
Physiological Effects of Neonicotinoid Insecticides on Non-Target Aquatic Animals—An Updated Review

 ,
,  , , ,
, , ,  , ,
, ,  and
and 
Abstract
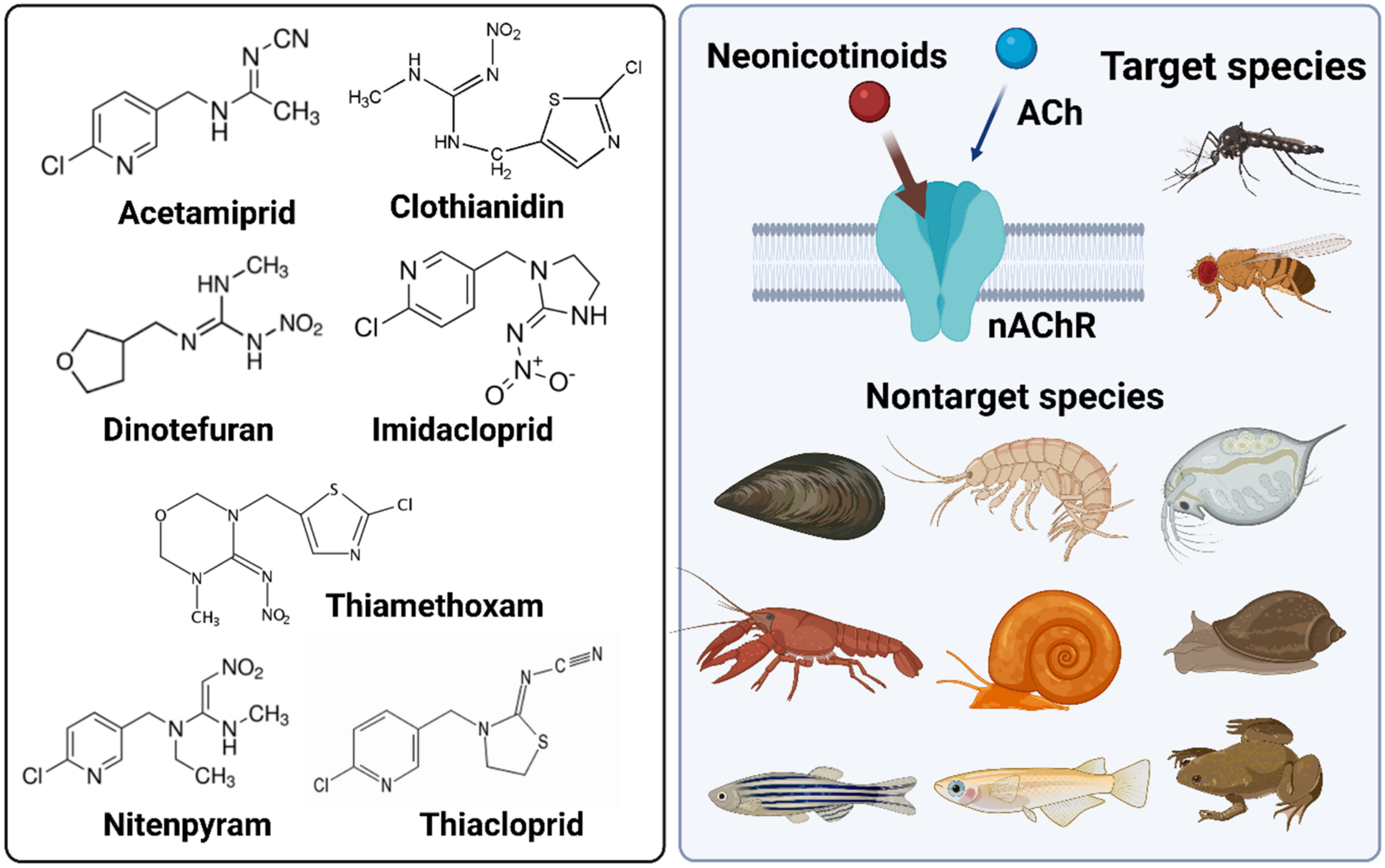
:1. Neonicotinoids—An Overview
2. Chemical and Physical Properties of Neonicotinoids
3. Interaction and Selectivity Mechanisms of Neonicotinoid Insecticides
4. Toxicity of Neonicotinoids towards Aquatic Invertebrates
5. The Potential Adverse Effects of Neonicotinoid Insecticides on Aquatic Vertebrates
6. Future Direction of Work
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Richardson, J.R.; Fitsanakis, V.; Westerink, R.H.; Kanthasamy, A.G. Neurotoxicity of pesticides. Acta Neuropathol. 2019, 138, 343–362. [Google Scholar] [CrossRef]
- Borsuah, J.F.; Messer, T.L.; Snow, D.D.; Comfort, S.D.; Mittelstet, A.R. Literature review: Global neonicotinoid insecticide occurrence in aquatic environments. Water 2020, 12, 3388. [Google Scholar] [CrossRef]
- Mahai, G.; Wan, Y.; Xia, W.; Yang, S.; He, Z.; Xu, S. Neonicotinoid insecticides in surface water from the central Yangtze River, china. Chemosphere 2019, 229, 452–460. [Google Scholar] [CrossRef]
- Hladik, M.L.; Kolpin, D.W.; Kuivila, K.M. Widespread occurrence of neonicotinoid insecticides in streams in a high corn and soybean producing region, USA. Environ. Pollut. 2014, 193, 189–196. [Google Scholar] [CrossRef]
- Goulson, D. An overview of the environmental risks posed by neonicotinoid insecticides. J. Appl. Ecol. 2013, 50, 977–987. [Google Scholar] [CrossRef]
- Jeschke, P.; Nauen, R.; Schindler, M.; Elbert, A. Overview of the status and global strategy for neonicotinoids. J. Agric. Food Chem. 2011, 59, 2897–2908. [Google Scholar] [CrossRef] [PubMed]
- Nauen, R.; Denholm, I. Resistance of insect pests to neonicotinoid insecticides: Current status and future prospects. Arch. Insect Biochem. Physiol. Publ. Collab. Entomol. Soc. Am. 2005, 58, 200–215. [Google Scholar] [CrossRef] [PubMed]
- Vogel, G. Where have all the insects gone? In Science; American Association for the Advancement of Science: Washington, DC, USA, 2017. [Google Scholar]
- Sánchez-Bayo, F.; Wyckhuys, K.A. Worldwide decline of the entomofauna: A review of its drivers. Biol. Conserv. 2019, 232, 8–27. [Google Scholar] [CrossRef]
- Feng, J.; Liu, R.; Chen, P.; Yuan, S.; Zhao, D.; Zhang, J.; Zheng, Z. Degradation of aqueous 3, 4-dichloroaniline by a novel dielectric barrier discharge plasma reactor. Environ. Sci. Pollut. Res. 2015, 22, 4447–4459. [Google Scholar] [CrossRef] [PubMed]
- Bonmatin, J.-M.; Giorio, C.; Sánchez-Bayo, F.; van Lexmond, M.B. An Update of the Worldwide Integrated Assessment (WIA) on Systemic Insecticides; Springer: New York, NY, USA, 2021. [Google Scholar]
- Desneux, N.; Decourtye, A.; Delpuech, J.-M. The sublethal effects of pesticides on beneficial arthropods. Annu. Rev. Entomol. 2007, 52, 81–106. [Google Scholar] [CrossRef]
- Sánchez-Bayo, F.; Goulson, D.; Pennacchio, F.; Nazzi, F.; Goka, K.; Desneux, N. Are bee diseases linked to pesticides?—A brief review. Environ. Int. 2016, 89, 7–11. [Google Scholar] [CrossRef]
- Cimino, A.M.; Boyles, A.L.; Thayer, K.A.; Perry, M.J. Effects of neonicotinoid pesticide exposure on human health: A systematic review. Environ. Health Perspect. 2017, 125, 155–162. [Google Scholar] [CrossRef] [Green Version]
- Pagano, M.; Stara, A.; Aliko, V.; Faggio, C. Impact of neonicotinoids to aquatic invertebrates—In vitro studies on mytilus galloprovincialis: A review. J. Mar. Sci. Eng. 2020, 8, 801. [Google Scholar] [CrossRef]
- Bonmatin, J.-M.; Noome, D.A.; Moreno, H.; Mitchell, E.A.; Glauser, G.; Soumana, O.S.; van Lexmond, M.B.; Sánchez-Bayo, F. A survey and risk assessment of neonicotinoids in water, soil and sediments of belize. Environ. Pollut. 2019, 249, 949–958. [Google Scholar] [CrossRef]
- Bonmatin, J.-M.; Giorio, C.; Girolami, V.; Goulson, D.; Kreutzweiser, D.; Krupke, C.; Liess, M.; Long, E.; Marzaro, M.; Mitchell, E.A. Environmental fate and exposure; neonicotinoids and fipronil. Environ. Sci. Pollut. Res. 2015, 22, 35–67. [Google Scholar] [CrossRef] [PubMed]
- Raby, M.; Nowierski, M.; Perlov, D.; Zhao, X.; Hao, C.; Poirier, D.G.; Sibley, P.K. Acute toxicity of 6 neonicotinoid insecticides to freshwater invertebrates. Environ. Toxicol. Chem. 2018, 37, 1430–1445. [Google Scholar] [CrossRef] [PubMed]
- Basley, K.; Goulson, D. Neonicotinoids thiamethoxam and clothianidin adversely affect the colonisation of invertebrate populations in aquatic microcosms. Environ. Sci. Pollut. Res. 2018, 25, 9593–9599. [Google Scholar] [CrossRef] [Green Version]
- Pisa, L.W.; Amaral-Rogers, V.; Belzunces, L.P.; Bonmatin, J.-M.; Downs, C.A.; Goulson, D.; Kreutzweiser, D.P.; Krupke, C.; Liess, M.; McField, M. Effects of neonicotinoids and fipronil on non-target invertebrates. Environ. Sci. Pollut. Res. 2015, 22, 68–102. [Google Scholar] [CrossRef] [Green Version]
- Starner, K.; Goh, K.S. Detections of the neonicotinoid insecticide imidacloprid in surface waters of three agricultural regions of california, USA, 2010–2011. Bull. Environ. Contam. Toxicol. 2012, 88, 316–321. [Google Scholar] [CrossRef] [PubMed]
- Jeschke, P.; Nauen, R. Neonicotinoids—From zero to hero in insecticide chemistry. Pest Manag. Sci. Former. Pestic. Sci. 2008, 64, 1084–1098. [Google Scholar] [CrossRef]
- Tomizawa, M.; Casida, J.E. Neonicotinoid Insecticides: Highlights of a Symposium on Strategic Molecular Designs; ACS Publications: Washington, DC, USA, 2011. [Google Scholar]
- Casida, J.E.; Durkin, K.A. Neuroactive insecticides: Targets, selectivity, resistance, and secondary effects. Annu. Rev. Entomol. 2013, 58, 99–117. [Google Scholar] [CrossRef]
- Gibbons, D.; Morrissey, C.; Mineau, P. A review of the direct and indirect effects of neonicotinoids and fipronil on vertebrate wildlife. Environ. Sci. Pollut. Res. 2015, 22, 103–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, X.; Li, H.; Xiong, J.; Mehler, W.T.; You, J. Developmental toxicity of a neonicotinoid insecticide, acetamiprid to zebrafish embryos. J. Agric. Food Chem. 2019, 67, 2429–2436. [Google Scholar] [CrossRef] [PubMed]
- Yamamuro, M.; Komuro, T.; Kamiya, H.; Kato, T.; Hasegawa, H.; Kameda, Y. Neonicotinoids disrupt aquatic food webs and decrease fishery yields. Science 2019, 366, 620–623. [Google Scholar] [CrossRef]
- Nauen, R.; Ebbinghaus-Kintscher, U.; Elbert, A.; Jeschke, P.; Tietjen, K. Acetylcholine receptors as sites for developing neonicotinoid insecticides. In Biochemical Sites of Insecticide Action and Resistance; Springer: New York, NY, USA, 2001; pp. 77–105. [Google Scholar]
- Tomizawa, M.; Yamamoto, I. Structure-activity relationships of nicotinoids and imidacloprid analogs. J. Pestic. Sci. 1993, 18, 91–98. [Google Scholar] [CrossRef] [Green Version]
- Tomizawa, M.; Casida, J.E. Selective toxicity of neonicotinoids attributable to specificity of insect and mammalian nicotinic receptors. Annu. Rev. Entomol. 2003, 48, 339–364. [Google Scholar] [CrossRef]
- Jeschke, P.; Nauen, R. Neonicotinoid Insecticides; Elsevier: Amsterdam, The Netherlands, 2005. [Google Scholar]
- Wexler, P.; Anderson, B.D.; Gad, S.C.; Hakkinen, P.B.; Kamrin, M.; De Peyster, A.; Locey, B.; Pope, C.; Mehendale, H.M.; Shugart, L.R. Encyclopedia of Toxicology; Academic Press: Cambridge, MA, USA, 2005; Volume 1. [Google Scholar]
- Seifert, J. Neonicotinoids; Elsevier: Amsterdam, The Netherlands, 2014. [Google Scholar]
- Crossthwaite, A.J.; Bigot, A.; Camblin, P.; Goodchild, J.; Lind, R.J.; Slater, R.; Maienfisch, P. The invertebrate pharmacology of insecticides acting at nicotinic acetylcholine receptors. J. Pestic. Sci. 2017, 42, D17–D19. [Google Scholar] [CrossRef] [Green Version]
- Beck, M.E.; Gutbrod, O.; Matthiesen, S. Insight into the binding mode of agonists of the nicotinic acetylcholine receptor from calculated electron densities. ChemPhysChem 2015, 16, 2760. [Google Scholar] [CrossRef] [Green Version]
- Teralı, K. An evaluation of neonicotinoids’ potential to inhibit human cholinesterases: Protein–ligand docking and interaction profiling studies. J. Mol. Graph. Model. 2018, 84, 54–63. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.-S.; Cheng, H.; Jiang, Y.; Melcher, K.; Xu, H.E. Ion channels gated by acetylcholine and serotonin: Structures, biology, and drug discovery. Acta Pharmacol. Sin. 2015, 36, 895–907. [Google Scholar] [CrossRef] [PubMed]
- Millar, N.S.; Denholm, I. Nicotinic acetylcholine receptors: Targets for commercially important insecticides. Invertebr. Neurosci. 2007, 7, 53–66. [Google Scholar] [CrossRef]
- Houchat, J.-N.; Cartereau, A.; Le Mauff, A.; Taillebois, E.; Thany, S.H. An overview on the effect of neonicotinoid insecticides on mammalian cholinergic functions through the activation of neuronal nicotinic acetylcholine receptors. Int. J. Environ. Res. Public Health 2020, 17, 3222. [Google Scholar] [CrossRef]
- Ihara, M.; Okajima, T.; Yamashita, A.; Oda, T.; Asano, T.; Matsui, M.; Sattelle, D.B.; Matsuda, K. Studies on an acetylcholine binding protein identify a basic residue in loop g on the β1 strand as a new structural determinant of neonicotinoid actions. Mol. Pharmacol. 2014, 86, 736–746. [Google Scholar] [CrossRef] [Green Version]
- Shimomura, M.; Yokota, M.; Ihara, M.; Akamatsu, M.; Sattelle, D.B.; Matsuda, K. Role in the selectivity of neonicotinoids of insect-specific basic residues in loop d of the nicotinic acetylcholine receptor agonist binding site. Mol. Pharmacol. 2006, 70, 1255–1263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siregar, P.; Suryanto, M.E.; Chen, K.H.-C.; Huang, J.-C.; Chen, H.-M.; Kurnia, K.A.; Santoso, F.; Hussain, A.; Ngoc Hieu, B.T.; Saputra, F. Exploiting the freshwater shrimp neocaridina denticulata as aquatic invertebrate model to evaluate nontargeted pesticide induced toxicity by investigating physiologic and biochemical parameters. Antioxidants 2021, 10, 391. [Google Scholar] [CrossRef]
- Sánchez-Bayo, F.; Tennekes, H.A. Time-cumulative toxicity of neonicotinoids: Experimental evidence and implications for environmental risk assessments. Int. J. Environ. Res. Public Health 2020, 17, 1629. [Google Scholar] [CrossRef] [Green Version]
- Rittschof, D.; McClellan-Green, P. Molluscs as multidisciplinary models in environment toxicology. Mar. Pollut. Bull. 2005, 50, 369–373. [Google Scholar] [CrossRef] [PubMed]
- Schuler, T.; Denholm, I.; Jouanin, L.; Clark, S.; Clark, A.; Poppy, G. Population-scale laboratory studies of the effect of transgenic plants on nontarget insects. Mol. Ecol. 2001, 10, 1845–1853. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Bayo, F.; Goka, K. Influence of light in acute toxicity bioassays of imidacloprid and zinc pyrithione to zooplankton crustaceans. Aquat. Toxicol. 2006, 78, 262–271. [Google Scholar] [CrossRef]
- Hayasaka, D.; Suzuki, K.; Nomura, T.; Nishiyama, M.; Nagai, T.; Sánchez-Bayo, F.; Goka, K. Comparison of acute toxicity of two neonicotinoid insecticides, imidacloprid and clothianidin, to five cladoceran species. J. Pestic. Sci. 2013, 38, 44–47. [Google Scholar] [CrossRef] [Green Version]
- Zein, M.A.; McElmurry, S.P.; Kashian, D.R.; Savolainen, P.T.; Pitts, D.K. Optical bioassay for measuring sublethal toxicity of insecticides in daphnia pulex. Environ. Toxicol. Chem. 2014, 33, 144–151. [Google Scholar] [CrossRef]
- Raby, M.; Zhao, X.; Hao, C.; Poirier, D.G.; Sibley, P.K. Chronic effects of an environmentally-relevant, short-term neonicotinoid insecticide pulse on four aquatic invertebrates. Sci. Total Environ. 2018, 639, 1543–1552. [Google Scholar] [CrossRef]
- Barbee, G.C.; Stout, M.J. Comparative acute toxicity of neonicotinoid and pyrethroid insecticides to non-target crayfish (procambarus clarkii) associated with rice–crayfish crop rotations. Pest Manag. Sci. Former. Pestic. Sci. 2009, 65, 1250–1256. [Google Scholar] [CrossRef] [PubMed]
- Van den Brink, P.J.; Van Smeden, J.M.; Bekele, R.S.; Dierick, W.; De Gelder, D.M.; Noteboom, M.; Roessink, I. Acute and chronic toxicity of neonicotinoids to nymphs of a mayfly species and some notes on seasonal differences. Environ. Toxicol. Chem. 2016, 35, 128–133. [Google Scholar] [CrossRef]
- Macaulay, S.J.; Hageman, K.J.; Alumbaugh, R.E.; Lyons, S.M.; Piggott, J.J.; Matthaei, C.D. Chronic toxicities of neonicotinoids to nymphs of the common new zealand mayfly deleatidium spp. Environ. Toxicol. Chem. 2019, 38, 2459–2471. [Google Scholar] [CrossRef] [PubMed]
- Roessink, I.; Merga, L.B.; Zweers, H.J.; Van den Brink, P.J. The neonicotinoid imidacloprid shows high chronic toxicity to mayfly nymphs. Environ. Toxicol. Chem. 2013, 32, 1096–1100. [Google Scholar] [CrossRef]
- Finnegan, M.C.; Baxter, L.R.; Maul, J.D.; Hanson, M.L.; Hoekstra, P.F. Comprehensive characterization of the acute and chronic toxicity of the neonicotinoid insecticide thiamethoxam to a suite of aquatic primary producers, invertebrates, and fish. Environ. Toxicol. Chem. 2017, 36, 2838–2848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rico, A.; Arenas-Sánchez, A.; Pasqualini, J.; García-Astillero, A.; Cherta, L.; Nozal, L.; Vighi, M. Effects of imidacloprid and a neonicotinoid mixture on aquatic invertebrate communities under mediterranean conditions. Aquat. Toxicol. 2018, 204, 130–143. [Google Scholar] [CrossRef]
- Camp, A.; Buchwalter, D. Can’t take the heat: Temperature-enhanced toxicity in the mayfly isonychia bicolor exposed to the neonicotinoid insecticide imidacloprid. Aquat. Toxicol. 2016, 178, 49–57. [Google Scholar] [CrossRef]
- Macaulay, S.J.; Buchwalter, D.B.; Matthaei, C.D. Water temperature interacts with the insecticide imidacloprid to alter acute lethal and sublethal toxicity to mayfly larvae. N. Z. J. Mar. Freshw. Res. 2020, 54, 115–130. [Google Scholar] [CrossRef]
- Macaulay, S.J.; Hageman, K.J.; Piggott, J.J.; Matthaei, C.D. Imidacloprid dominates the combined toxicities of neonicotinoid mixtures to stream mayfly nymphs. Sci. Total Environ. 2021, 761, 143263. [Google Scholar] [CrossRef]
- Bartlett, A.J.; Hedges, A.M.; Intini, K.D.; Brown, L.R.; Maisonneuve, F.J.; Robinson, S.A.; Gillis, P.L.; de Solla, S.R. Acute and chronic toxicity of neonicotinoid and butenolide insecticides to the freshwater amphipod, hyalella azteca. Ecotoxicol. Environ. Saf. 2019, 175, 215–223. [Google Scholar] [CrossRef] [PubMed]
- Bartlett, A.J.; Hedges, A.M.; Intini, K.D.; Brown, L.R.; Maisonneuve, F.J.; Robinson, S.A.; Gillis, P.L.; de Solla, S.R. Lethal and sublethal toxicity of neonicotinoid and butenolide insecticides to the mayfly, hexagenia spp. Environ. Pollut. 2018, 238, 63–75. [Google Scholar] [CrossRef] [PubMed]
- Chandran, N.N.; Fojtova, D.; Blahova, L.; Rozmankova, E.; Blaha, L. Acute and (sub) chronic toxicity of the neonicotinoid imidacloprid on chironomus riparius. Chemosphere 2018, 209, 568–577. [Google Scholar] [CrossRef] [PubMed]
- Wei, F.; Wang, D.; Li, H.; Xia, P.; Ran, Y.; You, J. Toxicogenomics provides insights to toxicity pathways of neonicotinoids to aquatic insect, chironomus dilutus. Environ. Pollut. 2020, 260, 114011. [Google Scholar] [CrossRef] [PubMed]
- Maloney, E.; Liber, K.; Headley, J.; Peru, K.; Morrissey, C. Neonicotinoid insecticide mixtures: Evaluation of laboratory-based toxicity predictions under semi-controlled field conditions. Environ. Pollut. 2018, 243, 1727–1739. [Google Scholar] [CrossRef] [PubMed]
- Cavallaro, M.C.; Morrissey, C.A.; Headley, J.V.; Peru, K.M.; Liber, K. Comparative chronic toxicity of imidacloprid, clothianidin, and thiamethoxam to chironomus dilutus and estimation of toxic equivalency factors. Environ. Toxicol. Chem. 2017, 36, 372–382. [Google Scholar] [CrossRef]
- Barmentlo, S.H.; Vriend, L.M.; van Grunsven, R.H.; Vijver, M.G. Environmental levels of neonicotinoids reduce prey consumption, mobility and emergence of the damselfly ischnura elegans. J. Appl. Ecol. 2019, 56, 2034–2044. [Google Scholar] [CrossRef] [Green Version]
- Raby, M.; Zhao, X.; Hao, C.; Poirier, D.G.; Sibley, P.K. Relative chronic sensitivity of neonicotinoid insecticides to ceriodaphnia dubia and daphnia magna. Ecotoxicol. Environ. Saf. 2018, 163, 238–244. [Google Scholar] [CrossRef]
- Prosser, R.; De Solla, S.; Holman, E.; Osborne, R.; Robinson, S.; Bartlett, A.; Maisonneuve, F.; Gillis, P. Sensitivity of the early-life stages of freshwater mollusks to neonicotinoid and butenolide insecticides. Environ. Pollut. 2016, 218, 428–435. [Google Scholar]
- Maloney, E.; Taillebois, E.; Gilles, N.; Morrissey, C.; Liber, K.; Servent, D.; Thany, S. Binding properties to nicotinic acetylcholine receptors can explain differential toxicity of neonicotinoid insecticides in chironomidae. Aquat. Toxicol. 2021, 230, 105701. [Google Scholar] [CrossRef]
- Ewere, E.E.; Reichelt-Brushett, A.; Benkendorff, K. The neonicotinoid insecticide imidacloprid, but not salinity, impacts the immune system of sydney rock oyster, saccostrea glomerata. Sci. Total Environ. 2020, 742, 140538. [Google Scholar] [CrossRef]
- Butcherine, P.; Kelaher, B.P.; Taylor, M.D.; Lawson, C.; Benkendorff, K. Acute toxicity, accumulation and sublethal effects of four neonicotinoids on juvenile black tiger shrimp (Penaeus monodon). Chemosphere 2021, 275, 129918. [Google Scholar] [CrossRef]
- Butcherine, P.; Kelaher, B.P.; Taylor, M.D.; Barkla, B.J.; Benkendorff, K. Impact of imidacloprid on the nutritional quality of adult black tiger shrimp (penaeus monodon). Ecotoxicol. Environ. Saf. 2020, 198, 110682. [Google Scholar] [CrossRef]
- Hook, S.E.; Doan, H.; Gonzago, D.; Musson, D.; Du, J.; Kookana, R.; Sellars, M.J.; Kumar, A. The impacts of modern-use pesticides on shrimp aquaculture: An assessment for north eastern australia. Ecotoxicol. Environ. Saf. 2018, 148, 770–780. [Google Scholar] [CrossRef]
- Bownik, A.; Pawłocik, M.; Sokołowska, N. Effects of neonicotinoid insecticide acetamiprid on swimming velocity, heart rate and thoracic limb movement of daphnia magna. Pol. J. Nat. Sci. 2017, 32, 481–493. [Google Scholar]
- Takács, E.; Klátyik, S.; Mörtl, M.; Rácz, G.; Kovács, K.; Darvas, B.; Székács, A. Effects of neonicotinoid insecticide formulations and their components on daphnia magna–the role of active ingredients and co-formulants. Int. J. Environ. Anal. Chem. 2017, 97, 885–900. [Google Scholar] [CrossRef]
- Vehovszky, Á.; Farkas, A.; Csikós, V.; Székács, A.; Mörtl, M.; Győri, J. Neonicotinoid insecticides are potential substrates of the multixenobiotic resistance (mxr) mechanism in the non-target invertebrate, dreissena sp. Aquat. Toxicol. 2018, 205, 148–155. [Google Scholar] [CrossRef] [Green Version]
- Stara, A.; Bellinvia, R.; Velisek, J.; Strouhova, A.; Kouba, A.; Faggio, C. Acute exposure of common yabby (cherax destructor) to the neonicotinoid pesticide. Sci. Total Environ. 2019, 665, 718–723. [Google Scholar] [CrossRef]
- Berghiche, H.; Touati, K.; Chouahda, S.; Soltani, N. Impact of the neonicotinoid insecticide, actara®, on the shrimp palaemon adspersus: Biomarkers measurement. In Euro-Mediterranean Conference for Environmental Integration; Springer: Cham, Switzerland, 2017; pp. 533–534. [Google Scholar]
- Al-Badran, A.A.; Fujiwara, M.; Mora, M.A. Effects of insecticides, fipronil and imidacloprid, on the growth, survival, and behavior of brown shrimp farfantepenaeus aztecus. PLoS ONE 2019, 14, e0223641. [Google Scholar] [CrossRef] [Green Version]
- Omar, H.; Samir, H.; Khalil, M.S.; Ghorab, M.A.; Zwiernik, M.J. Acute Water Column Effects Concentrations (lc50, lc90) for Three Commonly Used Insecticides, Two Neonicotinoids (Acetamiprid and Imidacloprid), and a Recently Registered Phenylpyrazole (Fipronil), Exposed to Common Commercially Cultured Shrimp (Marsupenaeus japonicus); Michigan State University: East Lansing, MI, USA, 2016. [Google Scholar]
- Stara, A.; Pagano, M.; Capillo, G.; Fabrello, J.; Sandova, M.; Vazzana, I.; Zuskova, E.; Velisek, J.; Matozzo, V.; Faggio, C. Assessing the effects of neonicotinoid insecticide on the bivalve mollusc mytilus galloprovincialis. Sci. Total Environ. 2020, 700, 134914. [Google Scholar] [CrossRef]
- Vehovszky, Á.; Farkas, A.; Ács, A.; Stoliar, O.; Székács, A.; Mörtl, M.; Győri, J. Neonicotinoid insecticides inhibit cholinergic neurotransmission in a molluscan (Lymnaea stagnalis) nervous system. Aquat. Toxicol. 2015, 167, 172–179. [Google Scholar] [CrossRef] [Green Version]
- Contardo-Jara, V.; Gessner, M.O. Uptake and physiological effects of the neonicotinoid imidacloprid and its commercial formulation confidor® in a widespread freshwater oligochaete. Environ. Pollut. 2020, 264, 114793. [Google Scholar] [CrossRef]
- Hano, T.; Ito, K.; Ohkubo, N.; Ito, M.; Watanabe, A.; Sakaji, H. Involvement of oxygenase confers higher resistance to neonicotinoid insecticides in estuarine resident sand shrimp crangon uritai than in kuruma prawn penaeus japonicus and mysid americamysis bahia. Fish. Sci. 2020, 86, 1079–1086. [Google Scholar] [CrossRef]
- De Esch, C.; Slieker, R.; Wolterbeek, A.; Woutersen, R.; de Groot, D. Zebrafish as potential model for developmental neurotoxicity testing: A mini review. Neurotoxicol. Teratol. 2012, 34, 545–553. [Google Scholar] [CrossRef]
- Aleström, P.; Holter, J.L.; Nourizadeh-Lillabadi, R. Zebrafish in functional genomics and aquatic biomedicine. Trends Biotechnol. 2006, 24, 15–21. [Google Scholar] [CrossRef]
- Padilla, S.; Cowden, J.; Hinton, D.E.; Yuen, B.; Law, S.; Kullman, S.W.; Johnson, R.; Hardman, R.C.; Flynn, K.; Au, D.W. Use of medaka in toxicity testing. Curr. Protoc. Toxicol. 2009, 39, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Luo, T.; Wang, X.; Jin, Y. Low concentrations of imidacloprid exposure induced gut toxicity in adult zebrafish (danio rerio). Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2021, 241, 108972. [Google Scholar] [CrossRef] [PubMed]
- Ge, W.; Yan, S.; Wang, J.; Zhu, L.; Chen, A.; Wang, J. Oxidative stress and DNA damage induced by imidacloprid in zebrafish (Denio rerio). J. Agric. Food Chem. 2015, 63, 1856–1862. [Google Scholar] [CrossRef] [PubMed]
- Crosby, E.B.; Bailey, J.M.; Oliveri, A.N.; Levin, E.D. Neurobehavioral impairments caused by developmental imidacloprid exposure in zebrafish. Neurotoxicol. Teratol. 2015, 49, 81–90. [Google Scholar] [CrossRef] [Green Version]
- Krishnan, A.; Clyburn, C.; Newcombe, P. Developmental effects of the pesticide imidacloprid on zebrafish body length and mortality. bioRxiv 2018, 395327. [Google Scholar] [CrossRef] [Green Version]
- Toğay, V.A.; Türel, G.Y.; Çelik, D.A.; Özgöçmen, M.; Tülüceoğlu, E.E.; Şen, İ.; Ayvaz, Y. DNA damage effect of cyprodinil and thiacloprid in adult zebrafish gills. Environ. Sci. Pollut. Res. 2021, 28, 14482–14487. [Google Scholar] [CrossRef]
- Yan, S.H.; Wang, J.H.; Zhu, L.S.; Chen, A.M.; Wang, J. Thiamethoxam induces oxidative stress and antioxidant response in zebrafish (Denio rerio) livers. Environ. Toxicol. 2016, 31, 2006–2015. [Google Scholar] [CrossRef]
- Yan, S.; Wang, J.; Zhu, L.; Chen, A.; Wang, J. Toxic effects of nitenpyram on antioxidant enzyme system and DNA in zebrafish (Danio rerio) livers. Ecotoxicol. Environ. Saf. 2015, 122, 54–60. [Google Scholar] [CrossRef]
- Wang, Y.; Lv, L.; Yu, Y.; Yang, G.; Xu, Z.; Wang, Q.; Cai, L. Single and joint toxic effects of five selected pesticides on the early life stages of zebrafish (Denio rerio). Chemosphere 2017, 170, 61–67. [Google Scholar] [CrossRef]
- Vignet, C.; Cappello, T.; Fu, Q.; Lajoie, K.; De Marco, G.; Clérandeau, C.; Mottaz, H.; Maisano, M.; Hollender, J.; Schirmer, K. Imidacloprid induces adverse effects on fish early life stages that are more severe in japanese medaka (Oryzias latipes) than in zebrafish (Denio rerio). Chemosphere 2019, 225, 470–478. [Google Scholar] [CrossRef] [PubMed]
- Vignet, C.; Lajoie, K.; Maisano, M.; Mottaz, H.; Clérandeau, C.; Cappello, T.; Schirmer, K.; Cachot, J. Response of Zebrafish and Japanese Medaka to Imidacloprid: A Comparative Study. In Proceedings of the SETAC Europe 27th Annual Meeting, Brussels, Belgium, 7–11 May 2017; Society of Environmental Toxicology and Chemistry Europe (SETAC Europe): Brussels, Belgium, 2017; p. 298. [Google Scholar]
- Hong, X.; Zhao, X.; Tian, X.; Li, J.; Zha, J. Changes of hematological and biochemical parameters revealed genotoxicity and immunotoxicity of neonicotinoids on chinese rare minnows (Gobiocypris raru). Environ. Pollut. 2018, 233, 862–871. [Google Scholar] [CrossRef] [PubMed]
- Tian, X.; Yang, W.; Wang, D.; Zhao, Y.; Yao, R.; Ma, L.; Ge, C.; Li, X.; Huang, Z.; He, L. Chronic brain toxicity response of juvenile chinese rare minnows (gobiocypris rarus) to the neonicotinoid insecticides imidacloprid and nitenpyram. Chemosphere 2018, 210, 1006–1012. [Google Scholar] [CrossRef]
- Li, X.; Mao, L.; Zhang, Y.; Wang, X.; Wang, Y.; Wu, X. Joint toxic impacts of cadmium and three pesticides on embryonic development of rare minnow (Gobiocypris rarus). Environ. Sci. Pollut. Res. 2020, 27, 36596–36604. [Google Scholar] [CrossRef]
- Paquet-Walsh, A.; Bertolo, A.; Landry, C.; Deschamps, L.; Boily, M. Interactive effects of neonicotinoids and natural ultraviolet radiation on yellow perch (Perca flavescens) larvae. Sci. Total Environ. 2019, 685, 690–701. [Google Scholar] [CrossRef]
- Marlatt, V.L.; Leung, T.Y.G.; Calbick, S.; Metcalfe, C.; Kennedy, C. Sub-lethal effects of a neonicotinoid, clothianidin, on wild early life stage sockeye salmon (Oncorhynchus nerka). Aquat. Toxicol. 2019, 217, 105335. [Google Scholar] [CrossRef]
- Iturburu, F.G.; Zömisch, M.; Panzeri, A.M.; Crupkin, A.C.; Contardo-Jara, V.; Pflugmacher, S.; Menone, M.L. Uptake, distribution in different tissues, and genotoxicity of imidacloprid in the freshwater fish australoheros facetus. Environ. Toxicol. Chem. 2017, 36, 699–708. [Google Scholar] [CrossRef] [PubMed]
- Houndji, M.A.; Imorou Toko, I.; Guedegba, L.; Yacouto, E.; Agbohessi, P.T.; Mandiki, S.N.; Scippo, M.-L.; Kestemont, P. Joint toxicity of two phytosanitary molecules, lambda-cyhalothrin and acetamiprid, on african catfish (Clarias gariepinus) juveniles. J. Environ. Sci. Health Part B 2020, 55, 669–676. [Google Scholar] [CrossRef]
- Robinson, S.; Richardson, S.; Dalton, R.; Maisonneuve, F.; Bartlett, A.; de Solla, S.; Trudeau, V.; Waltho, N. Assessment of sublethal effects of neonicotinoid insecticides on the life-history traits of 2 frog species. Environ. Toxicol. Chem. 2019, 38, 1967–1977. [Google Scholar] [CrossRef]
- Keller, P.G. Detection of Neonicotinoids in Northern Leopard Frog (Rana pipiens) Brains; University of South Dakota: Vermillion, SD, USA, 2021. [Google Scholar]
- Robinson, S.A.; Chlebak, R.J.; Young, S.D.; Dalton, R.L.; Gavel, M.J.; Prosser, R.S.; Bartlett, A.J.; de Solla, S.R. Clothianidin alters leukocyte profiles and elevates measures of oxidative stress in tadpoles of the amphibian, rana pipiens. Environ. Pollut. 2021, 284, 117149. [Google Scholar] [CrossRef] [PubMed]
- Lee-Jenkins, S.S.; Robinson, S.A. Effects of neonicotinoids on putative escape behavior of juvenile wood frogs (Lithobates sylvaticus) chronically exposed as tadpoles. Environ. Toxicol. Chem. 2018, 37, 3115–3123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saka, M.; Tada, N. Acute and chronic toxicity tests of systemic insecticides, four neonicotinoids and fipronil, using the tadpoles of the western clawed frog silurana tropicalis. Chemosphere 2021, 270, 129418. [Google Scholar] [CrossRef]
- Tian, X.; Hong, X.; Yan, S.; Li, X.; Wu, H.; Lin, A.; Yang, W. Neonicotinoids caused oxidative stress and DNA damage in juvenile chinese rare minnows (Gobiocypris rarus). Ecotoxicol. Environ. Saf. 2020, 197, 110566. [Google Scholar] [CrossRef]
- Vieira, C.E.D.; Pérez, M.R.; Acayaba, R.D.A.; Raimundo, C.C.M.; dos Reis Martinez, C.B. DNA damage and oxidative stress induced by imidacloprid exposure in different tissues of the neotropical fish prochilodus lineatus. Chemosphere 2018, 195, 125–134. [Google Scholar] [CrossRef]
- Velisek, J.; Stara, A. Effect of thiacloprid on early life stages of common carp (Cyprinus carpio). Chemosphere 2018, 194, 481–487. [Google Scholar] [CrossRef]


{kind=link}
| Type of Neonicotinoid | Species | Concentration and Exposure Time | Biological Effects | Reference | ||
|---|---|---|---|---|---|---|
| Crustaceans | ||||||
| Imidacloprid | Neocaridina denticulata | 0.03125, 0.0625, 0.125, 0.25, 0.5, and 1 ppm EC50 (96 h)—0.51 ppm | Reduced locomotor activity, heartbeat, and gill ventilation rate | [42] | ||
| MOSPILAN 20 SP AI: Acetamiprid | Daphnia magna | 25, 50 and 100 ppm 2–72 h LC50 (48 h)—49.8 ppm | Acetamiprid is a potent neuromodulator altering behavioral and physiological parameters of Daphnia magna | [73] | ||
| Imidacloprid, Clothianidin | Moina macrocopa, Daphnia pulex, Daphnia magna, Crenicichla reticulata, and Ceriodaphnia dubia, | 48-h acute immobilization tests EC50 (48 h) ppb | Clothianidin was observed to be four times less toxic than imidacloprid | [47] | ||
| IMI | Cl | |||||
| C. dubia | 571.62 | 1691.3 | ||||
| C. reticulata | 5552.9 | 29,474 | ||||
| D.magna | 43,265 | 67,564 | ||||
| D. pulex | 36,872 | 31,448 | ||||
| M. macrocopa | 45,271 | 61,106 | ||||
| Imidacloprid | Daphnia pulex | 0, 0.02, 0.04, 0.01 and 0.02 ppm, 90 min | Concentration-dependent behavioral effects at sub-lethal concentration and insecticide with the similar mode of action yield comparable results | [48] | ||
| Thiacloprid, Thiamethoxam, Clothianidin | Daphnia magna | 0–160 ppm, 48 h | ACTARA 240 SC® and CALYPSO 480 SC® were thrice less toxic than their active ingredients. APACHE 50 WG® was 46.5 times more toxic than its active ingredients Indication of probable antagonistic/synergistic interaction with the active ingredients | [74] | ||
| EC50 (ppm) | ||||||
| TLC | 5–13.5 | |||||
| TMC | 93–159 | |||||
| APACHE 50 WG®—AI: clothianidin CALYPSO 480 SC®—AI: thiacloprid, ACTARA 240 SC®—AI: thiamethoxam | CLO | 340 | ||||
| APACHE | 11.43 ± 3.74 | |||||
| CALYPSO | 27 ± 9.45 | |||||
| ACTARA | 226.72 ± 68.2 | |||||
| Imidacloprid, Thiamethoxam | Hexagenia spp., Hyalella azteca, Neocloeon triangulifer and Chironomus dilutus | 2.5, 5, and 10 ppb 24 h | Imidacloprid at ~9 ppb caused toxicity impact due to short-term pulse in sensitive insect spp. No persistent impact on test organisms after cessation of stressor | [49] | ||
| Hexagenia spp. EC50 (96 h) < 50 ppb N. triangulifer EC50 96 h < 10 ppb | ||||||
| Imidacloprid, Thiacloprid, Thiamethoxam | Plea minutissima, Caenis horaria, Cloeon dipterum Chaoborus obscuripes Asellus aquaticus Gammarus pulex | Acute: 0.3, 1, 3, 10, 30, 100, 300 ppm—24, 48, 72, and 96 h | Thiamethoxam and imidacloprid depicted comparable acute and chronic toxicity to C. dipterum winter generation; however, thiacloprid was observed two times as toxic | [51] | ||
| EC50 (ppm) | ||||||
| IMI | 18 | |||||
| TLC | 10 | |||||
| TMC | 20 | |||||
| Chronic: 0.01, 0.03, 0.1, 0.3, 1, 3 ppm Day 7, 14, 21, and 28 | ||||||
| EC50 (ppm) | ||||||
| IMI | 0.68 | |||||
| TLC | 0.29 | |||||
| TMC | 0.68 | |||||
| Imidacloprid, Clothianidin, Thiamethoxam. | Mayfly Deleatidium spp. | 0 to 4 ppb 28-d | Clothianidin and imidacloprid depicted strong chronic toxicity impact on Deleatidium nymphs | [52] | ||
| EC50 (ppb) | ||||||
| IMI | 0.19 | |||||
| CLO | 1.02 | |||||
| TMC | >4 | |||||
| Imidacloprid | Deleatidium spp. Coloburiscus humeralis | 9, 12, 15, 18, 21 and 24 °C 96 h | Survivor-ship of mayflies was synergistically decreased by combination of increasing temperatures and exposure to imidacloprid | [57] | ||
| EC50 (ppb) | ||||||
| Deleatidium | 8 | |||||
| Coloburiscus | 12.5 | |||||
| Thiamethoxam | Chironomus dilutus, Daphnia magna, Chironomus riparius, Chaoborus sp. | ≥80 ppm 24–48 h EC50 < 1 ppm | Invertebrates are highly sensitive, but existing environment concentrations are doubtful to surpass our determined Hazard conc. (HC5s) | [54] | ||
| Imidacloprid | Isonychia bicolor | 0.2, 1, 5, 25, 250 ppb 15, 18, 21, and 24 °C 1,4,7,10 days EC50 (96 h) 5.81 ppb | Temperature depicted highly modifying effect on aquatic insects’ toxicity | [56] | ||
| Imidacloprid | Macrocrustaceans Insects | Acute test: Macrocrustaceans 10, 30, 100, 300, 1000 ppb Insects: 1, 10, 30, 100, 300 ppb—4 days | Caddisfly and mayfly were highly sensitive to short-term exposure to imidacloprid. After long-term exposure 28 d of imidacloprid to arthropods the sensitivity value detected was (28-d EC10 = 0.03 ppb) | [53] | ||
| Chronic test: Macrocrustaceans 1, 3, 10, 30, 100 ppb; Insects: 0.3, 1, 3, 10, 30 ppb—28 days | ||||||
| Macrocrustaceans EC50—28 d—11.9, 15.4 ppb Insects EC50 28 d- 11.8, 3.46, 6.45, 0.12 ppb | ||||||
| Thiamethoxam, Acetamiprid, Imidacloprid Thiacloprid, Clothianidin, Dinotefuran | Hyalella azteca | Acute 7 d Chronic 28 d IMI and TMC 8–500 ppb ACT and CLO 0.08–5 ppb TLC and DFN 3–200 ppb FPF 0.6–40 ppb | The growth and survival of Hyalella azteca were altered after exposure to tested six neonicotinoids, with different toxicity amongst compounds | [59] | ||
| Acute EC50 (ppb) | Chronic EC50 (ppb) | |||||
| CLO | 4.0 | 3.5 | ||||
| ACT | 4.7 | 3.4 | ||||
| DFN | 60 | 30 | ||||
| TLC | 68 | 4.2 | ||||
| IMI | 230 | 4.3 | ||||
| TMC | 290 | 200 | ||||
| Thiamethoxam, Imidacloprid, Clothianidin, Acetamiprid, Dinotefuran, Thiacloprid | Hexagenia | Acute (96-h) 21-day clean water | Acute acetamiprid and thiacloprid caused persistent impacts; imidacloprid impacts were detected at environmental pertinent concentration | [60] | ||
| EC50 (ppb) | ||||||
| TMC | 630 | |||||
| IMI | 20 | |||||
| CLO | 24 | |||||
| ACT | 4.0 | |||||
| DFN | 82 | |||||
| TLC | 9.1 | |||||
| Imidacloprid | Chironomus riparius | Acute test 24-h (0.625, 201 1.25, 2.5, 5.0, 10, 20, 40 and 80 ppb) EC50 31.5 ppb Sub-chronic test 10 days 0.625, 1.25, 2.5, 5.0, and 10 ppb EC50 2.33 ppb Chronic test 28 days (0.0625, 0.125, and 0.625 ppb) EC50 3.11 ppb | Imidacloprid repressed larvae growth and affected emergence Effects on reduced/oxidized glutathione and oxidative stress were detected | [61] | ||
| Imidacloprid | Chironomus dilutus | 0.001, 0.01, 0.1, 0.4, 1, 2, 8, 40, and 80 ppb EC50 (96 h) 0.68 ppb 96 h | Death of organisms caused by DNA damage and oxidative stress | [62] | ||
| Clothianidin, Imidacloprid, Thiamethoxam | Limnocorrals | CLO (single compound = 0.71 ppb; in binary mixtures = 0.36 ppb), IMI (single compound = 0.50 ppb; in binary mixtures = 0.25 ppb), TMC (single compound = 8.91 ppb; in binary mixtures = 4.46 ppb)—28- and 56-days | Collective Chironomidae emergence and biomass difference was insignificant among control and neonicotinoid treatments groups on day 28 However, impact on collective biomass and emergence were substantial for IMI, CLO, and the CLO-TMX mixture at day 56 | [63] | ||
| EC50 (ppb) | ||||||
| CLO | 1.03 | |||||
| IMI | 1.03 | |||||
| TMC | 1.04 | |||||
| Clothianidin, Imidacloprid, Thiamethoxam | Chironomus dilutus | 0 ppb (control), 0.1, 0.3, 1.0, 3.3, and 10.0 ppb, 40 days | Clothianidin and imidacloprid exert similar toxicity to C. dilutus, Thiamethoxam induced analogous impact only at high concentrations | [64] | ||
| EC50 (ppb) | ||||||
| CLO | 0.28 | |||||
| IMI | 0.39 | |||||
| TMC | 4.13 | |||||
| Thiamethoxam, Clothianidin, Thiacloprid, Imidacloprid | Dreissena bugensis | 1, 10 ppm | Augmentation of chemostimulation, building up progressively in the organisms exposed to thiamethoxam clothianidin and imidacloprid | [75] | ||
| Thiacloprid | Ischnura elegans | 0 (control), 0.1, 1 and 10 ppb 40 days EC50 1.04 ppb | Environmental pertinent thiacloprid concentrations considerably decline I. elegans emergence | [65] | ||
| Dinotefuran, Clothianidin, Imidacloprid, Thiamethoxam, Acetamiprid, Thiacloprid | Ceriodaphnia dubia, Daphnia magna | 100, 50, 25, 12.5, 6.25, 3.12, 1.56 ppm— 7 days 50, 25, 12.5, 6.25, 3.12, 1.56, 0.78 ppm—21 days | Neonicotinoids depicted chronic toxicity to C. dubia and D. magna at > 1 ppm | [49] | ||
| EC50 (ppm) | ||||||
| D. magna | IMI | 4.59 | ||||
| C. dubia | ACT | 12.95 | ||||
| CLO | 14.52 | |||||
| IMI | 2.98 | |||||
| TLC | 2.06 | |||||
| CALYPSO 480 SC (CAL) AI: thiacloprid | Cherax destructor | 0.1, 0.5, 1, 5, 10, 25, and 50 ppm 24 h, 48 h, 72 h, and 96 h LC50 (96 h) 7.7 ppm | Antioxidant enzyme activity demonstrated considerable alteration in hepatopancreatic tissues | [76] | ||
| Clothianidin, Imidacloprid, Thiamethoxam | Chironomus riparius and Chironomus dilutus (larva and adult) | C. riparius: CLO = 100–217,000 ppm IMI = 100–217,000 ppm TMC = 640–38,230 ppm C. dilutus: CLO = 3400–24,000 ppm IMI = 3400–24,000 ppm TMC = 530–3510 ppm | Binding affinity varied depending on life stage and type of neonicotinoid competitor Differential neonicotinoid toxicity in insects is driven by nicotinic receptor binding | [68] | ||
| EC50 (ppb) | ||||||
| C. riparius | IMI | 12.94 | ||||
| CLO | 21.80 | |||||
| TMC | 55.50 | |||||
| C. dilutus | IMI | 4.63 | ||||
| CLO | 3.30 | |||||
| TMC | 45.0 | |||||
| Thiamethoxam, Clothianidin, Acetamiprid, Imidacloprid | Penaeus monodon | 5 ppb Uptake (8 days) elimination (4 days) | Depuration lowered tissue concentration of the AIs and decreased the activity of oxidative stress enzymes | [70] | ||
| LC50 (ppb) 48 h | ||||||
| TMC | 390 | |||||
| CLO | 190 | |||||
| ACT | >500 | |||||
| IMI | 408 | |||||
| Imidacloprid | Penaeus monodon | Acute conc. low 5 ppb—4d Chronic conc. high 30 ppb—21d | Chronic exposure to imidacloprid resulted in a substantial decrement in total lipid content and body weight. Composition of Fatty acid was altered in exposed shrimp compared to control | [71] | ||
| Imidacloprid | Penaeus monodon | 1, 10, 100, and 1000 ppb 48 h | Imidacloprid exposure decreased post-larval shrimp ability to seize live prey at environment pertinent conc. | [72] | ||
| EC50 ppb | ||||||
| IMI | 175 | |||||
| ACTARAAI: thiamethoxam | Palaemon adspersus | 0.5, 1, 2, 3, 4 and 5 ppm— 96 h | Thiamethoxam depicted a sensitive toxicity to shrimp at sub-lethal concentrations | [77] | ||
| Imidacloprid | Farfantepenaeus aztecus | 0.0, 0.5, 1.0, 15.0, 34.5, 320.0 ppb 36 days | F. aztecus exhibited less lethal effects on imidacloprid | [78] | ||
| Acetamiprid, Imidacloprid | Marsupenaeus japonicas | 50, 100, 200 and 400 ppm 48, 72 and 96 h | After 96 h of exposure acetamiprid showed least mortality | [79] | ||
| LC50 (ppm) 96 h | ||||||
| ACT | 214.33 | |||||
| IMI | 141.42 | |||||
| Molluscs | ||||||
| CALYPSO 480 SC (CAL) AI: thiacloprid | Mytilus galloprovincialis | 1, 10 and 100 ppm 20 days exposure 10 days recovery | Sub-chronic exposure to the neonicotinoid insecticide caused significant alterations in cell and tissue parameters | [80] | ||
| 96 h LC50—7.77 ppm | ||||||
| Imidacloprid, Clothianidin, Thiamethoxam | Planorbella pilsbryi Lampsilis fasciola | 7 days 10, 50, 100, 500, 1000, 5000, and 10,000 ppb or 28 days 10, 50, 100, 500, and 1000 ppb EC50 33.2 to 122.0 ppb | Growth was sensitive endpoint of exposure in comparison to mortality for juvenile snails | [67] | ||
| MOSPILAN, AI: acetamiprid, KOHINOR, AI: imidacloprid, APACS, AI: clothianidin, ACTARA, AI: thiamethoxam, CALYPSO, AI: thiacloprid, | Lymnaea stagnalis | 10–1000 ppm | Thiacloprid at 10 ppm was able to block almost 90% of excitatory post-synaptic potentials (EPSPs), whereas thiamethoxam 100 ppm lowered the synaptic responses by about 15% | [81] | ||
| Imidacloprid | Saccostrea glomerata | 0.01, 0.1, and 1 ppm 4 days | Imidacloprid causes stress at <0.1 ppm No synergistic impact of imidacloprid was observed with reduced salinity | [69] | ||
| Annelids | ||||||
| Imidacloprid and Commercial formulation CONFIDOR | Lumbriculus variegatus | 0.1, 1 and 10 IMI ppb 24 h and 5 d LC50 (24 h)—65 (IMI) and 88 (CONFIDOR) ppb | Activities of studied enzymes suggest imidacloprid exposure cause oxidative stress at environment relevant levels | [82] | ||
| Combo species tests | ||||||
| Clothianidin, Acetamiprid, Dinotefuran, Thiacloprid, Imidacloprid, Thiamethoxam | Lab cultured spp. Daphnia magna, Chironomus dilutus, Ceriodaphnia dubia, Hyalella azteca, Hexagenia spp., Neocloeon triangulifer and Lumbriculus variegatus Field collected spp. Ephemeroptera, Trichoptera, Coleoptera, Isopoda, Hemiptera, Odonata, Diptera, Plecoptera, Agnetina sp. and Paragnetina sp. | Target compounds in aqueous environmental matrices were measured without sample concentration by direct aqueous injection (injection volume of 90 µL), and where results exceeded the calibration range of an analyte 0.5–2 ppb for different analytes 48 h–96 h | Most sensitive insects Chironomus dilutus and Neocloeon triangulifer. Whereas Ceriodaphnia dubia and Daphnia magna were the least sensitive. Neonicotinoids except imidacloprid showed no harmful effect in terms of acute toxicity Imidacloprid, was found hazardous on invertebrate immobilization, and not lethality | [18] | ||
| Imidacloprid Mix. of five Neonicotinoids Imidacloprid, Thiacloprid, Clothianidin, Acetamiprid, Thiamethoxam | Macroinvertebrates—molluscs (5 taxa), insects (26 taxa), platyhelminthes (2 taxa), arachnid (1 taxon), annelids (3 taxa), crustacean (1 taxon) | (0.2, 1, 5, 25, 250 ppb) 1, 4, 7, 10 days (−20 °C) | Temperature emerged main environment factor affecting the sensitivity of invertebrates-neonicotinoid contamination | [55] | ||
| Acetamiprid, Clothianidin | Crangon uritai Penaeus japonicas Americamysis bahia | Crangon uritai 96-h LC50 ACT: 4500 ppb CLO: 360 ppb Penaeus japonicus 96-h LC50 ACT: 85 ppb CLO: 89 ppb Americamysis bahia 96-h LC50 ACT: 24 ppb CLO: 51 ppb | Treatments with the neonicotinoids and oxygenase inhibitor revealed increase in mortality in Crangon uritai but not in Penaeus japonicas and Americamysis bahia. It was concluded that oxygenase might interpret the high resistance of sand shrimp to neonicotinoid insecticides | [83] | ||
| Type of Neonicotinoid | Species | Concentration and Time | Biological Effects | Reference | |
|---|---|---|---|---|---|
| Fish | |||||
| Acetamiprid | Danio rerio | Embryo mortality and malformation Time checkpoints: 120 hpf Conc. of ACT: (54, 107, 263, 374, 433, 537, 644, 760, 848, and 974 ppm) Mortality EC50—518 ppm Malformation EC50—323 ppm (120 hpf) Embryo heart rate Time checkpoints: 48, 60, and 72 hpf Conc. of ACT: 107, 537, and 760 ppm Growth of zebrafish Time checkpoints: 120 hpf Conc. of ACT: 54, 107, 263, 374, and 433 ppm Embryo behaviors Time checkpoints: hourly between 17 and 27 hpf Conc. of ACT: 107, 537, 760, and 974 ppm Touch response Time checkpoint: 27, 36, and 48 hpf Conc. of ACT: 107, 537, 760, and 974 ppm Tail touch EC50—888 ppm Head touch response EC50—754 ppm (48 hpf) | Zebrafish embryos exhibited significant mortality (120 hpf) at 374 ppm in comparison to control groups p < 0.05, with absolute mortality at 760 ppm Acetamiprid caused different embryonic defects, namely, uninflated swim bladder, bent spine, yolk sac edema and pericardial edema Acetamiprid majorly decreased heart rate of zebrafish embryos at 48, 60, and 72 hpf for all treatments Body length of larval fish followed a dose-response relationship | [26] | |
| Imidacloprid | Danio rerio | 100 and 1000 ppb for 21 days | IMI at low concentration indicated toxicity in gut of adult zebrafish | [87] | |
| 300, 1250, and 5000 ppm 7, 14, 21, and 28 days | Imidacloprid induced oxidative stress and DNA damage in zebrafish | [88] | |||
| 2.79 or 3.72 ppm 4 h to 5 d after fertilization | Imidacloprid considerably reduced swimming activity in zebrafish | [89] | |||
| 0, 100, 1,000, and 10,000 ppb 1,2,3, 4, 5 dpf | Increased embryo mortality, and impairment of body length in a dose-dependent association to the imidacloprid concentration | [90] | |||
| Thiamethoxam | Danio rerio (livers) | 0.30, 1.25, and 5.00 ppm 7, 14, 21 and 28 days | Thiamethoxam could induce DNA damage and oxidative stress on the treated zebrafish | [92] | |
| Nitenpyram | Danio rerio (livers) | 0.6, 1.2, 2.5, and 5.0 ppm 28 d | Nitenpyram exposure affected the DNA damage and antioxidant enzymes activity in the zebrafish livers | [93] | |
|
Cyprodinil, Thiacloprid | Danio rerio | CY 0.31 and 0.155 ppm TLC 1.64 and 0.82 ppm 21 d | Cyprodinil and thiacloprid were identified as genotoxic agents damaging DNA | [91] | |
| Phoxim, Atrazine, Butachlor λ-cyhalothrin | Danio rerio | LC50—96 h (ppm) | Synergistic effect of pesticides in mixtures observed | [94] | |
| LCh | 0.0031–0.38 | ||||
| BUT | 0.45–1.93 | ||||
| ATZ | 6.09–34.19 | ||||
| Imidacloprid | Danio rerio Oryzias latipes | 0.2 to 2000 ppb D. rerio 5 dpf O. latipes 14 dpf | Imidacloprid exposure indicated sub-lethal effects in both species; with prominent impacts in medaka, e.g., lesions, reduced growth being and deformities | [95] | |
| Imidacloprid | Danio rerio Oryzias latipes | 0, 0.2, 2, 20, 200 and 2000 ppb D. rerio 5 dpf O. latipes 13 dpf | Remarkable difference observed in organism morphology: with major deformities in medaka, however, almost none observed in zebrafish | [96] | |
| Imidacloprid, Nitenpyram | Gobiocypris rarus (brains) | 0.1, 0.5 and 2.0 ppm 60 d | Nitenpyram and imidacloprid altered the antioxidant genes expression levels and induced oxidative stress in brains of juvenile Chinese rare minnow | [98] | |
| Thiamethoxam, Tebuconazole | Gobiocypris rarus | LC50—96 h (ppm) | Synergistic action and additive toxicity were observed | [99] | |
| TEB | 4.07 | ||||
| TMC | 351.9 | ||||
| Nitenpyram, Imidacloprid, Dinotefuran | Gobiocypris rarus | 0.1, 0.5, or 2.0 ppm 60 d | DNA damage and Oxidative stress was depicted after nitenpyram and imidacloprid exposure which cause adverse effects on juvenile G. rarus liver in dose-dependent manner | [109] | |
| Nitenpyram, Imidacloprid, Dinotefuran | Gobiocypris rarus | 0.1, 0.5, or 2.0 ppm 60 days | Imidacloprid can induce genotoxicity. Chronic dinotefuran and imidacloprid might significantly reduce the immune system of juvenile G. rarus | [97] | |
| Imidacloprid, Thiamethoxam | Perca flavescens larvae | Survival probability: 0, 8.33, and 23.32 ppt Protein in tissues: 8.33, and 23.32, 132.28 ppt AChE activity: 8.33, and 23.32, 132.28 ppt With and without UVR | Imidacloprid demonstrated rise in AChE activity and protein content. Imidacloprid and UVR both factors disturb signal transmission in the nervous system of fish larvae and reduction in lipid peroxidation | [100] | |
| Clothianidin | Oncorhynchus nerka | 0.15, 1.5, 15 and 150 ppb 4 months | Clothianidin at 0.15 ppb raised 17β-estradiol levels in O. nerka swim-up fry, testosterone levels were not affected Clothianidin (150 ppb) decreased Liver glucocorticoid gene expression to O. nerka swim-up fry | [101] | |
| Imidacloprid | Australoheros facetus | 100, 300, 2500 ppb 24 and 48 h | Imidacloprid was found in all the tested gut, gills, muscle and liver tissues | [102] | |
|
Acetamiprid, Lambda-cyhalothrin, ACT-LCh, ACER 35 EC® | Clarias gariepinus | LC50 96 h (ppm) | Additive and antagonistic results were observed with marked nervous system damage | [103] | |
| LCh | 0.00083 | ||||
| ACT | 265.7 | ||||
| ACT-LCh | 0.043 | ||||
| ACER 35 EC® | 0.21 | ||||
| Imidacloprid | Prochilodus lineatus | 1.25, 12.5, 125, and 1250 ppb 120 h | After IMI exposure liver and kidney were observed to be most affected organs, followed by the gills | [110] | |
| Thiacloprid | Cyprinus carpio | 4.5, 45, 225, and 450 ppb for 35 days | Thiacloprid caused reduced growth and delay in ontogenetic development of carp | [111] | |
| Frog | |||||
| Clothianidin, Thiamethoxam | Lithobates sylvaticus, Lithobates pipiens | 2.5 and 250 ppb 2 weeks | Neonicotinoid exposure did not show variation among controls and exposed groups for any of the parameters observed for either leopard or wood frogs | [104] | |
| Imidacloprid, Thiamethoxam | Lithobates sylvaticus | 1, 10, and 100 ppb 6 weeks | Frog’s ability was affected to respond to predators, significantly augmenting their vulnerability to predation | [107] | |
| Clothianidin | Rana pipiens | 0, 0.23, 1, 10 and 100 ppb 8 weeks | Increase in stress response such as oxidative stress and change in leukocyte profile were observed | [106] | |
| Acetamiprid, Clothianidin, Dinotefuran, Imidacloprid | Silurana tropicalis | 96 h semi-static test 0.1 and 1.0 ppm | Amphibians were not directly affected by insecticides alone through larval stages at concentrations that are probably present in paddy water | [108] | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Malhotra, N.; Chen, K.H.-C.; Huang, J.-C.; Lai, H.-T.; Uapipatanakul, B.; Roldan, M.J.M.; Macabeo, A.P.G.; Ger, T.-R.; Hsiao, C.-D. Physiological Effects of Neonicotinoid Insecticides on Non-Target Aquatic Animals—An Updated Review. Int. J. Mol. Sci. 2021, 22, 9591. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22179591
Malhotra N, Chen KH-C, Huang J-C, Lai H-T, Uapipatanakul B, Roldan MJM, Macabeo APG, Ger T-R, Hsiao C-D. Physiological Effects of Neonicotinoid Insecticides on Non-Target Aquatic Animals—An Updated Review. International Journal of Molecular Sciences. 2021; 22(17):9591. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22179591
Chicago/Turabian StyleMalhotra, Nemi, Kelvin H.-C. Chen, Jong-Chin Huang, Hong-Thih Lai, Boontida Uapipatanakul, Marri Jmelou M. Roldan, Allan Patrick G. Macabeo, Tzong-Rong Ger, and Chung-Der Hsiao. 2021. "Physiological Effects of Neonicotinoid Insecticides on Non-Target Aquatic Animals—An Updated Review" International Journal of Molecular Sciences 22, no. 17: 9591. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22179591