
Deletion of Cdk5 in Macrophages Ameliorates Anti-Inflammatory Response during Endotoxemia through Induction of C-Maf and Il-10


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
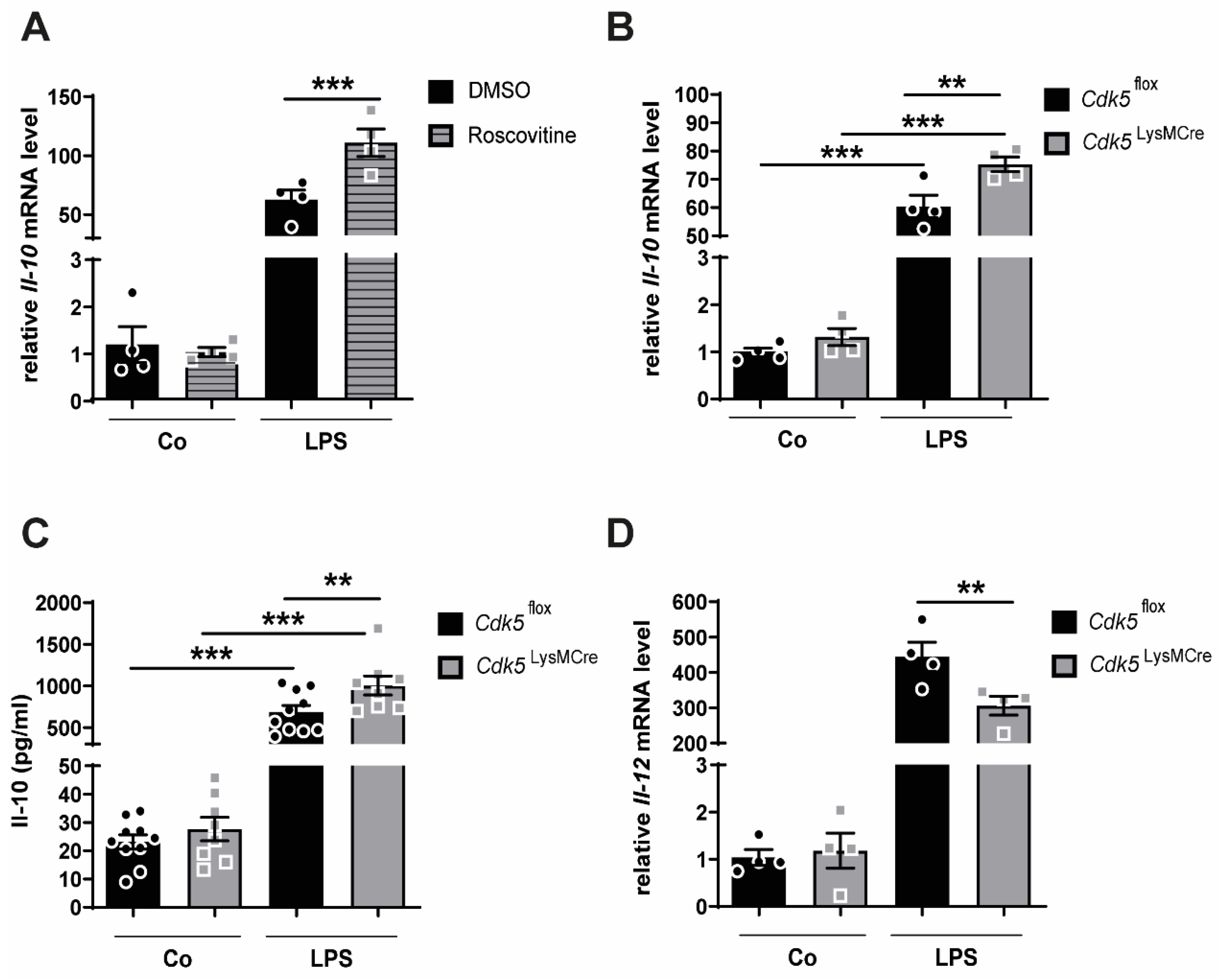
2.1. Inhibition of Cdk5 with Roscovitine or Cdk5 Genetic Deletion in Macrophages Enhances Il-10 Expression through C-Maf after Inflammatory Stimuli
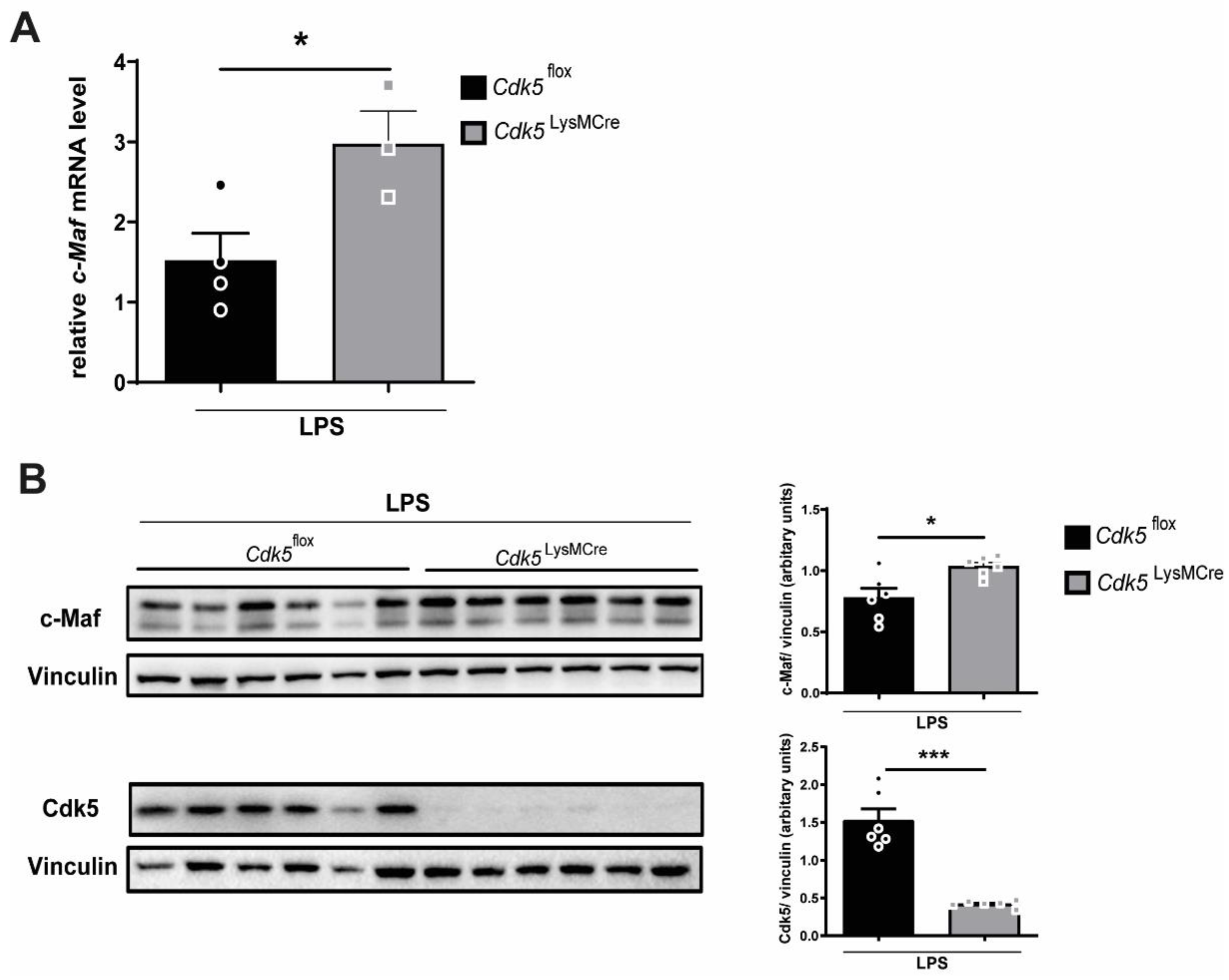
2.2. Genetic Deletion of Cdk5 in Macrophages Enhances Il-10 Levels through C-Maf in Mouse Lungs during Endotoxemia
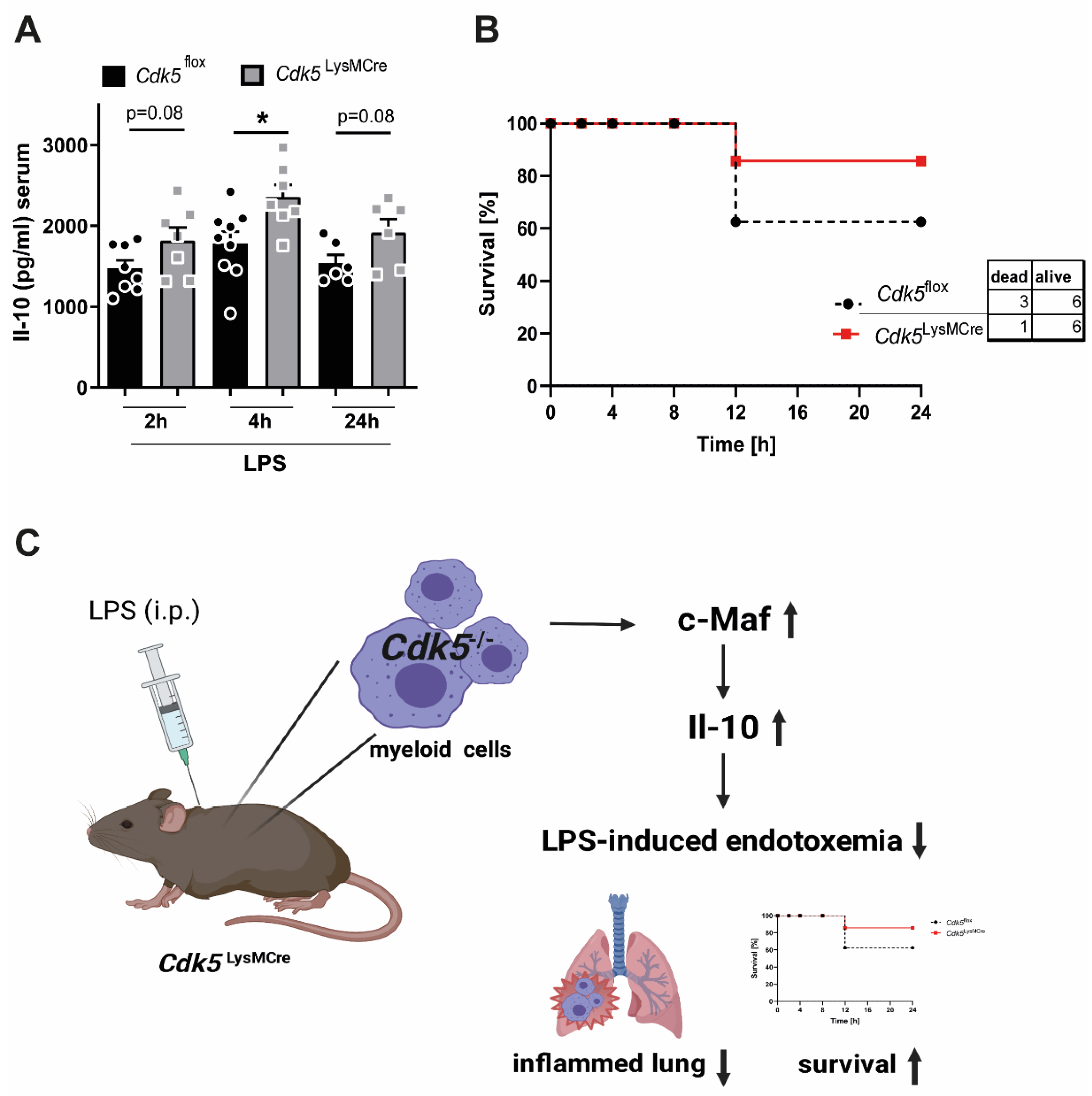
2.3. Cdk5LysMCre Mice Show an Ameliorated Disease Severity during Endotoxemia
3. Discussion
4. Materials and Methods
4.1. Mice
4.2. In Vivo Mouse Model
4.3. Measurements of Cytokine and Chemokine Concentrations
4.4. Blood Cell Anaylsis
4.5. Flow Cytometry
4.6. Cell Culture
4.7. Rna Isolation and Quantitative Rt-Pcr
4.8. Interleukin-10 ELISA
4.9. Immunoblot Analysis
4.10. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Moore, K.W.; Malefyt, R.d.W.; Coffman, R.L.; O’Garra, A. Interleukin-10 and the interleukin-10 receptor. Annu. Rev. Immunol. 2001, 19, 683–765. [Google Scholar] [CrossRef] [PubMed]
- Mosser, D.M.; Edwards, J.P. Exploring the full spectrum of macrophage activation. Nat. Rev. Immunol. 2008, 8, 958–969. [Google Scholar] [CrossRef]
- Kühn, R.; Löhler, J.; Rennick, D.; Rajewsky, K.; Müller, W. Interleukin-10-deficient mice develop chronic enterocolitis. Cell 1993, 75, 263–274. [Google Scholar] [CrossRef]
- Berg, D.J.; Kuhn, R.; Rajewsky, K.; Muller, W.; Menon, S.; Davidson, N.; Grunig, G.; Rennick, D. Interleukin-10 is a Central Regulator of the Response to LPS in Murine Models of Endotoxic Shock and the Shwartzman Reaction but not Endotoxin Tolerance. J. Clin. Investig. 1995, 96, 2339–2347. [Google Scholar] [CrossRef] [Green Version]
- Orraine, L.; Are, B.W.; Atthay, A.M. The acute respiratory distress syndrome. N. Engl. J. Med. 2000, 342, 1334–1349. [Google Scholar]
- Bernard, G.R.; Artigas, A.; Brigham, K.L.; Carlet, J.; Falke, K.; Hudson, L.; Lamy, M.; Legall, J.R.; Morris, A.; Spragg, R. Intensive Care Medicine Report of the American-European consensus conference on ARDS: Definitions, mechanisms, relevant outcomes and clinical trial coordination. Am. J. Respir. Crit. Care Med. 1994, 149, 818–824. [Google Scholar] [CrossRef] [PubMed]
- Herridge, M.S.; Cheung, A.M.; Tansey, C.M.; Matte-Martyn, A.; Diaz-Granados, N.; Al-Saidi, F.; Cooper, A.B.; Guest, C.B.; David Mazer, C.; Mehta, S.; et al. One-Year Outcomes in Survivors of the Acute Respiratory Distress Syndrome. N. Engl. J. Med. 2003, 348, 683–693. [Google Scholar] [CrossRef] [Green Version]
- Inoue, G. Effect of interleukin-10 (IL-10) on experimental LPS-induced acute lung injury. J. Infect. Chemother. 2000, 6, 51–60. [Google Scholar] [CrossRef]
- Cao, S.; Liu, J.; Chesi, M.; Bergsagel, P.L.; Ho, I.-C.; Donnelly, R.P.; Ma, X. Differential Regulation of IL-12 and IL-10 Gene Expression in Macrophages by the Basic Leucine Zipper Transcription Factor c-Maf Fibrosarcoma. J. Immunol. 2002, 169, 5715–5725. [Google Scholar] [CrossRef] [Green Version]
- Cao, S.; Liu, J.; Song, L.; Ma, X. The Protooncogene c-Maf Is an Essential Transcription Factor for IL-10 Gene Expression in Macrophages. J. Immunol. 2005, 174, 3484–3492. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.; Tong, Z.; Ding, C.; Luo, F.; Wu, S.; Wu, C.; Albeituni, S.; He, L.; Hu, X.; Tieri, D.; et al. Transcription factor c-Maf is a checkpoint that programs macrophages in lung cancer. J. Clin. Investig. 2020, 130, 2081–2096. [Google Scholar] [CrossRef] [Green Version]
- Nishizawa, M.; Kataoka, K.; Gotot, N.; Fujiwara, K.T.; Kawai, S. v-maf, a viral oncogene that encodes a “leucine zipper” motif (avian retrovirus/transformation/DNA binding protein). Proc. Natl. Acad. Sci. USA 1989, 86, 7711–7715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding Wu, Y.; Levy, D.E.; Ochando, J.C.; Jiangnan Xu, J.S.; Yang, Y.; Qiu, G.; Lal, G. c-Maf Regulates IL-10 Expression during Th17 Polarization. J. Immunol. Ref. 2009, 182, 6226–6236. [Google Scholar] [CrossRef] [Green Version]
- Apetoh, L.; Quintana, F.J.; Pot, C.; Joller, N.; Xiao, S.; Kumar, D.; Burns, E.J.; Sherr, D.H.; Weiner, H.L.; Kuchroo, V.K. The aryl hydrocarbon receptor interacts with c-Maf to promote the differentiation of type 1 regulatory T cells induced by IL-27. Nat. Immunol. 2010, 11, 854–861. [Google Scholar] [CrossRef] [Green Version]
- Ma, X.; Yan, W.; Zheng, H.; Du, Q.; Zhang, L.; Ban, Y.; Li, N.; Wei, F. Regulation of IL-10 and IL-12 production and function in macrophages and dendritic cells. F1000Research 2015, 4, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Na, Y.R.; Jung, D.; Gu, G.J.; Jang, A.R.; Suh, Y.H.; Seok, S.H. The early synthesis of p35 and activation of CDK5 in LPS-stimulated macrophages suppresses interleukin-10 production. Sci. Signal. 2015, 8, ra121. [Google Scholar] [CrossRef] [PubMed]
- Pareek, T.K.; Keller, J.; Kesavapany, S.; Pant, H.C.; Iadarola, M.J.; Brady, R.O.; Kulkarni, A.B. Cyclin-dependent kinase 5 activity regulates pain signaling. Proc. Natl. Acad. Sci. USA 2005, 103, 791–796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosales, J.L.; Ernst, J.D.; Hallows, J.; Lee, K.Y. GTP-dependent secretion from neutrophils is regulated by Cdk5. J. Biol. Chem. 2004, 279, 53932–53936. [Google Scholar] [CrossRef] [Green Version]
- Pareek, T.K.; Lam, E.; Zheng, X.; Askew, D.; Kulkarni, A.B.; Chance, M.R.; Huang, A.Y.; Cooke, K.R.; Letterio, J.J. Cyclin-dependent kinase 5 activity is required for T cell activation and induction of experimental autoimmune encephalomyelitis. J. Exp. Med. 2010, 207, 2507–2519. [Google Scholar] [CrossRef]
- Pfänder, P.; Fidan, M.; Burret, U.; Lipinski, L.; Vettorazzi, S. Cdk5 deletion enhances the anti-inflammatory potential of GC-mediated GR activation during inflammation. Front. Immunol. 2019, 10, 1554. [Google Scholar] [CrossRef] [PubMed]
- Aste-Amezaga, M.; Ma, X.; Sartori, A.; Trinchieri, G. Molecular Mechanisms of the Induction of IL-12 and Its Inhibition by IL-10. J. Immunol. 1998, 160, 5936–5944. [Google Scholar] [PubMed]
- Matthay, M.A.; Song, Y.; Bai, C.; Jones, K.D.; Edu, M.M. The acute respiratory distress syndrome in 2013. Transl. Respir. Med. 2013, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rossi, A.G.; Sawatzky, D.A.; Walker, A.; Ward, C.; Sheldrake, T.A.; Riley, N.A.; Caldicott, A.; Martinez-Losa, M.; Walker, T.R.; Duffin, R.; et al. Cyclin-dependent kinase inhibitors enhance the resolution of inflammation by promoting inflammatory cell apoptosis. Nat. Med. 2006, 12, 1056–1064. [Google Scholar] [CrossRef]
- Hoogendijk, A.J.; Roelofs, J.J.T.H.; Duitman, J.; van Lieshout, M.H.P.; Blok, D.C.; van der Poll, T.; Wieland, C.W. R-roscovitine reduces lung inflammation induced by lipoteichoic acid and Streptococcus pneumoniae. Mol. Med. 2012, 18, 1086–1095. [Google Scholar] [CrossRef] [Green Version]
- Rocques, N.; Abou Zeid, N.; Sii-Felice, K.; Lecoin, L.; Felder-Schmittbuhl, M.P.; Eychène, A.; Pouponnot, C. GSK-3-Mediated Phosphorylation Enhances Maf-Transforming Activity. Mol. Cell 2007, 28, 584–597. [Google Scholar] [CrossRef]
- Paglini, G.; Pigino, G.; Kunda, P.; Morfini, G.; Maccioni, R.; Quiroga, S.; Ferreira, A.; Cáceres, A. Evidence for the participation of the neuron-specific CDK5 activator P35 during laminin-enhanced axonal growth. J. Neurosci. 1998, 18, 9858–9869. [Google Scholar] [CrossRef] [PubMed]
- Morfini, G.; Szebenyi, G.; Brown, H.; Pant, H.C.; Pigino, G.; DeBoer, S.; Beffert, U.; Brady, S.T. A novel CDK5-dependent pathway for regulating GSK3 activity and kinesin-driven motility in neurons. EMBO J. 2004, 23, 2235–2245. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Kuchroo, V. Epigenetic and transcriptional mechanisms for the regulation of IL-10. Semin. Immunol. 2019, 44, 101324. [Google Scholar] [CrossRef]
- Duffin, R.; Leitch, A.E.; Sheldrake, T.A.; Hallett, J.M.; Meyer, C.; Fox, S.; Alessandri, A.L.; Martin, M.C.; Brady, H.J.; Teixeira, M.M.; et al. The CDK inhibitor, R-roscovitine, promotes eosinophil apoptosis by down-regulation of Mcl-1. FEBS Lett. 2009, 583, 2540–2546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liebl, J.; Zhang, S.; Moser, M.; Agalarov, Y.; Demir, C.S.; Hager, B.; Bibb, J.A.; Adams, R.H.; Kiefer, F.; Miura, N.; et al. Cdk5 controls lymphatic vessel development and function by phosphorylation of Foxc2. Nat. Commun. 2015, 6, 7274. [Google Scholar] [CrossRef] [Green Version]
- Vettorazzi, S.; Bode, C.; Dejager, L.; Frappart, L.; Shelest, E.; Klaen, C.; Tasdogan, A.; Reichardt, H.M.; Libert, C.; Schneider, M.; et al. ARTICLE Glucocorticoids limit acute lung inflammation in concert with inflammatory stimuli by induction of SphK1. Nat. Commun. 2015, 6, 7796. [Google Scholar] [CrossRef] [PubMed] [Green Version]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pfänder, P.; Eiers, A.-K.; Burret, U.; Vettorazzi, S. Deletion of Cdk5 in Macrophages Ameliorates Anti-Inflammatory Response during Endotoxemia through Induction of C-Maf and Il-10. Int. J. Mol. Sci. 2021, 22, 9648. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22179648
Pfänder P, Eiers A-K, Burret U, Vettorazzi S. Deletion of Cdk5 in Macrophages Ameliorates Anti-Inflammatory Response during Endotoxemia through Induction of C-Maf and Il-10. International Journal of Molecular Sciences. 2021; 22(17):9648. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22179648
Chicago/Turabian StylePfänder, Pauline, Ann-Kathrin Eiers, Ute Burret, and Sabine Vettorazzi. 2021. "Deletion of Cdk5 in Macrophages Ameliorates Anti-Inflammatory Response during Endotoxemia through Induction of C-Maf and Il-10" International Journal of Molecular Sciences 22, no. 17: 9648. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22179648