
Interleukin-21 in Viral Infections


Department of Immunology, Faculty of Medicine, Yamagata University, 2-2-2 Iida-nishi, Yamagata City 990-9585, Japan
Int. J. Mol. Sci. 2021, 22(17), 9521; https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22179521
Submission received: 29 July 2021
/
Revised: 24 August 2021
/
Accepted: 30 August 2021
/
Published: 1 September 2021
(This article belongs to the Special Issue The Role of Interleukin in Health and Diseases)
{kind=link}
{kind=link}
Abstract
:Interleukin (IL)-21 is a cytokine that affects the differentiation and function of lymphoid and myeloid cells and regulates both innate and adaptive immune responses. In addition to regulating the immune response to tumor and viral infections, IL-21 also has a profound effect on the development of autoimmune and inflammatory diseases. IL-21 is produced mainly from CD4+ T cells—in particular, follicular helper T (Tfh) cells—which have a great influence on the regulation of antibody production. It is also an important cytokine for the activation of CD8+ T cells, and its role in recovering the function of CD8+ T cells exhausted by chronic microbial infections and cancer has been clarified. Thus, IL-21 plays an extremely important role in viral infections, especially chronic viral infections. In this review, I will introduce the findings to date on how IL-21 is involved in some typical viral infections and the potential of treating viral diseases with IL-21.
1. Introduction
In 2000, a new type 1 cytokine family receptor, named the IL-21 receptor, was discovered through a database search [1,2]. At the same time, a novel cytokine, Interleukin (IL)-21, was identified as a ligand that binds to the IL-21 receptor [1]. The IL-21 receptor forms a heterodimer together with the common cytokine receptor γ chain (γc chain) and activates STAT3, mainly as a signaling molecule by stimulation with IL-21 [3,4]. The γc chain, originally identified as the IL-2 receptor γ chain in 1992 [5], was subsequently found to be shared by other cytokines, IL-4, IL-7, IL-9, IL-15 and IL-21, in addition to IL-2. Accordingly, it is called the γc chain [6]. The γc chain was found to be the causative gene of X-linked severe combined immunodeficiency (X-SCID) [7,8]. In X-SCID patients, in addition to impaired differentiation of T cells and NK cells, B cell dysfunction is considered to be caused by impaired signal transduction from IL-21 and IL-4 receptors [9].
IL-21 is produced from activated CD4+ T cells and NKT cells [1] and is thought to have various functions against various immune cells [10]. Later, it was found that IL-21 is produced from two types of activated CD4+ T cells. One is type 17 helper T (Th17) cells, whose differentiation has been reported to be affected by IL-21 [11,12,13,14,15,16,17]. The other type of activated CD4+ T cells are follicular helper T (Tfh) cells, and the IL-21 produced from these cells also plays an important role in Tfh cell differentiation and germinal center formation [18,19,20]. However, subsequent studies suggest that IL-6 and ICOSL, a co-stimulatory molecule, are essential for Tfh cell differentiation, and that IL-21 may play an auxiliary role [21,22].
IL-21 is a major cytokine produced by Tfh cells along with IL-4 and CXCL13, and IL-21, IL-4 and CD40L are major stimulators for the regulation of antibody production by T-cell-dependent activated B cells in the germinal center [22]. Affinity maturation, which is based on somatic hypermutation, and class switch recombination of antibody genes in germinal centers are extremely important for the production of high-affinity antibodies and the maintenance of memory B cells, which are essential for protecting against viral infections [23]. Meanwhile, IL-21 induces apoptosis in bystander B cells activated without antigen receptor stimulation [10]. Thus, it has become clear that IL-21 has important functions, such as controlling class switching on B cells and promoting final differentiation into plasma cells (Figure 1). Studies using gene-deficient mice showed that IL-21 promotes IgG1 class switching together with IL-4, but, unlike IL-4, it was found to suppress class switching to IgE [9,24]. In addition, in an in vitro study using human B cells, IL-21 promoted class switching to IgG1 and IgG3 [25]. Subsequent studies have shown that a deficiency of STAT3, the major signaling molecule from the IL-21 receptor, causes hyper-IgE syndrome, which has an autosomal inheritance pattern [26,27]. In addition, it was found that increased IgE was observed in approximately half of patients with an IL-21 receptor deficiency, although the level of immunoglobulin itself decreased [28]. These results suggest that IL-21 enhances the class switch to IgG1 and IgG3 and suppresses the class switch to IgE in humans. In addition, in studies using IL-21 or IL-21 receptor gene-deficient mice, it was reported that IL-21 induces somatic hypermutation in B cells and is involved in the enhanced affinity of antibodies [29,30].
IL-21 is not essential for the development of CD8+ T cells but is known to be involved in the proliferation and activation of CD8+ T cells and immunological memory formation, together with other γc cytokines such as IL-7 and IL-15 [31,32,33,34,35] (Figure 2). It has also been shown that IL-21 is involved in the differentiation of CD8+ effector memory T cells in Listeria infection [36]. Persistent viral infection exhausts activated CD8+ T cells [37,38,39], but even exhausted CD8+ T cells retain the ability to control chronic viral infections [40,41]. Therefore, the prevention of CD8+ T cell exhaustion and the recovery of exhausted CD8+ T cells are important for the treatment of chronic viral infections and tumors. Over the past 10 years, immune checkpoint inhibition therapy that inhibits the PD-L1-PD-1 and CD28-CTLA4 system has come into focus as an immunotherapy for tumors, targeting exhausted T cells, and is actively used as the fourth treatment method for tumors. In recent years, IL-21 was reported to maintain an antiviral immune response by maintaining CD8+ effector T cells and suppressing CD8+ T cell exhaustion [42]. In the future, it is expected that a treatment method combining an immune checkpoint inhibitor and IL-21 cytokine therapy will be developed as an approach to immunotherapy for chronic viral infections and tumors.
Given the outcomes of these different studies, IL-21 is considered to be a very important cytokine that is key to the antiviral immune response in many respects, including Tfh-mediated regulation of antibody function and activation of CD8+ T cells. This review summarizes recent findings regarding the role of IL-21 in viral infection and treatment.
2. Vaccinia Virus and Lymphocytic Choriomeningitis Virus (LCMV) Infection
In general, IL-21 regulates the immune response to viral infections via CD4+ T cells, B cells and CD8+ T cells. While there are reports that IL-21 has an important function in the survival of CD8 + T cells against acute vaccinia virus infection [33,35], there is also a report suggesting that IL-21 is not required [43]. On the other hand, in chronic LCMV infections, IL-21 produced by CD4+ T cells has been shown to maintain the antiviral activity of CD8+ T cells and regulate CD8+ T cell exhaustion [43,44,45]. Signal transduction from the IL-21 receptor activates Bcl-6 and Blimp-1 via STAT3-SOCS3 together with IL-10 and promotes the differentiation of CD8+ memory T cells during chronic LCMV infection [46]. It has also been reported that IL-21-induced basic leucine zipper transcription factor ATF-like (BATF) induces Blimp1 expression together with IRF4 and is important for maintaining the effector function of CD8+ T cells [47].
In recent years, Zander et al. reported that IL-21 produced from CD4+ T cells promotes the maintenance of CX3CR1+ CD8+ T cells, which are considered to be CD8+ effector T cells to LCMV infection, and its differentiation from progenitor cells [42]. Furthermore, IL-21 suppresses the exhaustion of CD8+ T cells to maintain the immune response against chronic viral infections. PD-1/PD-L1 checkpoint inhibition therapy partially suppresses the appearance of exhausted T cells in patients with chronic infections and cancer, but it has also been shown that the development of exhausted CD8+ T cells during chronic LCMV infection could not be reverted, even by PD-1/PD-L1 checkpoint inhibition, when the production of IL-21 by CD4+ T cells was inadequate [42]. In the future, treatment that combines immune checkpoint inhibition therapy and cytokine therapy with IL-21 may be developed for chronic viral infection and cancer. Regarding the mechanism by which IL-21 inhibits differentiation into exhausted T cells, Loschinski et al. showed in an in vitro study that IL-21 induces fatty acid oxidation and oxidative phosphorylation in mitochondria to induce memory T cell differentiation and to inhibit exhausted T cell differentiation [48].
3. HIV Infection
In HIV patients, serum IL-21 levels decrease with a decrease in CD4+ T cells [49,50]. IL-21 levels were higher in patients who could control HIV compared to those who could not, and it was shown to IL-21 restore the antiviral activity of CD8+ T cells, similar to that of LCMV infection [49,50,51]. It has also been shown that in patients who cannot control HIV, serum IL-21 is reduced and IL-21 receptor expression on CD8+ T cells is increased [52]. Thus, serum IL-21 concentration and IL-21 receptor expression on CD8+ T cells can serve as markers for understanding the viral control status of HIV patients. Interestingly, in HIV patients with a well-controlled viral load, HIV-specific CD8+ T cells also produce IL-21 in a CD4+ T-cell-independent manner [53,54]. Because IL-21 is also known to be produced by CD8+ T cells in patients with other autoimmune diseases [55,56,57], its association with the pathophysiology of HIV infection is interesting.
HIV infection affects the function of B cells, such as decreased BCR reactivity and proliferative capacity, and decreased CD21 expression [58,59]. In particular, memory B cell abnormalities are prominent [60], causing insufficient antibody production in HIV patients [61]. To control HIV infection, the production of broadly neutralizing antibodies (bnAbs) is important. Twenty percent of HIV patients can produce these bnAbs [62], and these antibodies have a neutralizing ability over 70% of the currently prevalent HIV strains [63,64,65,66]. However, it is unclear why such antibodies are produced only in some affected individuals.
An important point in developing a vaccine for HIV infection is the creation of bnAbs that are widely aware of HIV. Clinical trials of HIV vaccines under several protocols have progressed; however, only the clinical trials of HIV vaccines conducted in Thailand (RV144 trial) have shown limited activity in preventing HIV infection [67]. Consequently, attention has been focused on the involvement of HIV-specific CD4+ T cells induced by the vaccine used in the RV144 study [68,69,70,71]. In addition, because the protocols used in clinical trials conducted so far are varied, such as vaccine strategy, administration method, boost method, etc., it is important to examine how CD4+ T cells react under these conditions, especially regarding the responsiveness of Tfh cells [72].
Tfh cells are the preferred target of HIV infection and are known to be a reservoir for HIV replication [73,74,75]. In contrast, Tfh cells have been reported to proliferate even under chronic HIV infection [73,76,77]. These facts suggest that HIV infection may cause abnormalities in Tfh cells, resulting in impaired helper function in B cells. Colineau’s and Graff-Dubois’ groups analyzed the splenocytes of HIV patients and found that although Tfh cells were increased in HIV patients, cytokines such as IL-4 and IL-10 and co-stimulatory molecule expression decreased, and follicular regulatory T (Tfr) cells increased. These results show that Tfh cells in HIV patients cannot achieve normal B cell activation [78,79]. On the other hand, IL-21 is also a major cytokine produced by Tfh. There are conflicting reports that IL-21 increases in HIV patients [78] and decreases or does not change in HIV patients or SIV-infected macaques [80,81]. There seems to be a difference in the amount of IL-21 produced depending on the infection status and the sample.
Morita et al. found that CD4+ CXCR5+ T cells present in human peripheral blood show a similar phenotype to Tfh cells and stimulate B cells via IL-21 [82]. In the case of human physiology, it is usually difficult to examine Tfh cell status, so analysis of the Tfh cells’ counterpart present in the periphery has been actively conducted. Locci et al. reported that PD1+ CXCR3− CXCR5+ memory Tfh cells present in human peripheral blood have the most differentiation-inducing function for B cells, and their peripheral frequency correlates with bnAbs production in HIV patients [83]. Furthermore, Tfh cells, which produce IL-21 in peripheral blood, were decreased in HIV patients, and anti-HIV treatment restored peripheral Tfh cells [80]. These studies show that measuring peripheral Tfh cells in blood can estimate the status of Tfh cells in lymphoid tissues against HIV infection, providing an important indicator of HIV vaccine development. In addition, Yamamoto et al. also reported that PD1+ CXCR5+ Tfh cells are important as CD4-positive T cells were involved in bnAb production in an in vivo study using SHIVAD8-infected macaques [84].
Furthermore, Schultz et al. found that HIV-specific IL-21-producing CD4+ T cells in peripheral blood are Tfh cells circulating in peripheral blood, according to their analysis of gene expression patterns and phenotypes, and that these Tfh cells are increased in the RV144 study compared to other ineffective vaccine trials [85]. In addition, HIV-specific IL-21-producing peripheral Tfh cells correlate well with virus control in HIV patients; moreover, in poorly controlled HIV patients, peripheral Tfh cells are reprogrammed to the Th1 cell side by IL-2 signaling, resulting in decreased expression of Bcl6, CXCR5 and IL-21 [86,87]. In actual HIV infection, bnAbs are not acquired in the early stages and appear in some affected individuals in the later stages of HIV infection [88]. Although the details of the mechanism have not been clarified, it has been shown that IL-21 produced by Tfh cells plays an extremely important role in HIV-specific bnAb production [85]. If Tfh cells can properly produce IL-21 during HIV infection, bnAbs capable of controlling HIV may be produced.
In contrast, unlike bnAbs, antibodies that do not have neutralizing activity but cause innate immune activation in an Fc-dependent manner are elevated from the early stages of HIV infection and play an important role in HIV infection [89,90,91]. These antibodies are thought to confer resistance towards HIV infection through complement activation, opsonization and antibody-dependent cellular cytotoxicity (ADCC). Pusnik et al. reported that anti-HIV env-specific antibodies are produced from Blimp-1-positive B cells in an IL-21-producing CD4+ env-specific T cell-dependent manner, and that these antibodies are involved in Fc-dependent innate immune response activation [92]. It became clear that IL-21 produced from Tfh cells also plays an extremely important role in the production of anti-HIV antibodies that activate the innate immune response to HIV, further recognizing the importance of IL-21.
IL-21 also acts directly on HIV-infected CD4+ T cells and directly suppresses HIV replication by activating microRNA-29 via STAT3 [93]. This microRNA-29 activation by IL-21 shows strong natural resistance to HIV infection [94].
As described above, IL-21 resists HIV infection by activating CD8+ T cells, producing bnAbs and non-neutralizing innate immune-active antibodies and by directly suppressing HIV replication in CD4+ T cells. Animal experiments with SIV infection are being conducted to investigate the development of HIV therapy using IL-21. In SIV-infected rhesus macaques, the cytotoxic activity of SIV-specific CD8+ T cells decreases from the acute phase to the chronic phase, and dysbiosis and transition to the body of intestinal bacteria occurs due to a decrease in Th17 cells in the intestinal tract [95,96,97,98]. In addition, Micci et al. reported that IL-21-producing CD4+ T cells decreased with SIV infection, resulting in a decrease in Th17 cells [99]. Knowing that CD8+ T cells and Th17 cells are activated by IL-21, in vivo experiments using IL-21 have been conducted targeting these cells. In an experiment in which IL-21 was administered to rhesus macaques during the SIV chronic infection phase, the cytotoxic activity of NK cells and CD8+ T cells was enhanced, the anti-SIV antibody was increased, and IL-21 treatment against rhesus macaques during the SIV acute infection phase, the antiviral activity of CD8+ T cells and Th17 cells in the intestinal tract were maintained, but these treatments did not reduce the virus counts in either phase [100,101,102]. On the other hand, IL-21 exhibits various functions against NK cells. IL-21 promotes the proliferation of CD16+ NK cells and activates exhausted NK cells, while it has been also reported to limit the response of NK cells [1,31,103]. African green monkeys are non-susceptible to SIV infection, partly because their NK cells have been shown to undergo final differentiation in response to SIV infection and acquire antiviral activity [104]. Therefore, IL-21 may control the antiviral activity of NK cells. In an SIV infection study of rhesus macaques as a susceptible host, Harper et al. observed the promotion of the final differentiation of NK cells into NKG2alowCD16+ NK cells by a combination therapy of IL-21 and IFN-α in addition to antiretriviral therapy (ART), and it has been reported that the amount of SIV virus in the lymph nodes is reduced [105]. As will be described later, IL-21 has been clinically tested for malignant tumors using the activation of CD8+ T cells by IL-21, but this has not been successful due to side reactions such as liver damage. If side reactions can be controlled by limiting the method, duration and amount of IL-21 administration, stronger HIV treatment may be possible through regulation of the activity of various immune cells. In addition, by successfully administering IL-21 in parallel with vaccination, it may be possible to develop a vaccine that induces bnAbs through the appropriate activation of Tfh cells.
4. HBV Infection
Despite the availability of effective vaccines against HBV, there are still approximately 240 million chronic HBV infections worldwide [106]. Although HBV is often eliminated and cured in adults, infections in newborns and young people are much more likely to develop into chronic hepatitis than those in adults. The mechanism of chronicity, depending on the time of infection, is largely unknown. In a study using a mouse model of HBV infection, Publicover et al. showed that the immune system of adult mice produces more IL-21 than young mice [107]. Adult mice can eliminate viral antigens, whereas young mice cannot. IL-21 receptor-deficient mice also cannot eliminate viral antigens in the same way as young mice. Therefore, IL-21 is quite important for the anti-HBV immune response, suggesting that young patients may have reduced IL-21-mediated HBV-specific CD8 + T and B cell responsiveness. In human HBV hepatitis, there are more IL-21-producing CD4+ T cells in acute hepatitis than in chronic hepatitis, and the number is positively correlated with the number of HBV-specific CD8+ T cells [108]. In addition, virus elimination was significantly promoted in patients with elevated serum IL-21 levels 12 weeks after antiviral treatment [109]. Similar to other viral infections, IL-21 function is thought to be primarily focused on the recovery of exhausted CD8+ T cells [110].
Shen et al. used a mouse model to compare the immune response to persistently infectious HBV strains and non-persistently infectious HBV strains, and they discovered that IL-21 and IL-33 are induced in non-persistently infectious HBV strain-infected mice, whereas the induction does not occur in persistently infectious HBV strain-infected mice [111]. Therefore, by forcibly expressing IL-21 in hepatocytes, persistently infectious HBV strains were eliminated in a CD8+ T-cell-dependent manner [112,113]. It has also been shown that persistent infection with HBV can be prevented in mice in which IL-21 is transiently expressed in hepatocytes [114]. Thus, IL-21 is expected to enable the treatment and prevention of HBV infection via CD8+ T cells.
Appropriate antiviral antibody production is also an important antiviral defense response against HBV infections. However, with the transition to chronic hepatitis, exhaustion of CD4+ T cells is as prominent as the exhaustion of CD8+ T cells [115]. IL-21 produced from Tfh cells plays an important role in proper antibody production by B cells [10]. Interestingly, although IL-21 production from Tfh cells was significantly reduced in chronic HCV infections, antibody production by B cells was maintained [116]. This indicates that Tfh cells function IL-21-independently for the humoral immune response. Recently, Khanam et al. have reported that in chronic HBV infections, HBsAg-specific Tfh cells produce IL-27 and support the humoral immune response despite a deficiency in IL-21 production [117] (Figure 1). In the future, the difference in function between IL-21 and IL-27 in B cell differentiation will become clearer.
In contrast, IL-21 is also expected to exacerbate liver cirrhosis due to HBV infection [118]. Some patients with chronic HBV infection develop liver cirrhosis and cancer. Liver fibrosis that causes liver cirrhosis is caused by the production of α-smooth muscle actin (α-SMA) and collagen by the activation of hematopoietic stem cells (HSCs). Th17 cells are increased in liver cirrhosis [119,120] and are probably the source of increased serum IL-21 levels. Increased IL-21 may activate HSCs and exacerbate liver cirrhosis [121].
In this way, for HBV infection, IL-21 works to eliminate HBV in acute infection, but it is also involved in the transition from chronic hepatitis to liver cirrhosis, so great care should be taken when using IL-21 as a therapeutic agent [122].
5. HCV Infection
HCV causes chronic infection in 65–85% of affected people, and there are approximately 180 million chronic HCV patients worldwide [123]. Similar to HBV, IL-21’s function against HCV infection is thought to be mainly based on the recovery of exhausted CD8+ T cells [124]. It is clear that CD8+ T cells and CD4+ T cells play an important role in acute HCV infection [125,126].
In individuals in which neutralizing antibodies appear early in infection, spontaneous healing can be expected, whereas in individuals in which neutralizing antibodies appear late, HCV is persistently infected and induces chronic hepatitis [127,128,129]. Examining peripherally circulating Tfh cells instead of the Tfh cells in lymphoid tissues in patients with acute HCV infection, the number correlates with the amount of anti-HCV antibody, and when HCV is removed by anti-HCV drug treatment, the number of peripheral Tfh cells increases [130,131]. Salinas et al. reported that in HCV patients who heal spontaneously, peripheral Tfh cells that produce IL-21 appear early in infection, followed by anti-HCV E2 glycoprotein-specific antibodies [132].
6. Polyomavirus Infection
In human JC polyomavirus infection, when CD4+ T cells decrease due to AIDS, progressive multifocal leukoencephalopathy (PML) may develop as a brain lesion. Because the onset of PML is suppressed by JC polyomavirus-specific CD8+ T cells and CD4+ T cells, the helper function of CD4+ T cells against CD8+ T cells is considered to be important. Tissue-resident memory CD8+ T (CD8+ Trm) cells play an essential role in the immune response to polyomavirus infection according to a mouse central nervous system infection model. For the development and maintenance of CD8+ Trm cells, CD4+ T cells are required [133,134]; in particular, CXCR5hi PD-1hi CD4+ Tfh-like T cells produce IL-21 in the brain and are involved in the induction of CD8+ Trm cell differentiation [135] These CD4+ T cells express Tbet and Blimp1 and have been shown to possess both Th1 and Tfh cell traits.
7. SARS-CoV-2 Infection
Shortly after the first report of a new coronavirus infection in December 2019, the virus quickly spread worldwide. The acute respiratory disease caused by SARS-CoV-2, namely COVID-19, has caused many deaths worldwide [136,137]. Infection with SARS-CoV-2 activates the innate and adaptive immune system that responds to viral infections; some individuals recover without actual onset of the disease, whereas others develop severe symptoms and die. This heterogeneity is a major feature of COVID-19, and it is thought that various factors produce this heterogeneity in the innate and adaptive immune systems [138,139].
With respect to the acquired immune response during the acute and convalescent phases, many affected individuals produce IgG antibodies against the receptor-binding domain of the viral spike protein. These antibodies are important in preventing viral infections and are found to last for at least 6 months. In addition, spike protein-specific memory B cells appeared at the early stage of infection and further increased 6 months after infection [140,141,142]. Dan et al. showed that virus spike-specific CD4+ T cells and CD8+ T cells are halved in number 3–5 months after infection and that each component of each immune response shows a different trend, and there are individual differences in these trends [142].
The frequency of SARS-CoV-2-specific CD4+ T cells or CCR6− peripheral Tfh cells in COVID-19 patients has been shown to correlate with virus-neutralizing antibody levels [143,144]. In addition, it has been reported that spike protein-specific memory B cell differentiation and antibody affinity maturation in mildly recovered COVID-19 and severely ill patients correlate well with the function of IL-21-producing CD4+ T cells [145]. These results show that in SARS-CoV-2 infection, similar to other viral infections, Tfh cell-induced IL-21-mediated germinal center B cell differentiation is crucial for the production of useful virus-neutralizing antibodies. As a result of analyzing B cells that differentiated in the germinal center of patients with severe COVID-19, IgG2-producing B cells that were affected by type 1 IFN at the beginning of ICU hospitalization were dominant; however, the number of IgG1- and IgA1-producing B cells affected by IL-21 and TGF-β increased as the disease progressed, and eventually, the number of IgA2-producing B cells strongly affected by TGF-β increased [146]. Furthermore, IgA2 affected by TGF-β in patients with severe COVID-19 showed unstable SARS-CoV-2 recognition. TGF-β causes pulmonary fibrosis, which is the most avoidable complication in the chronic post-inflammatory phase of SARS-CoV-2 infection. In severe COVID-19, IL-21 and TGF-β may be imbalanced, and the pathological condition is exacerbated. The development of therapeutic methods targeting these cytokines is expected in the future. A treatment method aimed at activating NK cells and CD8+ T cells using IL-15 for COVID-19 has been proposed [147], and Wilz argued that IL-21 should be added to this treatment [148]. If the heterogeneity of the immune response to SARS-CoV-2 infection becomes more clearly understood in the future, the development of new therapeutic methods targeting cytokines will be further promoted.
8. Clinical Application of IL-21
Initially, IL-21 was clinically tested for malignant melanoma, acute myeloid leukemia, metastatic renal cancer, etc., using the antitumor activity of the activation of CD8-positive T cells [149]. Some effects on tumors were observed in Phase 1 and Phase 2 clinical trials, but many were discontinued due to side reactions such as liver damage [150]. A clinical trial combining anti-EGF receptor antibody and IL-21 for metastatic colorectal cancer was also conducted, achieving stable disease in 60% of patients and observing immune cell activation [151]. However, clinical studies were discontinued when various side reactions occurred, and IL-21 was reported to be involved in the development of inflammatory bowel disease and subsequent colorectal tumors [152,153,154].
9. Concluding Remarks
IL-21 is a multifunctional cytokine that acts on various immune cells. In particular, the proliferation and activation of CD8+ T cells, the differentiation and maintenance of CD8+ Trm cells and the activation of exhausted CD8+ T cells are considered to be some of the most important functions of IL-21 as an immune response to resist viral infections, especially chronic viral infections. On the other hand, IL-21 is a major cytokine produced by Tfh cells, and it has been clarified that it has an effect of promoting optimal antibody production on B cells through class switch recombination and somatic hypermutation. Thus, IL-21 plays an extremely important role, namely the regulation of antibody production for the early convergence of acute viral infections and the recovery of exhausted T cells to resist chronic infections. Evidently, IL-21 can potentially be utilized in vaccine development for the production of optimal neutralizing antibodies and the induction of T cell immune memory. However, clinical trials of IL-21 aimed at antitumor effects have been unsuccessful, and it has been found that its clinical application is not easy. In the future, I hope that treatment and vaccine development research for viral infections using the functions of IL-21 will progress, such as the development of therapeutic strategies that limit the action to immune cells such as B cells and CD8+ T cells.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
I declare no conflict of interest.
Abbreviations
| IL | interleukin |
| Tfh | follicular helper T |
| Th17 | type 17 helper T |
| LCMV | lymphocytic choriomeningitis virus |
| bnAbs | broadly neutralizing antibodies |
| Tfr | follicular regulatory T |
| CD8+ Trm | CD8+ tissue-resident memory T |
References
- Parrish-Novak, J.; Dillon, S.R.; Nelson, A.; Hammond, A.; Sprecher, C.; Gross, J.A.; Johnston, J.; Madden, K.; Xu, W.; West, J.; et al. Interleukin 21 and its receptor are involved in NK cell expansion and regulation of lymphocyte function. Nature 2000, 408, 57–63. [Google Scholar] [CrossRef]
- Ozaki, K.; Kikly, K.; Michalovich, D.; Young, P.R.; Leonard, W.J. Cloning of a type I cytokine receptor most related to the IL-2 receptor beta chain. Proc. Natl. Acad. Sci. USA 2000, 97, 11439–11444. [Google Scholar] [CrossRef] [Green Version]
- Asao, H.; Okuyama, C.; Kumaki, S.; Ishii, N.; Tsuchiya, S.; Foster, D.; Sugamura, K. Cutting edge: The common γ-chain is an indispensable subunit of the IL-21 receptor complex. J. Immunol. 2001, 167, 1–5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Habib, T.; Senadheera, S.; Weinberg, K.; Kaushansky, K. The common γ chain (γ c) is a required signaling component of the IL-21 receptor and supports IL-21-induced cell proliferation via JAK3. Biochemistry 2002, 41, 8725–8731. [Google Scholar] [CrossRef] [PubMed]
- Takeshita, T.; Asao, H.; Ohtani, K.; Ishii, N.; Kumaki, S.; Tanaka, N.; Munakata, H.; Nakamura, M.; Sugamura, K. Cloning of the γ chain of the human IL-2 receptor. Science 1992, 257, 379–382. [Google Scholar] [CrossRef] [PubMed]
- Sugamura, K.; Asao, H.; Kondo, M.; Tanaka, N.; Ishii, N.; Nakamura, M.; Takeshita, T. The common γ-chain for multiple cytokine receptors. Adv. Immunol. 1995, 59, 225–277. [Google Scholar] [CrossRef]
- Noguchi, M.; Yi, H.; Rosenblatt, H.M.; Filipovich, A.H.; Adelstein, S.; Modi, W.S.; McBride, O.W.; Leonard, W.J. Interleukin-2 receptor γ chain mutation results in X-linked severe combined immunodeficiency in humans. Cell 1993, 73, 147–157. [Google Scholar] [CrossRef]
- Leonard, W.J. Cytokines and immunodeficiency diseases. Nat. Rev. Immunol. 2001, 1, 200–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ozaki, K.; Spolski, R.; Feng, C.G.; Qi, C.F.; Cheng, J.; Sher, A.; Morse, H.C., 3rd; Liu, C.; Schwartzberg, P.L.; Leonard, W.J. A critical role for IL-21 in regulating immunoglobulin production. Science 2002, 298, 1630–1634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spolski, R.; Leonard, W.J. Interleukin-21: Basic biology and implications for cancer and autoimmunity. Annu. Rev. Immunol. 2008, 26, 57–79. [Google Scholar] [CrossRef] [Green Version]
- Fantini, M.C.; Rizzo, A.; Fina, D.; Caruso, R.; Becker, C.; Neurath, M.F.; Macdonald, T.T.; Pallone, F.; Monteleone, G. IL-21 regulates experimental colitis by modulating the balance between Treg and Th17 cells. Eur. J. Immunol. 2007, 37, 3155–3163. [Google Scholar] [CrossRef] [PubMed]
- Korn, T.; Bettelli, E.; Gao, W.; Awasthi, A.; Jäger, A.; Strom, T.B.; Oukka, M.; Kuchroo, V.K. IL-21 initiates an alternative pathway to induce proinflammatory T(H)17 cells. Nature 2007, 448, 484–487. [Google Scholar] [CrossRef] [PubMed]
- Nurieva, R.; Yang, X.O.; Martinez, G.; Zhang, Y.; Panopoulos, A.D.; Ma, L.; Schluns, K.; Tian, Q.; Watowich, S.S.; Jetten, A.M.; et al. Essential autocrine regulation by IL-21 in the generation of inflammatory T cells. Nature 2007, 448, 480–483. [Google Scholar] [CrossRef] [PubMed]
- Onoda, T.; Rahman, M.; Nara, H.; Araki, A.; Makabe, K.; Tsumoto, K.; Kumagai, I.; Kudo, T.; Ishii, N.; Tanaka, N.; et al. Human CD4+ central and effector memory T cells produce IL-21: Effect on cytokine-driven proliferation of CD4+ T cell subsets. Int. Immunol. 2007, 19, 1191–1199. [Google Scholar] [CrossRef] [Green Version]
- Wei, L.; Laurence, A.; Elias, K.M.; O’Shea, J.J. IL-21 is produced by Th17 cells and drives IL-17 production in a STAT3-dependent manner. J. Biol. Chem. 2007, 282, 34605–34610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, L.; Ivanov, I.I.; Spolski, R.; Min, R.; Shenderov, K.; Egawa, T.; Levy, D.E.; Leonard, W.J.; Littman, D.R. IL-6 programs T(H)-17 cell differentiation by promoting sequential engagement of the IL-21 and IL-23 pathways. Nat. Immunol. 2007, 8, 967–974. [Google Scholar] [CrossRef] [PubMed]
- Suto, A.; Kashiwakuma, D.; Kagami, S.; Hirose, K.; Watanabe, N.; Yokote, K.; Saito, Y.; Nakayama, T.; Grusby, M.J.; Iwamoto, I.; et al. Development and characterization of IL-21-producing CD4+ T cells. J. Exp. Med. 2008, 205, 1369–1379. [Google Scholar] [CrossRef] [PubMed]
- Chtanova, T.; Tangye, S.G.; Newton, R.; Frank, N.; Hodge, M.R.; Rolph, M.S.; Mackay, C.R. T follicular helper cells express a distinctive transcriptional profile, reflecting their role as non-Th1/Th2 effector cells that provide help for B cells. J. Immunol. 2004, 173, 68–78. [Google Scholar] [CrossRef]
- Nurieva, R.I.; Chung, Y.; Hwang, D.; Yang, X.O.; Kang, H.S.; Ma, L.; Wang, Y.H.; Watowich, S.S.; Jetten, A.M.; Tian, Q.; et al. Generation of T follicular helper cells is mediated by interleukin-21 but independent of T helper 1, 2, or 17 cell lineages. Immunity 2008, 29, 138–149. [Google Scholar] [CrossRef] [Green Version]
- Vogelzang, A.; McGuire, H.M.; Yu, D.; Sprent, J.; Mackay, C.R.; King, C. A fundamental role for interleukin-21 in the generation of T follicular helper cells. Immunity 2008, 29, 127–137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vinuesa, C.G.; Linterman, M.A.; Yu, D.; MacLennan, I.C. Follicular Helper T Cells. Annu. Rev. Immunol. 2016, 34, 335–368. [Google Scholar] [CrossRef]
- Crotty, S. T Follicular Helper Cell Biology: A Decade of Discovery and Diseases. Immunity 2019, 50, 1132–1148. [Google Scholar] [CrossRef]
- Victora, G.D.; Nussenzweig, M.C. Germinal centers. Annu. Rev. Immunol. 2012, 30, 429–457. [Google Scholar] [CrossRef]
- Suto, A.; Nakajima, H.; Hirose, K.; Suzuki, K.; Kagami, S.; Seto, Y.; Hoshimoto, A.; Saito, Y.; Foster, D.C.; Iwamoto, I. Interleukin 21 prevents antigen-induced IgE production by inhibiting germ line C(ε) transcription of IL-4-stimulated B cells. Blood 2002, 100, 4565–4573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pène, J.; Gauchat, J.F.; Lécart, S.; Drouet, E.; Guglielmi, P.; Boulay, V.; Delwail, A.; Foster, D.; Lecron, J.C.; Yssel, H. Cutting edge: IL-21 is a switch factor for the production of IgG1 and IgG3 by human B cells. J. Immunol. 2004, 172, 5154–5157. [Google Scholar] [CrossRef] [Green Version]
- Holland, S.M.; DeLeo, F.R.; Elloumi, H.Z.; Hsu, A.P.; Uzel, G.; Brodsky, N.; Freeman, A.F.; Demidowich, A.; Davis, J.; Turner, M.L.; et al. STAT3 mutations in the hyper-IgE syndrome. N. Engl. J. Med. 2007, 357, 1608–1619. [Google Scholar] [CrossRef] [PubMed]
- Minegishi, Y.; Saito, M.; Tsuchiya, S.; Tsuge, I.; Takada, H.; Hara, T.; Kawamura, N.; Ariga, T.; Pasic, S.; Stojkovic, O.; et al. Dominant-negative mutations in the DNA-binding domain of STAT3 cause hyper-IgE syndrome. Nature 2007, 448, 1058–1062. [Google Scholar] [CrossRef] [PubMed]
- Cagdas, D.; Mayr, D.; Baris, S.; Worley, L.; Langley, D.B.; Metin, A.; Aytekin, E.S.; Atan, R.; Kasap, N.; Bal, S.K.; et al. Genomic Spectrum and Phenotypic Heterogeneity of Human IL-21 Receptor Deficiency. J. Clin. Immunol. 2021, 41, 1272–1290. [Google Scholar] [CrossRef]
- Linterman, M.A.; Beaton, L.; Yu, D.; Ramiscal, R.R.; Srivastava, M.; Hogan, J.J.; Verma, N.K.; Smyth, M.J.; Rigby, R.J.; Vinuesa, C.G. IL-21 acts directly on B cells to regulate Bcl-6 expression and germinal center responses. J. Exp. Med. 2010, 207, 353–363. [Google Scholar] [CrossRef] [Green Version]
- Zotos, D.; Coquet, J.M.; Zhang, Y.; Light, A.; D’Costa, K.; Kallies, A.; Corcoran, L.M.; Godfrey, D.I.; Toellner, K.M.; Smyth, M.J.; et al. IL-21 regulates germinal center B cell differentiation and proliferation through a B cell-intrinsic mechanism. J. Exp. Med. 2010, 207, 365–378. [Google Scholar] [CrossRef] [Green Version]
- Kasaian, M.T.; Whitters, M.J.; Carter, L.L.; Lowe, L.D.; Jussif, J.M.; Deng, B.; Johnson, K.A.; Witek, J.S.; Senices, M.; Konz, R.F.; et al. IL-21 limits NK cell responses and promotes antigen-specific T cell activation: A mediator of the transition from innate to adaptive immunity. Immunity 2002, 16, 559–569. [Google Scholar] [CrossRef] [Green Version]
- Moroz, A.; Eppolito, C.; Li, Q.; Tao, J.; Clegg, C.H.; Shrikant, P.A. IL-21 enhances and sustains CD8+ T cell responses to achieve durable tumor immunity: Comparative evaluation of IL-2, IL-15, and IL-21. J. Immunol. 2004, 173, 900–909. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, R.; Spolski, R.; Finkelstein, S.E.; Oh, S.; Kovanen, P.E.; Hinrichs, C.S.; Pise-Masison, C.A.; Radonovich, M.F.; Brady, J.N.; Restifo, N.P.; et al. Synergy of IL-21 and IL-15 in regulating CD8+ T cell expansion and function. J. Exp. Med. 2005, 201, 139–148. [Google Scholar] [CrossRef] [Green Version]
- Barker, B.R.; Gladstone, M.N.; Gillard, G.O.; Panas, M.W.; Letvin, N.L. Critical role for IL-21 in both primary and memory anti-viral CD8+ T-cell responses. Eur. J. Immunol. 2010, 40, 3085–3096. [Google Scholar] [CrossRef]
- Novy, P.; Huang, X.; Leonard, W.J.; Yang, Y. Intrinsic IL-21 signaling is critical for CD8 T cell survival and memory formation in response to vaccinia viral infection. J. Immunol. 2011, 186, 2729–2738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gazi, M.Y.; Takeda, Y.; Nara, H.; Araki, A.; Nemoto, N.; Asao, H. Interleukin-21 reduces Listeria monocytogenes secondary infection via CD8+ effector memory T cells. Yamagata Med. J. 2021, 39, 45–51. [Google Scholar] [CrossRef]
- Zajac, A.J.; Blattman, J.N.; Murali-Krishna, K.; Sourdive, D.J.; Suresh, M.; Altman, J.D.; Ahmed, R. Viral immune evasion due to persistence of activated T cells without effector function. J. Exp. Med. 1998, 188, 2205–2213. [Google Scholar] [CrossRef]
- Kahan, S.M.; Wherry, E.J.; Zajac, A.J. T cell exhaustion during persistent viral infections. Virology 2015, 479–480, 180–193. [Google Scholar] [CrossRef] [PubMed]
- Wherry, E.J.; Kurachi, M. Molecular and cellular insights into T cell exhaustion. Nat. Rev. Immunol. 2015, 15, 486–499. [Google Scholar] [CrossRef] [PubMed]
- Schmitz, J.E.; Kuroda, M.J.; Santra, S.; Sasseville, V.G.; Simon, M.A.; Lifton, M.A.; Racz, P.; Tenner-Racz, K.; Dalesandro, M.; Scallon, B.J.; et al. Control of viremia in simian immunodeficiency virus infection by CD8+ lymphocytes. Science 1999, 283, 857–860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, X.; Bauer, D.E.; Tuttleton, S.E.; Lewin, S.; Gettie, A.; Blanchard, J.; Irwin, C.E.; Safrit, J.T.; Mittler, J.; Weinberger, L.; et al. Dramatic rise in plasma viremia after CD8(+) T cell depletion in simian immunodeficiency virus-infected macaques. J. Exp. Med. 1999, 189, 991–998. [Google Scholar] [CrossRef] [Green Version]
- Zander, R.; Schauder, D.; Xin, G.; Nguyen, C.; Wu, X.; Zajac, A.; Cui, W. CD4+ T Cell Help Is Required for the Formation of a Cytolytic CD8+ T Cell Subset that Protects against Chronic Infection and Cancer. Immunity 2019, 51, 1028–1042.e4. [Google Scholar] [CrossRef] [PubMed]
- Fröhlich, A.; Kisielow, J.; Schmitz, I.; Freigang, S.; Shamshiev, A.T.; Weber, J.; Marsland, B.J.; Oxenius, A.; Kopf, M. IL-21R on T cells is critical for sustained functionality and control of chronic viral infection. Science 2009, 324, 1576–1580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elsaesser, H.; Sauer, K.; Brooks, D.G. IL-21 is required to control chronic viral infection. Science 2009, 324, 1569–1572. [Google Scholar] [CrossRef] [Green Version]
- Yi, J.S.; Du, M.; Zajac, A.J. A vital role for interleukin-21 in the control of a chronic viral infection. Science 2009, 324, 1572–1576. [Google Scholar] [CrossRef] [PubMed]
- Cui, W.; Liu, Y.; Weinstein, J.S.; Craft, J.; Kaech, S.M. An interleukin-21-interleukin-10-STAT3 pathway is critical for functional maturation of memory CD8+ T cells. Immunity 2011, 35, 792–805. [Google Scholar] [CrossRef] [Green Version]
- Xin, G.; Schauder, D.M.; Lainez, B.; Weinstein, J.S.; Dai, Z.; Chen, Y.; Esplugues, E.; Wen, R.; Wang, D.; Parish, I.A.; et al. A Critical Role of IL-21-Induced BATF in Sustaining CD8-T-Cell-Mediated Chronic Viral Control. Cell Rep. 2015, 13, 1118–1124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loschinski, R.; Böttcher, M.; Stoll, A.; Bruns, H.; Mackensen, A.; Mougiakakos, D. IL-21 modulates memory and exhaustion phenotype of T-cells in a fatty acid oxidation-dependent manner. Oncotarget 2018, 9, 13125–13138. [Google Scholar] [CrossRef] [Green Version]
- Iannello, A.; Tremblay, C.; Routy, J.P.; Boulassel, M.R.; Toma, E.; Ahmad, A. Decreased levels of circulating IL-21 in HIV-infected AIDS patients: Correlation with CD4+ T-cell counts. Viral Immunol. 2008, 21, 385–388. [Google Scholar] [CrossRef]
- Iannello, A.; Boulassel, M.R.; Samarani, S.; Debbeche, O.; Tremblay, C.; Toma, E.; Routy, J.P.; Ahmad, A. Dynamics and consequences of IL-21 production in HIV-infected individuals: A longitudinal and cross-sectional study. J. Immunol. 2010, 184, 114–126. [Google Scholar] [CrossRef] [Green Version]
- Chevalier, M.F.; Jülg, B.; Pyo, A.; Flanders, M.; Ranasinghe, S.; Soghoian, D.Z.; Kwon, D.S.; Rychert, J.; Lian, J.; Muller, M.I.; et al. HIV-1-specific interleukin-21+ CD4+ T cell responses contribute to durable viral control through the modulation of HIV-specific CD8+ T cell function. J. Virol. 2011, 85, 733–741. [Google Scholar] [CrossRef] [Green Version]
- Dalel, J.; Ung, S.K.; Hayes, P.; Black, S.L.; Joseph, S.; King, D.F.; Makinde, J.; Gilmour, J. IAVI Protocol C investigators list. HIV-1 infection and the lack of viral control are associated with greater expression of interleukin-21 receptor on CD8+ T cells. AIDS 2021, 35, 1167–1177. [Google Scholar] [CrossRef]
- Williams, L.D.; Bansal, A.; Sabbaj, S.; Heath, S.L.; Song, W.; Tang, J.; Zajac, A.J.; Goepfert, P.A. Interleukin-21-producing HIV-1-specific CD8 T cells are preferentially seen in elite controllers. J. Virol. 2011, 85, 2316–2324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, L.D.; Amatya, N.; Bansal, A.; Sabbaj, S.; Heath, S.L.; Sereti, I.; Goepfert, P.A. Immune activation is associated with CD8 T cell interleukin-21 production in HIV-1-infected individuals. J. Virol. 2014, 88, 10259–10263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ortega, C.; Fernández-A, S.; Carrillo, J.M.; Romero, P.; Molina, I.J.; Moreno, J.C.; Santamaría, M. IL-17-producing CD8+ T lymphocytes from psoriasis skin plaques are cytotoxic effector cells that secrete Th17-related cytokines. J. Leukoc. Biol. 2009, 86, 435–443. [Google Scholar] [CrossRef]
- Dolff, S.; Abdulahad, W.H.; Westra, J.; Doornbos-van der Meer, B.; Limburg, P.C.; Kallenberg, C.G.; Bijl, M. Increase in IL-21 producing T-cells in patients with systemic lupus erythematosus. Arthritis Res. Ther. 2011, 13, R157. [Google Scholar] [CrossRef] [Green Version]
- Mittal, A.; Murugaiyan, G.; Beynon, V.; Hu, D.; Weiner, H.L. IL-27 induction of IL-21 from human CD8+ T cells induces granzyme B in an autocrine manner. Immunol. Cell Biol. 2012, 90, 831–835. [Google Scholar] [CrossRef]
- Buckner, C.M.; Moir, S.; Ho, J.; Wang, W.; Posada, J.G.; Kardava, L.; Funk, E.K.; Nelson, A.K.; Li, Y.; Chun, T.W.; et al. Characterization of plasmablasts in the blood of HIV-infected viremic individuals: Evidence for nonspecific immune activation. J. Virol. 2013, 87, 5800–5811. [Google Scholar] [CrossRef] [Green Version]
- Moir, S.; Fauci, A.S. B-cell responses to HIV infection. Immunol. Rev. 2017, 275, 33–48. [Google Scholar] [CrossRef] [PubMed]
- Kardava, L.; Moir, S.; Shah, N.; Wang, W.; Wilson, R.; Buckner, C.M.; Santich, B.H.; Kim, L.J.; Spurlin, E.E.; Nelson, A.K.; et al. Abnormal B cell memory subsets dominate HIV-specific responses in infected individuals. J. Clin. Investig. 2014, 124, 3252–3262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kernéis, S.; Launay, O.; Turbelin, C.; Batteux, F.; Hanslik, T.; Boëlle, P.Y. Long-term immune responses to vaccination in HIV-infected patients: A systematic review and meta-analysis. Clin. Infect. Dis. 2014, 58, 1130–1139. [Google Scholar] [CrossRef] [PubMed]
- Kwong, P.D.; Mascola, J.R. Human antibodies that neutralize HIV-1: Identification, structures, and B cell ontogenies. Immunity 2012, 37, 412–425. [Google Scholar] [CrossRef] [Green Version]
- Walker, L.M.; Phogat, S.K.; Chan-Hui, P.Y.; Wagner, D.; Phung, P.; Goss, J.L.; Wrin, T.; Simek, M.D.; Fling, S.; Mitcham, J.L.; et al. Broad and potent neutralizing antibodies from an African donor reveal a new HIV-1 vaccine target. Science 2009, 326, 285–289. [Google Scholar] [CrossRef] [Green Version]
- Walker, L.M.; Huber, M.; Doores, K.J.; Falkowska, E.; Pejchal, R.; Julien, J.P.; Wang, S.K.; Ramos, A.; Chan-Hui, P.Y.; Moyle, M.; et al. Broad neutralization coverage of HIV by multiple highly potent antibodies. Nature 2011, 477, 466–470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scheid, J.F.; Mouquet, H.; Ueberheide, B.; Diskin, R.; Klein, F.; Oliveira, T.Y.; Pietzsch, J.; Fenyo, D.; Abadir, A.; Velinzon, K.; et al. Sequence and structural convergence of broad and potent HIV antibodies that mimic CD4 binding. Science 2011, 333, 1633–1637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, J.; Ofek, G.; Laub, L.; Louder, M.K.; Doria-Rose, N.A.; Longo, N.S.; Imamichi, H.; Bailer, R.T.; Chakrabarti, B.; Sharma, S.K.; et al. Broad and potent neutralization of HIV-1 by a gp41-specific human antibody. Nature 2012, 491, 406–412. [Google Scholar] [CrossRef] [Green Version]
- Rerks-Ngarm, S.; Pitisuttithum, P.; Nitayaphan, S.; Kaewkungwal, J.; Chiu, J.; Paris, R.; Premsri, N.; Namwat, C.; de Souza, M.; Adams, E.; et al. Vaccination with ALVAC and AIDSVAX to prevent HIV-1 infection in Thailand. N. Engl. J. Med. 2009, 361, 2209–2220. [Google Scholar] [CrossRef]
- Burton, D.R.; Ahmed, R.; Barouch, D.H.; Butera, S.T.; Crotty, S.; Godzik, A.; Kaufmann, D.E.; McElrath, M.J.; Nussenzweig, M.C.; Pulendran, B.; et al. A Blueprint for HIV Vaccine Discovery. Cell Host Microbe. 2012, 12, 396–407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.H.; Rerks-Ngarm, S.; Excler, J.L.; Michael, N.L. HIV vaccines: Lessons learned and the way forward. Curr. Opin. HIV AIDS 2010, 5, 428–434. [Google Scholar] [CrossRef] [Green Version]
- McMichael, A.J.; Haynes, B.F. Lessons learned from HIV-1 vaccine trials: New priorities and directions. Nat. Immunol. 2012, 13, 423–427. [Google Scholar] [CrossRef] [Green Version]
- Streeck, H.; D’Souza, M.P.; Littman, D.R.; Crotty, S. Harnessing CD4⁺ T cell responses in HIV vaccine development. Nat. Med. 2013, 19, 143–149. [Google Scholar] [CrossRef] [Green Version]
- Pissani, F.; Schulte, B.; Eller, M.A.; Schultz, B.T.; Ratto-Kim, S.; Marovich, M.; Thongcharoen, P.; Sriplienchan, S.; Rerks-Ngarm, S.; Pitisuttithum, P.; et al. Modulation of Vaccine-Induced CD4 T Cell Functional Profiles by Changes in Components of HIV Vaccine Regimens in Humans. J. Virol. 2018, 92, e01143-18. [Google Scholar] [CrossRef] [Green Version]
- Perreau, M.; Savoye, A.L.; De Crignis, E.; Corpataux, J.M.; Cubas, R.; Haddad, E.K.; De Leval, L.; Graziosi, C.; Pantaleo, G. Follicular helper T cells serve as the major CD4 T cell compartment for HIV-1 infection, replication, and production. J. Exp. Med. 2013, 210, 143–156. [Google Scholar] [CrossRef] [PubMed]
- Pallikkuth, S.; Sharkey, M.; Babic, D.Z.; Gupta, S.; Stone, G.W.; Fischl, M.A.; Stevenson, M.; Pahwa, S. Peripheral T Follicular Helper Cells Are the Major HIV Reservoir within Central Memory CD4 T Cells in Peripheral Blood from Chronically HIV-Infected Individuals on Combination Antiretroviral Therapy. J. Virol. 2015, 90, 2718–2728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García, M.; Górgolas, M.; Cabello, A.; Estrada, V.; Ligos, J.M.; Fernández-Guerrero, M.; Barros, C.; López-Bernaldo, J.C.; De La Hera, F.J.; Montoya, M.; et al. Peripheral T follicular helper Cells Make a Difference in HIV Reservoir Size between Elite Controllers and Patients on Successful cART. Sci. Rep. 2017, 7, 16799. [Google Scholar] [CrossRef] [Green Version]
- Lindqvist, M.; van Lunzen, J.; Soghoian, D.Z.; Kuhl, B.D.; Ranasinghe, S.; Kranias, G.; Flanders, M.D.; Cutler, S.; Yudanin, N.; Muller, M.I.; et al. Expansion of HIV-specific T follicular helper cells in chronic HIV infection. J. Clin. Investig. 2012, 122, 3271–3280. [Google Scholar] [CrossRef]
- Pissani, F.; Streeck, H. Emerging concepts on T follicular helper cell dynamics in HIV infection. Trends Immunol. 2014, 35, 278–286. [Google Scholar] [CrossRef] [Green Version]
- Colineau, L.; Rouers, A.; Yamamoto, T.; Xu, Y.; Urrutia, A.; Pham, H.P.; Cardinaud, S.; Samri, A.; Dorgham, K.; Coulon, P.G.; et al. HIV-Infected Spleens Present Altered Follicular Helper T Cell (Tfh) Subsets and Skewed B Cell Maturation. PLoS ONE 2015, 10, e0140978. [Google Scholar] [CrossRef] [Green Version]
- Graff-Dubois, S.; Rouers, A.; Moris, A. Impact of Chronic HIV/SIV Infection on T Follicular Helper Cell Subsets and Germinal Center Homeostasis. Front. Immunol. 2016, 7, 501. [Google Scholar] [CrossRef] [Green Version]
- Boswell, K.L.; Paris, R.; Boritz, E.; Ambrozak, D.; Yamamoto, T.; Darko, S.; Wloka, K.; Wheatley, A.; Narpala, S.; McDermott, A.; et al. Loss of circulating CD4 T cells with B cell helper function during chronic HIV infection. PLoS Pathog. 2014, 10, e1003853. [Google Scholar] [CrossRef]
- Moukambi, F.; Rabezanahary, H.; Rodrigues, V.; Racine, G.; Robitaille, L.; Krust, B.; Andreani, G.; Soundaramourty, C.; Silvestre, R.; Laforge, M.; et al. Early Loss of Splenic Tfh Cells in SIV-Infected Rhesus Macaques. PLoS Pathog. 2015, 11, e1005287. [Google Scholar] [CrossRef]
- Morita, R.; Schmitt, N.; Bentebibel, S.E.; Ranganathan, R.; Bourdery, L.; Zurawski, G.; Foucat, E.; Dullaers, M.; Oh, S.; Sabzghabaei, N.; et al. Human blood CXCR5(+)CD4(+) T cells are counterparts of T follicular cells and contain specific subsets that differentially support antibody secretion. Immunity 2011, 34, 108–121. [Google Scholar] [CrossRef] [Green Version]
- Locci, M.; Havenar-Daughton, C.; Landais, E.; Wu, J.; Kroenke, M.A.; Arlehamn, C.L.; Su, L.F.; Cubas, R.; Davis, M.M.; Sette, A.; et al. Human circulating PD-1+CXCR3-CXCR5+ memory Tfh cells are highly functional and correlate with broadly neutralizing HIV antibody responses. Immunity 2013, 39, 758–769. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, T.; Lynch, R.M.; Gautam, R.; Matus-Nicodemos, R.; Schmidt, S.D.; Boswell, K.L.; Darko, S.; Wong, P.; Sheng, Z.; Petrovas, C.; et al. Quality and quantity of TFH cells are critical for broad antibody development in SHIVAD8 infection. Sci. Transl. Med. 2015, 7, 298ra120. [Google Scholar] [CrossRef] [Green Version]
- Schultz, B.T.; Teigler, J.E.; Pissani, F.; Oster, A.F.; Kranias, G.; Alter, G.; Marovich, M.; Eller, M.A.; Dittmer, U.; Robb, M.L.; et al. Circulating HIV-Specific Interleukin-21(+)CD4(+) T Cells Represent Peripheral Tfh Cells with Antigen-Dependent Helper Functions. Immunity 2016, 44, 167–178. [Google Scholar] [CrossRef] [Green Version]
- Buranapraditkun, S.; Pissani, F.; Teigler, J.E.; Schultz, B.T.; Alter, G.; Marovich, M.; Robb, M.L.; Eller, M.A.; Martin, J.; Deeks, S.; et al. Preservation of Peripheral T Follicular Helper Cell Function in HIV Controllers. J. Virol. 2017, 91, e00497-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cubas, R.; van Grevenynghe, J.; Wills, S.; Kardava, L.; Santich, B.H.; Buckner, C.M.; Muir, R.; Tardif, V.; Nichols, C.; Procopio, F.; et al. Reversible Reprogramming of Circulating Memory T Follicular Helper Cell Function during Chronic HIV Infection. J. Immunol. 2015, 195, 5625–5636. [Google Scholar] [CrossRef]
- Landais, E.; Moore, P.L. Development of broadly neutralizing antibodies in HIV-1 infected elite neutralizers. Retrovirology 2018, 15, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hessell, A.J.; Hangartner, L.; Hunter, M.; Havenith, C.E.; Beurskens, F.J.; Bakker, J.M.; Lanigan, C.M.; Landucci, G.; Forthal, D.N.; Parren, P.W.; et al. Fc receptor but not complement binding is important in antibody protection against HIV. Nature 2007, 449, 101–104. [Google Scholar] [CrossRef] [PubMed]
- Bournazos, S.; Klein, F.; Pietzsch, J.; Seaman, M.S.; Nussenzweig, M.C.; Ravetch, J.V. Broadly neutralizing anti-HIV-1 antibodies require Fc effector functions for in vivo activity. Cell 2014, 158, 1243–1253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ackerman, M.E.; Mikhailova, A.; Brown, E.P.; Dowell, K.G.; Walker, B.D.; Bailey-Kellogg, C.; Suscovich, T.J.; Alter, G. Polyfunctional HIV-Specific Antibody Responses Are Associated with Spontaneous HIV Control. PLoS Pathog. 2016, 12, e1005315. [Google Scholar] [CrossRef]
- Pušnik, J.; Fischinger, S.; Dittmer, U.; Esser, S.; van Gils, M.J.; Sanders, R.W.; Alter, G.; Streeck, H. Production of HIV-1 Env-specific antibodies mediating innate immune functions depends on cognate IL-21- secreting CD4+ T cells. J. Virol. 2021, 95, e02097-20. [Google Scholar] [CrossRef]
- Adoro, S.; Cubillos-Ruiz, J.R.; Chen, X.; Deruaz, M.; Vrbanac, V.D.; Song, M.; Park, S.; Murooka, T.T.; Dudek, T.E.; Luster, A.D.; et al. IL-21 induces antiviral microRNA-29 in CD4 T cells to limit HIV-1 infection. Nat. Commun. 2015, 6, 7562. [Google Scholar] [CrossRef] [Green Version]
- Ortega, P.A.S.; Saulle, I.; Mercurio, V.; Ibba, S.V.; Lori, E.M.; Fenizia, C.; Masetti, M.; Trabattoni, D.; Caputo, S.L.; Vichi, F.; et al. Interleukin 21 (IL-21)/microRNA-29 (miR-29) axis is associated with natural resistance to HIV-1 infection. AIDS 2018, 32, 2453–2461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brenchley, J.M.; Price, D.A.; Schacker, T.W.; Asher, T.E.; Silvestri, G.; Rao, S.; Kazzaz, Z.; Bornstein, E.; Lambotte, O.; Altmann, D.; et al. Microbial translocation is a cause of systemic immune activation in chronic HIV infection. Nat. Med. 2006, 12, 1365–1371. [Google Scholar] [CrossRef] [Green Version]
- Favre, D.; Lederer, S.; Kanwar, B.; Ma, Z.M.; Proll, S.; Kasakow, Z.; Mold, J.; Swainson, L.; Barbour, J.D.; Baskin, C.R.; et al. Critical loss of the balance between Th17 and T regulatory cell populations in pathogenic SIV infection. PLoS Pathog. 2009, 5, e1000295. [Google Scholar] [CrossRef]
- Estes, J.D.; Harris, L.D.; Klatt, N.R.; Tabb, B.; Pittaluga, S.; Paiardini, M.; Barclay, G.R.; Smedley, J.; Pung, R.; Oliveira, K.M.; et al. Damaged intestinal epithelial integrity linked to microbial translocation in pathogenic simian immunodeficiency virus infections. PLoS Pathog. 2010, 6, e1001052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roberts, E.R.; Carnathan, D.G.; Li, H.; Shaw, G.M.; Silvestri, G.; Betts, M.R. Collapse of Cytolytic Potential in SIV-Specific CD8+ T Cells Following Acute SIV Infection in Rhesus Macaques. PLoS Pathog. 2016, 12, e1006135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Micci, L.; Cervasi, B.; Ende, Z.S.; Iriele, R.I.; Reyes-Aviles, E.; Vinton, C.; Else, J.; Silvestri, G.; Ansari, A.A.; Villinger, F.; et al. Paucity of IL-21-producing CD4(+) T cells is associated with Th17 cell depletion in SIV infection of rhesus macaques. Blood 2012, 120, 3925–3935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pallikkuth, S.; Rogers, K.; Villinger, F.; Dosterii, M.; Vaccari, M.; Franchini, G.; Pahwa, R.; Pahwa, S. Interleukin-21 administration to rhesus macaques chronically infected with simian immunodeficiency virus increases cytotoxic effector molecules in T cells and NK cells and enhances B cell function without increasing immune activation or viral replication. Vaccine 2011, 29, 9229–9238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pallikkuth, S.; Micci, L.; Ende, Z.S.; Iriele, R.I.; Cervasi, B.; Lawson, B.; McGary, C.S.; Rogers, K.A.; Else, J.G.; Silvestri, G.; et al. Maintenance of intestinal Th17 cells and reduced microbial translocation in SIV-infected rhesus macaques treated with interleukin (IL)-21. PLoS Pathog. 2013, 9, e1003471. [Google Scholar] [CrossRef] [Green Version]
- Méndez-Lagares, G.; Lu, D.; Merriam, D.; Baker, C.A.; Villinger, F.; Van Rompay, K.K.A.; McCune, J.M.; Hartigan-O’Connor, D.J. IL-21 Therapy Controls Immune Activation and Maintains Antiviral CD8+ T Cell Responses in Acute Simian Immunodeficiency Virus Infection. AIDS Res. Hum. Retrovir. 2017, 33, S81–S92. [Google Scholar] [CrossRef] [Green Version]
- Seo, H.; Jeon, I.; Kim, B.S.; Park, M.; Bae, E.A.; Song, B.; Koh, C.H.; Shin, K.S.; Kim, I.K.; Choi, K.; et al. IL-21-mediated reversal of NK cell exhaustion facilitates anti-tumour immunity in MHC class I-deficient tumours. Nat. Commun. 2017, 8, 15776. [Google Scholar] [CrossRef]
- Huot, N.; Rascle, P.; Petitdemange, C.; Contreras, V.; Stürzel, C.M.; Baquero, E.; Harper, J.L.; Passaes, C.; Legendre, R.; Varet, H.; et al. SIV-induced terminally differentiated adaptive NK cells in lymph nodes associated with enhanced MHC-E restricted activity. Nat. Commun. 2021, 12, 1282. [Google Scholar] [CrossRef]
- Harper, J.; Huot, N.; Micci, L.; Tharp, G.; King, C.; Rascle, P.; Shenvi, N.; Wang, H.; Galardi, C.; Upadhyay, A.A.; et al. IL-21 and IFNα therapy rescues terminally differentiated NK cells and limits SIV reservoir in ART-treated macaques. Nat. Commun. 2021, 12, 2866. [Google Scholar] [CrossRef] [PubMed]
- Schweitzer, A.; Horn, J.; Mikolajczyk, R.T.; Krause, G.; Ott, J.J. Estimations of worldwide prevalence of chronic hepatitis B virus infection: A systematic review of data published between 1965 and 2013. Lancet 2015, 386, 1546–1555. [Google Scholar] [CrossRef]
- Publicover, J.; Goodsell, A.; Nishimura, S.; Vilarinho, S.; Wang, Z.E.; Avanesyan, L.; Spolski, R.; Leonard, W.J.; Cooper, S.; Baron, J.L. IL-21 is pivotal in determining age-dependent effectiveness of immune responses in a mouse model of human hepatitis B. J. Clin. Investig. 2011, 121, 1154–1162. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Liu, M.; Cheng, L.W.; Gao, X.Y.; Fu, J.J.; Kong, G.; Feng, X.; Pan, X.C. HBcAg-specific IL-21-producing CD4+ T cells are associated with relative viral control in patients with chronic hepatitis B. Scand. J. Immunol. 2013, 78, 439–446. [Google Scholar] [CrossRef] [Green Version]
- Ma, S.W.; Huang, X.; Li, Y.Y.; Tang, L.B.; Sun, X.F.; Jiang, X.T.; Zhang, Y.X.; Sun, J.; Liu, Z.H.; Abbott, W.G.; et al. High serum IL-21 levels after 12 weeks of antiviral therapy predict HBeAg seroconversion in chronic hepatitis B. J. Hepatol. 2012, 56, 775–781. [Google Scholar] [CrossRef]
- Tang, L.; Chen, C.; Gao, X.; Zhang, W.; Yan, X.; Zhou, Y.; Guo, L.; Zheng, X.; Wang, W.; Yang, F.; et al. Interleukin 21 Reinvigorates the Antiviral Activity of Hepatitis B Virus (HBV)-Specific CD8+ T Cells in Chronic HBV Infection. J. Infect. Dis. 2019, 219, 750–759. [Google Scholar] [CrossRef]
- Shen, Z.; Yang, H.; Yang, S.; Wang, W.; Cui, X.; Zhou, X.; Liu, W.; Pan, S.; Liu, Y.; Zhang, J.; et al. Hepatitis B virus persistence in mice reveals IL-21 and IL-33 as regulators of viral clearance. Nat. Commun. 2017, 8, 2119. [Google Scholar] [CrossRef] [Green Version]
- Shen, Z.; Liu, J.; Wu, J.; Zhu, Y.; Li, G.; Wang, J.; Luo, M.; Deng, Q.; Zhang, J.; Xie, Y. IL-21-based therapies induce clearance of hepatitis B virus persistence in mouse models. Theranostics 2019, 9, 3798–3811. [Google Scholar] [CrossRef]
- Shen, Z.; Wu, J.; Gao, Z.; Wang, J.; Zhu, H.; Mao, R.; Wang, X.; Zhang, J.; Xie, Y.; Liu, J. Characterization of IL-21-expressing recombinant hepatitis B virus (HBV) as a therapeutic agent targeting persisting HBV infection. Theranostics 2020, 10, 5600–5612. [Google Scholar] [CrossRef] [PubMed]
- Shen, Z.; Gao, Z.; Gu, C.; Wu, J.; Wang, J.; Zhang, J.; Xie, Y.; Liu, J. Prior transient exposure to interleukin-21 delivered by recombinant adeno-associated virus vector protects mice from hepatitis B virus persistence. Antivir. Res. 2021, 190, 105076. [Google Scholar] [CrossRef] [PubMed]
- Ye, B.; Liu, X.; Li, X.; Kong, H.; Tian, L.; Chen, Y. T-cell exhaustion in chronic hepatitis B infection: Current knowledge and clinical significance. Cell Death Dis. 2015, 6, e1694. [Google Scholar] [CrossRef] [Green Version]
- Spaan, M.; Kreefft, K.; de Graav, G.N.; Brouwer, W.P.; de Knegt, R.J.; ten Kate, F.J.; Baan, C.C.; Vanwolleghem, T.; Janssen, H.L.; Boonstra, A. CD4+ CXCR5+ T cells in chronic HCV infection produce less IL-21, yet are efficient at supporting B cell responses. J. Hepatol. 2015, 62, 303–310. [Google Scholar] [CrossRef]
- Khanam, A.; Ayithan, N.; Tang, L.; Poonia, B.; Kottilil, S. IL-21-Deficient T Follicular Helper Cells Support B Cell Responses Through IL-27 in Patients with Chronic Hepatitis B. Front. Immunol. 2021, 11, 599648. [Google Scholar] [CrossRef]
- Bao, S.; Zheng, J.; Shi, G. The role of T helper 17 cells in the pathogenesis of hepatitis B virus-related liver cirrhosis. Mol. Med. Rep. 2017, 16, 3713–3719. [Google Scholar] [CrossRef] [Green Version]
- Sun, H.Q.; Zhang, J.Y.; Zhang, H.; Zou, Z.S.; Wang, F.S.; Jia, J.H. Increased Th17 cells contribute to disease progression in patients with HBV-associated liver cirrhosis. J. Viral Hepat. 2012, 19, 396–403. [Google Scholar] [CrossRef]
- Yang, C.; Cui, F.; Chen, L.M.; Gong, X.Y.; Qin, B. Correlation between Th17 and nTreg cell frequencies and the stages of progression in chronic hepatitis B. Mol. Med. Rep. 2016, 13, 853–859. [Google Scholar] [CrossRef] [Green Version]
- Feng, G.; Zhang, J.Y.; Zeng, Q.L.; Yu, X.; Zhang, Z.; Lv, S.; Xu, X.; Wang, F.S. Interleukin-21 mediates hepatitis B virus-associated liver cirrhosis by activating hepatic stellate cells. Hepatol. Res. 2014, 44, E198–E205. [Google Scholar] [CrossRef]
- Li, Y.; Tang, L.; Hou, J. Role of interleukin-21 in HBV infection: Friend or foe? Cell Mol. Immunol. 2015, 12, 303–308. [Google Scholar] [CrossRef] [Green Version]
- Thrift, A.P.; El-Serag, H.B.; Kanwal, F. Global epidemiology and burden of HCV infection and HCV-related disease. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 122–132. [Google Scholar] [CrossRef]
- Peña-Asensio, J.; Calvo, H.; Torralba, M.; Miquel, J.; Sanz-de-Villalobos, E.; Larrubia, J.R. Gamma-Chain Receptor Cytokines & PD-1 Manipulation to Restore HCV-Specific CD8+ T Cell Response during Chronic Hepatitis C. Cells 2021, 10, 538. [Google Scholar] [CrossRef]
- Grakoui, A.; Shoukry, N.H.; Woollard, D.J.; Han, J.H.; Hanson, H.L.; Ghrayeb, J.; Murthy, K.K.; Rice, C.M.; Walker, C.M. HCV persistence and immune evasion in the absence of memory T cell help. Science 2003, 302, 659–662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shoukry, N.H.; Grakoui, A.; Houghton, M.; Chien, D.Y.; Ghrayeb, J.; Reimann, K.A.; Walker, C.M. Memory CD8+ T cells are required for protection from persistent hepatitis C virus infection. J. Exp. Med. 2003, 197, 1645–1655. [Google Scholar] [CrossRef]
- Osburn, W.O.; Fisher, B.E.; Dowd, K.A.; Urban, G.; Liu, L.; Ray, S.C.; Thomas, D.L.; Cox, A.L. Spontaneous control of primary hepatitis C virus infection and immunity against persistent reinfection. Gastroenterology 2010, 138, 315–324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osburn, W.O.; Snider, A.E.; Wells, B.L.; Latanich, R.; Bailey, J.R.; Thomas, D.L.; Cox, A.L.; Ray, S.C. Clearance of hepatitis C infection is associated with the early appearance of broad neutralizing antibody responses. Hepatology 2014, 59, 2140–2151. [Google Scholar] [CrossRef] [PubMed]
- Kinchen, V.J.; Massaccesi, G.; Flyak, A.I.; Mankowski, M.C.; Colbert, M.D.; Osburn, W.O.; Ray, S.C.; Cox, A.L.; Crowe, J.E., Jr.; Bailey, J.R. Plasma deconvolution identifies broadly neutralizing antibodies associated with hepatitis C virus clearance. J. Clin. Investig. 2019, 129, 4786–4796. [Google Scholar] [CrossRef] [PubMed]
- Raziorrouh, B.; Sacher, K.; Tawar, R.G.; Emmerich, F.; Neumann-Haefelin, C.; Baumert, T.F.; Thimme, R.; Boettler, T. Virus-Specific CD4+ T Cells Have Functional and Phenotypic Characteristics of Follicular T-Helper Cells in Patients with Acute and Chronic HCV Infections. Gastroenterology 2016, 150, 696–706.e3. [Google Scholar] [CrossRef]
- Smits, M.; Zoldan, K.; Ishaque, N.; Gu, Z.; Jechow, K.; Wieland, D.; Conrad, C.; Eils, R.; Fauvelle, C.; Baumert, T.F.; et al. Follicular T helper cells shape the HCV-specific CD4+ T cell repertoire after virus elimination. J. Clin. Investig. 2020, 130, 998–1009. [Google Scholar] [CrossRef] [Green Version]
- Salinas, E.; Boisvert, M.; Upadhyay, A.A.; Bédard, N.; Nelson, S.A.; Bruneau, J.; Derdeyn, C.A.; Marcotrigiano, J.; Evans, M.J.; Bosinger, S.E.; et al. Early T follicular helper cell activity accelerates hepatitis C virus-specific B cell expansion. J. Clin. Investig. 2021, 131, e140590. [Google Scholar] [CrossRef] [PubMed]
- Mockus, T.E.; Shwetank; Lauver, M.D.; Ren, H.M.; Netherby, C.S.; Salameh, T.; Kawasawa, Y.I.; Yue, F.; Broach, J.R.; Lukacher, A.E. CD4 T cells control development and maintenance of brain-resident CD8 T cells during polyomavirus infection. PLoS Pathog. 2018, 14, e1007365. [Google Scholar] [CrossRef] [PubMed]
- Ren, H.M.; Lukacher, A.E. IL-21 in Homeostasis of Resident Memory and Exhausted CD8 T Cells during Persistent Infection. Int. J. Mol. Sci. 2020, 21, 6966. [Google Scholar] [CrossRef] [PubMed]
- Ren, H.M.; Kolawole, E.M.; Ren, M.; Jin, G.; Netherby-Winslow, C.S.; Wade, Q.; Shwetank; Rahman, Z.S.M.; Evavold, B.D.; Lukacher, A.E. IL-21 from high-affinity CD4 T cells drives differentiation of brain-resident CD8 T cells during persistent viral infection. Sci. Immunol. 2020, 5, eabb5590. [Google Scholar] [CrossRef]
- Shah, V.K.; Firmal, P.; Alam, A.; Ganguly, D.; Chattopadhyay, S. Overview of Immune Response During SARS-CoV-2 Infection: Lessons from the Past. Front. Immunol. 2020, 11, 1949. [Google Scholar] [CrossRef]
- Hu, B.; Guo, H.; Zhou, P.; Shi, Z.L. Characteristics of SARS-CoV-2 and COVID-19. Nat. Rev. Microbiol. 2021, 19, 141–154. [Google Scholar] [CrossRef]
- Schultze, J.L.; Aschenbrenner, A.C. COVID-19 and the human innate immune system. Cell 2021, 184, 1671–1692. [Google Scholar] [CrossRef]
- Sette, A.; Crotty, S. Adaptive immunity to SARS-CoV-2 and COVID-19. Cell 2021, 184, 861–880. [Google Scholar] [CrossRef]
- Hartley, G.E.; Edwards, E.S.J.; Aui, P.M.; Varese, N.; Stojanovic, S.; McMahon, J.; Peleg, A.Y.; Boo, I.; Drummer, H.E.; Hogarth, P.M.; et al. Rapid generation of durable B cell memory to SARS-CoV-2 spike and nucleocapsid proteins in COVID-19 and convalescence. Sci. Immunol. 2020, 5, eabf8891. [Google Scholar] [CrossRef]
- Byazrova, M.; Yusubalieva, G.; Spiridonova, A.; Efimov, G.; Mazurov, D.; Baranov, K.; Baklaushev, V.; Filatov, A. Pattern of circulating SARS-CoV-2-specific antibody-secreting and memory B-cell generation in patients with acute COVID-19. Clin. Transl. Immunol. 2021, 10, e1245. [Google Scholar] [CrossRef] [PubMed]
- Dan, J.M.; Mateus, J.; Kato, Y.; Hastie, K.M.; Yu, E.D.; Faliti, C.E.; Grifoni, A.; Ramirez, S.I.; Haupt, S.; Frazier, A.; et al. Immunological memory to SARS-CoV-2 assessed for up to 8 months after infection. Science 2021, 371, eabf4063. [Google Scholar] [CrossRef]
- Grifoni, A.; Weiskopf, D.; Ramirez, S.I.; Mateus, J.; Dan, J.M.; Moderbacher, C.R.; Rawlings, S.A.; Sutherland, A.; Premkumar, L.; Jadi, R.S.; et al. Targets of T Cell Responses to SARS-CoV-2 Coronavirus in Humans with COVID-19 Disease and Unexposed Individuals. Cell 2020, 181, 1489–1501.e15. [Google Scholar] [CrossRef] [PubMed]
- Juno, J.A.; Tan, H.X.; Lee, W.S.; Reynaldi, A.; Kelly, H.G.; Wragg, K.; Esterbauer, R.; Kent, H.E.; Batten, C.J.; Mordant, F.L.; et al. Humoral and circulating follicular helper T cell responses in recovered patients with COVID-19. Nat. Med. 2020, 26, 1428–1434. [Google Scholar] [CrossRef] [PubMed]
- Pušnik, J.; Richter, E.; Schulte, B.; Dolscheid-Pommerich, R.; Bode, C.; Putensen, C.; Hartmann, G.; Alter, G.; Streeck, H. Memory B cells targeting SARS-CoV-2 spike protein and their dependence on CD4+ T cell help. Cell Rep. 2021, 35, 109320. [Google Scholar] [CrossRef]
- Ferreira-Gomes, M.; Kruglov, A.; Durek, P.; Heinrich, F.; Tizian, C.; Heinz, G.A.; Pascual-Reguant, A.; Du, W.; Mothes, R.; Fan, C.; et al. SARS-CoV-2 in severe COVID-19 induces a TGF-β-dominated chronic immune response that does not target itself. Nat. Commun. 2021, 12, 1961. [Google Scholar] [CrossRef]
- Kandikattu, H.K.; Venkateshaiah, S.U.; Kumar, S.; Mishra, A. IL-15 immunotherapy is a viable strategy for COVID-19. Cytokine Growth Factor Rev. 2020, 54, 24–31. [Google Scholar] [CrossRef]
- Wilz, S.W. A clinical trial of IL-15 and IL-21 combination therapy for COVID-19 is warranted. Cytokine Growth Factor Rev. 2021, 58, 49–50. [Google Scholar] [CrossRef]
- Spolski, R.; Leonard, W.J. Interleukin-21: A double-edged sword with therapeutic potential. Nat. Rev. Drug Discov. 2014, 13, 379–395. [Google Scholar] [CrossRef]
- Conlon, K.C.; Miljkovic, M.D.; Waldmann, T.A. Cytokines in the Treatment of Cancer. J. Interferon Cytokine Res. 2019, 39, 6–21. [Google Scholar] [CrossRef]
- Steele, N.; Anthony, A.; Saunders, M.; Esmarck, B.; Ehrnrooth, E.; Kristjansen, P.E.; Nihlén, A.; Hansen, L.T.; Cassidy, J. A phase 1 trial of recombinant human IL-21 in combination with cetuximab in patients with metastatic colorectal cancer. Br. J. Cancer 2012, 106, 793–798. [Google Scholar] [CrossRef] [Green Version]
- Stolfi, C.; Rizzo, A.; Franzè, E.; Rotondi, A.; Fantini, M.C.; Sarra, M.; Caruso, R.; Monteleone, I.; Sileri, P.; Franceschilli, L.; et al. Involvement of interleukin-21 in the regulation of colitis-associated colon cancer. J. Exp. Med. 2011, 208, 2279–2290. [Google Scholar] [CrossRef] [PubMed]
- Jauch, D.; Martin, M.; Schiechl, G.; Kesselring, R.; Schlitt, H.J.; Geissler, E.K.; Fichtner-Feigl, S. Interleukin 21 controls tumour growth and tumour immunosurveillance in colitis-associated tumorigenesis in mice. Gut 2011, 60, 1678–1686. [Google Scholar] [CrossRef] [Green Version]
- Araki, A.; Jin, L.; Nara, H.; Takeda, Y.; Nemoto, N.; Gazi, M.Y.; Asao, H. IL-21 Enhances the Development of Colitis-Associated Colon Cancer: Possible Involvement of Activation-Induced Cytidine Deaminase Expression. J. Immunol. 2019, 202, 3326–3333. [Google Scholar] [CrossRef] [PubMed]
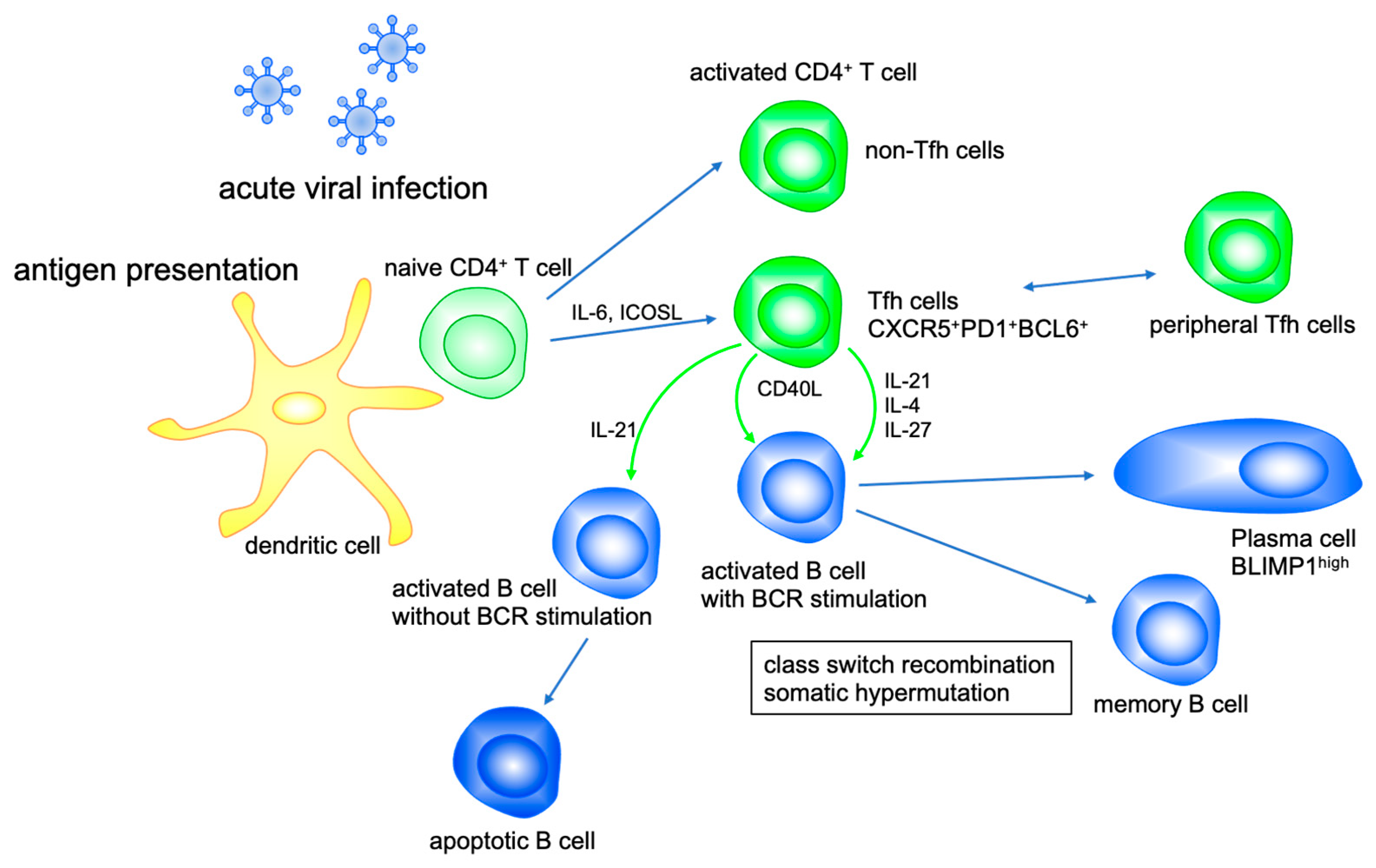
Figure 1.
IL-21 modulates B cell function. Tfh cell activates B cell differentiation with CD40L, IL-21, IL-4 and IL-27. IL-21 promotes class switch recombination to IgG1 and IgG3 and suppresses that to IgE. IL-21 enhances somatic hypermutation and production of high-affinity antibodies by plasma cells, whereas IL-21 induces apoptosis of bystander B cells activated without antigen receptor stimulation.
Figure 1.
IL-21 modulates B cell function. Tfh cell activates B cell differentiation with CD40L, IL-21, IL-4 and IL-27. IL-21 promotes class switch recombination to IgG1 and IgG3 and suppresses that to IgE. IL-21 enhances somatic hypermutation and production of high-affinity antibodies by plasma cells, whereas IL-21 induces apoptosis of bystander B cells activated without antigen receptor stimulation.

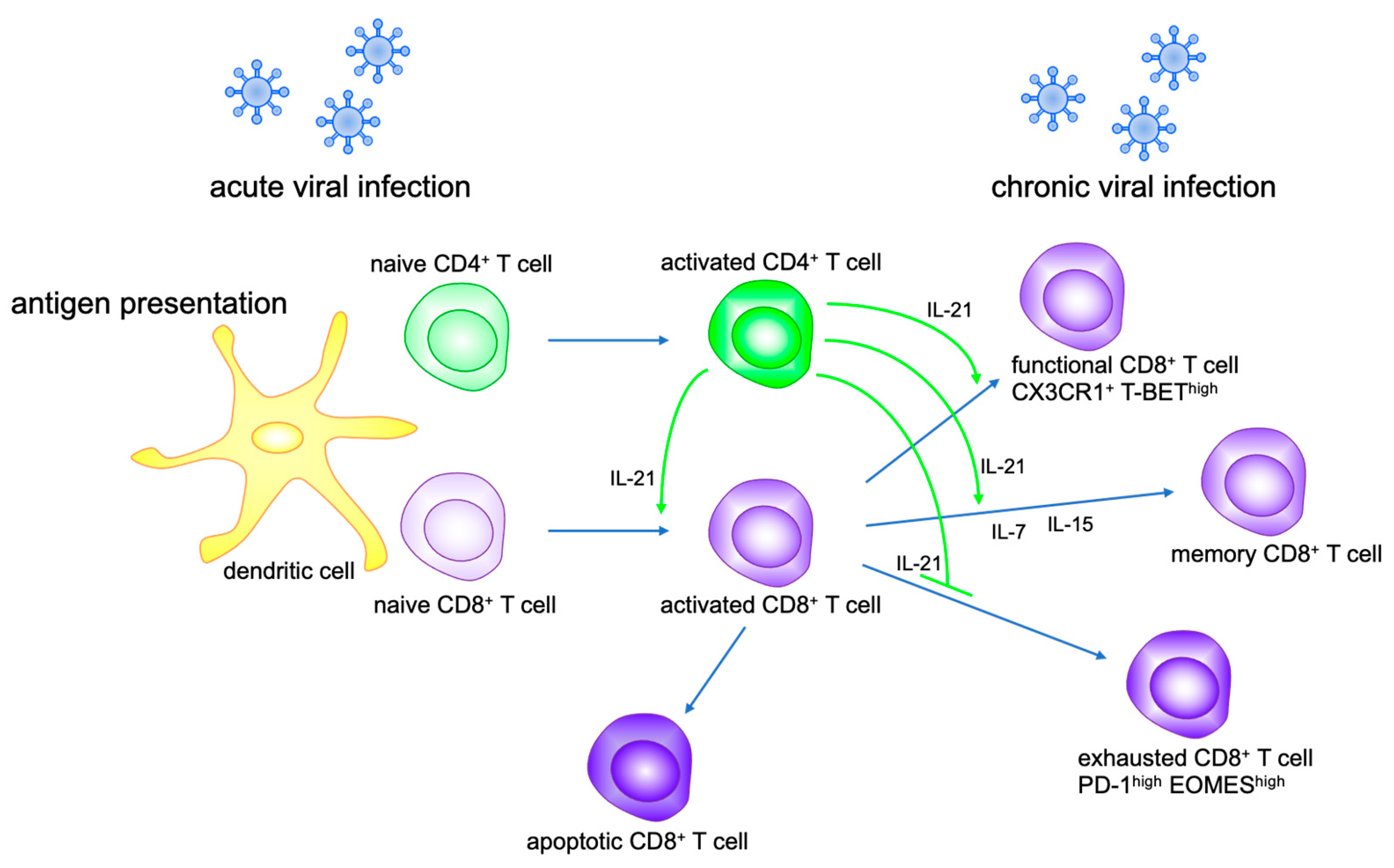
Figure 2.
IL-21 modulates CD8+ T cell function. IL-21 secreted by activated CD4+ T cells promotes CD8+ T cell activation and proliferation, and differentiation to memory CD8+ T cells with IL-7 and IL-15. IL-21 maintains functional CD8+ T cells and prevents CD8+ T exhaustion in chronic viral infection.
Figure 2.
IL-21 modulates CD8+ T cell function. IL-21 secreted by activated CD4+ T cells promotes CD8+ T cell activation and proliferation, and differentiation to memory CD8+ T cells with IL-7 and IL-15. IL-21 maintains functional CD8+ T cells and prevents CD8+ T exhaustion in chronic viral infection.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Asao, H. Interleukin-21 in Viral Infections. Int. J. Mol. Sci. 2021, 22, 9521. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22179521
AMA Style
Asao H. Interleukin-21 in Viral Infections. International Journal of Molecular Sciences. 2021; 22(17):9521. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22179521
Chicago/Turabian StyleAsao, Hironobu. 2021. "Interleukin-21 in Viral Infections" International Journal of Molecular Sciences 22, no. 17: 9521. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22179521
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.