
Visual Cortex Engagement in Retinitis Pigmentosa

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
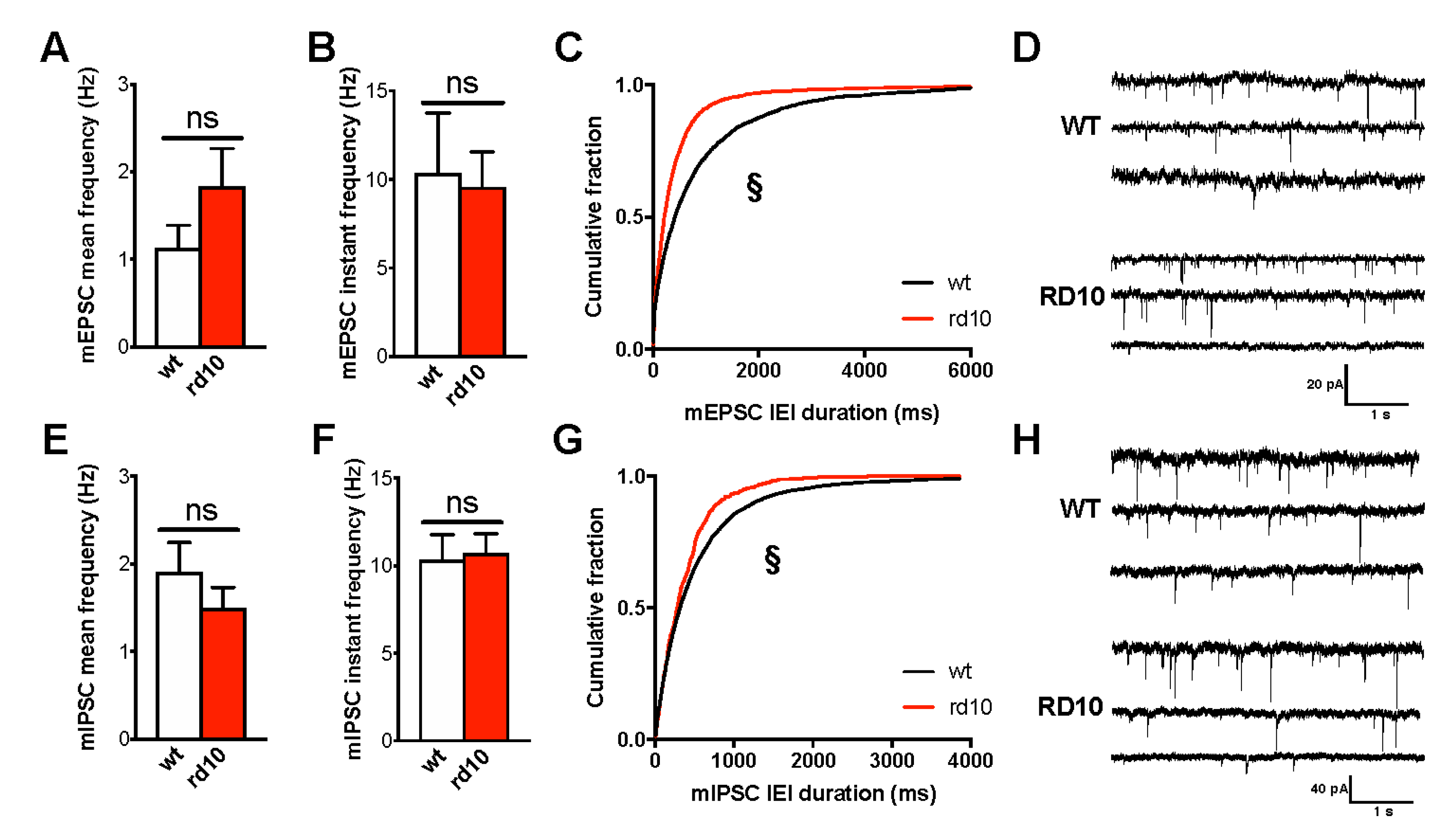
2.1. Altered Excitation/Inhibition Balance in the Visual Cortex of rd10 Animals
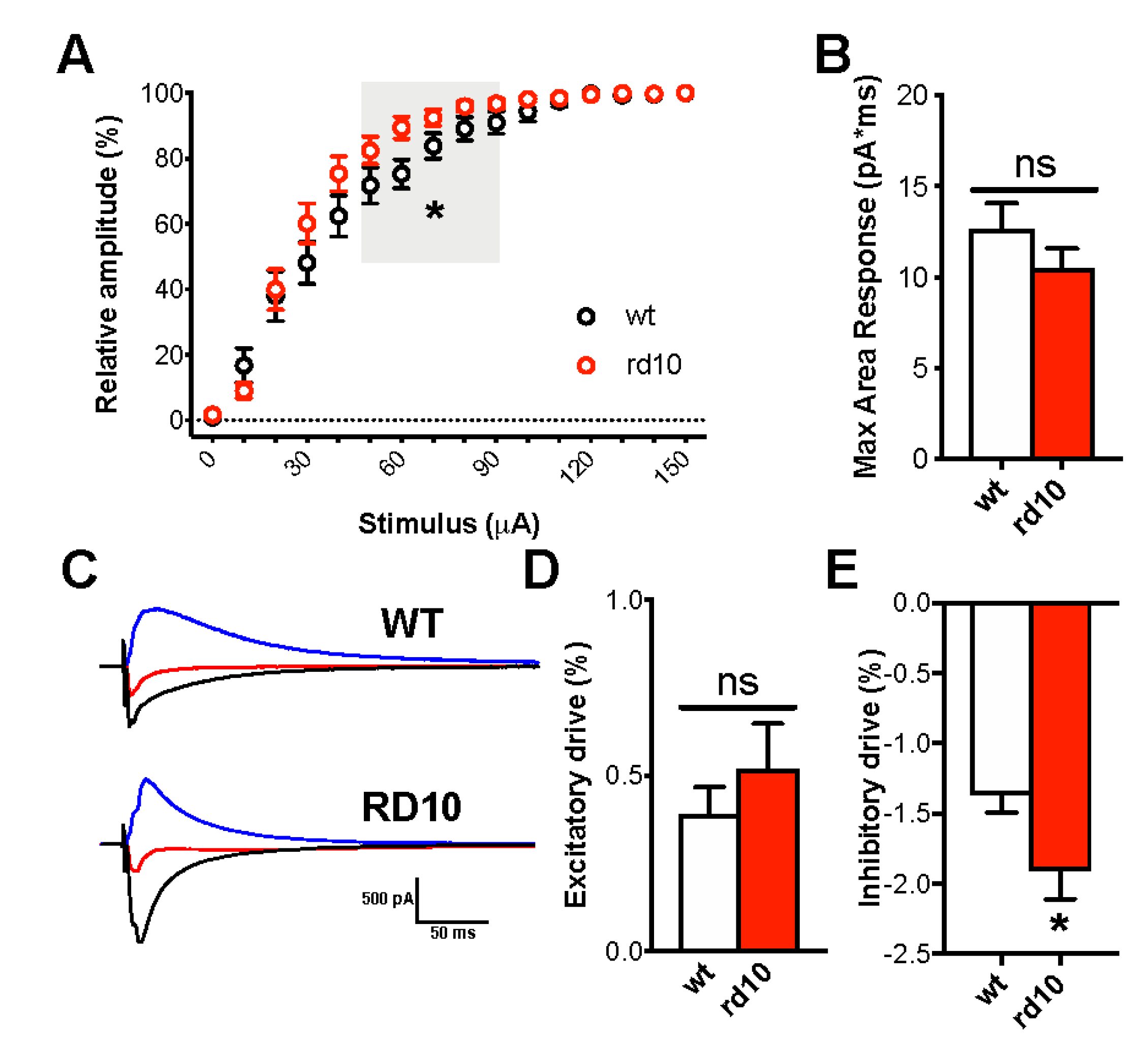
2.2. Rd Mutation Shifts the Balance towards Inhibitory Overdrive in the Visual Cortex
2.3. Thalamic Synaptic Processing in RP Animals
2.4. Lowering Inhibition in the Visual Cortex of rd10 Animals Hampers Visual Acuity
3. Discussion
4. Material and Methods
4.1. Animals
4.2. Western Blot
4.3. Analysis of Neurotransmitter Release in Visual Cortex Synaptosomes
4.4. In Vitro Electrophysiology
4.5. Drug Administration
4.6. Behavioral Assessment of Visual Acuity and Contrast Sensitivity
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hartong, D.T.; Berson, E.L.; Dryja, T.P. Retinitis Pigmentosa. Lancet 2006, 368, 1795–1809. [Google Scholar] [CrossRef]
- Dias, M.F.; Joo, K.; Kemp, J.A.; Fialho, S.L.; da Silva Cunha, A., Jr.; Woo, S.J.; Kwon, Y.J. Molecular Genetics and Emerging Therapies for Retinitis Pigmentosa: Basic Research and Clinical Perspectives. Prog. Retin. Eye Res. 2018, 63, 107–131. [Google Scholar] [CrossRef]
- O’Neal, T.B.; Luther, E.E. Retinitis Pigmentosa. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2020. [Google Scholar]
- Rayapudi, S.; Schwartz, S.G.; Wang, X.; Chavis, P. Vitamin A and Fish Oils for Retinitis Pigmentosa. Cochrane Database Syst. Rev. 2013, CD008428. [Google Scholar] [CrossRef] [Green Version]
- Aguirre, G.K.; Komáromy, A.M.; Cideciyan, A.V.; Brainard, D.H.; Aleman, T.S.; Roman, A.J.; Avants, B.B.; Gee, J.C.; Korczykowski, M.; Hauswirth, W.W.; et al. Canine and Human Visual Cortex Intact and Responsive despite Early Retinal Blindness from RPE65 Mutation. PLoS Med. 2007, 4, e230. [Google Scholar] [CrossRef]
- Cohen, E.D. Prosthetic Interfaces with the Visual System: Biological Issues. J. Neural Eng. 2007, 4, R14–R31. [Google Scholar] [CrossRef]
- Lagali, P.S.; Balya, D.; Awatramani, G.B.; Münch, T.A.; Kim, D.S.; Busskamp, V.; Cepko, C.L.; Roska, B. Light-Activated Channels Targeted to ON Bipolar Cells Restore Visual Function in Retinal Degeneration. Nat. Neurosci. 2008, 11, 667–675. [Google Scholar] [CrossRef]
- Strettoi, E.; Gargini, C.; Novelli, E.; Sala, G.; Piano, I.; Gasco, P.; Ghidoni, R. Inhibition of Ceramide Biosynthesis Preserves Photoreceptor Structure and Function in a Mouse Model of Retinitis Pigmentosa. Proc. Natl. Acad. Sci. USA 2010, 107, 18706–18711. [Google Scholar] [CrossRef] [Green Version]
- Thyagarajan, S.; van Wyk, M.; Lehmann, K.; Löwel, S.; Feng, G.; Wässle, H. Visual Function in Mice with Photoreceptor Degeneration and Transgenic Expression of Channelrhodopsin 2 in Ganglion Cells. J. Neurosci. 2010, 30, 8745–8758. [Google Scholar] [CrossRef]
- Maya-Vetencourt, J.F.; Ghezzi, D.; Antognazza, M.R.; Colombo, E.; Mete, M.; Feyen, P.; Desii, A.; Buschiazzo, A.; Di Paolo, M.; Di Marco, S.; et al. A Fully Organic Retinal Prosthesis Restores Vision in a Rat Model of Degenerative Blindness. Nat. Mater. 2017, 16, 681–689. [Google Scholar] [CrossRef]
- Marc, R.E.; Jones, B.W.; Watt, C.B.; Strettoi, E. Neural Remodeling in Retinal Degeneration. Prog. Retin. Eye Res. 2003, 22, 607–655. [Google Scholar] [CrossRef]
- Strettoi, E.; Pignatelli, V.; Rossi, C.; Porciatti, V.; Falsini, B. Remodeling of Second-Order Neurons in the Retina of Rd/rd Mutant Mice. Vision Res. 2003, 43, 867–877. [Google Scholar] [CrossRef] [Green Version]
- Mazzoni, F.; Novelli, E.; Strettoi, E. Retinal Ganglion Cells Survive and Maintain Normal Dendritic Morphology in a Mouse Model of Inherited Photoreceptor Degeneration. J. Neurosci. 2008, 28, 14282–14292. [Google Scholar] [CrossRef]
- Margolis, D.J.; Newkirk, G.; Euler, T.; Detwiler, P.B. Functional Stability of Retinal Ganglion Cells after Degeneration-Induced Changes in Synaptic Input. J. Neurosci. 2008, 28, 6526–6536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stasheff, S.F. Emergence of Sustained Spontaneous Hyperactivity and Temporary Preservation of OFF Responses in Ganglion Cells of the Retinal Degeneration (rd1) Mouse. J. Neurophysiol. 2008, 99, 1408–1421. [Google Scholar] [CrossRef] [PubMed]
- Menzler, J.; Zeck, G. Network Oscillations in Rod-Degenerated Mouse Retinas. J. Neurosci. 2011, 31, 2280–2291. [Google Scholar] [CrossRef] [Green Version]
- Sekirnjak, C.; Jepson, L.H.; Hottowy, P.; Sher, A.; Dabrowski, W.; Litke, A.M.; Chichilnisky, E.J. Changes in Physiological Properties of Rat Ganglion Cells during Retinal Degeneration. J. Neurophysiol. 2011, 105, 2560–2571. [Google Scholar] [CrossRef] [Green Version]
- Sauvé, Y.; Girman, S.V.; Wang, S.; Keegan, D.J.; Lund, R.D. Preservation of Visual Responsiveness in the Superior Colliculus of RCS Rats after Retinal Pigment Epithelium Cell Transplantation. Neuroscience 2002, 114, 389–401. [Google Scholar] [CrossRef]
- Alexander, K.R.; Rajagopalan, A.S.; Seiple, W.; Zemon, V.M.; Fishman, G.A. Contrast Response Properties of Magnocellular and Parvocellular Pathways in Retinitis Pigmentosa Assessed by the Visual Evoked Potential. Investig. Ophthalmol. Vis. Sci. 2005, 46, 2967–2973. [Google Scholar] [CrossRef] [PubMed]
- Milam, A.H.; Li, Z.Y.; Fariss, R.N. Histopathology of the Human Retina in Retinitis Pigmentosa. Prog. Retin. Eye Res. 1998, 17, 175–205. [Google Scholar]
- Schoth, F.; Burgel, U.; Dorsch, R.; Reinges, M.H.T.; Krings, T. Diffusion Tensor Imaging in Acquired Blind Humans. Neurosci. Lett. 2006, 398, 178–182. [Google Scholar] [CrossRef]
- Ohno, N.; Murai, H.; Suzuki, Y.; Kiyosawa, M.; Tokumaru, A.M.; Ishii, K.; Ohno-Matsui, K. Alteration of the Optic Radiations Using Diffusion-Tensor MRI in Patients with Retinitis Pigmentosa. Br. J. Ophthalmol. 2015, 99, 1051–1054. [Google Scholar] [CrossRef]
- Gothe, J.; Brandt, S.A.; Irlbacher, K.; Röricht, S.; Sabel, B.A.; Meyer, B.-U. Changes in Visual Cortex Excitability in Blind Subjects as Demonstrated by Transcranial Magnetic Stimulation. Brain 2002, 125, 479–490. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.-J.; Mahadevappa, M.; Roizenblatt, R.; Weiland, J.; Humayun, M. Neural Responses Elicited by Electrical Stimulation of the Retina. Trans. Am. Ophthalmol. Soc. 2006, 104, 252–259. [Google Scholar]
- Gekeler, F.; Messias, A.; Ottinger, M.; Bartz-Schmidt, K.U.; Zrenner, E. Phosphenes Electrically Evoked with DTL Electrodes: A Study in Patients with Retinitis Pigmentosa, Glaucoma, and Homonymous Visual Field Loss and Normal Subjects. Investig. Ophthalmol. Vis. Sci. 2006, 47, 4966–4974. [Google Scholar] [CrossRef]
- Naycheva, L.; Schatz, A.; Röck, T.; Willmann, G.; Messias, A.; Bartz-Schmidt, K.U.; Zrenner, E.; Gekeler, F. Phosphene Thresholds Elicited by Transcorneal Electrical Stimulation in Healthy Subjects and Patients with Retinal Diseases. Investig. Ophthalmol. Vis. Sci. 2012, 53, 7440–7448. [Google Scholar] [CrossRef] [PubMed]
- Jacobson, S.G.; Knighton, R.W.; Levene, R.M. Dark- and Light-Adapted Visual Evoked Cortical Potentials in Retinitis Pigmentosa. Doc. Ophthalmol. 1985, 60, 189–196. [Google Scholar] [CrossRef]
- Janáky, M.; Pálffy, A.; Horváth, G.; Tuboly, G.; Benedek, G. Pattern-Reversal Electroretinograms and Visual Evoked Potentials in Retinitis Pigmentosa. Doc. Ophthalmol. 2008, 117, 27–36. [Google Scholar] [CrossRef]
- Huang, X.; Zhou, F.-Q.; Dan, H.-D.; Shen, Y. Abnormal Intrinsic Brain Activity in Individuals with Peripheral Vision Loss because of Retinitis Pigmentosa Using Amplitude of Low-Frequency Fluctuations. Neuroreport 2018, 29, 1323–1332. [Google Scholar] [CrossRef] [PubMed]
- Amendola, T.; Fiore, M.; Aloe, L. Postnatal Changes in Nerve Growth Factor and Brain Derived Neurotrophic Factor Levels in the Retina, Visual Cortex, and Geniculate Nucleus in Rats with Retinitis Pigmentosa. Neurosci. Lett. 2003, 345, 37–40. [Google Scholar] [CrossRef]
- Chen, K.; Wang, Y.; Liang, X.; Zhang, Y.; Ng, T.K.; Chan, L.L.H. Electrophysiology Alterations in Primary Visual Cortex Neurons of Retinal Degeneration (S334ter-Line-3) Rats. Sci. Rep. 2016, 6, 26793. [Google Scholar] [CrossRef] [PubMed]
- Lunghi, C.; Galli-Resta, L.; Binda, P.; Cicchini, G.M.; Placidi, G.; Falsini, B.; Morrone, M.C. Visual Cortical Plasticity in Retinitis Pigmentosa. Investig. Ophthalmol. Vis. Sci. 2019, 60, 2753–2763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Begenisic, T.; Mazziotti, R.; Sagona, G.; Lupori, L.; Sale, A.; Galli, L.; Baroncelli, L. Preservation of Visual Cortex Plasticity in Retinitis Pigmentosa. Neuroscience 2020, 424, 205–210. [Google Scholar] [CrossRef]
- Chang, B.; Hawes, N.L.; Hurd, R.E.; Davisson, M.T.; Nusinowitz, S.; Heckenlively, J.R. Retinal Degeneration Mutants in the Mouse. Vision Res. 2002, 42, 517–525. [Google Scholar] [CrossRef] [Green Version]
- Gargini, C.; Terzibasi, E.; Mazzoni, F.; Strettoi, E. Retinal Organization in the Retinal Degeneration 10 (rd10) Mutant Mouse: A Morphological and ERG Study. J. Comp. Neurol. 2007, 500, 222–238. [Google Scholar] [CrossRef] [Green Version]
- Sale, A.; Maya Vetencourt, J.F.; Medini, P.; Cenni, M.C.; Baroncelli, L.; De Pasquale, R.; Maffei, L. Environmental Enrichment in Adulthood Promotes Amblyopia Recovery through a Reduction of Intracortical Inhibition. Nat. Neurosci. 2007, 10, 679–681. [Google Scholar] [CrossRef]
- Baroncelli, L.; Bonaccorsi, J.; Milanese, M.; Bonifacino, T.; Giribaldi, F.; Manno, I.; Cenni, M.C.; Berardi, N.; Bonanno, G.; Maffei, L.; et al. Enriched Experience and Recovery from Amblyopia in Adult Rats: Impact of Motor, Social and Sensory Components. Neuropharmacology 2012, 62, 2388–2397. [Google Scholar] [CrossRef]
- Hensch, T.K.; Fagiolini, M. Excitatory-Inhibitory Balance and Critical Period Plasticity in Developing Visual Cortex. Prog. Brain Res. 2005, 147, 115–124. [Google Scholar]
- Sale, A.; Berardi, N.; Spolidoro, M.; Baroncelli, L.; Maffei, L. GABAergic Inhibition in Visual Cortical Plasticity. Front. Cell. Neurosci. 2010, 4, 10. [Google Scholar] [CrossRef] [Green Version]
- Isaacson, J.S.; Scanziani, M. How Inhibition Shapes Cortical Activity. Neuron 2011, 72, 231–243. [Google Scholar] [CrossRef] [Green Version]
- Nahmani, M.; Erisir, A. VGluT2 Immunochemistry Identifies Thalamocortical Terminals in Layer 4 of Adult and Developing Visual Cortex. J. Comp. Neurol. 2005, 484, 458–473. [Google Scholar] [CrossRef]
- Sherman, S.M. Thalamus Plays a Central Role in Ongoing Cortical Functioning. Nat. Neurosci. 2016, 19, 533–541. [Google Scholar] [CrossRef]
- Fernandez, F.; Morishita, W.; Zuniga, E.; Nguyen, J.; Blank, M.; Malenka, R.C.; Garner, C.C. Pharmacotherapy for Cognitive Impairment in a Mouse Model of Down Syndrome. Nat. Neurosci. 2007, 10, 411–413. [Google Scholar] [CrossRef]
- Landfield, P.W.; Baskin, R.K.; Pitler, T.A. Brain Aging Correlates: Retardation by Hormonal-Pharmacological Treatments. Science 1981, 214, 581–584. [Google Scholar] [CrossRef]
- Gias, C.; Vugler, A.; Lawrence, J.; Carr, A.J.; Chen, L.L.; Ahmado, A.; Semo, M.; Coffey, P.J. Degeneration of Cortical Function in the Royal College of Surgeons Rat. Vision Res. 2011, 51, 2176–2185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holopigian, K.; Shuwairi, S.M.; Greenstein, V.C.; Winn, B.J.; Zhang, X.; Carr, R.E.; Hood, D.C. Multifocal Visual Evoked Potentials to Cone Specific Stimuli in Patients with Retinitis Pigmentosa. Vision Res. 2005, 45, 3244–3252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Versendaal, D.; Levelt, C.N. Inhibitory Interneurons in Visual Cortical Plasticity. Cell. Mol. Life Sci. 2016, 73, 3677–3691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Somogyi, P.; Kisvárday, Z.F.; Martin, K.A.; Whitteridge, D. Synaptic Connections of Morphologically Identified and Physiologically Characterized Large Basket Cells in the Striate Cortex of Cat. Neuroscience 1983, 10, 261–294. [Google Scholar] [CrossRef]
- Fagiolini, M.; Pizzorusso, T.; Berardi, N.; Domenici, L.; Maffei, L. Functional Postnatal Development of the Rat Primary Visual Cortex and the Role of Visual Experience: Dark Rearing and Monocular Deprivation. Vision Res. 1994, 34, 709–720. [Google Scholar] [CrossRef]
- Crair, M.C.; Gillespie, D.C.; Stryker, M.P. The Role of Visual Experience in the Development of Columns in Cat Visual Cortex. Science 1998, 279, 566–570. [Google Scholar] [CrossRef] [Green Version]
- White, L.E.; Coppola, D.M.; Fitzpatrick, D. The Contribution of Sensory Experience to the Maturation of Orientation Selectivity in Ferret Visual Cortex. Nature 2001, 411, 1049–1052. [Google Scholar] [CrossRef]
- Ko, H.; Mrsic-Flogel, T.D.; Hofer, S.B. Emergence of Feature-Specific Connectivity in Cortical Microcircuits in the Absence of Visual Experience. J. Neurosci. 2014, 34, 9812–9816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montey, K.L.; Quinlan, E.M. Recovery from Chronic Monocular Deprivation Following Reactivation of Thalamocortical Plasticity by Dark Exposure. Nat. Commun. 2011, 2, 317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erchova, I.; Vasalauskaite, A.; Longo, V.; Sengpiel, F. Enhancement of Visual Cortex Plasticity by Dark Exposure. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2017, 372, 20160159. [Google Scholar] [CrossRef]
- Mathis, T.; Housset, M.; Eandi, C.; Beguier, F.; Touhami, S.; Reichman, S.; Augustin, S.; Gondouin, P.; Sahel, J.-A.; Kodjikian, L.; et al. Activated Monocytes Resist Elimination by Retinal Pigment Epithelium and Downregulate Their OTX2 Expression via TNF-α. Aging Cell 2017, 16, 173–182. [Google Scholar] [CrossRef] [PubMed]
- Sugiyama, S.; Di Nardo, A.A.; Aizawa, S.; Matsuo, I.; Volovitch, M.; Prochiantz, A.; Hensch, T.K. Experience-Dependent Transfer of Otx2 Homeoprotein into the Visual Cortex Activates Postnatal Plasticity. Cell 2008, 134, 508–520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beurdeley, M.; Spatazza, J.; Lee, H.H.C.; Sugiyama, S.; Bernard, C.; Di Nardo, A.A.; Hensch, T.K.; Prochiantz, A. Otx2 Binding to Perineuronal Nets Persistently Regulates Plasticity in the Mature Visual Cortex. J. Neurosci. 2012, 32, 9429–9437. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, N.; Ikeda, Y.; Notomi, S.; Ishikawa, K.; Murakami, Y.; Hisatomi, T.; Enaida, H.; Ishibashi, T. Clinical Evidence of Sustained Chronic Inflammatory Reaction in Retinitis Pigmentosa. Ophthalmology 2013, 120, 100–105. [Google Scholar] [CrossRef]
- Guadagni, V.; Novelli, E.; Piano, I.; Gargini, C.; Strettoi, E. Pharmacological Approaches to Retinitis Pigmentosa: A Laboratory Perspective. Prog. Retin. Eye Res. 2015, 48, 62–81. [Google Scholar] [CrossRef]
- Apulei, J.; Kim, N.; Testa, D.; Ribot, J.; Morizet, D.; Bernard, C.; Jourdren, L.; Blugeon, C.; Di Nardo, A.A.; Prochiantz, A. Non-Cell Autonomous OTX2 Homeoprotein Regulates Visual Cortex Plasticity through Gadd45b/g. Cereb. Cortex 2018, 29, 2384–2395. [Google Scholar] [CrossRef]
- Milanese, M.; Zappettini, S.; Onofri, F.; Musazzi, L.; Tardito, D.; Bonifacino, T.; Messa, M.; Racagni, G.; Usai, C.; Benfenati, F.; et al. Abnormal Exocytotic Release of Glutamate in a Mouse Model of Amyotrophic Lateral Sclerosis. J. Neurochem. 2011, 116, 1028–1042. [Google Scholar] [CrossRef]
- Fleck, M.W.; Barrionuevo, G.; Palmer, A.M. Synaptosomal and Vesicular Accumulation of L-Glutamate, L-Aspartate and D-Aspartate. Neurochem. Int. 2001, 39, 217–225. [Google Scholar] [CrossRef]
- Raiteri, L.; Zappettini, S.; Milanese, M.; Fedele, E.; Raiteri, M.; Bonanno, G. Mechanisms of Glutamate Release Elicited in Rat Cerebrocortical Nerve Endings by “Pathologically” Elevated Extraterminal K+ Concentrations. J. Neurochem. 2007, 103, 952–961. [Google Scholar] [CrossRef]
- Giralt, M.T.; Bonanno, G.; Raiteri, M. GABA Terminal Autoreceptors in the Pars Compacta and in the Pars Reticulata of the Rat Substantia Nigra Are GABAB. Eur. J. Pharmacol. 1990, 175, 137–144. [Google Scholar] [CrossRef]
- Cacciante, F.; Gennaro, M.; Sagona, G.; Mazziotti, R.; Lupori, L.; Cerri, E.; Putignano, E.; Butt, M.; Do, M.-H.T.; McKew, J.C.; et al. Cyclocreatine Treatment Ameliorates the Cognitive, Autistic and Epileptic Phenotype in a Mouse Model of Creatine Transporter Deficiency. Sci. Rep. 2020, 10, 18361. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pietra, G.; Bonifacino, T.; Talamonti, D.; Bonanno, G.; Sale, A.; Galli, L.; Baroncelli, L. Visual Cortex Engagement in Retinitis Pigmentosa. Int. J. Mol. Sci. 2021, 22, 9412. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22179412
Pietra G, Bonifacino T, Talamonti D, Bonanno G, Sale A, Galli L, Baroncelli L. Visual Cortex Engagement in Retinitis Pigmentosa. International Journal of Molecular Sciences. 2021; 22(17):9412. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22179412
Chicago/Turabian StylePietra, Gianluca, Tiziana Bonifacino, Davide Talamonti, Giambattista Bonanno, Alessandro Sale, Lucia Galli, and Laura Baroncelli. 2021. "Visual Cortex Engagement in Retinitis Pigmentosa" International Journal of Molecular Sciences 22, no. 17: 9412. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22179412