related to a disorder of excitatory and/or inhibitory neurotransmitters, such as upregulation of glutamate and acetylcholine, and downregulation of GABA and serotonin. According to this concept, about 40 anti-seizure drugs are used for symptomatic treatment of epilepsy [54]. However, over one third of epileptic patients are resistant to multiple antiseizure drug therapies. Drug-resistant epilepsy can be defined as a failure of multiple efficient treatments of tolerated, chosen, and appropriately used anti-epileptic drug guidelines [55]. Based on the inducement and hypothetical mechanisms of drug-resistant epilepsy, recent speculations of mechanisms of drug resistance include: (1) Drug target variation: drug targets evoke the alteration of neurotransmitter receptors, voltage-dependent ion channels, and transporters participating in the metabolic pathway involved in the metabolism of neurotransmitters [56]. (2) Genetic mechanisms: The epigenome is a dynamic process, and endogenous mutations in receptor genes may be considered to cause the occurrence of drug-resistant epilepsy [57]. The roles in microbiome are also of great interest in epileptic disorders [58], and earlier research demonstrated that the effects of ketogenic diets for treatment of seizures may be involved in microbial processes [59]. (3) The drug target is missed because most of the anti-epileptic drugs focus on the neuronal inhibition and excitation but do not pay attention to the real pathogenesis caused by encephalitis or cancer. In view of patients with drug-resistant epilepsy, the alternative treatment methods mainly include: (1) surgical treatment via removal of epileptic foci [60], where with the development of epileptic area localization and imaging technology, as well as the in-depth research on the resistance mechanism of epilepsy, comprehensive treatment based on surgery will continue to be improved and promoted, (2) transcranial magnetic stimulation, where the combination of transcranial magnetic stimulation (TMS) and motor cortical EEG enables biomarkers to provide cortical stimulation and suppression measures that are particularly relevant to epilepsy [61], (3) an embedded stimulating electrode, applied to interfere with the synchronization process of abnormal nerve cells’ discharge [62] or for provocation of the electrical current to the vagus nerve [63], and (4) ketogenic diets, which imitate the fasting state by taking in fat as the chief fuel source therapy [64].
4. Cannabinoid System as a Potential Therapeutic Target for Treating Epilepsy
4.1. Endocannabinoid System
The endocannabinoid system (ECS) is involved in regulation of excitatory and inhibitory synaptic transmission in the brain [65,66], and consists of two G protein-coupled receptors, CB1R and CB2R, with two known endogenous cannabinoid ligands, namely 2-arachidonoylglycerol (2-AG) and N-arachidonoylethanolamide (NAN), respectively [10,11]. In the past few years, scientists have drawn attention to using a treatment focused on ECS [67,68]. There is already a comprehensive review indicating the roles of ECS dysfunction-induced neuroinflammation in epilepsy [69]. Anticonvulsant-like effects of cannabinoid receptor agonists are dependent on CB1R. CB1 agonists could increase ATP-sensitive K+ channel (KATP) activation by decreasing mitochondrial ATP levels. CB1R-mediated regulation in neuronal excitability can exert antiseizure effects [70]. Microglia are evoked by pathogens, products of damaged/inflammatory neurons, and destruction of the blood–brain barrier, as well as diverse chemical menace signals. However, there is also a contradiction about suppressing epilepsy by adjusting the activity of CB1Rs to affect excitability. Owing to the extensive distribution and high level of CB1 in the CNS under physiological conditions, a risk of side-effects develops when CB1Rs are activated and targeted.
4.2. Cannabinoids’ Effects on Epilepsy
Tetrahydrocannabinol (THC) and cannabidiol (CBD) have been the most researched at present, especially in psychoactive pharmacology. THC and CBD have many similarities in structure, and their structure–activity relationship has an impact on mental activity. It has been reported that both have certain anticonvulsant effects. THC mainly activates the GPR55 receptor, and partially activates the CB1Rs and CB2Rs. However, THC can cause a series of psychological effects, such as coordination problems, slower reaction times, memory loss, anxiety, and addiction. CBD can reduce these side-effects caused by THC. Studies have reported that CBD treats the frequency of spontaneous seizures in DS mice mainly by improving the excitability of hippocampal interneurons, and the excitability of vertebral neurons in the dentate gyrus to strong depolarization stimulation is also reduced [67]. The pharmacological activity of CBD is mainly focused on blocking the effect of GPR55 receptors to inhibit the effect of neurotransmission and significantly reduce the α-amino-3-hydroxy-5-methyl-4-isoxazole propionic acid receptor (AMPAR)-mediated amplitude and frequency of induced excitatory postsynaptic currents (EPSCs) and miniature EPSCs (mEPSCs) [71]. CBD is also used clinically to treat epileptic seizures caused by Lennox-Gastaut syndrome [19]. A proportion of patients allocated to CBD treatment in these trials were receiving clobazam (CLB). Bialer and Perucca showed an antiseizure effect of CBD independent of an interaction with CLB. A greater antiseizure effect and greater adverse effects were observed when CBD was combined with CLB [72]. However, the therapeutic effect of CBD on other types of epilepsy and the complete mechanism of action remains unclear, but it may involve a non-CLB mechanism. The peripheral side-effect of CBD is mainly diarrhea. There is only indirect proof implying that CBD could modulate endocannabinoid signaling, but no promising data indicating a direct binding or interaction between CBD and CB receptors. CBD can also partially activate CB2Rs [73]. Negative allosteric modulation of CB2R activity by CBD might interpret its action for antiseizure and other neural disorders, which provides us with novel insights to develop its medical application [74]. Regarding CB2R, CBD-DMH, a modification on different pharmacophoric sties, was considered to promote a conformational change in CB2R, which favors G protein-dependent signaling rather than β-arrestin-dependent signaling [75]. However, there are very few modulators of CB1R and CB2R reported in the literature.
4.3. CB2R Effects on Preclinical Epilepsy
Since CB2Rs exhibit low expression levels in the brain under normal conditions, but are highly inducible during various disease states (including epilepsy), they appear to be an important substrate for neuroprotection [76]. Targeting CB2Rs will likely offer a novel therapeutic strategy for treating epileptic seizures without the typical CB1R-mediated side-effects [77,78]. Emerging evidence has indicated that CB2Rs are involved in epileptic activity in animal models. In an acute pentylenetetrazol (PTZ) rat seizure model, pretreatment with palmitoyl ethanolamide (PEA) increased the latency of seizure initiation and reduced the duration of seizures, and this antiseizure effect was attenuated by the CB2R antagonist (AM630), suggesting that CB2Rs mediate PEA’s effect [79]. In developing rats, Huizenga et al. examined the antiseizure effects of a variety of cannabinoid ligands, and found that either combined CB1R/CB2R or selective CB1R agonists exhibited antiseizure effects in either chemo-convulsing methyl-6,7-dimethoxy-4-ethyl-beta-carboline-3-carboxylate or PTZ seizure models of 10-day postnatal rats [80]. Although the CB2R selective agonist HU-308 did not show an antiseizure effect, the CB2R selective antagonist AM630 did increase seizure severity [80]. In addition, a recent report showed that CB1R knockout (KO) mice did not have an epilepsy phenotype, but co-KO of CB1R and CB2R caused animal epilepsy [81], suggesting that CB2R plays a role in stabilizing the neuronal system. Recent studies have also explored the effects of modulating CB2R activity on seizure susceptibility (Table 1). The activation of CB2Rs decreases excitatory synaptic transmission in the CNS. The new roles for CB2R have been identified in inducing hippocampal pyramidal cell hyperpolarization and inhibiting epileptic seizures [12]. The CB2Rs expressed on hippocampal CA3 neurons also play a critical role in reduced neuronal excitation and oscillations [82]. WIN 55212-2, a non-selective CB receptor agonist, shows striking antiseizure effects in a rat epileptic model [83], and CB1R and CB2R double-knockout mice show spontaneous or manual-evoked seizures [81]. CB2R knockout mice including both heterozygous and homozygous exhibit enhanced epileptic susceptibility, and a reduction in CB2R activity is associated with increased susceptibility [84], suggesting that absent CB2Rs can increase seizure susceptibility. The administration of caryophyllene, a CB2R agonist, was found to improve seizure activity in a mouse model [85]. Collectively, these lines of evidence support the idea that the activation of CB2Rs exhibits the antiseizure role. On the other hand, some studies reported different responses by CB2R agonists. For instance, HU-308 [80] and JHW133 [84] show no significant effect on mice seizure occurrence. Moreover, CB2R agonist AM1241 increases seizure intensity in a PTZ model [86]. Additionally, CB2R antagonists AM630 and SR144528 can increase seizure susceptibility [80]. Therefore, it is likely that the altered CB2R activity can affect seizure susceptibility, although the underlying mechanisms are still unclear. The different effects of CB2R-mediated modulations may come from the differences in using different types of CB2R ligands, and be based on different experimental designs, including species, epileptic model types, and dosage. We recently found that the commercially available CB2R agonists showed different effects on pancreatic acinar cell Ca2+ oscillations [87]. Nevertheless, numerous lines of evidence have demonstrated that CB2R agonists ameliorate a variety of epileptic seizures, suggesting that CB2R is a potential therapeutic target for treating epilepsy.
4.4. CB2R Mediated Anti-Epileptic Effects through a Reduction of Neural Excitability and Synchronization
As a crucial neuro-modulatory system of the brain, the midbrain dopaminergic system plays an important role in neuronal excitability. Temporal lobe epilepsy, involving pathological erethism of the hippocampus, is associated with VTA dopamine neuron activation [88]. The abnormally high synchronous activity of neuronal firing will cause phased impulse stimulation, causing more dopamine neurons to produce impulse firing, so that the dopaminergic system is in a super-reactive state during seizures’ occurrence [89]. This suggests that the dopaminergic system is vital in epileptic brains, and the evidence for a relationship between epilepsy and the dopaminergic system was previously described by Rezaei [2]. Pilocarpine-induced epileptic rats exhibit a significant enhancement in activity of dopaminergic neurons [88]. In the PTZ kindling model of epilepsy in mice, dopamine neurons within the VTA display hyperactivity when compared to saline-injected controls [90]. Anti-seizure treatments (antiseizure drugs or brain stimulation) are applied to downregulate the neuronal excitability for controlling epileptic seizures.
Stimulation of Gs-linked G protein-coupled receptors (GPCRs) triggers adenylate cyclase (AC) to produce cAMP from ATP. Activation of this signal pathway causes calcium influx through plasma membrane channels in a calmodulin-dependent manner [91]. Recent studies using transcriptomic analysis show that the transcription of a set of genes related to cAMP signaling is changed in patients suffering from drug-resistant temporal lobe epilepsy [92,93]. When seizures occur, levels of cAMP increase in the brain. The appearance of cAMP signaling can be divided into temporary influence on neuronal excitability, including ion channel or receptor phosphorylation [75], and long-term effects on epileptogenesis, such as effects regulated by cAMP response factor binding protein, CREB [94]. By coupling to Gi proteins, the activation of CB2Rs causes inhibition of AC activity and downregulation of cAMP release [95]. 5-HA1A-CB2 heteroreceptors were characterized in cortical primary cultures of neurons, and 5-HT1AR-CB2 heteroreceptor complex expression and functionality are significantly enhanced in the brain after cerebral ischemia, especially during the neonatal term, also suggesting that this heteromer is associated with NHIBD pathophysiology [96]. Therefore, CB2R activation may suppress the occurrence of epilepsy via decreasing excitability of the CNS by reducing the level of cAMP.
Most K+ channels are controlled by various physiological mediators, such as transmembrane voltage, and intracellular Ca2+ and G proteins. The roles of K+ in membrane physiology have been extensively investigated in rodent models, and the basic electrophysiological properties and bursting patterns of primate central neurons are generally similar to those reported for the rodent [97]. K+ channels are very important in regulating the intrinsic excitability of neurons, and they are the main contributor to neuronal membrane repolarization [98]. K+ channels are a promising target for the development of novel anti-epileptic drugs, and activation of the K+ channels can be used for restoring control on neuronal excitability in patients with epilepsy [99,100]. Kv7 (M-) channels are an unusual type of K+ channel which is different from those that repolarize an individual’s action potential. The M-channels are partially activated within the resting membrane potential of neurons, and further activated by membrane depolarization. The rate of M-channel open and close is slower than other types of K+ channels that contribute to the repolarization of action potentials. Activation of M-channels can inhibit highly synchronized neuronal firing that may reduce hyper-excitation and seizure activity [101]. The K+ channel opener retigabine is a compelling and selective opener of M-type K+ channels and is approved for therapy of drug-resistant focal and focal-to-bilateral tonic-clonic seizures [102]. In VTA DA neurons, G protein-coupled receptor signals regulate the excitability of neurons through a few ion channels, such as G protein-gated internally rectified K+ channels (GIRKs) [50,51,52]. The CB2R agonist JWH133 has been confirmed to effectively modulate the excitability of neurons by regulating voltage-dependent M-type K+ channels [53].
4.5. Glia CB2R-Mediated Anti-Epileptic Effects via Inflammation and Excitability
More and more lines of evidence show that CB2Rs are relevant to both immune cell competence at the peripheral region [103] and brain cells in the CNS. Actually, neurons, microglia, and astrocytes cells express CB2Rs [32,104,105], which are capable of modulating central neural-immune function and impact the related diseases [106,107]. It is critical to accentuate that CB2R has inducible expression in several immune cells under activated neuroinflammatory conditions. It means that CB2R levels may be upregulated in the CNS and increased in inflamed brain parenchyma due to the invasion of peripheral immune cells (such as peripheral T cells) that express CB2Rs [107]. During activated processes, microglia increase the expression of an array of membrane surfaces of CB2Rs that may be essential in microglial production and/or degeneration within the brain. Both CB1 and CB2 receptors were expressed on microglia using an in vitro assay, including immunoglobulin superfamily receptors, cell component receptors, toll-related receptors, opioid receptors, and cannabinoid receptors [13]. A notable example is that neuropathic pain upregulates CB2Rs in microglia in rat spinal cord, while chronic inflammatory pain does not [108]. When there is an inflammation response in the body, microglia CB2R is rapidly upregulated and activated, which effectively inhibits the release of harmful factors, including TNF-α and free radicals [103].
In patients with epilepsy, medial temporal lobe sclerosis, cortical dysplasia, encephalitis, and glioma, the astrocytes show increased expression of CB2Rs, which may change in K+ currents during seizures. It may lead to overexcitation and changes in a series of enzymatic pathways, which means that astrocytes may change the M-type K+ current by adjusting the expression of CB2Rs to change the intensity of epilepsy. We believe that the role of the astrocyte/astrocytic CB2R–cAMP signaling pathway in controlling epileptogenesis is worthy of further exploration.
5. Conclusions and Perspective
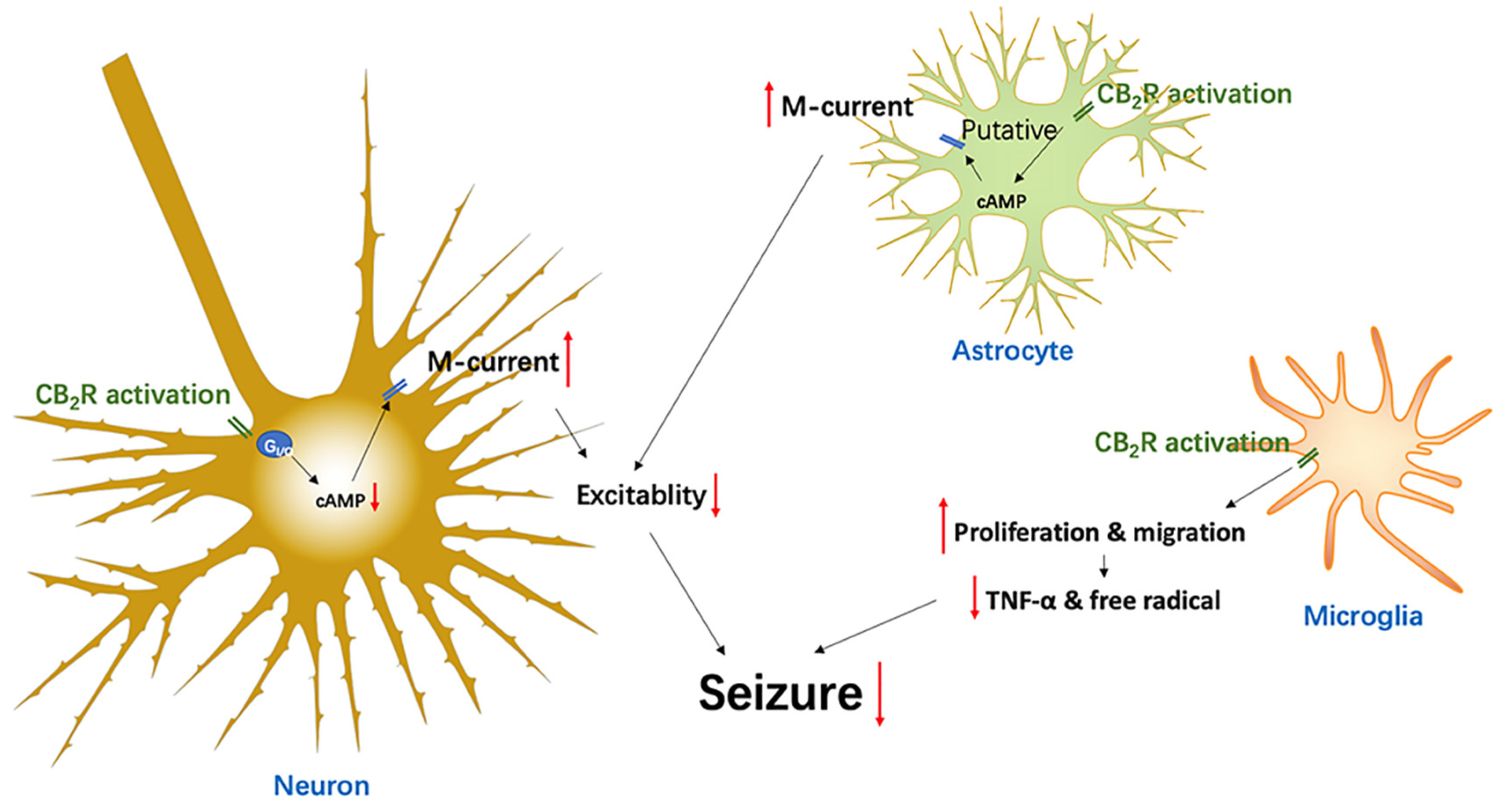
In early studies, CB2Rs were found in the peripheral region, while CB1Rs were mainly expressed in the CNS, which led to a question of the existence of the CB2Rs in the CNS. Nowadays, with the development of more sensitive detection technologies, CB2Rs have been found in multiple brain regions in the CNS, though at a low level of expression compared to CB1Rs. However, CB2R expression and function are rapidly and profoundly increased under pathological conditions in the CNS. This attractive feature means that CB2R is considered as a disease-associated target, suggesting that it will greatly reduce the occurrence of CB1R’s side-effects through modulating the activity of CB2Rs to improve the neurological disorders. Drug-resistant epilepsy seriously affects the quality of life of patients, which highlights the need to invent more effective treatments. Although the underlying mechanisms of drug-resistant epilepsy are still unclear, several novel medicines to improve drug-resistant epilepsy have been developed. One example is the CB1 agonist, THC, which has been replaced by CBD due to its psychoactive side-effect. Epidiolex has been recently approved for the treatment of rare, severe forms of epilepsy by the USA’s Food and Drug Administration (FDA) [109]. Although this shows a promise of targeting the endocannabinoid system as a novel anti-seizure treatment, some harmful side-effects, including somnolence, diarrhea, appetite inhibition, and increased levels of hepatic transaminase [104] and blood pressure [105], limit its use. Considering that CB2Rs exhibit low expression levels in the brain under normal conditions, but are highly inducible during various disease states (including epilepsy), they appear to be an important substrate for neuroprotection [76]. Targeting CB2Rs will likely offer a novel therapeutic strategy for treating epileptic seizures without the typical CB1R-mediated side-effects [77]. Figure 1 summarizes the potential mechanisms of CB2R activation to inhibit seizures, that include reduced neuronal excitability by downregulating cAMP, and consequently enhanced M-currents in both neurons and astrocytes. In addition, CB2Rs can also regulate immune function and slow down neuroinflammatory responses. Together, it suggests that the CB2R may be an important target for controlling epileptic seizures.
Author Contributions
Conceptualization, J.W. and X.J.; investigation, X.J. and Y.Z.; writing—original draft preparation, X.J.; writing—review and editing, J.W. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the 2020 Li Ka Shing Foundation Cross-Disciplinary Research Grant (2020LKSFG01A) and The Shantou City Science and Technology plan for Medical and Health Project (200622165260881).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
Authors thank Stanley Lin for his careful editing and correction of English language in this paper.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Espinosa-Jovel, C.; Toledano, R.; Aledo-Serrano, A.; Garcia-Morales, I.; Gil-Nagel, A. Epidemiological profile of epilepsy in low income populations. Seizure 2018, 56, 67–72. [Google Scholar] [CrossRef] [Green Version]
- Rezaei, M.; Sadeghian, A.; Roohi, N.; Shojaei, A.; Mirnajafi-Zadeh, J. Epilepsy and dopaminergic system. Physiol. Pharmacol. 2017, 21, 1–14. [Google Scholar]
- Devinsky, O.; Vezzani, A.; O’Brien, T.J.; Jette, N.; Scheffer, I.E.; de Curtis, M.; Perucca, P. Epilepsy. Nat. Rev. Dis. Primers 2018, 4, 18024. [Google Scholar] [CrossRef] [PubMed]
- Vezzani, A.; Balosso, S.; Ravizza, T. Neuroinflammatory pathways as treatment targets and biomarkers in epilepsy. Nat. Rev. Neurol. 2019, 15, 459–472. [Google Scholar] [CrossRef]
- Wallace, M.J.; Blair, R.E.; Falenski, K.W.; Martin, B.R.; DeLorenzo, R.J. The endogenous cannabinoid system regulates seizure frequency and duration in a model of temporal lobe epilepsy. J. Pharm. Exp. 2003, 307, 129–137. [Google Scholar] [CrossRef] [Green Version]
- Deshpande, L.S.; Sombati, S.; Blair, R.E.; Carter, D.S.; Martin, B.R.; DeLorenzo, R.J. Cannabinoid CB1 receptor antagonists cause status epilepticus-like activity in the hippocampal neuronal culture model of acquired epilepsy. Neurosci. Lett. 2007, 411, 11–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kozan, R.; Ayyildiz, M.; Agar, E. The effects of intracerebroventricular AM-251, a CB1-receptor antagonist, and ACEA, a CB1-receptor agonist, on penicillin-induced epileptiform activity in rats. Epilepsia 2009, 50, 1760–1767. [Google Scholar] [CrossRef] [PubMed]
- Cakil, D.; Yildirim, M.; Ayyildiz, M.; Agar, E. The effect of co-administration of the NMDA blocker with agonist and antagonist of CB1-receptor on penicillin-induced epileptiform activity in rats. Epilepsy Res. 2011, 93, 128–137. [Google Scholar] [CrossRef]
- Capasso, A. Do Cannabinoids Confer Neuroprotection Against Epilepsy? An Overview. Open Neurol. J. 2017, 11, 61–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neale, M. Efficacy and safety of cannabis for treating children with refractory epilepsy. Nurs. Child Young People 2017, 29, 32–37. [Google Scholar] [CrossRef]
- De Caro, C.; Leo, A.; Citraro, R.; De Sarro, C.; Russo, R.; Calignano, A.; Russo, E. The potential role of cannabinoids in epilepsy treatment. Expert Rev. Neurother. 2017, 17, 1069–1079. [Google Scholar] [CrossRef] [PubMed]
- Sugaya, Y.; Yamazaki, M.; Uchigashima, M.; Kobayashi, K.; Watanabe, M.; Sakimura, K.; Kano, M. Crucial Roles of the Endocannabinoid 2-Arachidonoylglycerol in the Suppression of Epileptic Seizures. Cell Rep. 2016, 16, 1405–1415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bie, B.; Wu, J.; Foss, J.F.; Naguib, M. An overview of the cannabinoid type 2 receptor system and its therapeutic potential. Curr. Opin. Anaesthesiol. 2018, 31, 407–414. [Google Scholar] [CrossRef]
- Qian, W.J.; Yin, N.; Gao, F.; Miao, Y.; Li, Q.; Li, F.; Sun, X.H.; Yang, X.L.; Wang, Z. Cannabinoid CB1 and CB2 receptors differentially modulate L- and T-type Ca(2+) channels in rat retinal ganglion cells. Neuropharmacology 2017, 124, 143–156. [Google Scholar] [CrossRef] [PubMed]
- Yang, K.; Wang, C.; Sun, T. The Roles of Intracellular Chaperone Proteins, Sigma Receptors, in Parkinson’s Disease (PD) and Major Depressive Disorder (MDD). Front. Pharm. 2019, 10, 528. [Google Scholar] [CrossRef] [Green Version]
- Kreitzer, A.C.; Regehr, W.G. Retrograde inhibition of presynaptic calcium influx by endogenous cannabinoids at excitatory synapses onto Purkinje cells. Neuron 2001, 29, 717–727. [Google Scholar] [CrossRef] [Green Version]
- Wilson, R.I.; Nicoll, R.A. Endogenous cannabinoids mediate retrograde signalling at hippocampal synapses. Nature 2001, 410, 588–592. [Google Scholar] [CrossRef]
- Wilson, R.I.; Kunos, G.; Nicoll, R.A. Presynaptic specificity of endocannabinoid signaling in the hippocampus. Neuron 2001, 31, 453–462. [Google Scholar] [CrossRef] [Green Version]
- Devinsky, O.; Patel, A.D.; Cross, J.H.; Villanueva, V.; Wirrell, E.C.; Privitera, M.; Greenwood, S.M.; Roberts, C.; Checketts, D.; VanLandingham, K.E.; et al. Effect of Cannabidiol on Drop Seizures in the Lennox-Gastaut Syndrome. N. Engl. J. Med. 2018, 378, 1888–1897. [Google Scholar] [CrossRef] [Green Version]
- Arslan, G.; Ayyildiz, M.; Agar, E. The interaction between ghrelin and cannabinoid systems in penicillin-induced epileptiform activity in rats. Neuropeptides 2014, 48, 345–352. [Google Scholar] [CrossRef]
- Devinsky, O.; Marsh, E.; Friedman, D.; Thiele, E.; Laux, L.; Sullivan, J.; Miller, I.; Flamini, R.; Wilfong, A.; Filloux, F.; et al. Cannabidiol in patients with treatment-resistant epilepsy: An open-label interventional trial. Lancet Neurol. 2016, 15, 270–278. [Google Scholar] [CrossRef]
- Devinsky, O.; Cross, J.H.; Wright, S. Trial of Cannabidiol for Drug-Resistant Seizures in the Dravet Syndrome. N. Engl. J. Med. 2017, 377, 699–700. [Google Scholar] [CrossRef] [Green Version]
- Szaflarski, M.; Hansen, B.; Bebin, E.M.; Szaflarski, J.P. Social correlates of health status, quality of life, and mood states in patients treated with cannabidiol for epilepsy. Epilepsy Behav. 2017, 70, 364–369. [Google Scholar] [CrossRef]
- Munro, S.; Thomas, K.L.; Abu-Shaar, M. Molecular characterization of a peripheral receptor for cannabinoids. Nature 1993, 365, 61–65. [Google Scholar] [CrossRef]
- Schatz, A.R.; Lee, M.; Condie, R.B.; Pulaski, J.T.; Kaminski, N.E. Cannabinoid receptors CB1 and CB2: A characterization of expression and adenylate cyclase modulation within the immune system. Toxicol. Appl. Pharm. 1997, 142, 278–287. [Google Scholar] [CrossRef]
- Galiegue, S.; Mary, S.; Marchand, J.; Dussossoy, D.; Carriere, D.; Carayon, P.; Bouaboula, M.; Shire, D.; Le Fur, G.; Casellas, P. Expression of central and peripheral cannabinoid receptors in human immune tissues and leukocyte subpopulations. Eur. J. Biochem. 1995, 232, 54–61. [Google Scholar] [CrossRef]
- Griffin, G.; Wray, E.J.; Tao, Q.; McAllister, S.D.; Rorrer, W.K.; Aung, M.M.; Martin, B.R.; Abood, M.E. Evaluation of the cannabinoid CB2 receptor-selective antagonist, SR144528: Further evidence for cannabinoid CB2 receptor absence in the rat central nervous system. Eur. J. Pharm. 1999, 377, 117–125. [Google Scholar] [CrossRef]
- Stella, N. Cannabinoid and cannabinoid-like receptors in microglia, astrocytes, and astrocytomas. Glia 2010, 58, 1017–1030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skaper, S.D.; Buriani, A.; Dal Toso, R.; Petrelli, L.; Romanello, S.; Facci, L.; Leon, A. The ALIAmide palmitoylethanolamide and cannabinoids, but not anandamide, are protective in a delayed postglutamate paradigm of excitotoxic death in cerebellar granule neurons. Proc. Natl. Acad. Sci. USA 1996, 93, 3984–3989. [Google Scholar] [CrossRef] [Green Version]
- Lanciego, J.L.; Barroso-Chinea, P.; Rico, A.J.; Conte-Perales, L.; Callen, L.; Roda, E.; Gomez-Bautista, V.; Lopez, I.P.; Lluis, C.; Labandeira-Garcia, J.L.; et al. Expression of the mRNA coding the cannabinoid receptor 2 in the pallidal complex of Macaca fascicularis. J. Psychopharmacol. 2011, 25, 97–104. [Google Scholar] [CrossRef]
- Liu, Q.R.; Pan, C.H.; Hishimoto, A.; Li, C.Y.; Xi, Z.X.; Llorente-Berzal, A.; Viveros, M.P.; Ishiguro, H.; Arinami, T.; Onaivi, E.S.; et al. Species differences in cannabinoid receptor 2 (CNR2 gene): Identification of novel human and rodent CB2 isoforms, differential tissue expression and regulation by cannabinoid receptor ligands. Genes Brain Behav. 2009, 8, 519–530. [Google Scholar] [CrossRef]
- Lu, Q.; Straiker, A.; Lu, Q.; Maguire, G. Expression of CB2 cannabinoid receptor mRNA in adult rat retina. Vis. Neurosci. 2000, 17, 91–95. [Google Scholar] [CrossRef]
- Garcia-Gutierrez, M.S.; Garcia-Bueno, B.; Zoppi, S.; Leza, J.C.; Manzanares, J. Chronic blockade of cannabinoid CB(2) receptors induces anxiolytic-like actions associated to alterations in GABA(A) receptors. Br. J. Pharm. 2011, 165, 951–964. [Google Scholar] [CrossRef] [Green Version]
- Navarrete, F.; Perez-Ortiz, J.M.; Manzanares, J. Cannabinoid CB(2) receptor-mediated regulation of impulsive-like behaviour in DBA/2 mice. Br. J. Pharm. 2012, 165, 260–273. [Google Scholar] [CrossRef] [Green Version]
- Van Sickle, M.D.; Duncan, M.; Kingsley, P.J.; Mouihate, A.; Urbani, P.; Mackie, K.; Stella, N.; Makriyannis, A.; Piomelli, D.; Davison, J.S.; et al. Identification and functional characterization of brainstem cannabinoid CB2 receptors. Science 2005, 310, 329–332. [Google Scholar] [CrossRef] [Green Version]
- Viscomi, M.T.; Oddi, S.; Latini, L.; Pasquariello, N.; Florenzano, F.; Bernardi, G.; Molinari, M.; Maccarrone, M. Selective CB2 receptor agonism protects central neurons from remote axotomy-induced apoptosis through the PI3K/Akt pathway. J. Neurosci. 2009, 29, 4564–4570. [Google Scholar] [CrossRef] [Green Version]
- Sherwood, T.A.; Nong, L.; Agudelo, M.; Newton, C.; Widen, R.; Klein, T.W. Identification of transcription start sites and preferential expression of select CB2 transcripts in mouse and human B lymphocytes. J. Neuroimmune Pharm. 2009, 4, 476–488. [Google Scholar] [CrossRef]
- Canseco-Alba, A.; Schanz, N.; Sanabria, B.; Zhao, J.; Lin, Z.; Liu, Q.R.; Onaivi, E.S. Behavioral effects of psychostimulants in mutant mice with cell-type specific deletion of CB2 cannabinoid receptors in dopamine neurons. Behav. Brain Res. 2019, 360, 286–297. [Google Scholar] [CrossRef]
- Agudo, J.; Martin, M.; Roca, C.; Molas, M.; Bura, A.S.; Zimmer, A.; Bosch, F.; Maldonado, R. Deficiency of CB2 cannabinoid receptor in mice improves insulin sensitivity but increases food intake and obesity with age. Diabetologia 2010, 53, 2629–2640. [Google Scholar] [CrossRef] [Green Version]
- Ignatowska-Jankowska, B.; Jankowski, M.M.; Swiergiel, A.H. Cannabidiol decreases body weight gain in rats: Involvement of CB2 receptors. Neurosci. Lett. 2011, 490, 82–84. [Google Scholar] [CrossRef] [PubMed]
- Emadi, L.; Jonaidi, H.; Hosseini Amir Abad, E. The role of central CB2 cannabinoid receptors on food intake in neonatal chicks. J. Comp. Physiol. A Neuroethol. Sens. Neural Behav. Physiol. 2011, 197, 1143–1147. [Google Scholar] [CrossRef]
- Flake, N.M.; Zweifel, L.S. Behavioral effects of pulp exposure in mice lacking cannabinoid receptor 2. J. Endod. 2012, 38, 86–90. [Google Scholar] [CrossRef]
- Garcia-Gutierrez, M.S.; Manzanares, J. Overexpression of CB2 cannabinoid receptors decreased vulnerability to anxiety and impaired anxiolytic action of alprazolam in mice. J. Psychopharmacol. 2011, 25, 111–120. [Google Scholar] [CrossRef] [Green Version]
- Hwang, E.S.; Kim, H.B.; Lee, S.; Kim, M.J.; Kim, K.J.; Han, G.; Han, S.Y.; Lee, E.A.; Yoon, J.H.; Kim, D.O.; et al. Antidepressant-like effects of beta-caryophyllene on restraint plus stress-induced depression. Behav. Brain Res. 2020, 380, 112439. [Google Scholar] [CrossRef]
- Ortega-Alvaro, A.; Aracil-Fernandez, A.; Garcia-Gutierrez, M.S.; Navarrete, F.; Manzanares, J. Deletion of CB2 cannabinoid receptor induces schizophrenia-related behaviors in mice. Neuropsychopharmacology 2011, 36, 1489–1504. [Google Scholar] [CrossRef] [Green Version]
- Xi, Z.X.; Peng, X.Q.; Li, X.; Song, R.; Zhang, H.Y.; Liu, Q.R.; Yang, H.J.; Bi, G.H.; Li, J.; Gardner, E.L. Brain cannabinoid CB(2) receptors modulate cocaine’s actions in mice. Nat. Neurosci. 2011, 14, 1160–1166. [Google Scholar] [CrossRef] [Green Version]
- Navarrete, F.; Rodriguez-Arias, M.; Martin-Garcia, E.; Navarro, D.; Garcia-Gutierrez, M.S.; Aguilar, M.A.; Aracil-Fernandez, A.; Berbel, P.; Minarro, J.; Maldonado, R.; et al. Role of CB2 Cannabinoid Receptors in the Rewarding, Reinforcing, and Physical Effects of Nicotine. Neuropsychopharmacology 2013, 38, 2515–2524. [Google Scholar] [CrossRef] [Green Version]
- Ortega-Alvaro, A.; Ternianov, A.; Aracil-Fernandez, A.; Navarrete, F.; Garcia-Gutierrez, M.S.; Manzanares, J. Role of cannabinoid CB receptor in the reinforcing actions of ethanol. Addict. Biol. 2013, 20, 43–55. [Google Scholar] [CrossRef]
- Miller, L.K.; Devi, L.A. The highs and lows of cannabinoid receptor expression in disease: Mechanisms and their therapeutic implications. Pharm. Rev. 2011, 63, 461–470. [Google Scholar] [CrossRef]
- Onaivi, E.S.; Ishiguro, H.; Gong, J.P.; Patel, S.; Meozzi, P.A.; Myers, L.; Perchuk, A.; Mora, Z.; Tagliaferro, P.A.; Gardner, E.; et al. Functional expression of brain neuronal CB2 cannabinoid receptors are involved in the effects of drugs of abuse and in depression. Ann. N. Y. Acad. Sci. 2008, 1139, 434–449. [Google Scholar] [CrossRef] [Green Version]
- Onaivi, E.S.; Ishiguro, H.; Gu, S.; Liu, Q.R. CNS effects of CB2 cannabinoid receptors: Beyond neuro-immuno-cannabinoid activity. J. Psychopharmacol. 2012, 26, 92–103. [Google Scholar] [CrossRef] [Green Version]
- Spyridakos, D.; Papadogkonaki, S.; Dionysopoulou, S.; Mastrodimou, N.; Polioudaki, H.; Thermos, K. Effect of acute and subchronic administration of (R)-WIN55,212-2 induced neuroprotection and anti inflammatory actions in rat retina: CB1 and CB2 receptor involvement. Neurochem. Int. 2021, 142, 104907. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Gao, F.; Larsen, B.; Gao, M.; Luo, Z.; Chen, D.; Ma, X.; Qiu, S.; Zhou, Y.; Xie, J.; et al. Mechanisms of cannabinoid CB2 receptor-mediated reduction of dopamine neuronal excitability in mouse ventral tegmental area. EBioMedicine 2019, 42, 225–237. [Google Scholar] [CrossRef] [Green Version]
- Loscher, W.; Potschka, H.; Sisodiya, S.M.; Vezzani, A. Drug Resistance in Epilepsy: Clinical Impact, Potential Mechanisms, and New Innovative Treatment Options. Pharm. Rev. 2020, 72, 606–638. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zhu, X.; Peng, A.; Lai, W.; He, S.; Qiu, X.; Zou, X.; Chen, L. Predictors of drug-resistance in epilepsy with auditory features. Epilepsy Res. 2020, 164, 106353. [Google Scholar] [CrossRef]
- Kolker, S. Metabolism of amino acid neurotransmitters: The synaptic disorder underlying inherited metabolic diseases. J. Inherit. Metab. Dis. 2018, 41, 1055–1063. [Google Scholar] [CrossRef]
- Epi25 Collaborative. Ultra-Rare Genetic Variation in the Epilepsies: A Whole-Exome Sequencing Study of 17,606 Individuals. Am. J. Hum. Genet. 2019, 105, 267–282. [Google Scholar] [CrossRef] [Green Version]
- Iannone, L.F.; Preda, A.; Blottiere, H.M.; Clarke, G.; Albani, D.; Belcastro, V.; Carotenuto, M.; Cattaneo, A.; Citraro, R.; Ferraris, C.; et al. Microbiota-gut brain axis involvement in neuropsychiatric disorders. Expert Rev. Neurother. 2019, 19, 1037–1050. [Google Scholar] [CrossRef]
- Olson, C.A.; Vuong, H.E.; Yano, J.M.; Liang, Q.Y.; Nusbaum, D.J.; Hsiao, E.Y. The Gut Microbiota Mediates the Anti-Seizure Effects of the Ketogenic Diet. Cell 2018, 174, 497. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Tymchuk, S.; Barry, J.; Muppidi, S.; Le, S. Antibody Prevalence in Epilepsy before Surgery (APES) in drug-resistant focal epilepsy. Epilepsia 2021, 62, 720–728. [Google Scholar] [CrossRef]
- Walton, D.; Spencer, D.C.; Nevitt, S.J.; Michael, B.D. Transcranial magnetic stimulation for the treatment of epilepsy. Cochrane Database Syst. Rev. 2021, 4, CD011025. [Google Scholar]
- Fox, K.C.R.; Shi, L.; Baek, S.; Raccah, O.; Foster, B.L.; Saha, S.; Margulies, D.S.; Kucyi, A.; Parvizi, J. Intrinsic network architecture predicts the effects elicited by intracranial electrical stimulation of the human brain. Nat. Hum. Behav. 2020, 4, 1039–1052. [Google Scholar] [CrossRef]
- Lim, Z.; Wong, K.; Downs, J.; Bebbington, K.; Demarest, S.; Leonard, H. Vagus nerve stimulation for the treatment of refractory epilepsy in the CDKL5 Deficiency Disorder. Epilepsy Res. 2018, 146, 36–40. [Google Scholar] [CrossRef]
- Giordano, C.; Marchio, M.; Timofeeva, E.; Biagini, G. Neuroactive peptides as putative mediators of antiepileptic ketogenic diets. Front. Neurol. 2014, 5, 63. [Google Scholar] [CrossRef]
- Schonhofen, P.; Bristot, I.J.; Crippa, J.A.; Hallak, J.E.C.; Zuardi, A.W.; Parsons, R.B.; Klamt, F. Cannabinoid-Based Therapies and Brain Development: Potential Harmful Effect of Early Modulation of the Endocannabinoid System. CNS Drugs 2018, 32, 697–712. [Google Scholar] [CrossRef] [Green Version]
- Senn, L.; Cannazza, G.; Biagini, G. Receptors and Channels Possibly Mediating the Effects of Phytocannabinoids on Seizures and Epilepsy. Pharmaceuticals 2020, 13, 174. [Google Scholar] [CrossRef]
- Kaplan, J.S.; Stella, N.; Catterall, W.A.; Westenbroek, R.E. Cannabidiol attenuates seizures and social deficits in a mouse model of Dravet syndrome. Proc. Natl. Acad. Sci. USA 2017, 114, 11229–11234. [Google Scholar] [CrossRef] [Green Version]
- Kwan Cheung, K.A.; Peiris, H.; Wallace, G.; Holland, O.J.; Mitchell, M.D. The interplay between the endocannabinoid system, epilepsy and cannabinoids. Int. J. Mol. Sci. 2019, 20, 6079. [Google Scholar] [CrossRef] [Green Version]
- Zhu, X.; Liu, J.; Huang, S.; Zhu, W.; Wang, Y.; Chen, O.; Xue, J. Neuroprotective effects of isoliquiritigenin against cognitive impairment via suppression of synaptic dysfunction, neuronal injury, and neuroinflammation in rats with kainic acid-induced seizures. Int. Immunopharmacol. 2019, 72, 358–366. [Google Scholar] [CrossRef]
- Haj-Mirzaian, A.; Ramezanzadeh, K.; Afshari, K.; Mousapour, P.; Abbasi, N.; Haj-Mirzaian, A.; Nikbakhsh, R.; Haddadi, N.-S.; Dehpour, A.R. Activation of ATP-sensitive K-channel promotes the anticonvulsant properties of cannabinoid receptor agonist through mitochondrial ATP level reduction. Epilepsy Behav. 2019, 93, 1–6. [Google Scholar] [CrossRef]
- Yu, Y.; Yang, Z.; Jin, B.; Qin, X.; Zhu, X.; Sun, J.; Huo, L.; Wang, R.; Shi, Y.; Jia, Z. Cannabidiol inhibits febrile seizure by modulating AMPA receptor kinetics through its interaction with the N-terminal domain of GluA1/GluA2. Pharmacol. Res. 2020, 161, 105128. [Google Scholar] [CrossRef]
- Bialer, M.; Perucca, E. Does cannabidiol have antiseizure activity independent of its interactions with clobazam? An appraisal of the evidence from randomized controlled trials. Epilepsia 2020, 61, 1082–1089. [Google Scholar] [CrossRef]
- Tham, M.; Yilmaz, O.; Alaverdashvili, M.; Kelly, M.E.; Denovan-Wright, E.M.; Laprairie, R.B. Allosteric and orthosteric pharmacology of cannabidiol and cannabidiol-dimethylheptyl at the type 1 and type 2 cannabinoid receptors. Br. J. Pharmacol. 2019, 176, 1455–1469. [Google Scholar] [CrossRef] [Green Version]
- Laprairie, R.; Bagher, A.; Kelly, M.; Denovan-Wright, E. Cannabidiol is a negative allosteric modulator of the cannabinoid CB1 receptor. Br. J. Pharmacol. 2015, 172, 4790–4805. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Gao, B.; Zheng, F.; Lu, S.; Li, Y.; Xiong, Y.; Yang, Q.; Yang, Y.; Fu, P.; Xiao, F.; et al. The Phosphodiesterase 10A Inhibitor PF-2545920 Enhances Hippocampal Excitability and Seizure Activity Involving the Upregulation of GluA1 and NR2A in Post-synaptic Densities. Front. Mol. Neurosci. 2017, 10, 100. [Google Scholar] [CrossRef] [Green Version]
- Cassano, T.; Calcagnini, S.; Pace, L.; De Marco, F.; Romano, A.; Gaetani, S. Cannabinoid Receptor 2 Signaling in Neurodegenerative Disorders: From Pathogenesis to a Promising Therapeutic Target. Front. Neurosci. 2017, 11, 30. [Google Scholar] [CrossRef] [Green Version]
- Xin, Q.; Xu, F.; Taylor, D.H.; Zhao, J.F.; Wu, J. The impact of cannabinoid type 2 receptors (CB2Rs) in neuroprotection against neurological disorders. Acta Pharm. Sin. 2020, 41, 1507–1518. [Google Scholar] [CrossRef]
- Wu, J. Cannabis, cannabinoid receptors, and endocannabinoid system: Yesterday, today, and tomorrow. Acta Pharm. Sin. 2019, 40, 297–299. [Google Scholar] [CrossRef]
- Aghaei, I.; Rostampour, M.; Shabani, M.; Naderi, N.; Motamedi, F.; Babaei, P.; Khakpour-Taleghani, B. Palmitoylethanolamide attenuates PTZ-induced seizures through CB1 and CB2 receptors. Epilepsy Res. 2015, 117, 23–28. [Google Scholar] [CrossRef]
- Huizenga, M.N.; Wicker, E.; Beck, V.C.; Forcelli, P.A. Anticonvulsant effect of cannabinoid receptor agonists in models of seizures in developing rats. Epilepsia 2017, 58, 1593–1602. [Google Scholar] [CrossRef]
- Rowley, S.; Sun, X.; Lima, I.V.; Tavenier, A.; de Oliveira, A.C.P.; Dey, S.K.; Danzer, S.C. Cannabinoid receptor 1/2 double-knockout mice develop epilepsy. Epilepsia 2017, 58, e162–e166. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Kim, J. Distinct roles of neuronal and microglial CB2 cannabinoid receptors in the mouse hippocampus. Neuroscience 2017, 363, 11–25. [Google Scholar] [CrossRef]
- Rizzo, V.; Carletti, F.; Gambino, G.; Schiera, G.; Cannizzaro, C.; Ferraro, G.; Sardo, P. Role of CB2 receptors and cGMP pathway on the cannabinoid-dependent antiepileptic effects in an in vivo model of partial epilepsy. Epilepsy Res. 2014, 108, 1711–1718. [Google Scholar] [CrossRef]
- Shapiro, L.; Wong, J.C.; Escayg, A. Reduced cannabinoid 2 receptor activity increases susceptibility to induced seizures in mice. Epilepsia 2019, 60, 2359–2369. [Google Scholar] [CrossRef]
- da Silva Oliveira, G.L.; da Silva, J.; Dos Santos, C.L.d.S.A.P.; Feitosa, C.M.; de Castro Almeida, F.R. Anticonvulsant, Anxiolytic and Antidepressant Properties of the beta-caryophyllene in Swiss Mice: Involvement of Benzodiazepine-GABAAergic, Serotonergic and Nitrergic Systems. Curr. Mol. Pharm. 2021, 14, 36–51. [Google Scholar] [CrossRef]
- de Carvalho, C.R.; Hoeller, A.A.; Franco, P.L.; Martini, A.P.; Soares, F.M.; Lin, K.; Prediger, R.D.; Whalley, B.J.; Walz, R. The cannabinoid CB2 receptor-specific agonist AM1241 increases pentylenetetrazole-induced seizure severity in Wistar rats. Epilepsy Res. 2016, 127, 160–167. [Google Scholar] [CrossRef]
- Xia, K.K.; Shen, J.X.; Huang, Z.B.; Song, H.M.; Gao, M.; Chen, D.J.; Zhang, S.J.; Wu, J. Heterogeneity of cannabinoid ligand-induced modulations in intracellular Ca(2+) signals of mouse pancreatic acinar cells in vitro. Acta Pharm. Sin. 2019, 40, 410–417. [Google Scholar] [CrossRef]
- Cifelli, P.; Grace, A.A. Pilocarpine-induced temporal lobe epilepsy in the rat is associated with increased dopamine neuron activity. Int. J. Neuropsychopharmacol. 2012, 15, 957–964. [Google Scholar] [CrossRef] [Green Version]
- Floresco, S.B.; West, A.R.; Ash, B.; Moore, H.; Grace, A.A. Afferent modulation of dopamine neuron firing differentially regulates tonic and phasic dopamine transmission. Nat. Neurosci. 2003, 6, 968–973. [Google Scholar] [CrossRef]
- Ahmadi, M.; Dufour, J.P.; Seifritz, E.; Mirnajafi-Zadeh, J.; Saab, B.J. The PTZ kindling mouse model of epilepsy exhibits exploratory drive deficits and aberrant activity amongst VTA dopamine neurons in both familiar and novel space. Behav. Brain Res. 2017, 330, 1–7. [Google Scholar] [CrossRef]
- Boczek, T.; Yu, Q.; Zhu, Y.; Dodge-Kafka, K.L.; Goldberg, J.L.; Kapiloff, M.S. cAMP at Perinuclear mAKAPα Signalosomes Is Regulated by Local Ca2+ Signaling in Primary Hippocampal Neurons. eNeuro 2021, 8. [Google Scholar] [CrossRef]
- Kjaer, C.; Barzaghi, G.; Bak, L.K.; Goetze, J.P.; Yde, C.W.; Woldbye, D.; Pinborg, L.H.; Jensen, L.J. Transcriptome analysis in patients with temporal lobe epilepsy. Brain 2019, 142, e55. [Google Scholar] [CrossRef] [PubMed]
- Guelfi, S.; Botia, J.A.; Thom, M.; Ramasamy, A.; Perona, M.; Stanyer, L.; Martinian, L.; Trabzuni, D.; Smith, C.; Walker, R.; et al. Transcriptomic and genetic analyses reveal potential causal drivers for intractable partial epilepsy. Brain 2019, 142, 1616–1630. [Google Scholar] [CrossRef]
- Gunaydin, C.; Arslan, G.; Bilge, S.S. Proconvulsant effect of trans-cinnamaldehyde in pentylenetetrazole-induced kindling model of epilepsy: The role of TRPA1 channels. Neurosci. Lett. 2020, 721, 134823. [Google Scholar] [CrossRef]
- Ebrahimi-Ghiri, M.; Khakpai, F. The link between cannabinoids and depression. In The Neuroscience of Depression; Elsevier: Amsterdam, The Netherlands, 2021; pp. 293–300. [Google Scholar]
- Franco, R.; Villa, M.; Morales, P.; Reyes-Resina, I.; Gutierrez-Rodriguez, A.; Jimenez, J.; Jagerovic, N.; Martinez-Orgado, J.; Navarro, G. Increased expression of cannabinoid CB2 and serotonin 5-HT1A heteroreceptor complexes in a model of newborn hypoxic-ischemic brain damage. Neuropharmacology 2019, 152, 58–66. [Google Scholar] [CrossRef]
- Foust, A.J.; Yu, Y.; Popovic, M.; Zecevic, D.; McCormick, D.A. Somatic membrane potential and Kv1 channels control spike repolarization in cortical axon collaterals and presynaptic boutons. J. Neurosci. 2011, 31, 15490–15498. [Google Scholar] [CrossRef] [Green Version]
- Xiao, Y.; Yang, J.; Ji, W.; He, Q.; Mao, L.; Shu, Y. A- and D-type potassium currents regulate axonal action potential repolarization in midbrain dopamine neurons. Neuropharmacology 2021, 185, 108399. [Google Scholar] [CrossRef]
- Kohling, R.; Wolfart, J. Potassium Channels in Epilepsy. Cold Spring Harb. Perspect. Med. 2016, 6, a022871. [Google Scholar]
- Blomster, L.V.; Strobaek, D.; Hougaard, C.; Klein, J.; Pinborg, L.H.; Mikkelsen, J.D.; Christophersen, P. Quantification of the functional expression of the Ca(2+) -activated K(+) channel KCa 3.1 on microglia from adult human neocortical tissue. Glia 2016, 64, 2065–2078. [Google Scholar] [CrossRef] [Green Version]
- Brown, D. M-currents: An update. Trends Neurosci. 1988, 11, 294–299. [Google Scholar] [CrossRef]
- Gunthorpe, M.J.; Large, C.H.; Sankar, R. The mechanism of action of retigabine (ezogabine), a first-in-class K+ channel opener for the treatment of epilepsy. Epilepsia 2012, 53, 412–424. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Wolk, B.; Nolan, R.; Scott, C.E.; Kendall, D.A. Characterization of Subtype Selective Cannabinoid CB2 Receptor Agonists as Potential Anti-Inflammatory Agents. Pharmaceuticals 2021, 14, 378. [Google Scholar] [CrossRef] [PubMed]
- Devinsky, O.; Cross, J.H.; Laux, L.; Marsh, E.; Miller, I.; Nabbout, R.; Scheffer, I.E.; Thiele, E.A.; Wright, S. Cannabidiol in Dravet Syndrome Study, G. Trial of Cannabidiol for Drug-Resistant Seizures in the Dravet Syndrome. N. Engl. J. Med. 2017, 376, 2011–2020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thiele, E.A.; Marsh, E.D.; French, J.A.; Mazurkiewicz-Beldzinska, M.; Benbadis, S.R.; Joshi, C.; Lyons, P.D.; Taylor, A.; Roberts, C.; Sommerville, K.; et al. Cannabidiol in patients with seizures associated with Lennox-Gastaut syndrome (GWPCARE4): A randomised, double-blind, placebo-controlled phase 3 trial. Lancet 2018, 391, 1085–1096. [Google Scholar] [CrossRef]
- Little, J.P.; Villanueva, E.B.; Klegeris, A. Therapeutic potential of cannabinoids in the treatment of neuroinflammation associated with Parkinson’s disease. Mini Rev. Med. Chem. 2011, 11, 582–590. [Google Scholar] [CrossRef] [Green Version]
- Cabral, G.A.; Marciano-Cabral, F. Cannabinoid receptors in microglia of the central nervous system: Immune functional relevance. J. Leukoc. Biol. 2005, 78, 1192–1197. [Google Scholar] [CrossRef] [PubMed]
- Shang, Y.; Tang, Y. The central cannabinoid receptor type-2 (CB2) and chronic pain. Int. J. Neurosci. 2017, 127, 812–823. [Google Scholar] [CrossRef]
- Food, U.; Administration, D. FDA Approves First Drug Comprised of an Active Ingredient Derived from Marijuana to Treat Rare, Severe Forms of Epilepsy; U.S. Food & Drugs: Washington, DC, USA, 2018; p. 25. [Google Scholar]
Figure 1.
Diagram epitomizing the CB2R-associated mechanisms in the modulation of epileptic seizures.
Figure 1.
Diagram epitomizing the CB2R-associated mechanisms in the modulation of epileptic seizures.


{kind=link}
Table 1.
Preclinical evidence for CB2R-mediated modulations in epilepsy.
| Modulation Approaches | Seizure or Epilepsy Model | Response | References |
|---|---|---|---|
| - | PTZ induced seizures in heterozygous and homozygous CB2 knockout mice | Susceptibility | [84] |
| - | Handling-induced and spontaneous seizures | Susceptibility | [81] |
| 2-Arachidonoylglycerol (2-AG) | Kainate induced seizures in CB1R and CB2R double-knockout mice | Suppressed excitatory | [12] |
| CB2 agonist beta caryophyllene | PTZ induced seizures in mice | Anticonvulsant activity | [85] |
| CB2 agonist beta caryophyllene | Induced by kainic acid (KA) seizure in mice | Decreased the seizure scores | [85] |
| CB2 antagonist AM630 | Kainate induced seizures in CB1R knockout mice | Increased seizure susceptibility | [80] |
| CB2 agonist AM1241 | PTZ induced seizures in rats | Increased seizure severity | [86] |
| CB2 agonist HU-308 | DMCM and PTZ induced seizures in rats | No significant effect on seizure severity | [80] |
| CB2R agonist JWH133 | PTZ induced seizures in mice | No significant effect on seizure severity | [84] |
| CB2R antagonist SR144528 | PTZ induced seizures in mice | Increases seizure susceptibility | [84] |
| CB1/2R mixed agonist WIN 55,212-2 | DMCM and PTZ induced seizures in rats | Anticonvulsant effects | [80] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Ji, X.; Zeng, Y.; Wu, J. The CB2 Receptor as a Novel Therapeutic Target for Epilepsy Treatment. Int. J. Mol. Sci. 2021, 22, 8961. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22168961
AMA Style
Ji X, Zeng Y, Wu J. The CB2 Receptor as a Novel Therapeutic Target for Epilepsy Treatment. International Journal of Molecular Sciences. 2021; 22(16):8961. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22168961
Chicago/Turabian StyleJi, Xiaoyu, Yang Zeng, and Jie Wu. 2021. "The CB2 Receptor as a Novel Therapeutic Target for Epilepsy Treatment" International Journal of Molecular Sciences 22, no. 16: 8961. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22168961
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.