
Allicin May Promote Reversal of T-Cell Dysfunction in Periodontitis via the PD-1 Pathway

 ,
,  ,
,  , , and
, , and
Abstract
:1. Introduction
2. Results
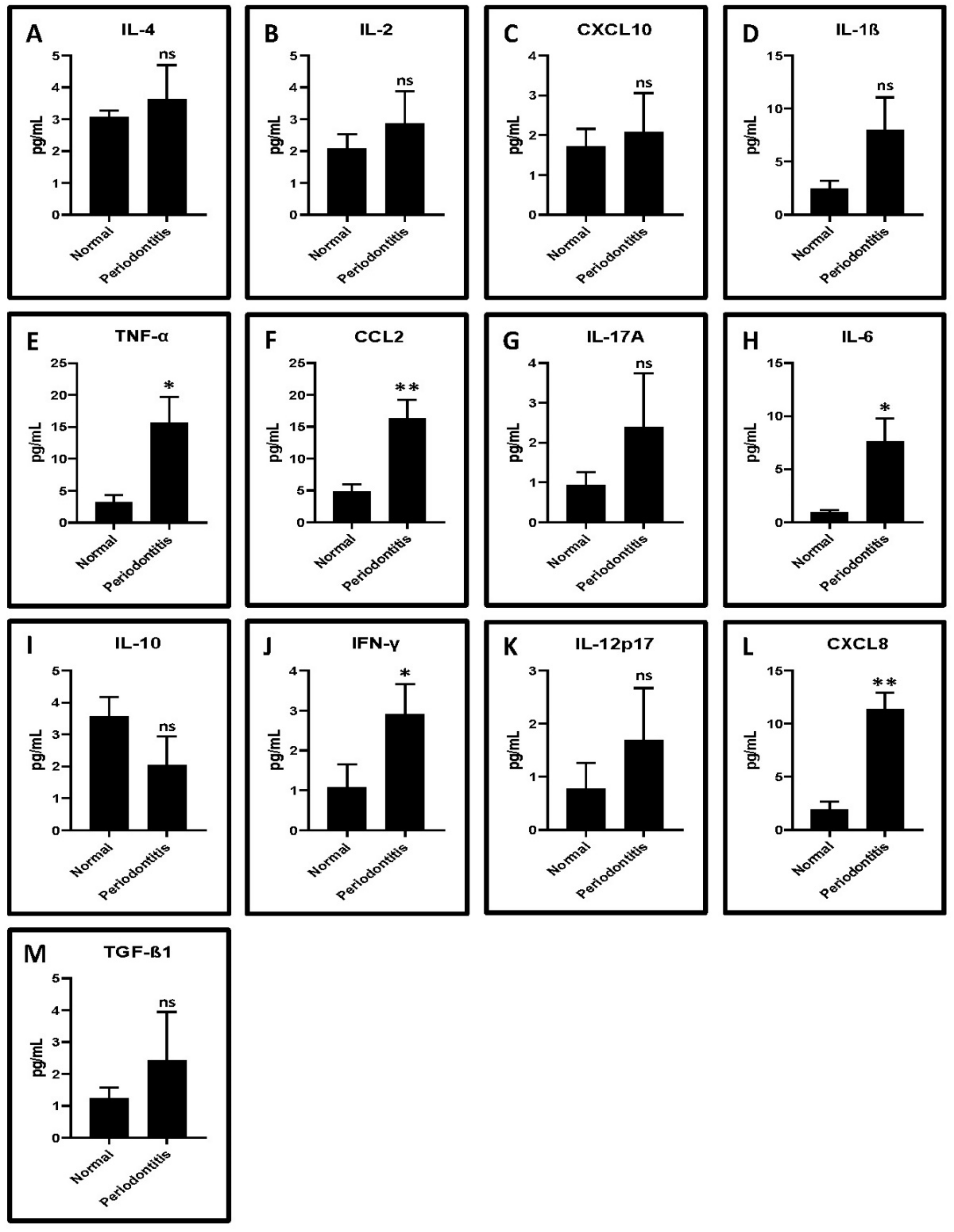
2.1. GCF from Periodontitis Patients Show Higher Levels of Proinflammatory Cytokines Compared to Healthy Controls
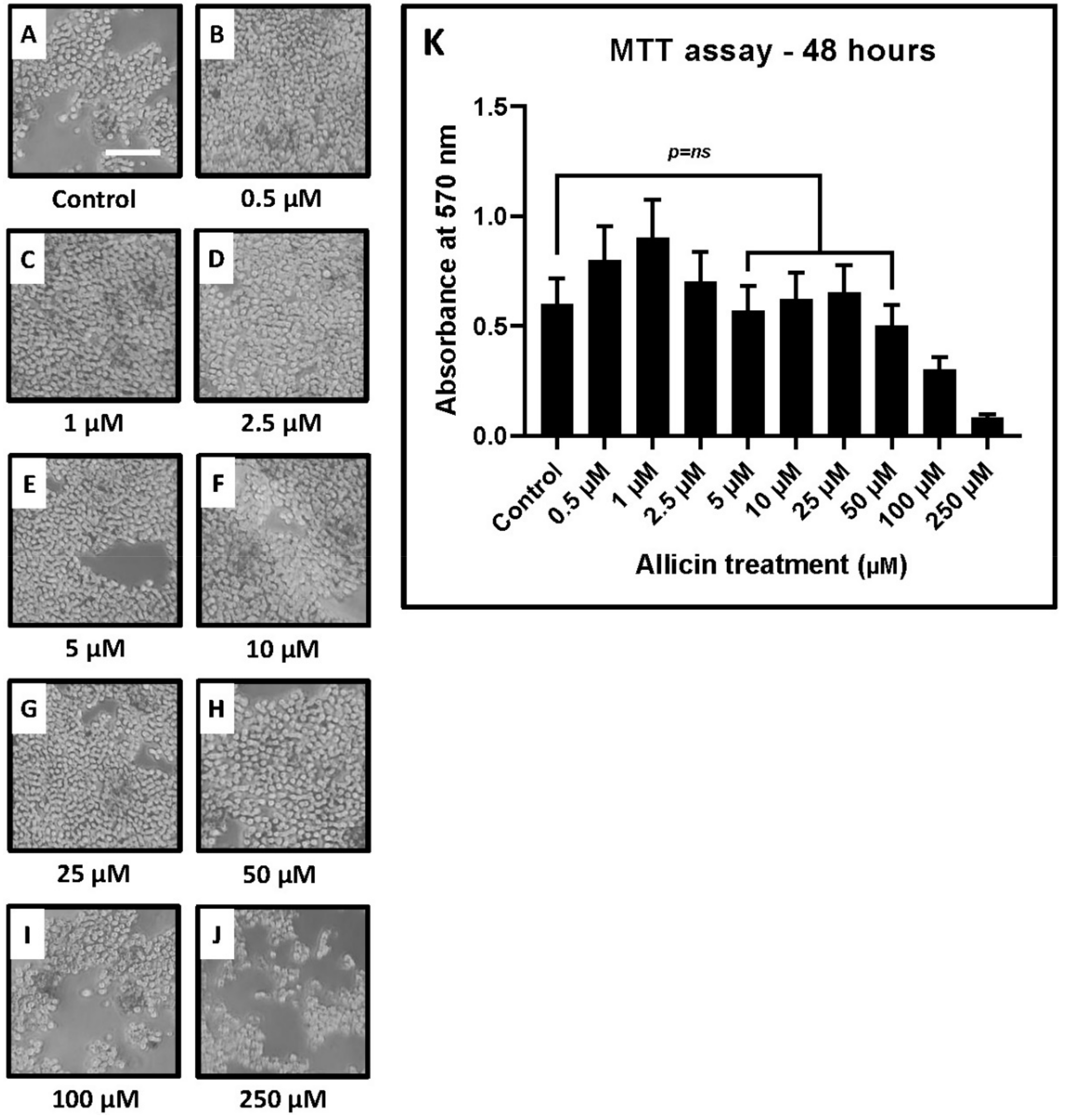
2.2. MTT Assay for Evaluation of Cytotoxic Concentrations of Allicin
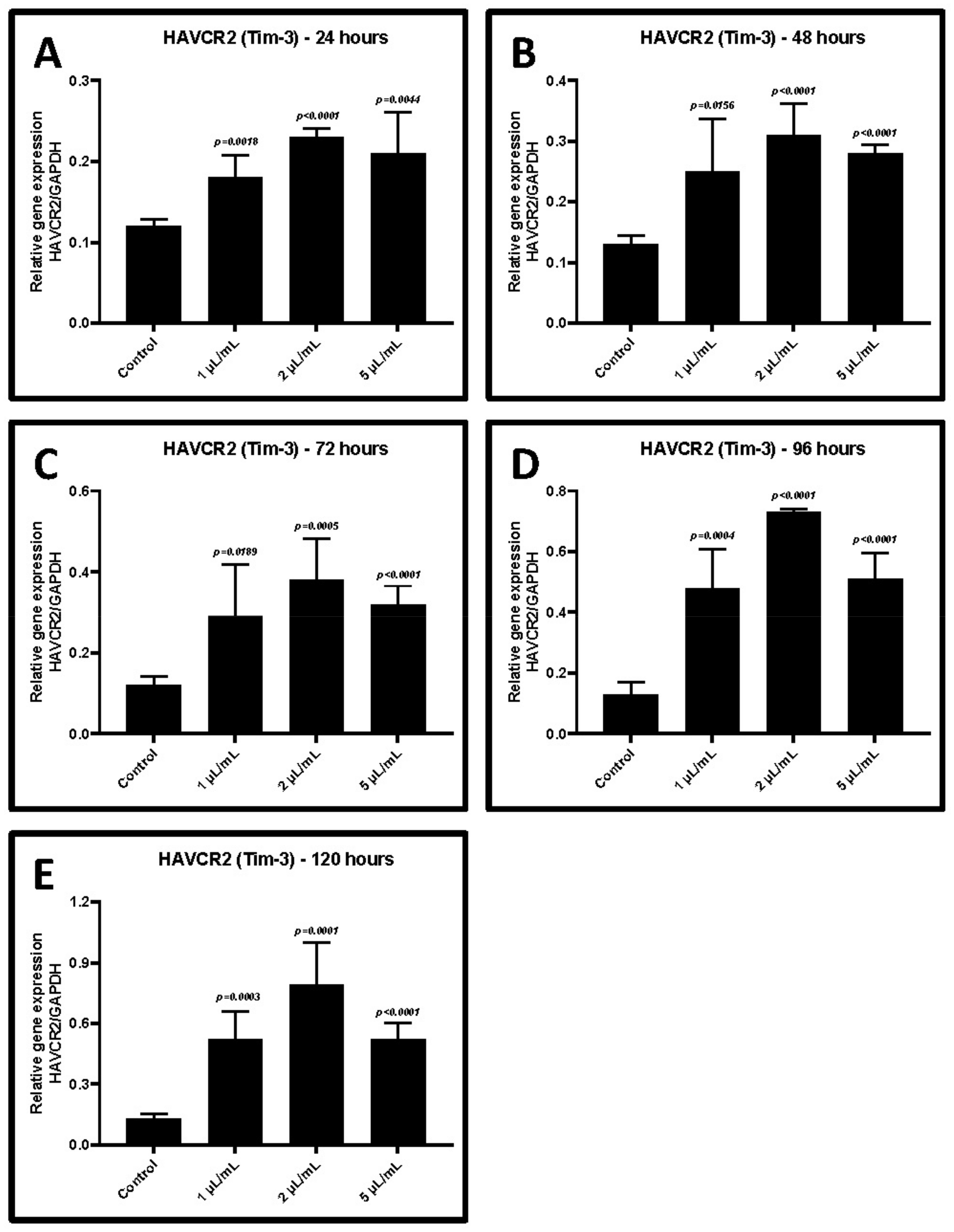
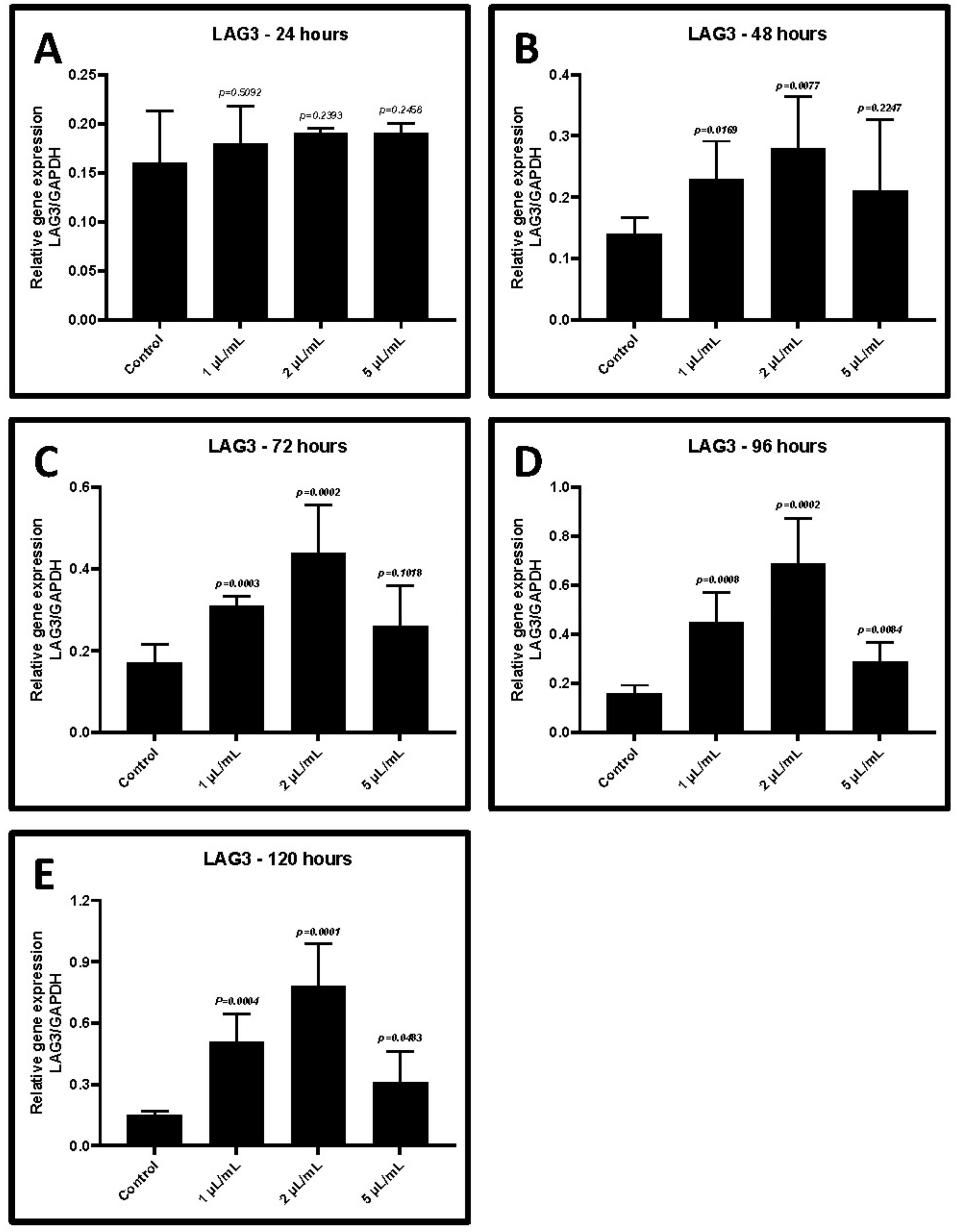
2.3. Treatment of CD3+ T Cells with GCF from Periodontitis Increases the Gene Expression of TIM-3 and LAG-3
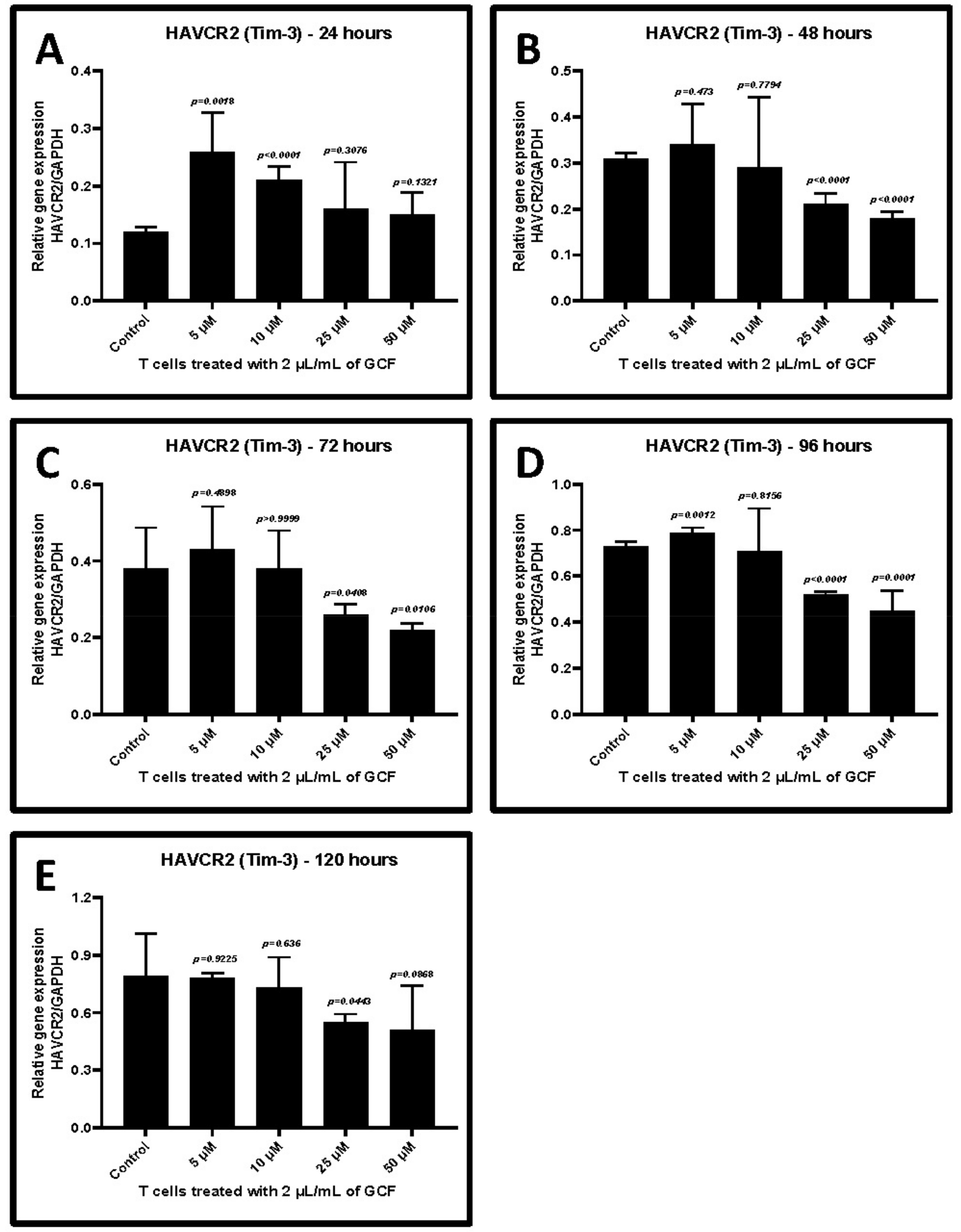
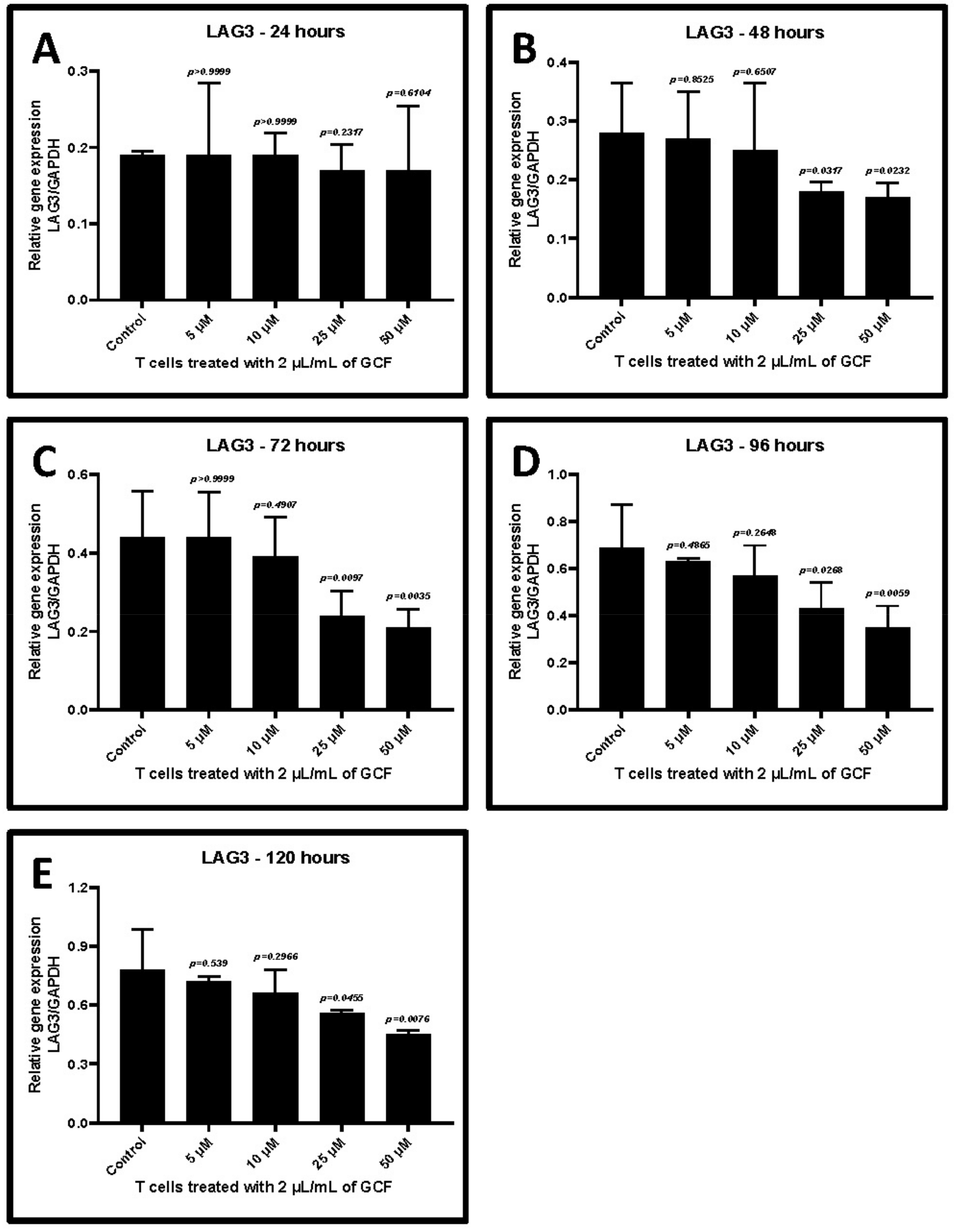
2.4. Allicin Treatment Decreased GCF-Induced Upregulation of TIM-3 and LAG-3
3. Discussion
4. Materials and Methods
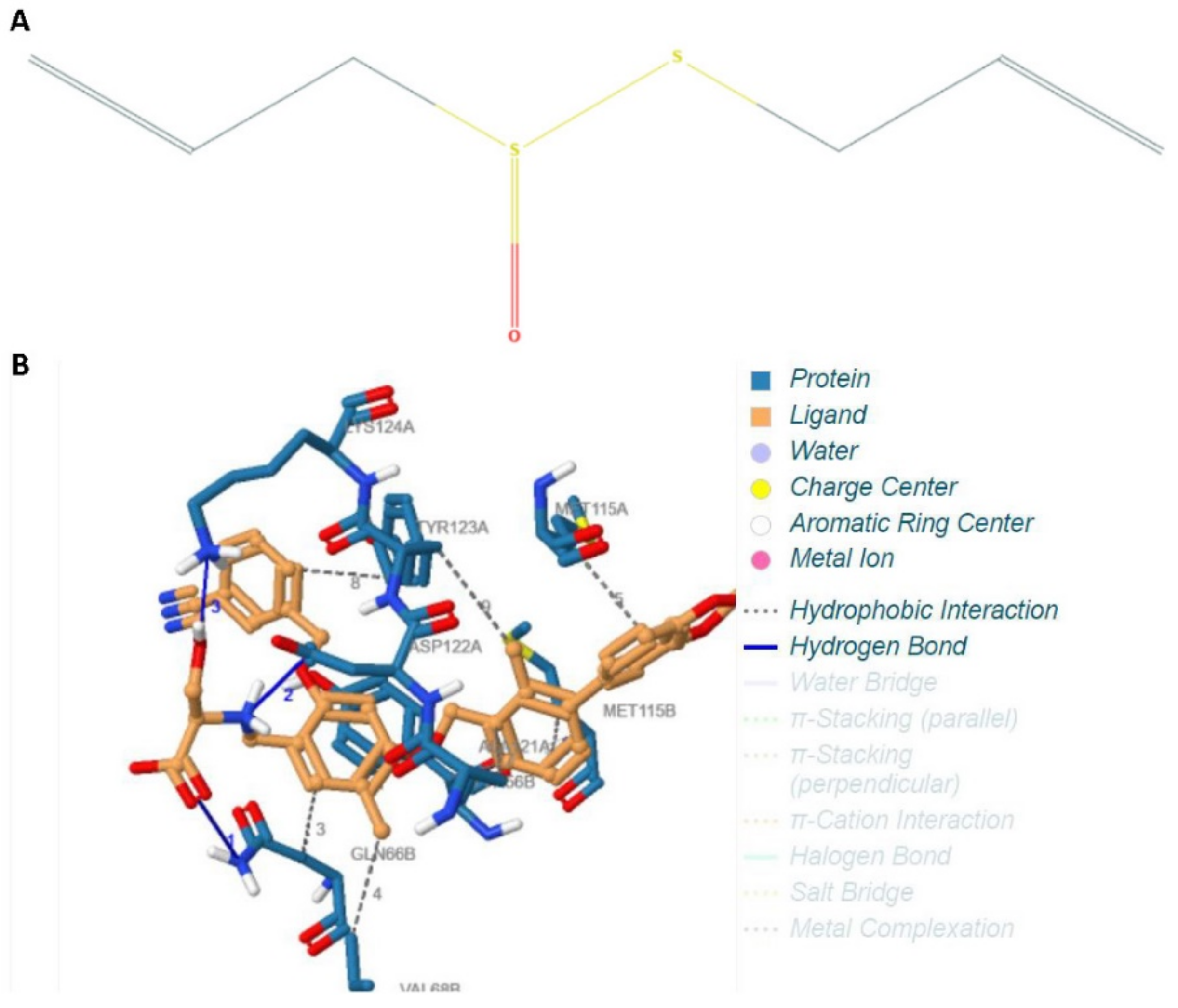
4.1. Protocol and Methodology of the In Silico Experiment
4.2. Patient Selection
4.3. Gingival Crevicular Fluid and Blood Sample Collection
4.4. Cytometric Bead Array for the Estimation of Cytokine Levels in the GCF Samples
4.5. Magnetic Cell Sorting of CD3+ T Cells and T-Cell Culture
4.6. MTT Assay for Analysis of Allicin Cytotoxicity
4.7. Treatment of T Cells with GCF and Allicin
4.8. RT-PCR for Gene Expression Analysis
4.9. Analysis of the Data and Statistics
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Armitage, G.C.; Cullinan, M.P. Comparison of the clinical features of chronic and aggressive periodontitis. Periodontol. 2000 2010, 53, 12–27. [Google Scholar] [CrossRef] [PubMed]
- Grossi, S.G.; Genco, R.J. Periodontal disease and diabetes mellitus: A two-way relationship. Ann. Periodontol. 1998, 3, 51–61. [Google Scholar] [CrossRef] [PubMed]
- Scannapieco, F.A. Position paper of The American Academy of Periodontology: Periodontal disease as a potential risk factor for systemic diseases. J. Periodontol. 1998, 69, 841–850. [Google Scholar] [PubMed]
- Offenbacher, S.; Katz, V.; Fertik, G.; Collins, J.; Boyd, D.; Maynor, G.; McKaig, R.; Beck, J. Periodontal Infection as a Possible Risk Factor for Preterm Low Birth Weight. J. Periodontol. 1996, 67, 1103–1113. [Google Scholar] [CrossRef]
- Ryder, M.I. Comparison of neutrophil functions in aggressive and chronic periodontitis. Periodontol. 2000 2010, 53, 124–137. [Google Scholar] [CrossRef]
- Moretta, L.; Mingari, M.C.; Moretta, A. Human T cell subpopulations in normal and pathologic conditions. Immunol. Rev. 1979, 45, 163–193. [Google Scholar] [CrossRef]
- Petit, M.D.A.; Hovenkamp, E.; Hamann, D.; Roos, M.T.L.; Van Der Velden, U.; Miedema, F.; Loos, B.G. Phenotypical and functional analysis of T cells in periodontitis. J. Periodontal. Res. 2001, 36, 214–220. [Google Scholar] [CrossRef]
- Vernal, R.; Dutzan, N.; Chaparro, A.; Puente, J.; Antonieta Valenzuela, M.; Gamonal, J. Levels of interleukin-17 in gingival crevicular fluid and in supernatants of cellular cultures of gingival tissue from patients with chronic periodontitis. J. Clin. Periodontol. 2005, 32, 383–389. [Google Scholar] [CrossRef]
- Cochran, D.L. Inflammation and Bone Loss in Periodontal Disease. J. Periodontol. 2008, 79, 1569–1576. [Google Scholar] [CrossRef]
- Yi, J.S.; Cox, M.A.; Zajac, A.J. T-cell exhaustion: Characteristics, causes and conversion. Immunology 2010, 129, 474–481. [Google Scholar] [CrossRef]
- Zajac, A.J.; Blattman, J.N.; Murali-Krishna, K.; Sourdive, D.J.; Suresh, M.; Altman, J.D.; Ahmed, R. Viral immune evasion due to persistence of activated T cells without effector function. J. Exp. Med. 1998, 188, 2205–2213. [Google Scholar] [CrossRef]
- Jin, H.-T.; Jeong, Y.H.; Park, H.J.; Ha, S.-J. Mechanism of T cell exhaustion in a chronic environment. BMB Rep. 2011, 44, 217–231. [Google Scholar] [CrossRef] [Green Version]
- Barber, D.L.; Wherry, E.J.; Masopust, D.; Zhu, B.; Allison, J.P.; Sharpe, A.H.; Freeman, G.J.; Ahmed, R. Restoring function in exhausted CD8 T cells during chronic viral infection. Nature 2006, 439, 682–687. [Google Scholar] [CrossRef]
- Lu, D.; Ni, Z.; Liu, X.; Feng, S.; Dong, X.; Shi, X.; Zhai, J.; Mai, S.; Jiang, J.; Wang, Z.; et al. Beyond T Cells: Understanding the Role of PD-1/PD-L1 in Tumor-Associated Macrophages. J. Immunol. Res. 2019, 2019, 1919082. [Google Scholar] [CrossRef] [Green Version]
- Blackburn, S.D.; Shin, H.; Haining, W.N.; Zou, T.; Workman, C.J.; Polley, A.; Betts, M.R.; Freeman, G.J.; Vignali, D.A.A.; Wherry, E.J. Coregulation of CD8+ T cell exhaustion by multiple inhibitory receptors during chronic viral infection. Nat. Immunol. 2009, 10, 29–37. [Google Scholar] [CrossRef] [Green Version]
- Groeger, S.; Domann, E.; Gonzales, J.R.; Chakraborty, T.; Meyle, J. B7-H1 and B7-DC receptors of oral squamous carcinoma cells are upregulated by Porphyromonas gingivalis. Immunobiology 2011, 216, 1302–1310. [Google Scholar] [CrossRef]
- Wang, H.-S.; Yang, F.-H.; Li, Y.; Pei, F.; Kulkarni, A.B.; Chen, Z.; Zhang, L. The expression of PD-1 and LAG-3 in periapical lesions. Am. J. Transl. Res. 2018, 10, 2677–2684. [Google Scholar]
- Zhu, D.; Liu, F.; Dai, F.; Luo, X.; Hong, B. Expression of programmed death-1 and programmed death ligand-1 in the peripheral T-lymphocytes from patients with chronic periodontitis. Zhonghua Kou Qiang Yi Xue Za Zhi 2014, 49, 216–219. [Google Scholar]
- Bailly, C. The implication of the PD-1/PD-L1 checkpoint in chronic periodontitis suggests novel therapeutic opportunities with natural products. Jpn. Dent. Sci. Rev. 2020, 56, 90–96. [Google Scholar] [CrossRef]
- Kang, N.S.; Moon, E.Y.; Cho, C.G.; Pyo, S. Immunomodulating effect of garlic component, allicin, on murine peritoneal macrophages. Nutr. Res. 2001, 21, 617–626. [Google Scholar] [CrossRef]
- Arreola, R.; Quintero-Fabián, S.; López-Roa, R.I.; Flores-Gutiérrez, E.O.; Reyes-Grajeda, J.P.; Carrera-Quintanar, L.; Ortuño-Sahagún, D. Immunomodulation and Anti-Inflammatory Effects of Garlic Compounds. J. Immunol. Res. 2015, 2015, 401630. [Google Scholar] [CrossRef] [PubMed]
- Patya, M. Allicin stimulates lymphocytes and elicits an antitumor effect: A possible role of p21ras. Int. Immunol. 2004, 16, 275–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cimasoni, G. The crevicular fluid. Monogr. Oral Sci. 1974, 3, 1–122. [Google Scholar] [PubMed]
- Subbarao, K.C.; Nattuthurai, G.S.; Sundararajan, S.K.; Sujith, I.; Joseph, J.; Syedshah, Y.P. Gingival Crevicular Fluid: An Overview. J. Pharm. Bioallied Sci. 2019, 11, S135–S139. [Google Scholar] [CrossRef]
- Figueredo, C.M.; Ribeiro, M.S.; Fischer, R.G.; Gustafsson, A. Increased interleukin-1beta concentration in gingival crevicular fluid as a characteristic of periodontitis. J. Periodontol. 1999, 70, 1457–1463. [Google Scholar] [CrossRef]
- Afacan, B.; Öztürk, V.Ö.; Paşalı, Ç.; Bozkurt, E.; Köse, T.; Emingil, G. Gingival crevicular fluid and salivary HIF-1α, VEGF, and TNF-α levels in periodontal health and disease. J. Periodontol. 2019, 90, 788–797. [Google Scholar] [CrossRef]
- Sharpe, A.H.; Wherry, E.J.; Ahmed, R.; Freeman, G.J. The function of programmed cell death 1 and its ligands in regulating autoimmunity and infection. Nat. Immunol. 2007, 8, 239–245. [Google Scholar] [CrossRef]
- Fourcade, J.; Sun, Z.; Benallaoua, M.; Guillaume, P.; Luescher, I.F.; Sander, C.; Kirkwood, J.M.; Kuchroo, V.; Zarour, H.M. Upregulation of Tim-3 and PD-1 expression is associated with tumor antigen-specific CD8+ T cell dysfunction in melanoma patients. J. Exp. Med. 2010, 207, 2175–2186. [Google Scholar] [CrossRef]
- Fallah-Rostami, F.; Tabari, M.A.; Esfandiari, B.; Aghajanzadeh, H.; Behzadi, M.Y. Immunomodulatory activity of aged garlic extract against implanted fibrosarcoma tumor in mice. N. Am. J. Med. Sci. 2013, 5, 207–212. [Google Scholar] [CrossRef] [Green Version]
- Torres-Palazzolo, C.; Ramirez, D.; Locatelli, D.; Manucha, W.; Castro, C.; Camargo, A. Bioaccessibility and permeability of bioactive compounds in raw and cooked garlic. J. Food Compos. Anal. 2018, 70, 49–53. [Google Scholar] [CrossRef] [Green Version]
- Alam, R.T.M.; Fawzi, E.M.; Alkhalf, M.I.; Alansari, W.S.; Aleya, L.; Abdel-Daim, M.M. Anti-Inflammatory, Immunomodulatory, and Antioxidant Activities of Allicin, Norfloxacin, or Their Combination against Pasteurella multocida Infection in Male New Zealand Rabbits. Oxid. Med. Cell. Longev. 2018, 2018, 1780956. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Wang, D.; Song, G.; Zuo, C.; Qiao, X.; Qin, S. The effect of combination therapy of allicin and fenofibrate on high fat diet-induced vascular endothelium dysfunction and liver damage in rats. Lipids Health Dis. 2010, 9, 131. [Google Scholar] [CrossRef] [Green Version]
- Lang, A.; Lahav, M.; Sakhnini, E.; Barshack, I.; Fidder, H.H.; Avidan, B.; Bardan, E.; Hershkoviz, R.; Bar-Meir, S.; Chowers, Y. Allicin inhibits spontaneous and TNF-alpha induced secretion of proinflammatory cytokines and chemokines from intestinal epithelial cells. Clin. Nutr. 2004, 23, 1199–1208. [Google Scholar] [CrossRef]
- Humphrey, W.; Dalke, A.; Schulten, K. VMD: Visual molecular dynamics. J. Mol. Graph. 1996, 14, 33–38. [Google Scholar] [CrossRef]
- Fährrolfes, R.; Bietz, S.; Flachsenberg, F.; Meyder, A.; Nittinger, E.; Otto, T.; Volkamer, A.; Rarey, M. ProteinsPlus: A web portal for structure analysis of macromolecules. Nucleic Acids Res. 2017, 45, W337–W343. [Google Scholar] [CrossRef] [Green Version]
- Papapanou, P.N.; Sanz, M.; Buduneli, N.; Dietrich, T.; Feres, M.; Fine, D.H.; Flemmig, T.F.; Garcia, R.; Giannobile, W.V.; Graziani, F.; et al. Periodontitis: Consensus report of workgroup 2 of the 2017 World Workshop on the Classification of Periodontal and Peri-Implant Diseases and Conditions. J. Clin. Periodontol. 2018, 45, S162–S170. [Google Scholar] [CrossRef]
- Karthikeyan, B.V.; Pradeep, A.R. Gingival crevicular fluid and serum leptin: Their relationship to periodontal health and disease. J. Clin. Periodontol. 2007, 34, 467–472. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Rank Pttern | Sub—Rank | Run | Binding Energy | Cluster Rmsd | Reference Rmsd |
|---|---|---|---|---|---|
| 1 RANKING | 1 | 7 | −7.10 | 0.00 | 118.43 |
| 1 RANKING | 2 | 3 | −7.10 | 0.67 | 118.33 |
| 1 RANKING | 3 | 10 | −7.10 | 0.21 | 118.40 |
| 1 RANKING | 4 | 6 | −7.02 | 0.88 | 118.31 |
| 2 RANKING | 1 | 8 | −6.84 | 0.00 | 129.46 |
| 3 RANKING | 1 | 1 | −6.70 | 0.00 | 129.39 |
| 3 RANKING | 2 | 4 | −6.63 | 0.63 | 129.38 |
| 4 RANKING | 1 | 9 | −5.88 | 0.00 | 130.20 |
| 5 RANKING | 1 | 2 | −5.56 | 0.00 | 122.91 |
| 6 RANKING | 1 | 5 | −5.27 | 0.00 | 118.23 |
| Cytokines | Normal | Periodontitis | p-Value |
|---|---|---|---|
| IL-4 | 3.06 ± 0.21 | 3.64 ± 1.06 | 0.4999 |
| IL-2 | 2.09 ± 0.44 | 2.86 ± 1.03 | 0.37 |
| CXCL10 | 1.71 ± 0.45 | 2.08 ± 0.97 | 0.6387 |
| IL-1β | 2.49 ± 0.73 | 7.97 ± 3.08 | 0.065 |
| TNF-α | 3.24 ± 1.06 | 15.63 ± 4.08 | 0.0101 * |
| CCL2 | 4.88 ± 1.04 | 16.29 ± 2.92 | 0.0036 * |
| IL-17A | 0.93 ± 0.34 | 2.39 ± 1.34 | 0.2094 |
| IL-6 | 0.97 ± 0.18 | 7.59 ± 2.19 | 0.0101 * |
| IL-10 | 3.57 ± 0.59 | 2.034 ± 0.9 | 0.0831 |
| IFN-γ | 1.08 ± 0.57 | 2.90 ± 0.76 | 0.0311 * |
| IL-12p17 | 0.78 ± 0.48 | 1.69 ± 0.97 | 0.2755 |
| CXCL8 | 1.92 ± 0.76 | 11.32 ± 1.58 | 0.0006 * |
| TGF-β1 | 1.25 ± 0.33 | 2.43 ± 1.51 | 0.3474 |
| Treatment | OD at 48 h of Allicin Treatment (Mean ± SD) | p-Value |
|---|---|---|
| Control w/o treatment | 0.62 ± 0.12 | - |
| 0.5 μM allicin | 0.78 ± 0.15 | 0.0488 |
| 1 μM allicin | 0.86 ± 0.17 | 0.0386 |
| 2.5 μM allicin | 0.73 ± 0.14 | 0.0479 |
| 5 μM allicin | 0.57 ± 0.11 | 0.7625 |
| 10 μM allicin | 0.62 ± 0.12 | 0.8462 |
| 25 μM allicin | 0.65 ± 0.13 | 0.6405 |
| 50 μM allicin | 0.50 ± 0.09 | 0.3169 |
| 100 μM allicin | 0.34 ± 0.06 | 0.0162 |
| 250 μM allicin | 0.08 ± 0.02 | 0.0016 |
| GCF Treatment | Control w/o Treatment | 1 μL/mL | 2 μL/mL | 5 μL/mL |
|---|---|---|---|---|
| 24 h | 0.12 ± 0.01 | 0.18 ± 0.03 | 0.23 ± 0.01 | 0.21 ± 0.05 |
| p-value | - | 0.0018 | <0.0001 | 0.0044 |
| 48 h | 0.13 ± 0.01 | 0.25 ± 0.09 | 0.31 ± 0.05 | 0.28 ± 0.01 |
| p-value | - | 0.0156 | <0.0001 | <0.0001 |
| 72 h | 0.12 ± 0.02 | 0.29 ± 0.13 | 0.38 ± 0.10 | 0.32 ± 0.04 |
| p-value | - | 0.0189 | 0.0005 | <0.0001 |
| 96 h | 0.13 ± 0.04 | 0.48 ± 0.13 | 0.73 ± 0.01 | 0.51 ± 0.08 |
| p-value | - | 0.0004 | <0.0001 | <0.0001 |
| 120 h | 0.13 ± 0.02 | 0.52 ± 0.14 | 0.79 ± 0.21 | 0.52 ± 0.08 |
| p-value | - | 0.0003 | 0.0001 | <0.0001 |
| GCF Treatment | Control w/o Treatment | 1 μL/mL | 2 μL/mL | 5 μL/mL |
|---|---|---|---|---|
| 24 h | 0.16 ± 0.05 | 0.18 ± 0.04 | 0.19 ± 0.001 | 0.19 ± 0.01 |
| p-value | - | 0.5092 | 0.2393 | 0.2453 |
| 48 h | 0.14 ± 0.03 | 0.23 ± 0.06 | 0.28 ± 0.08 | 0.21 ± 0.12 |
| p-value | - | 0.0169 | 0.0077 | 0.2247 |
| 72 h | 0.17 ± 0.04 | 0.31 ± 0.02 | 0.44 ± 0.12 | 0.26 ± 0.10 |
| p-value | - | 0.0003 | 0.0002 | 0.1018 |
| 96 h | 0.16 ± 0.03 | 0.45 ± 0.12 | 0.69 ± 0.18 | 0.29 ± 0.08 |
| p-value | - | 0.0008 | 0.0002 | 0.0084 |
| 120 h | 0.15 ± 0.02 | 0.51 ± 0.13 | 0.78 ± 0.21 | 0.31 ± 0.15 |
| p-value | - | 0.0004 | 0.0001 | 0.0483 |
| Treatment with Allicin | Control w/o Treatment | 5 μM | 10 μM | 25 μM | 50 μM |
|---|---|---|---|---|---|
| 24 h | 0.12 ± 0.01 | 0.26 ± 0.07 | 0.21 ± 0.02 | 0.16 ± 0.08 | 0.15 ± 0.04 |
| p-value | - | 0.0018 | <0.0001 | 0.3078 | 0.1321 |
| 48 h | 0.31 ± 0.01 | 0.34 ± 0.09 | 0.29 ± 0.15 | 0.21 ± 0.02 | 0.18 ± 0.01 |
| p-value | - | 0.473 | 0.7794 | <0.0001 | <0.0001 |
| 72 h | 0.38 ± 0.11 | 0.43 ± 0.11 | 0.38 ± 0.10 | 0.26 ± 0.03 | 0.22 ± 0.02 |
| p-value | - | 0.4898 | >0.9999 | 0.0408 | 0.0108 |
| 96 h | 0.73 ± 0.02 | 0.79 ± 0.02 | 0.71 ± 0.18 | 0.52 ± 0.01 | 0.45 ± 0.09 |
| p-value | - | 0.0012 | 0.8156 | <0.0001 | 0.0001 |
| 120 h | 0.79 ± 0.22 | 0.78 ± 0.03 | 0.73 ± 0.16 | 0.55 ± 0.04 | 0.51 ± 0.23 |
| p-value | - | 0.9225 | 0.636 | 0.0443 | 0.0868 |
| Treatment with Allicin | Control w/o Treatment | 5 μM | 10 μM | 25 μM | 50 μM |
|---|---|---|---|---|---|
| 24 h | 0.19 ± 0.005 | 0.19 ± 0.10 | 0.19 ± 0.03 | 0.17 ± 0.03 | 0.17 ± 0.08 |
| p-value | - | >0.9999 | >0.9999 | 0.2317 | 0.8104 |
| 48 h | 0.28 ± 0.08 | 0.27 ± 0.08 | 0.25 ± 0.11 | 0.18 ± 0.02 | 0.17 ± 0.02 |
| p-value | - | 0.8525 | 0.6507 | 0.0317 | 0.0232 |
| 72 h | 0.44 ± 0.12 | 0.44 ± 0.11 | 0.39 ± 0.10 | 0.24 ± 0.06 | 0.21 ± 0.05 |
| p-value | - | >0.9999 | 0.4907 | 0.0097 | 0.0035 |
| 96 h | 0.69 ± 0.18 | 0.63 ± 0.01 | 0.57 ± 0.13 | 0.43 ± 0.11 | 0.35 ± 0.09 |
| p-value | - | 0.4885 | 0.2648 | 0.0266 | 0.0059 |
| 120 h | 0.78 ± 0.21 | 0.72 ± 0.03 | 0.66 ± 0.12 | 0.56 ± 0.02 | 0.45 ± 0.02 |
| p-value | - | 0.539 | 0.2966 | 0.0455 | 0.0076 |
| Gene | Forward Primer | Reverse Primer |
|---|---|---|
| TIM-3 | 5′-TCC AAG GAT GCT TAC CAC CAG-3′ | 5′-AAC ACA AAT ATC CAC ATT GGC-3′ |
| LAG-3 | 5′-TCA CAG TGA CTC CCA AAT CCT T-3′ | 5′-GCT CCA CAC AAA GCG TTC TT-3′ |
| GAPDH | 5′-ATG GGG AAG GTG AAG GTC G-3′ | 5′-GGG GTC ATT GAT GGC AAC AAT A-3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Patil, S.; Sayed, M.E.; Mugri, M.H.; Alsharif, K.F.; Salman, A.; Bhandi, S.; Baeshen, H.A.; Balaji, T.M.; Yadalam, P.K.; Varadarajan, S.; et al. Allicin May Promote Reversal of T-Cell Dysfunction in Periodontitis via the PD-1 Pathway. Int. J. Mol. Sci. 2021, 22, 9162. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22179162
Patil S, Sayed ME, Mugri MH, Alsharif KF, Salman A, Bhandi S, Baeshen HA, Balaji TM, Yadalam PK, Varadarajan S, et al. Allicin May Promote Reversal of T-Cell Dysfunction in Periodontitis via the PD-1 Pathway. International Journal of Molecular Sciences. 2021; 22(17):9162. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22179162
Chicago/Turabian StylePatil, Shankargouda, Mohammed E. Sayed, Maryam H. Mugri, Khalaf F. Alsharif, Arif Salman, Shilpa Bhandi, Hosam Ali Baeshen, Thodur Madapusi Balaji, Pradeep Kumar Yadalam, Saranya Varadarajan, and et al. 2021. "Allicin May Promote Reversal of T-Cell Dysfunction in Periodontitis via the PD-1 Pathway" International Journal of Molecular Sciences 22, no. 17: 9162. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22179162