
circRNA-Mediated Inhibin–Activin Balance Regulation in Ovarian Granulosa Cell Apoptosis and Follicular Atresia

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
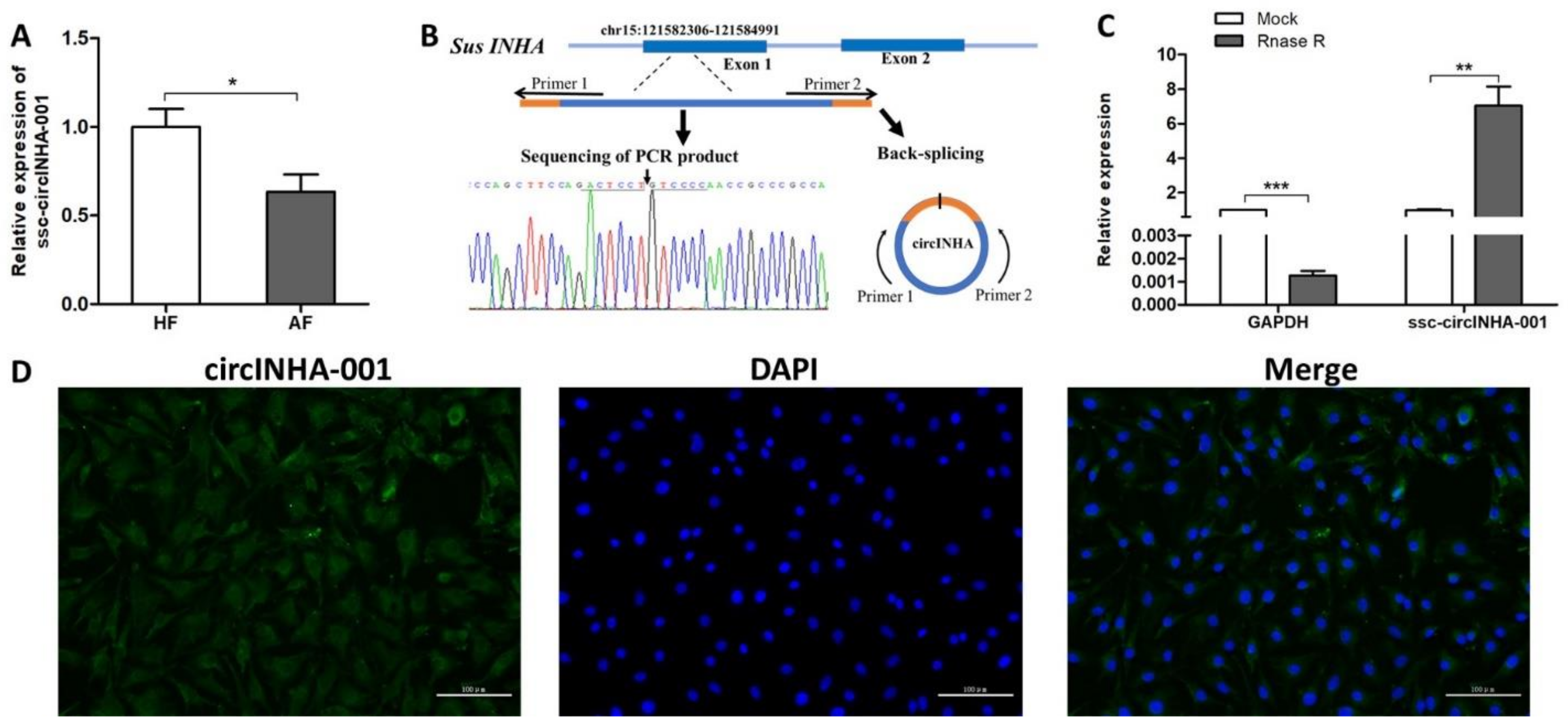
2.1. Identification and Validation of ssc-circINHA-001
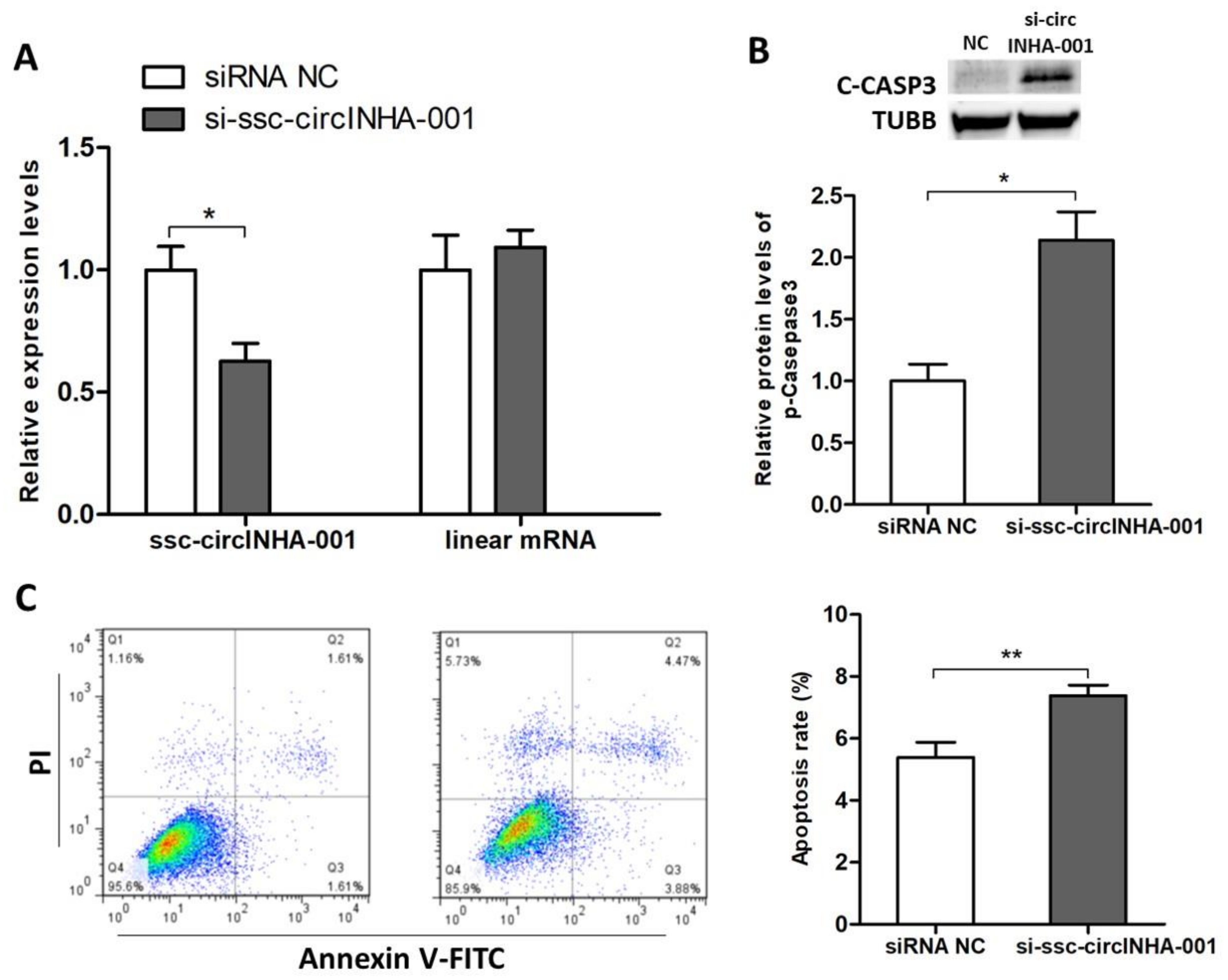
2.2. ssc-circINHA-001 Inhibits GC Apoptosis
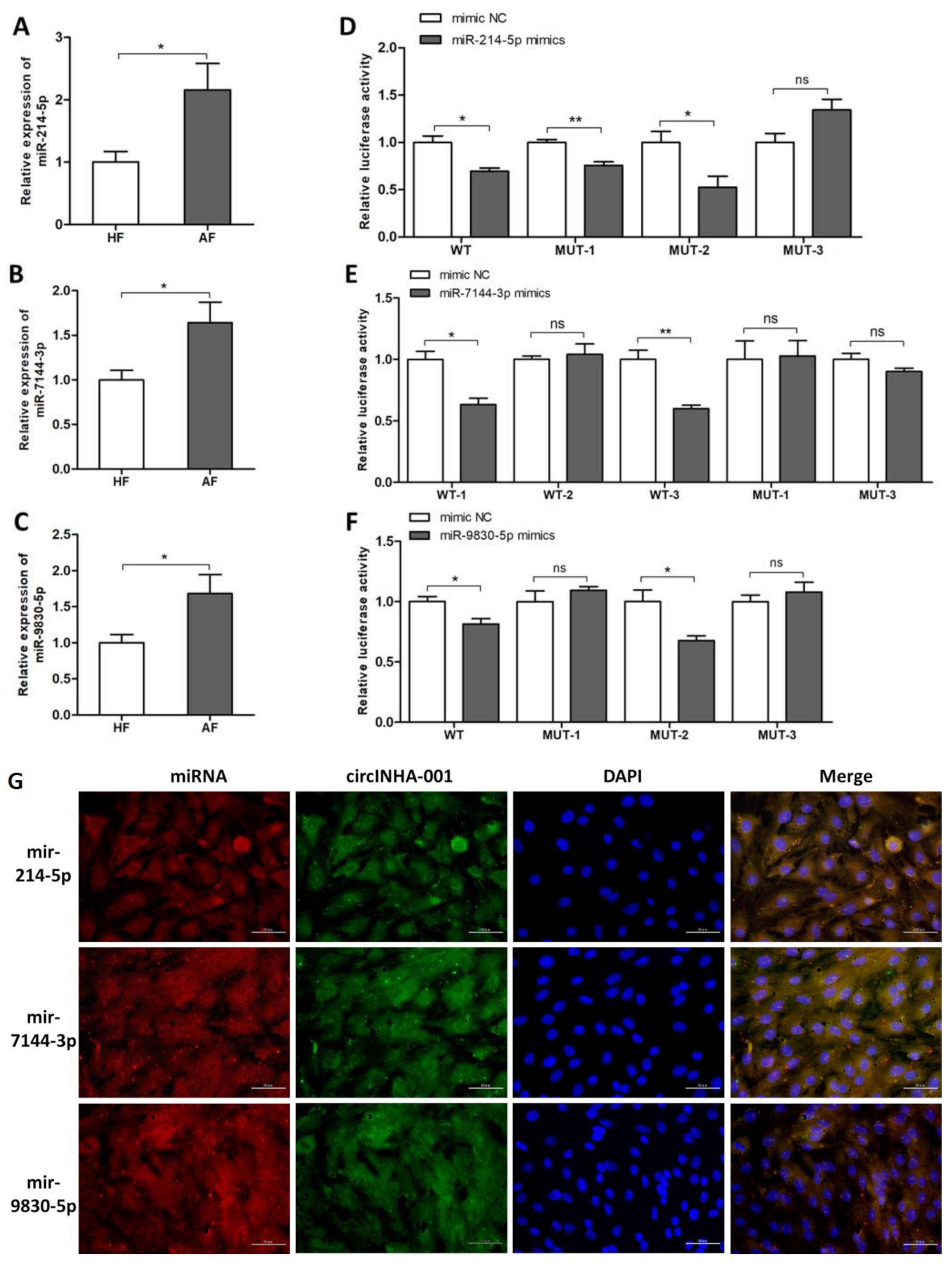
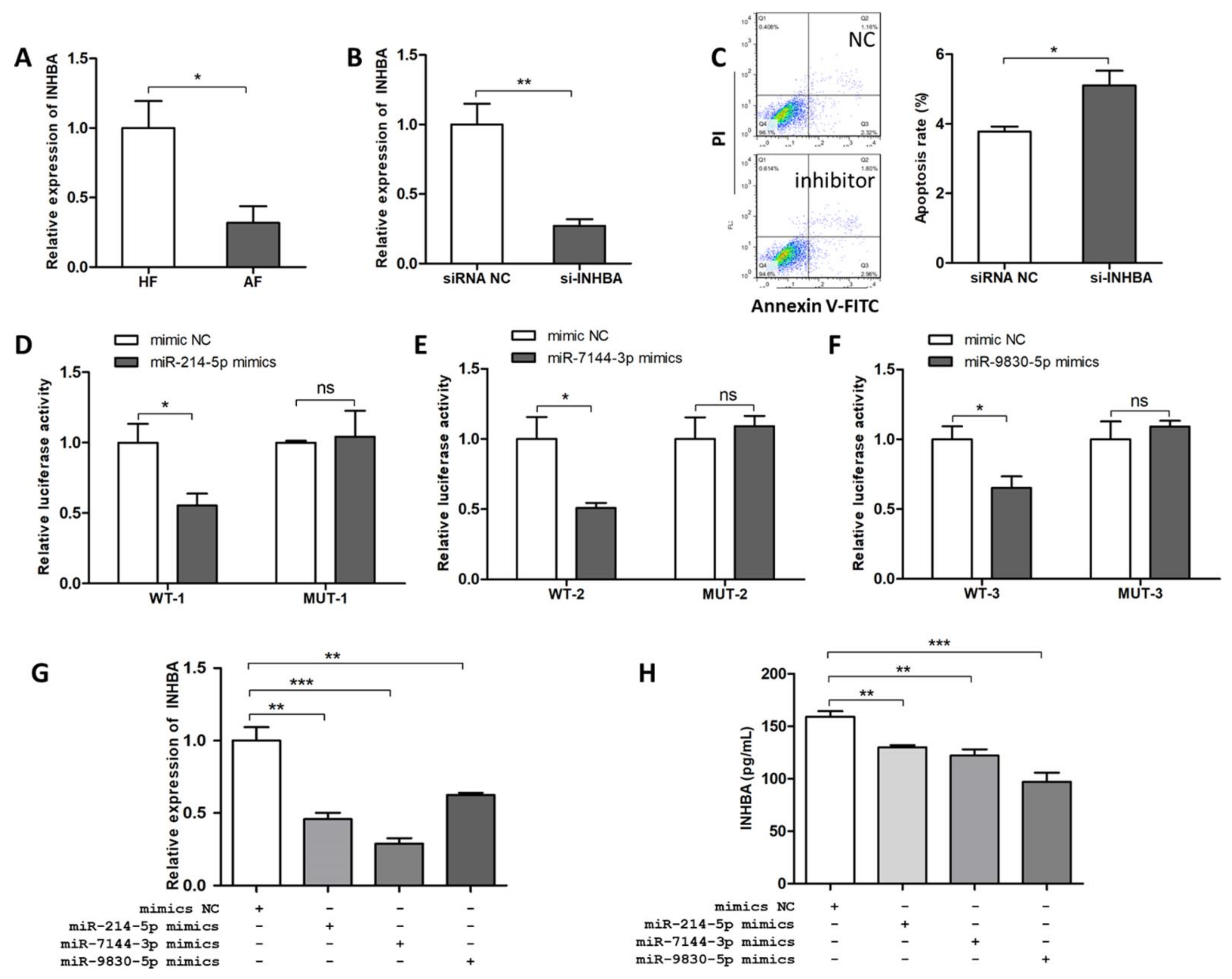
2.3. ssc-circINHA-001 is a Common Target of miR-214-5p/miR-7144-3p/miR-9830-5p
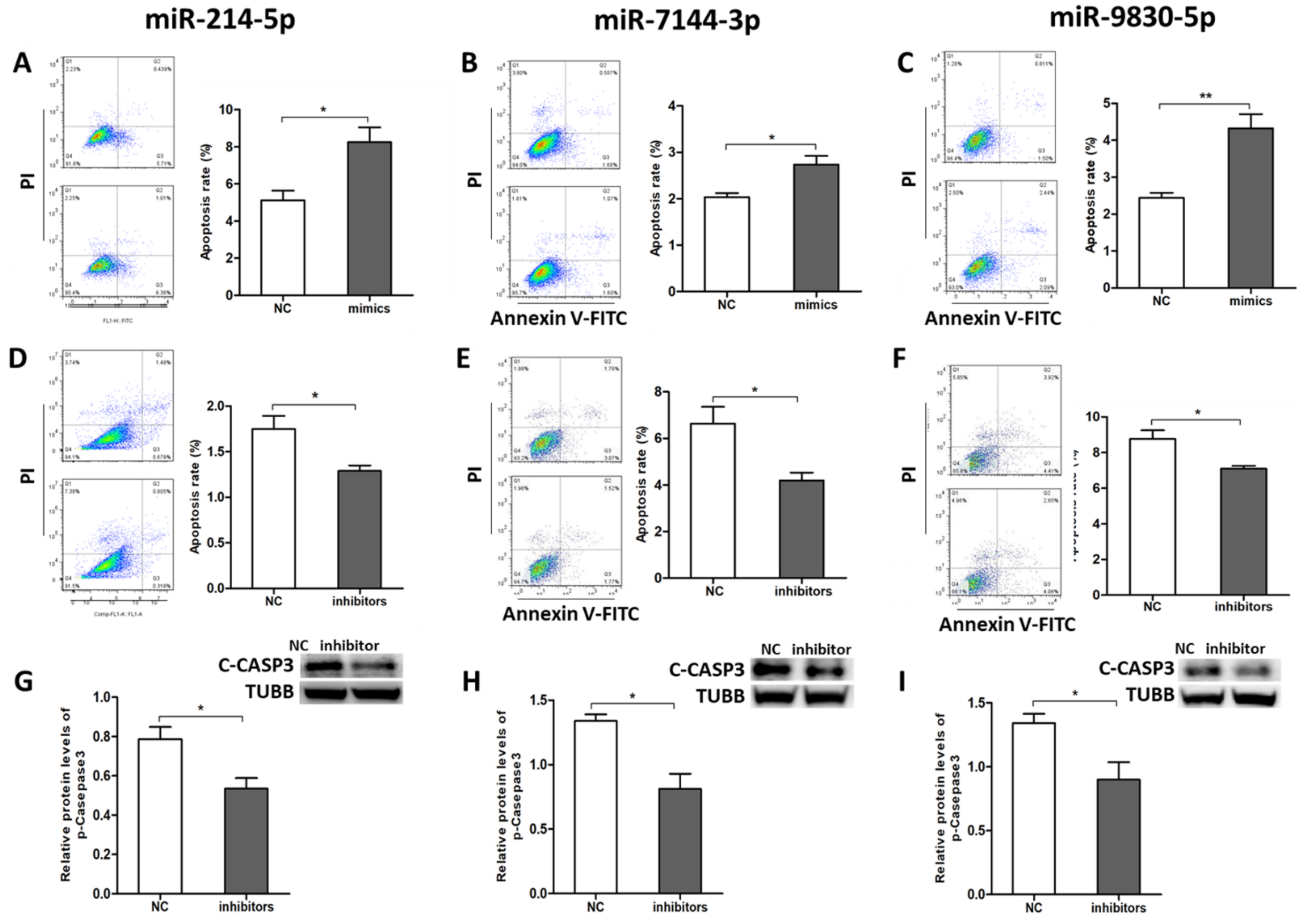
2.4. miR-214-5p/miR-7144-3p/miR-9830-5p Enhanced GC Apoptosis
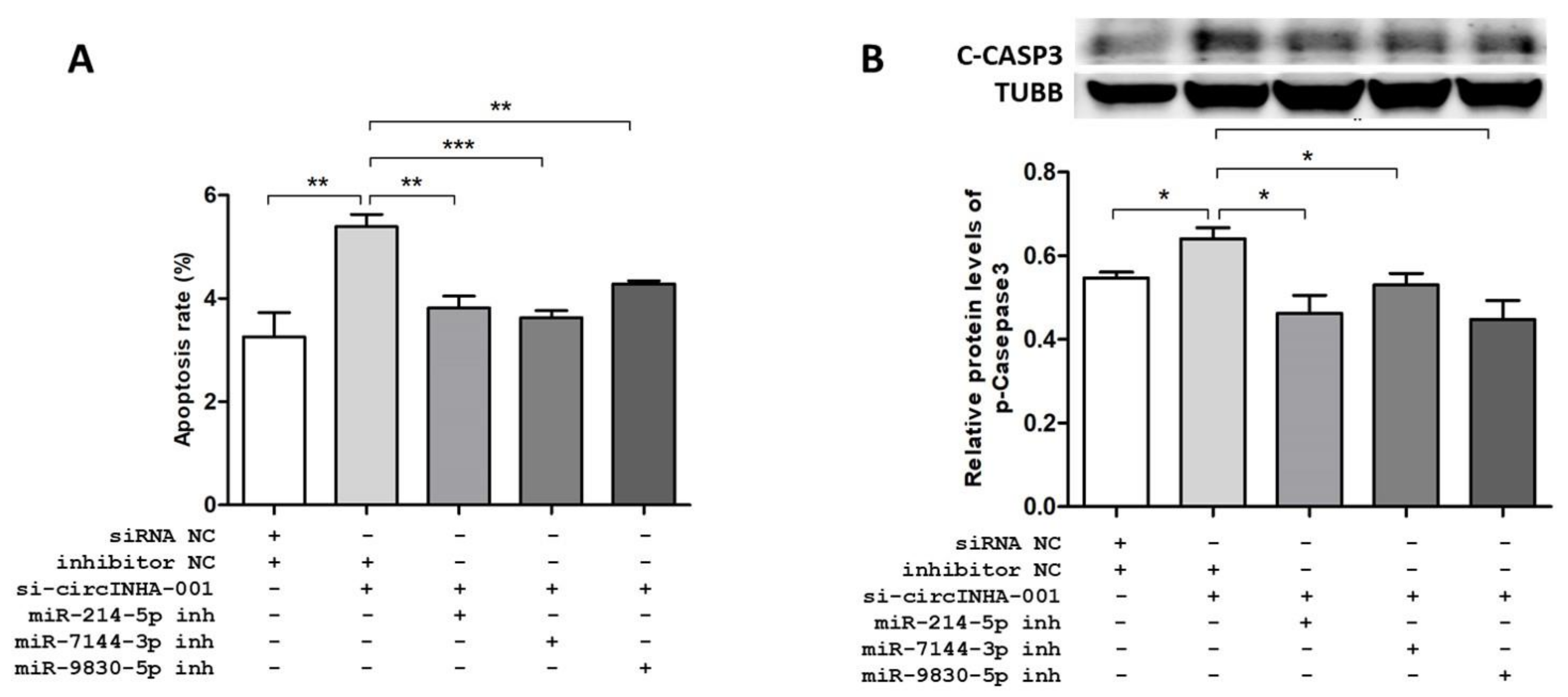
2.5. ssc-circINHA-001 Affecst GC Apoptosis through miR-214-5p/miR-7144-3p/miR-9830-5p
2.6. INHBA Regulates Follicular Atresia and GC Apoptosis and Is Targeted by miR-214-5p/miR-7144-3p/miR-9830-5p
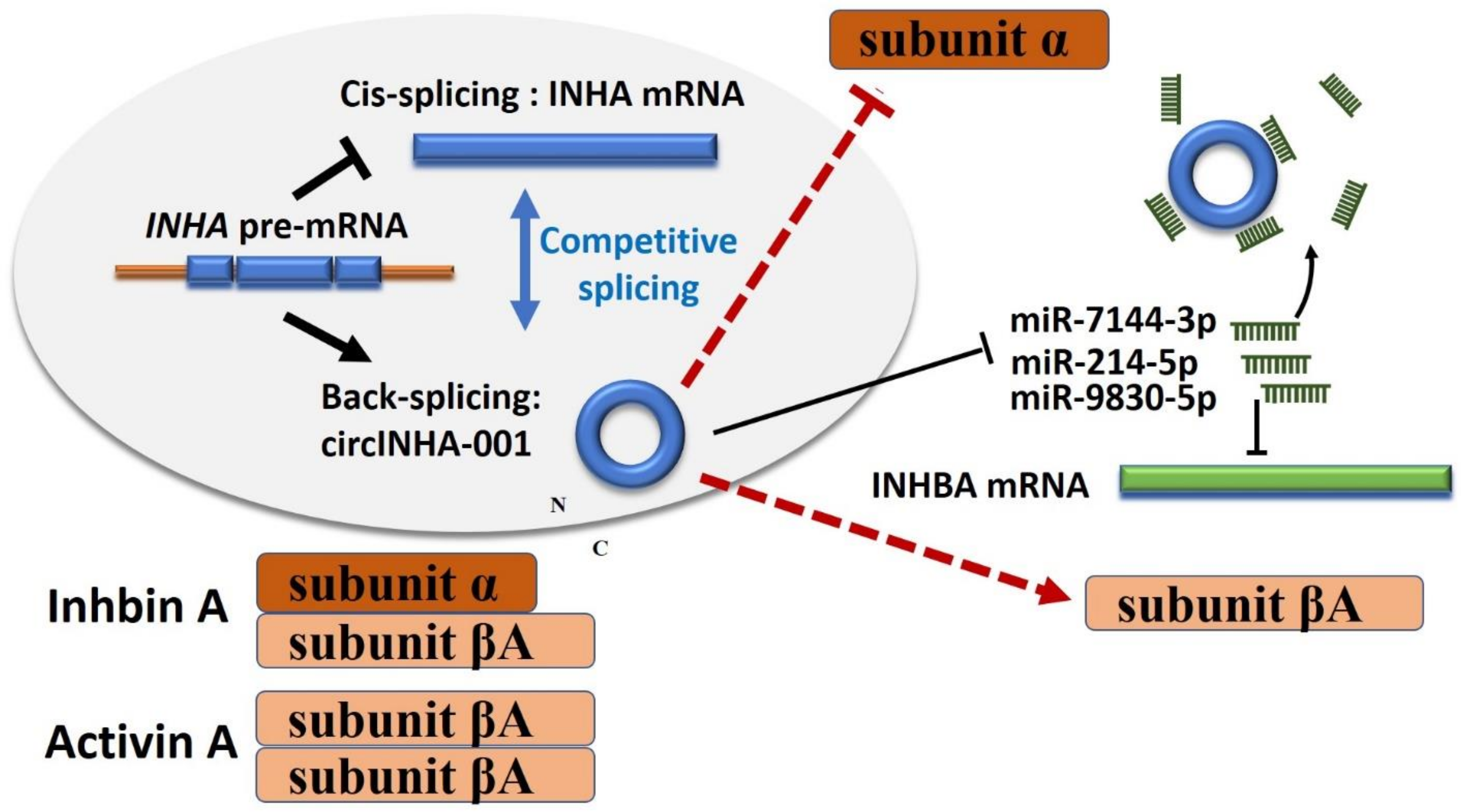
3. Discussion
4. Materials and Methods
4.1. Follicle Collection and Isolation
4.2. Cell Culture and Transfection
4.3. RNA Preparation and qRT-PCR
4.4. RNase R Treatment and RT-PCR Product Sequencing
4.5. Dual Luciferase Activity Assay
4.6. Western Blotting
4.7. FISH
4.8. Apoptosis Assay
4.9. ELISA Assay
4.10. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
References
- Orisaka, M.; Tajima, K.; Tsang, B.K.; Kotsuji, F. Oocyte-granulosa-theca cell interactions during preantral follicular development. J. Ovarian Res. 2009, 2, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Tao, Q.; Shang, J.; Xu, Y.; Zhang, L.; Ma, Y.; Zhu, W.; Yang, M.; Ding, Y.; Yin, Z. MiR-26a promotes apoptosis of porcine granulosa cells by targeting the 3β-hydroxysteroid-Δ24-reductase gene. Asian-Australas. J. Anim. Sci. 2020, 33, 547–555. [Google Scholar] [CrossRef] [PubMed]
- Johnson, P.A.; Woodcock, J.R.; Kent, T.R. Effect of activin A and inhibin A on expression of the inhibin/activin beta-B-subunit and gonadotropin receptors in granulosa cells of the hen. Gen. Comp. Endocrinol. 2006, 147, 102–107. [Google Scholar] [CrossRef] [PubMed]
- Clifford, R.L.; Singer, C.A.; John, A.E. Epigenetics and miRNA emerge as key regulators of smooth muscle cell phenotype and function. Pulm. Pharmacol. Ther. 2013, 26, 75–85. [Google Scholar] [CrossRef] [Green Version]
- Lin, F.; Li, R.; Pan, Z.X.; Zhou, B.; Yu, D.B.; Wang, X.G.; Ma, X.S.; Han, J.; Shen, M.; Liu, H.L. miR-26b promotes granulosa cell apoptosis by targeting ATM during follicular atresia in porcine ovary. PLoS ONE 2012, 7, e38640. [Google Scholar] [CrossRef] [Green Version]
- Nicolet, B.P.; Engels, S.; Aglialoro, F.; van den Akker, E.; von Lindern, M.; Wolkers, M.C. Circular RNA expression in human hematopoietic cells is widespread and cell-type specific. Nucleic Acids Res. 2018, 46, 8168–8180. [Google Scholar] [CrossRef]
- Memczak, S.; Jens, M.; Elefsinioti, A.; Torti, F.; Krueger, J.; Rybak, A.; Maier, L.; Mackowiak, S.D.; Gregersen, L.H.; Munschauer, M.; et al. Circular RNAs are a large class of animal RNAs with regulatory potency. Nature 2013, 495, 333–338. [Google Scholar] [CrossRef]
- Legnini, I.; Di Timoteo, G.; Rossi, F.; Morlando, M.; Briganti, F.; Sthandier, O.; Fatica, A.; Santini, T.; Andronache, A.; Wade, M.; et al. Circ-ZNF609 is a circular RNA that can be translated and functions in myogenesis. Mol. Cell 2017, 66, 22–37.e9. [Google Scholar] [CrossRef] [Green Version]
- Guo, T.Y.; Huang, L.; Yao, W.; Du, X.; Li, Q.Q.; Ma, M.L.; Li, Q.F.; Liu, H.L.; Zhang, J.B.; Pan, Z.X. The potential biological functions of circular RNAs during the initiation of atresia in pig follicles. Domest. Anim. Endocrinol. 2019, 72, 106401. [Google Scholar] [CrossRef]
- Kertesz, M.; Iovino, N.; Unnerstall, U.; Gaul, U.; Segal, E. The role of site accessibility in microRNA target recognition. Nat. Genet. 2007, 39, 1278–1284. [Google Scholar] [CrossRef]
- Enright, A.J.; John, B.; Gaul, U.; Tuschl, T.; Marks, D.S. MicroRNA targets in drosophila. Genome Biol. 2003, 4, 1–27. [Google Scholar] [CrossRef] [Green Version]
- Sturm, M.; Hackenberg, M.; Langenberger, D.; Frishman, D. TargetSpy: A supervised machine learning approach for microRNA target prediction. BMC Bioinform. 2010, 11, 292. [Google Scholar] [CrossRef] [Green Version]
- Cocquerelle, C.; Mascrez, B.; Hétuin, D.; Bailleul, B. Mis-splicing yields circular RNA molecules. FASEB J. 1993, 7, 155–160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caiment, F.; Gaj, S.; Claessen, S.; Kleinjans, J. High-throughput data integration of RNA-miRNA-circRNA reveals novel insights into mechanisms of benzo[a]pyrene-induced carcinogenicity. Nucleic Acids Res. 2015, 43, 2525–2534. [Google Scholar] [CrossRef] [Green Version]
- Stella, M.; Falzone, L.; Caponnetto, A.; Gattuso, G.; Barbagallo, C.; Battaglia, R.; Mirabella, F.; Broggi, G.; Altieri, R.; Certo, F.; et al. Serum extracellular vesicle-derived circHIPK3 and circSMARCA5 are two novel diagnostic biomarkers for glioblastoma multiforme. Pharmaceuticals 2021, 14, 618. [Google Scholar] [CrossRef]
- van Zonneveld, A.J.; Kölling, M.; Bijkerk, R.; Lorenzen, J.M. Circular RNAs in kidney disease and cancer. Nat. Rev. Nephrol. 2021. [Google Scholar] [CrossRef]
- Li, J.; Sun, C.; Cui, H.; Sun, J.; Zhou, P. Role of circRNAs in neurodevelopment and neurodegenerative diseases. J. Mol. Neurosci. 2021. [Google Scholar] [CrossRef]
- Wang, K.; Gao, X.Q.; Wang, T.; Zhou, L.Y. The function and therapeutic potential of circular RNA in cardiovascular diseases. Cardiovasc. Drugs Ther. 2021. [Google Scholar] [CrossRef]
- Zaiou, M. circRNAs signature as potential diagnostic and prognostic biomarker for diabetes mellitus and related cardiovascular complications. Cells 2020, 9, 659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, H.; Li, Y.; Niringiyumukiza, J.D.; Su, P.; Xiang, W. Circular RNA involvement in aging: An emerging player with great potential. Mech. Ageing Dev. 2019, 178, 16–24. [Google Scholar] [CrossRef] [PubMed]
- Che, Q.; Liu, M.; Xu, J.; Liu, Y.; Cao, X.; Dong, X.; Liu, S. Characterization of circular RNA expression profiles in cumulus cells from patients with polycystic ovary syndrome. Fertil. Steril. 2019, 111, 1243–1251.e1. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Peng, X.; Lv, X.; Liu, L.; Li, X.; He, X.; Lv, F.; Pan, Y.; Wang, L.; Liu, K. High throughput circRNAs sequencing profile of follicle fluid exosomes of polycystic ovary syndrome patients. J. Cell. Physiol. 2019, 234, 15537–15547. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, I.; Karedath, T.; Andrews, S.S.; Al, I.K.; Mohamoud, Y.A.; Querleu, D.; Rafii, A.; Malek, J.A. Altered expression pattern of circular RNAs in primary and metastatic sites of epithelial ovarian carcinoma. Oncotarget 2016, 7, 36366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ning, L.; Long, B.; Zhang, W.; Yu, M.; Wang, S.; Cao, D.; Yang, J.; Shen, K.; Huang, Y.; Lang, J. Circular RNA profiling reveals circEXOC6B and circN4BP2L2 as novel prognostic biomarkers in epithelial ovarian cancer. Int. J. Oncol. 2018, 53, 2637–2646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, G.; Yan, J.; Guo, J.; Tang, Z. Identification of ovarian circular RNAs and differential expression analysis between MeiShan and large white pigs. Animals 2020, 10, 1114. [Google Scholar] [CrossRef] [PubMed]
- Pan, X.; Gong, W.; He, Y.; Li, N.; Zhang, H.; Zhang, Z.; Li, J.; Yuan, X. Ovary-derived circular RNAs profile analysis during the onset of puberty in gilts. BMC Genom. 2021, 22, 445. [Google Scholar] [CrossRef]
- Guo, T.; Zhang, J.; Yao, W.; Du, X.; Li, Q.; Huang, L.; Ma, M.; Li, Q.; Liu, H.; Pan, Z. CircINHA resists granulosa cell apoptosis by upregulating CTGF as a ceRNA of miR-10a-5p in pig ovarian follicles. Biochim. Biophys. Acta (BBA)-Gene Regul. Mech. 2019, 1862, 194420. [Google Scholar] [CrossRef] [PubMed]
- Guan, X.; Zong, Z.; Liu, Y.; Chen, S.; Wang, L.; Zhao, Y. circPUM1 promotes tumorigenesis and progression of ovarian cancer by sponging miR-615-5p and miR-6753-5p. Mol. Ther.-Nucleic Acids 2019, 18, 882–892. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y. Selection of Genes Related to Bovine Follicular Development and Study on the Functions of SIRT1 and INHBA Gene. Ph.D. Dissertation, Jilin Agricultural University, Changchun, China, 2018. [Google Scholar]
- Cui, Z.; Liu, L.; Zhu, Q.; Wang, Y.; Zhao, X. Inhibin A regulates follicular development via hormone secretion and granulosa cell behaviors in laying hens. Cell Tissue Res. 2020, 381, 337–350. [Google Scholar] [CrossRef] [PubMed]
- Ma, M.; Zhang, J.; Gao, X.; Yao, W.; Li, Q.; Pan, Z. miR-361-5p mediates SMAD4 to promote porcine granulosa cell apoptosis through VEGFA. Biomolecules 2020, 10, 1281. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Liu, Y.; Yao, W.; Li, Q.; Liu, H.; Pan, Z. Initiation of follicular atresia: Gene networks during early atresia in pig ovaries. Reproduction 2018, 156, 23–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Yao, W.; Yao, Y.; Du, X.; Zhou, J.; Ma, B.; Liu, H.; Li, Q.; Pan, Z. MiR-92a inhibits porcine ovarian granulosa cell apoptosis by targeting Smad7 gene. FEBS Lett. 2014, 588, 4497–4503. [Google Scholar] [CrossRef] [PubMed] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, M.; Wang, H.; Zhang, Y.; Zhang, J.; Liu, J.; Pan, Z. circRNA-Mediated Inhibin–Activin Balance Regulation in Ovarian Granulosa Cell Apoptosis and Follicular Atresia. Int. J. Mol. Sci. 2021, 22, 9113. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22179113
Ma M, Wang H, Zhang Y, Zhang J, Liu J, Pan Z. circRNA-Mediated Inhibin–Activin Balance Regulation in Ovarian Granulosa Cell Apoptosis and Follicular Atresia. International Journal of Molecular Sciences. 2021; 22(17):9113. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22179113
Chicago/Turabian StyleMa, Mengnan, Huiming Wang, Yi Zhang, Jinbi Zhang, Jingge Liu, and Zengxiang Pan. 2021. "circRNA-Mediated Inhibin–Activin Balance Regulation in Ovarian Granulosa Cell Apoptosis and Follicular Atresia" International Journal of Molecular Sciences 22, no. 17: 9113. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22179113