
Isoleucilactucin Ameliorates Coal Fly Ash-Induced Inflammation through the NF-κB and MAPK Pathways in MH-S Cells




Abstract
:1. Introduction
2. Results
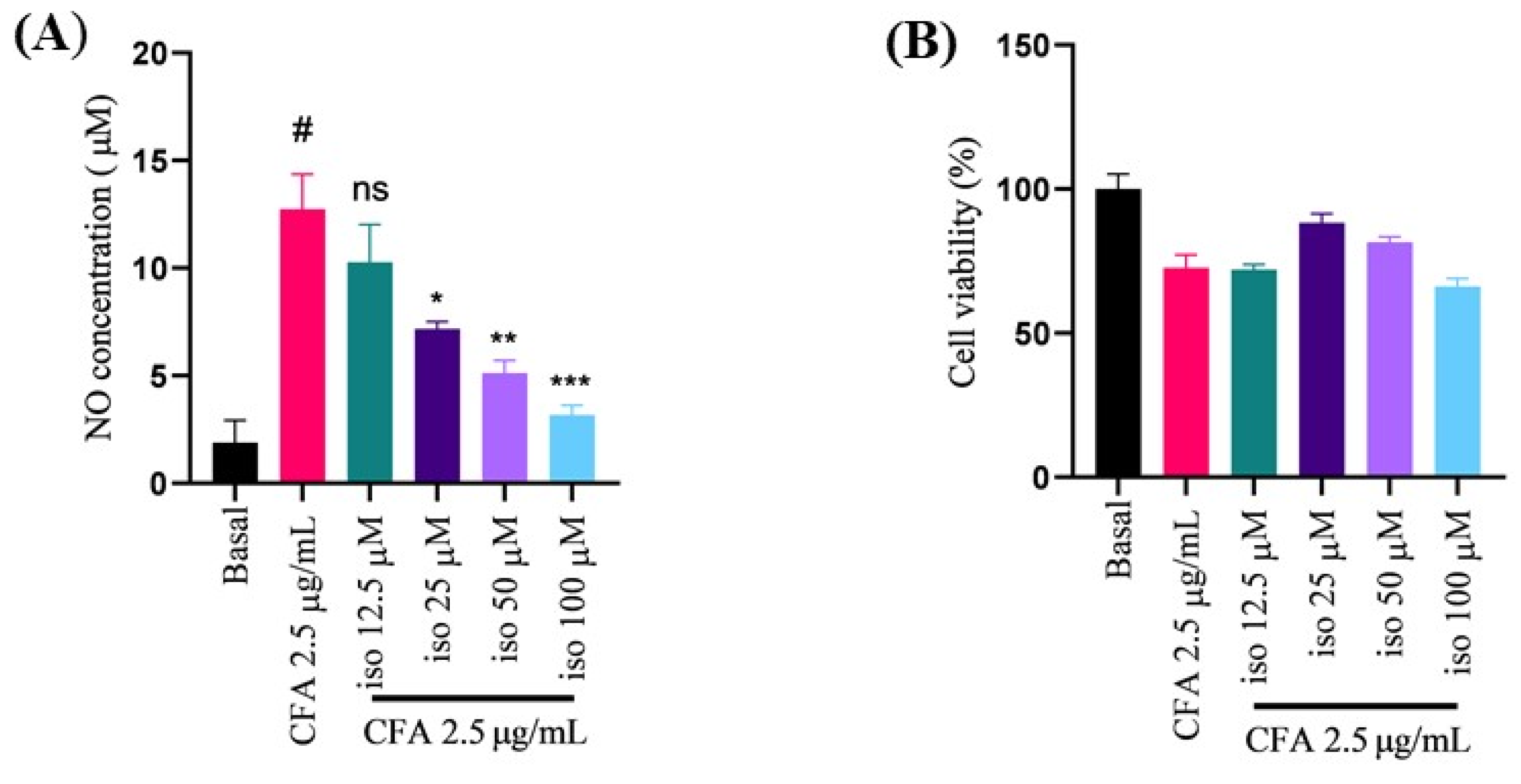
2.1. Isoleucilactucin Protected against CFA-Induced Nitric Oxide Production and Cell Death in Alveolar Macrophage (MH-S) Cells
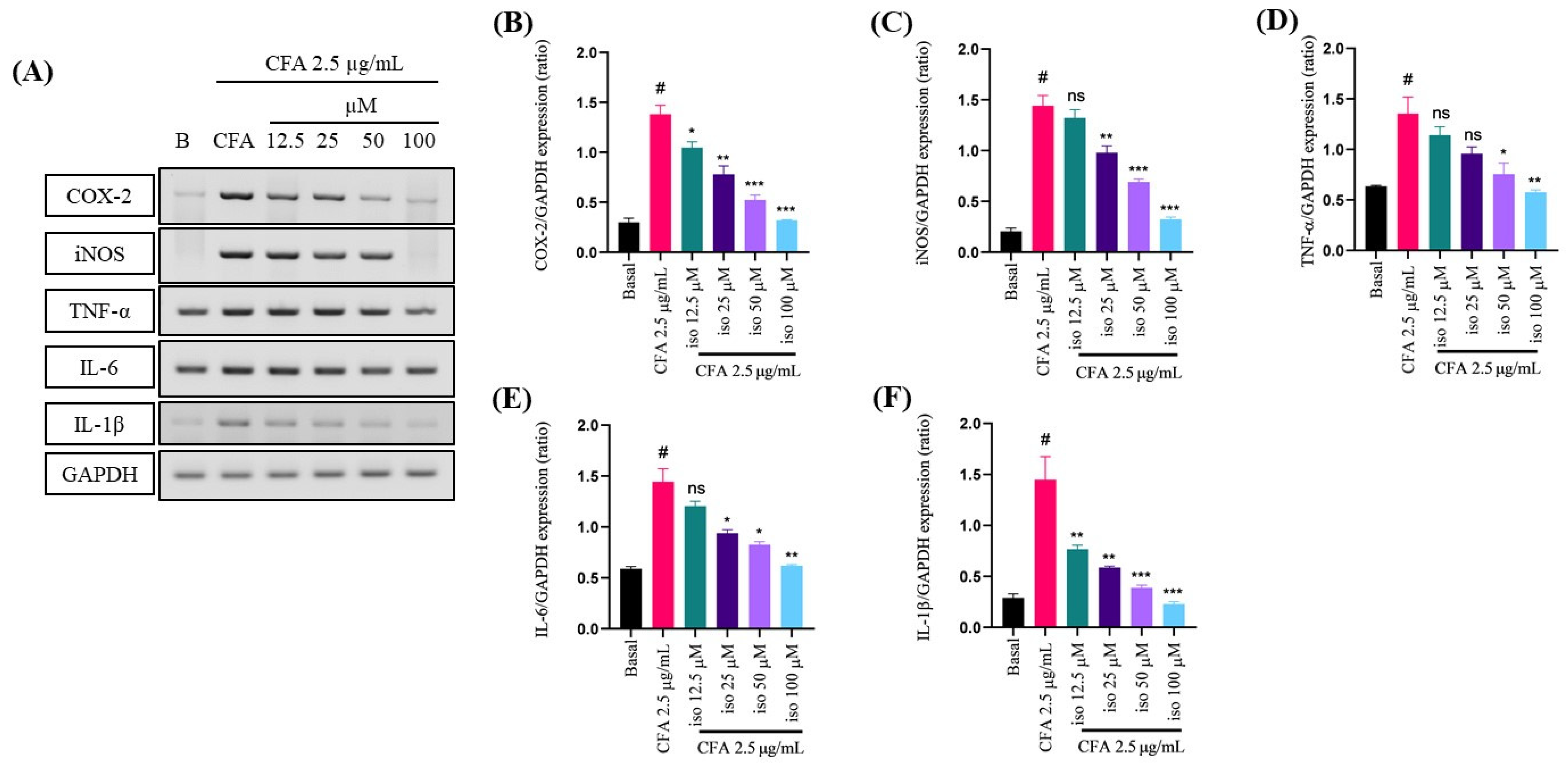
2.2. Suppressive Effect of Isoleucilactucin on CFA-Induced Proinflammatory Cytokines in MH-S Cells
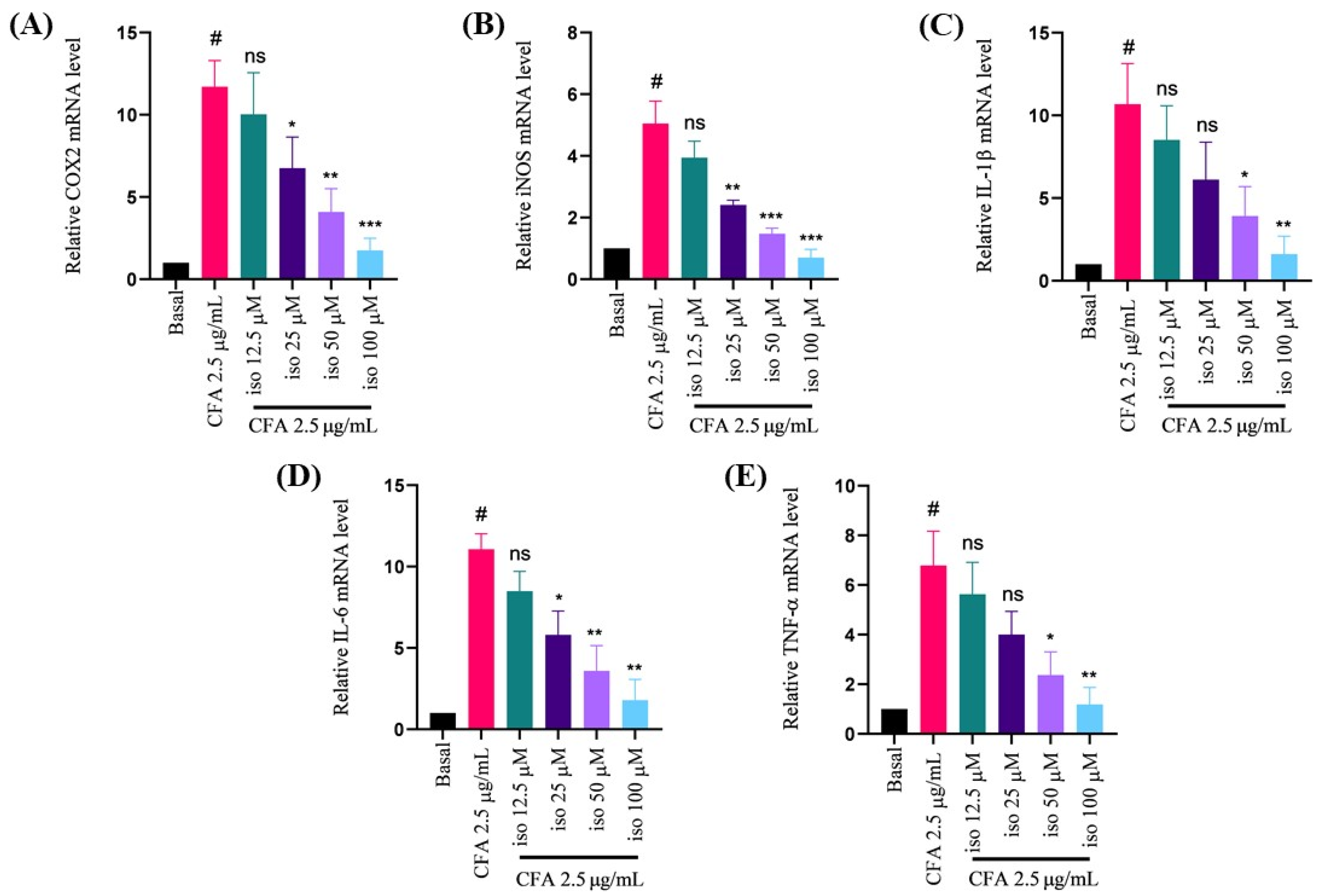
2.3. Isoleucilactucin Ameliorated CFA-Induced mRNA Expression of Proinflammatory Cytokines in MH-S Cells
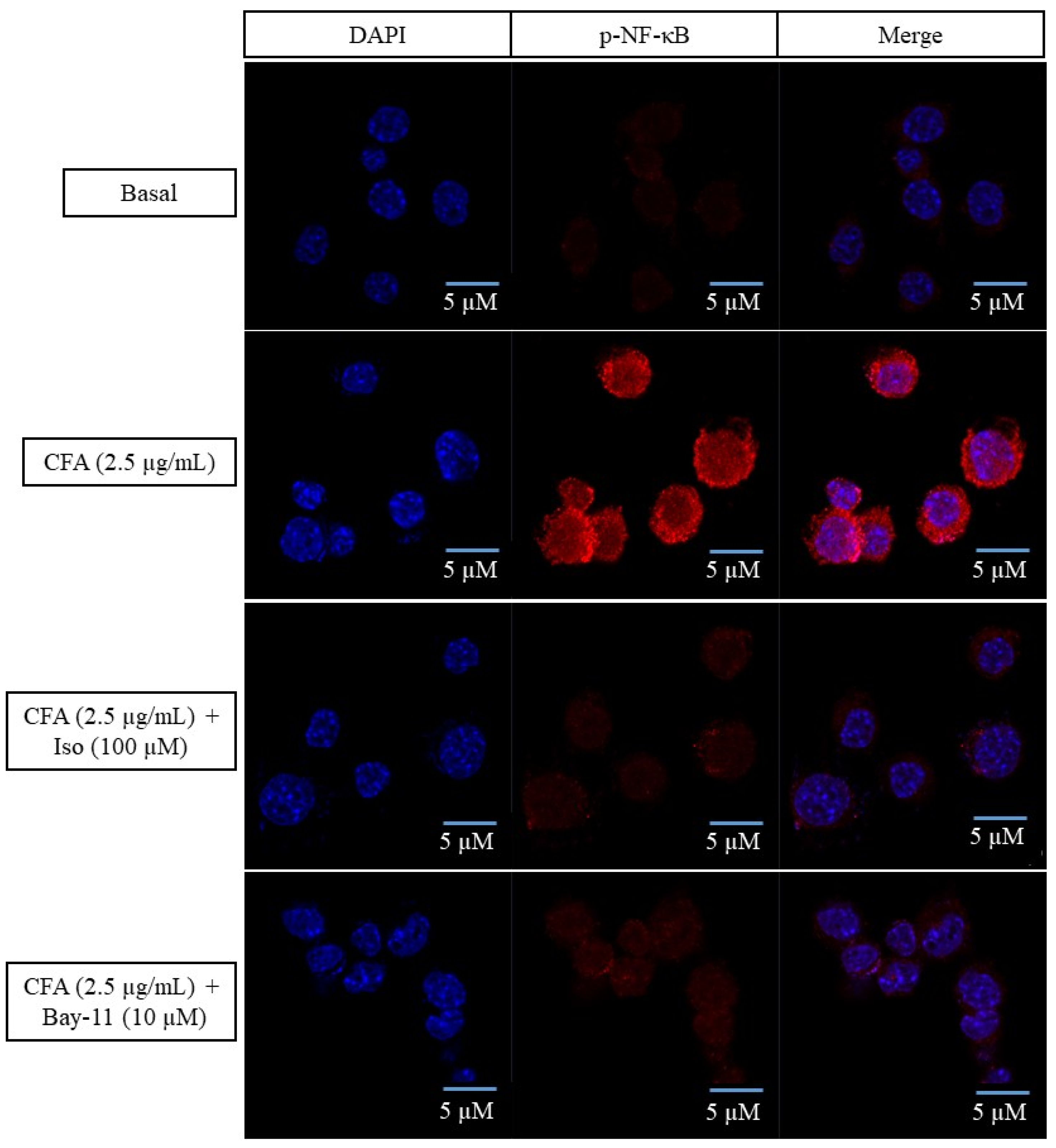
2.4. Isoleucilactucin Inhibited the Translocation of Nuclear Factor-Kappa B in CFA-Treated MH-S Cells
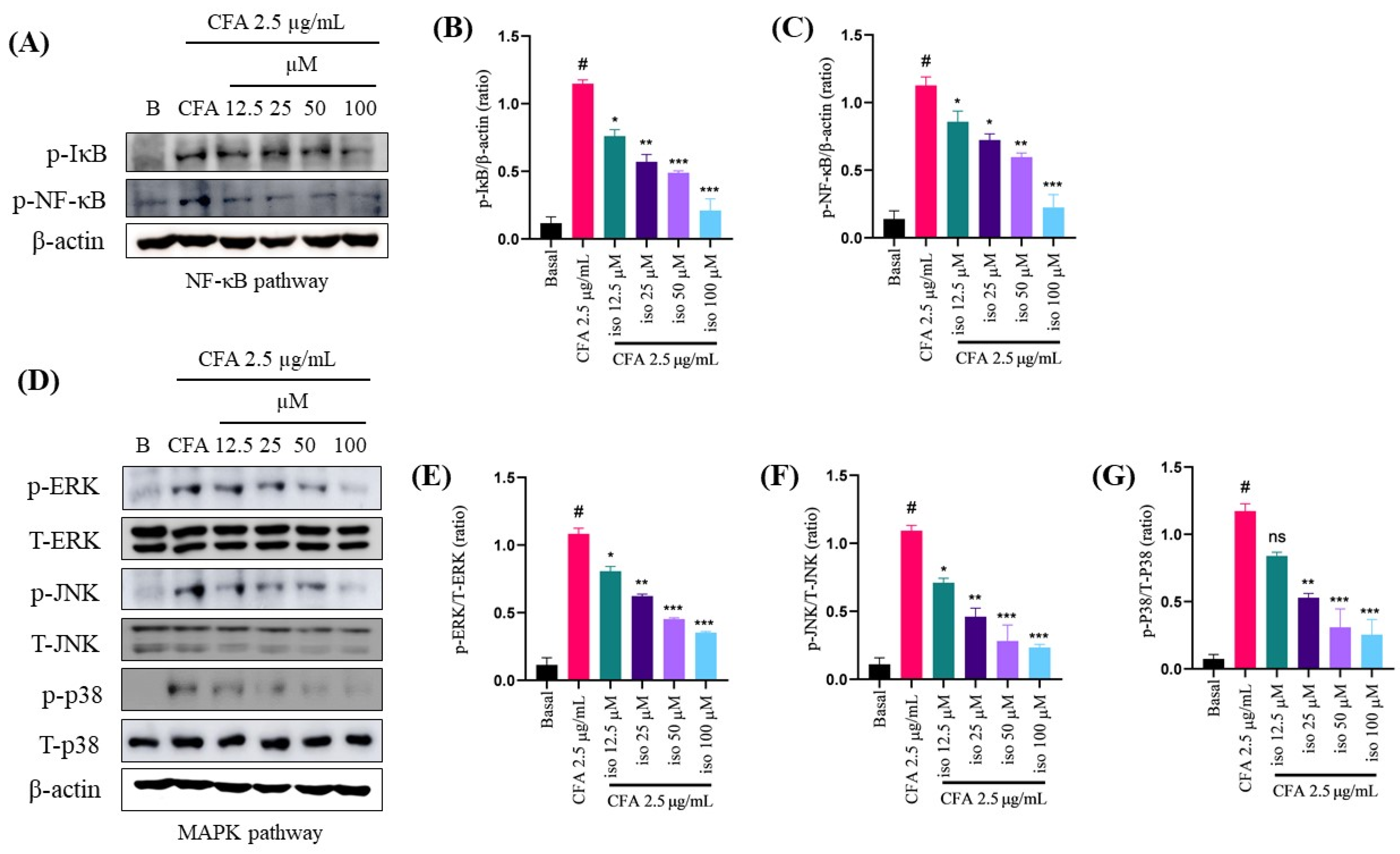
2.5. Isoleucilactucin Inhibits the Activation of Nuclear Factor-Kappa B and Mitogen-Activated Protein Kinase Signaling Pathways in CFA-Treated MH-S Cells
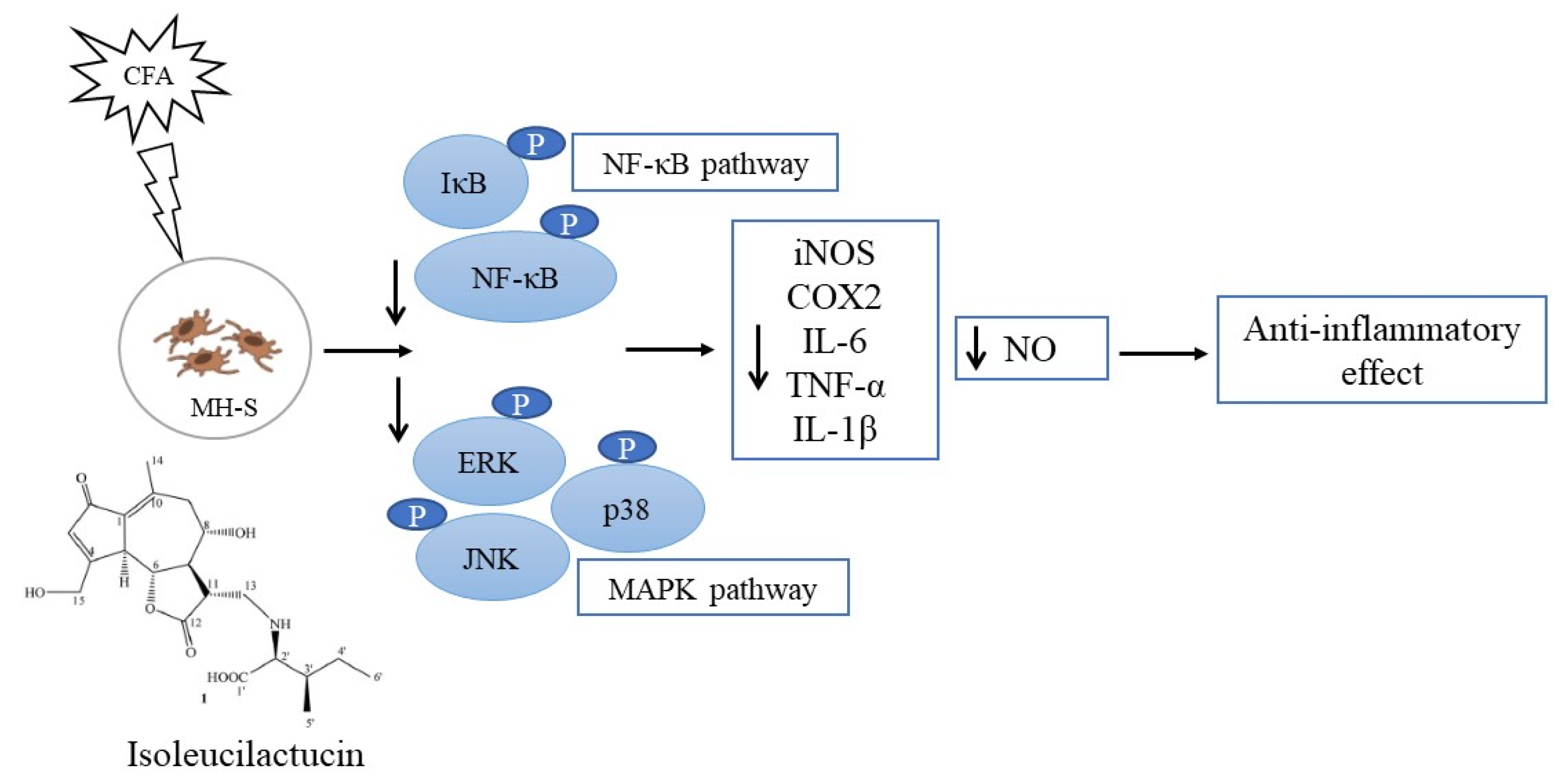
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Plant Material Collection
4.3. Plan Material Extraction and Isolation
4.4. Cell Culture and Treatment
4.5. Nitric Oxide Assay
4.6. Cell Viability Assay
4.7. PCR Analysis
4.8. Immunofluorescence Analysis
4.9. Western Blot Analysis
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| CFA | coal fly ash |
| MH-S | alveolar macrophages |
| NO | nitric oxide |
| PCR | polymerase chain reaction |
| COX2 | cyclooxygenase-2 |
| iNOS | inducible NO synthase |
| IL-6 | interleukin-6 |
| IL-1β | interleukin-1 beta |
| TNF-α | tumor necrosis factor-alpha |
| IκB | inhibitor of kappa B |
| p-NF-κB | phosphorylated nuclear factor-kappa B |
| MAPK | mitogen-activated protein kinase |
| JNK | c-Jun N-terminal kinase |
| ERK | extracellular signal-regulated kinase |
| COPD | chronic obstructive pulmonary disease |
| DMSO | dimethyl sulfoxide |
| RPMI | roswell park memorial institute medium |
| FBS | fetal bovine serum |
| IF | immunofluorescence |
| DAPI | 4, 6-diamidino-2-phenylindole |
References
- Saba, E.; Lee, Y.-S.; Yang, W.-K.; Lee, Y.Y.; Kim, M.; Woo, S.-M.; Kim, K.; Kwon, Y.-S.; Kim, T.-H.; Kwak, D.; et al. Effects of a herbal formulation, KGC3P, and its individual component, nepetin, on coal fly dust-induced airway inflammation. Sci. Rep. 2020, 10, 14036. [Google Scholar] [CrossRef]
- Ullah, H.M.A.; Elfadl, A.K.; Park, S.; Kim, Y.D.; Chung, M.-J.; Son, J.-Y.; Yun, H.-H.; Park, J.-M.; Yim, J.-H.; Jung, S.-J.; et al. Nogo-A is critical for pro-inflammatory gene regulation in myocytes and macrophages. Cells 2021, 10, 282. [Google Scholar] [CrossRef]
- Aziz, N.; Kim, M.-Y.; Cho, J.Y. Anti-inflammatory effects of luteolin: A review of in vitro, in vivo, and in silico studies. J. Ethnopharmacol. 2018, 225, 342–358. [Google Scholar] [CrossRef] [PubMed]
- Yu, T.; Yi, Y.-S.; Yang, Y.; Oh, J.; Jeong, D.; Cho, J.Y. The pivotal role of TBK1 in inflammatory responses mediated by macrophages. Mediat. Inflamm. 2012, 2012, 979105. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.H.; Park, J.G.; Sung, G.-H.; Yang, S.; Yang, W.S.; Kim, E.; Kim, J.H.; Ha, V.T.; Kim, H.G.; Yi, Y.-S.; et al. Kaempferol, a dietary flavonoid, ameliorates acute inflammatory and nociceptive symptoms in gastritis, pancreatitis, and abdominal pain. Mol. Nutr. Food Res. 2015, 59, 1400–1405. [Google Scholar] [CrossRef] [PubMed]
- Ullah, H.M.A.; Lee, Y.Y.; Kim, S.D.; Rhee, M.H. Duchesnea indica extract attenuates coal fly ash-induced inflammation in murine alveolar macrophages through the NF-Kappa B pathway. Evid.-Based Complement. Altern. Med. 2021, 2021, 5546052. [Google Scholar] [CrossRef]
- Cho, J.Y.; Baik, K.U.; Jung, J.H.; Park, M.H. In vitro anti-inflammatory effects of cynaropicrin, a sesquiterpene lactone, from Saussurea lappa. Eur. J. Pharmacol. 2000, 398, 399–407. [Google Scholar] [CrossRef]
- Lee, Y.Y.; Yang, W.K.; Han, J.E.; Kwak, D.; Kim, T.H.; Saba, E.; Kim, S.D.; Lee, Y.C.; Kim, J.S.; Kim, S.H.; et al. Hypericum ascyron L. extract reduces particulate matter-induced airway inflammation in mice. Phytother. Res. 2021, 35, 1621–1633. [Google Scholar] [CrossRef] [PubMed]
- Piao, C.H.; Fan, Y.; Nguyen, T.V.; Shin, H.S.; Kim, H.T.; Song, C.H.; Chai, O.H. PM2.5 exacerbates oxidative stress and inflammatory response through the Nrf2/NF-κB signaling pathway in OVA-induced allergic rhinitis mouse model. Int. J. Mol. Sci. 2021, 22, 8173. [Google Scholar] [CrossRef]
- Jiang, X.-Q.; Mei, X.-D.; Feng, D. Air pollution and chronic airway diseases: What should people know and do? J. Thorac. Dis. 2016, 8, E31–E40. [Google Scholar] [CrossRef]
- Ansari, S.; Hosseinzadeh, H.; Dennis, S.; Zwar, N. Activating primary care COPD patients with multi-morbidity through tailored self-management support. NPJ Prim. Care Respir. Med. 2020, 30, 12. [Google Scholar] [CrossRef] [PubMed]
- Zanobetti, A.; Franklin, M.; Koutrakis, P.; Schwartz, J. Fine particulate air pollution and its components in association with cause-specific emergency admissions. Environ. Health 2009, 8, 58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewtas, J. Air pollution combustion emissions: Characterization of causative agents and mechanisms associated with cancer, reproductive, and cardiovascular effects. Mutat. Res./Rev. Mutat. Res. 2007, 636, 95–133. [Google Scholar] [CrossRef]
- Shi, J.; Chen, R.; Yang, C.; Lin, Z.; Cai, J.; Xia, Y.; Wang, C.; Li, H.; Johnson, N.; Xu, X.; et al. Association between fine particulate matter chemical constituents and airway inflammation: A panel study among healthy adults in China. Environ. Res. 2016, 150, 264–268. [Google Scholar] [CrossRef]
- Deng, Q.; Deng, L.; Miao, Y.; Guo, X.; Li, Y. Particle deposition in the human lung: Health implications of particulate matter from different sources. Environ. Res. 2019, 169, 237–245. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Huang, J.; Wang, L.; Chen, C.; Yang, D.; Jin, M.; Bai, C.; Song, Y. Urban particulate matter triggers lung inflammation via the ROS-MAPK-NF-κB signaling pathway. J. Thorac. Dis. 2017, 9, 4398–4412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ullah, H.M.A.; Kim, J.; Rehman, N.U.; Kim, H.-J.; Ahn, M.-J.; Chung, H.J. A simple and sensitive liquid chromatography with tandem mass spectrometric method for the simultaneous determination of anthraquinone glycosides and their aglycones in rat plasma: Application to a pharmacokinetic study of rumex acetosa extract. Pharmaceutics 2018, 10, 100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ullah, H.M.A.; Zaman, S.; Juhara, F.; Akter, L.; Tareq, S.M.; Masum, E.H.; Bhattacharjee, R. Evaluation of antinociceptive, in-vivo & in-vitro anti-inflammatory activity of ethanolic extract of Curcuma zedoaria rhizome. BMC Complement. Altern. Med. 2014, 14, 346. [Google Scholar] [CrossRef] [Green Version]
- Katiyar, C.; Gupta, A.; Kanjilal, S.; Katiyar, S. Drug discovery from plant sources: An integrated approach. Ayu 2012, 33, 10–19. [Google Scholar] [CrossRef]
- Park, S.; Nhiem, N.X.; Park, J.H.; Bhattarai, K.R.; Chae, H.-J.; Kim, H.-R.; Kim, S.H. Isolation of amylase regulators from the leaves of Ixeridium dentatum. Nat. Prod. Res. 2021, 35, 744–749. [Google Scholar] [CrossRef]
- Sanjeewa, K.K.A.; Jayawardena, T.U.; Kim, S.-Y.; Lee, H.G.; Je, J.-G.; Jee, Y.; Jeon, Y.-J. Sargassum horneri (Turner) inhibit urban particulate matter-induced inflammation in MH-S lung macrophages via blocking TLRs mediated NF-κB and MAPK activation. J. Ethnopharmacol. 2020, 249, 112363. [Google Scholar] [CrossRef]
- Crosswhite, P.; Sun, Z. Nitric oxide, oxidative stress and inflammation in pulmonary arterial hypertension. J. Hypertens. 2010, 28, 201–212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clancy, R.M.; Abramson, S.B. Nitric oxide: A novel mediator of inflammation. Proc. Soc. Exp. Biol. Med. 1995, 210, 93–101. [Google Scholar] [CrossRef] [PubMed]
- Tuteja, N.; Chandra, M.; Tuteja, R.; Misra, M.K. Nitric oxide as a unique bioactive signaling messenger in physiology and pathophysiology. J. Biomed. Biotechnol. 2004, 2004, 498591. [Google Scholar] [CrossRef] [PubMed]
- Yu, Q.; Zeng, K.; Ma, X.; Song, F.; Jiang, Y.; Tu, P.; Wang, X. Resokaempferol-mediated anti-inflammatory effects on activated macrophages via the inhibition of JAK2/STAT3, NF-κB and JNK/p38 MAPK signaling pathways. Int. Immunopharmacol. 2016, 38, 104–114. [Google Scholar] [CrossRef] [PubMed]
- Liou, C.-J.; Len, W.-B.; Wu, S.-J.; Lin, C.-F.; Wu, X.-L.; Huang, W.-C. Casticin inhibits COX-2 and iNOS expression via suppression of NF-κB and MAPK signaling in lipopolysaccharide-stimulated mouse macrophages. J. Ethnopharmacol. 2014, 158, 310–316. [Google Scholar] [CrossRef] [PubMed]
- Herath, K.H.I.N.M.; Kim, H.J.; Lee, J.H.; Je, J.G.; Yu, H.-S.; Jeon, Y.-J.; Kim, H.J.; Jee, Y. Sargassum horneri (Turner) C. Agardh containing polyphenols attenuates particulate matter-induced inflammatory response by blocking TLR-mediated MYD88-dependent MAPK signaling pathway in MLE-12 cells. J. Ethnopharmacol. 2021, 265, 113340. [Google Scholar] [CrossRef]
- Ricciardolo, F.L.M.; Sterk, P.J.; Gaston, B.; Folkerts, G. Nitric oxide in health and disease of the respiratory system. Physiol. Rev. 2004, 84, 731–765. [Google Scholar] [CrossRef]
- Aquilano, K.; Baldelli, S.; Rotilio, G.; Ciriolo, M.R. Role of Nitric Oxide Synthases in Parkinson’s Disease: A Review on the Antioxidant and Anti-inflammatory Activity of Polyphenols. Neurochem. Res. 2008, 33, 2416–2426. [Google Scholar] [CrossRef]
- Korhonen, R.; Lahti, A.; Kankaanranta, H.; Moilanen, E. Nitric oxide production and signaling in inflammation. Curr. Drug Targets-Inflamm. Allergy 2005, 4, 471–479. [Google Scholar] [CrossRef]
- Kalayarasan, S.; Sriram, N.; Sudhandiran, G. Diallyl sulfide attenuates bleomycin-induced pulmonary fibrosis: Critical role of iNOS, NF-κB, TNF-α and IL-1β. Life Sci. 2008, 82, 1142–1153. [Google Scholar] [CrossRef] [PubMed]
- Minghetti, L. Cyclooxygenase-2 (COX-2) in inflammatory and degenerative brain diseases. J. Neuropathol. Exp. Neurol. 2004, 63, 901–910. [Google Scholar] [CrossRef] [Green Version]
- Ulivi, V.; Giannoni, P.; Gentili, C.; Cancedda, R.; Descalzi, F. p38/NF-κB-dependent expression of COX-2 during differentiation and inflammatory response of chondrocytes. J. Cell. Biochem. 2008, 104, 1393–1406. [Google Scholar] [CrossRef]
- Needleman, P.; Manning, P.T. Interactions between the inducible cyclooxygenase (COX-2) and nitric oxide synthase (iNOS) pathways: Implications for therapeutic intervention in osteoarthritis. Osteoarthr. Cartil. 1999, 7, 367–370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yasuda, T.; Kanno, M.; Kawamoto, M.; Yuge, O.; Ninomiya, Y. Suppression of inducible nitric oxide synthase and cyclooxygenase-2 gene expression by 22(R)-hydroxycholesterol requires de novo protein synthesis in activated macrophages. J. Steroid Biochem. Mol. Biol. 2005, 97, 376–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pierce, J.W.; Schoenleber, R.; Jesmok, G.; Best, J.; Moore, S.A.; Collins, T.; Gerritsen, M.E. Novel inhibitors of cytokine-induced IκBα phosphorylation and endothelial cell adhesion molecule expression show anti-inflammatory effects in vivo. J. Biol. Chem. 1997, 272, 21096–21103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zanotto-Filho, A.; Delgado-Cañedo, A.; Schröder, R.; Becker, M.; Klamt, F.; Moreira, J.C.F. The pharmacological NFκB inhibitors BAY117082 and MG132 induce cell arrest and apoptosis in leukemia cells through ROS-mitochondria pathway activation. Cancer Lett. 2010, 288, 192–203. [Google Scholar] [CrossRef]
- Lee, J.; Rhee, M.H.; Kim, E.; Cho, J.Y. BAY 11-7082 Is a Broad-Spectrum Inhibitor with Anti-Inflammatory Activity against Multiple Targets. Mediat. Inflamm. 2012, 2012, 416036. [Google Scholar] [CrossRef]
- Lee, D.; Lee, S.R.; Kang, K.S.; Kim, K.H. Bioactive phytochemicals from mulberry: Potential anti-inflammatory effects in lipopolysaccharide-stimulated RAW 264.7 macrophages. Int. J. Mol. Sci. 2021, 22, 8120. [Google Scholar] [CrossRef]
- Yoon, Y.P.; Lee, H.J.; Lee, D.-U.; Lee, S.K.; Hong, J.-H.; Lee, C.J. Effects of lupenone, lupeol, and taraxerol derived from Adenophora triphylla on the gene expression and production of airway MUC5AC mucin. Tuberc. Respir. Dis. 2015, 78, 210–217. [Google Scholar] [CrossRef]
- He, J.; Lu, X.; Wei, T.; Dong, Y.; Cai, Z.; Tang, L.; Liu, M. Asperuloside and asperulosidic acid Exert an anti-Inflammatory effect via suppression of the NF-κB and MAPK signaling pathways in LPS-Induced RAW 264.7 macrophages. Int. J. Mol. Sci. 2018, 19, 2027. [Google Scholar] [CrossRef] [Green Version]
- Liang, D.; Li, F.; Fu, Y.; Cao, Y.; Song, X.; Wang, T.; Wang, W.; Guo, M.; Zhou, E.; Li, D.; et al. Thymol inhibits LPS-stimulated inflammatory response via down-regulation of NF-κB and MAPK signaling pathways in mouse mammary epithelial cells. Inflammation 2014, 37, 214–222. [Google Scholar] [CrossRef]
- Panday, A.; Inda, M.E.; Bagam, P.; Sahoo, M.K.; Osorio, D.; Batra, S. Transcription factor NF-κB: An update on intervention strategies. Arch. Immunol. Ther. Exp. 2016, 64, 463–483. [Google Scholar] [CrossRef]
- Schmid, J.A.; Birbach, A. IκB kinase β (IKKβ/IKK2/IKBKB)—A key molecule in signaling to the transcription factor NF-κB. Cytokine Growth Factor Rev. 2008, 19, 157–165. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.-I.; Burckart, G.J. Nuclear factor kappa B: Important transcription factor and therapeutic target. J. Clin. Pharmacol. 1998, 38, 981–993. [Google Scholar] [CrossRef]
- Lang, R.; Hammer, M.; Mages, J. DUSP meet immunology: Dual specificity MAPK phosphatases in control of the inflammatory response. J. Immunol. 2006, 177, 7497–7504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.; Nhiem, N.X.; Lee, T.H.; Kim, N.; Kim, S.Y.; Chae, H.-J.; Kim, S.H. Isolation of two new bioactive sesquiterpene lactone glycosides from the roots of Ixeris dentata. Bioorganic Med. Chem. Lett. 2015, 25, 4562–4566. [Google Scholar] [CrossRef] [PubMed]
- Saba, E.; Irfan, M.; Jeong, D.H.; Ameer, K.; Lee, Y.Y.; Park, C.-K.; Hong, S.-B.; Rhee, M.H. Mediation of antiinflammatory effects of Rg3-enriched red ginseng extract from Korean Red Ginseng via retinoid X receptor α–peroxisome-proliferating receptor γ nuclear receptors. J. Ginseng Res. 2019, 43, 442–451. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| H | Primer Sequence |
|---|---|
| GAPDH * | F: 5′-CACTCACGGCAAATTCAACGGCAC-3′ R: 5′-GACTCCACGACATACTCAGCAC-3′ |
| iNOS * | F: 5′-CCCTTCCGAAGTTTCTGGCAGCAGC-3′ R: 5′-GGCTGTCAGAGCCTCGTGGCTTTGG-3′ |
| COX-2 * | F: 5′-CACTACATCCTGACCCACTT-3′ R: 5′-ATGCTCCTGCTTGAGTATGT-3′ |
| TNF-α * | F: 5′-TTGACCTCAGCGCTGAGTTG-3′ R: 5′-CCTGTAGCCCACGTCGTAGC-3′ |
| IL-1β * | F: 5′-CTGTGGAGAAGCTGTGGCAG-3′ R: 5′-GGGATCCACACTCTCCAGCT-3′ |
| IL-6 * | F: 5′-GTACTCCAGAAGACCAGAGG-3′ R: 5’-TGCTGGTGACAACCACGGCC-3′ |
| GAPDH ** | F: 5′-CACTCACGGCAAATTCAACGGCAC-3′ R: 5′-GACTCCACGACATACTCAGCAC-3′ |
| iNOS ** | F: 5′-GGCAGCCTGTGAGACCTTTG-3′ R: 5′-GCATTGGAAGTGAAGCGTTTC-3′ |
| COX-2 ** | F: 5′-GGGTGTCCCTTCACTTCTTTCA-3′ R: 5′-TGGGAGGCACTTGCATTGA-3′ |
| TNF-α ** | F: 5′-TGCCTATGTCTCAGCCTCTTC-3′ R: 5′-GAGGCCATTTGGGAACTTCT-3′ |
| IL-1β ** | F: 5′-CAACCAACAAGTGATATTCTCCATG-3′ R: 5′-GATCCACACTCTCCAGCTGCA-3′ |
| IL-6 ** | F: 5′-TCCAGTTGCCTTCTTGGGAC-3′ R: 5′-GTGTAATTAAGCCTCCGACTTG-3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ullah, H.M.A.; Kwon, T.-H.; Park, S.; Kim, S.D.; Rhee, M.H. Isoleucilactucin Ameliorates Coal Fly Ash-Induced Inflammation through the NF-κB and MAPK Pathways in MH-S Cells. Int. J. Mol. Sci. 2021, 22, 9506. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22179506
Ullah HMA, Kwon T-H, Park S, Kim SD, Rhee MH. Isoleucilactucin Ameliorates Coal Fly Ash-Induced Inflammation through the NF-κB and MAPK Pathways in MH-S Cells. International Journal of Molecular Sciences. 2021; 22(17):9506. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22179506
Chicago/Turabian StyleUllah, H. M. Arif, Tae-Hyung Kwon, SeonJu Park, Sung Dae Kim, and Man Hee Rhee. 2021. "Isoleucilactucin Ameliorates Coal Fly Ash-Induced Inflammation through the NF-κB and MAPK Pathways in MH-S Cells" International Journal of Molecular Sciences 22, no. 17: 9506. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22179506