
In Search of Monocot Phosphodiesterases: Identification of a Calmodulin Stimulated Phosphodiesterase from Brachypodium distachyon

 , , , and
, , , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
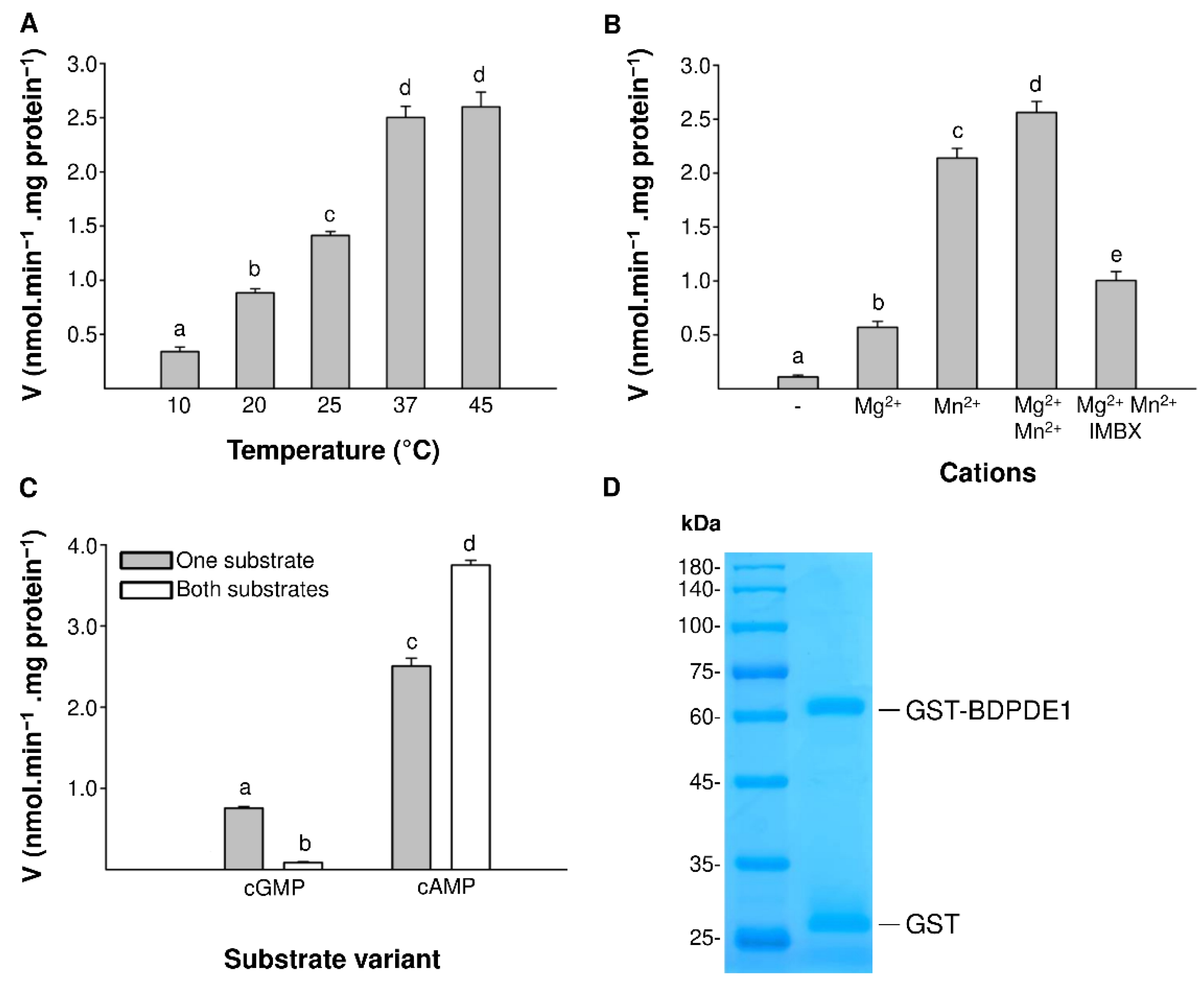
2.1. Identification and Characterization of PDE Activity in BDPDE1
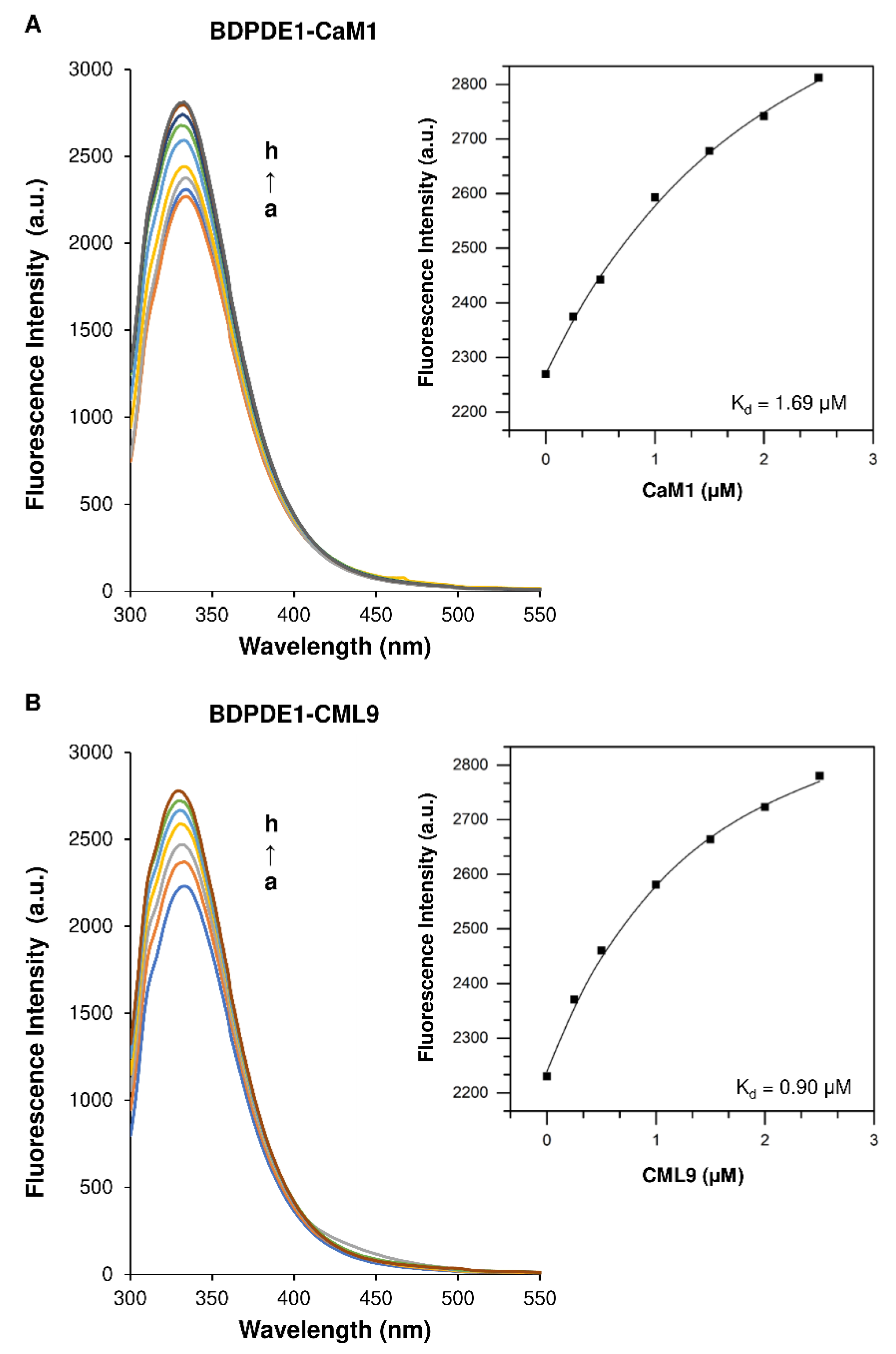
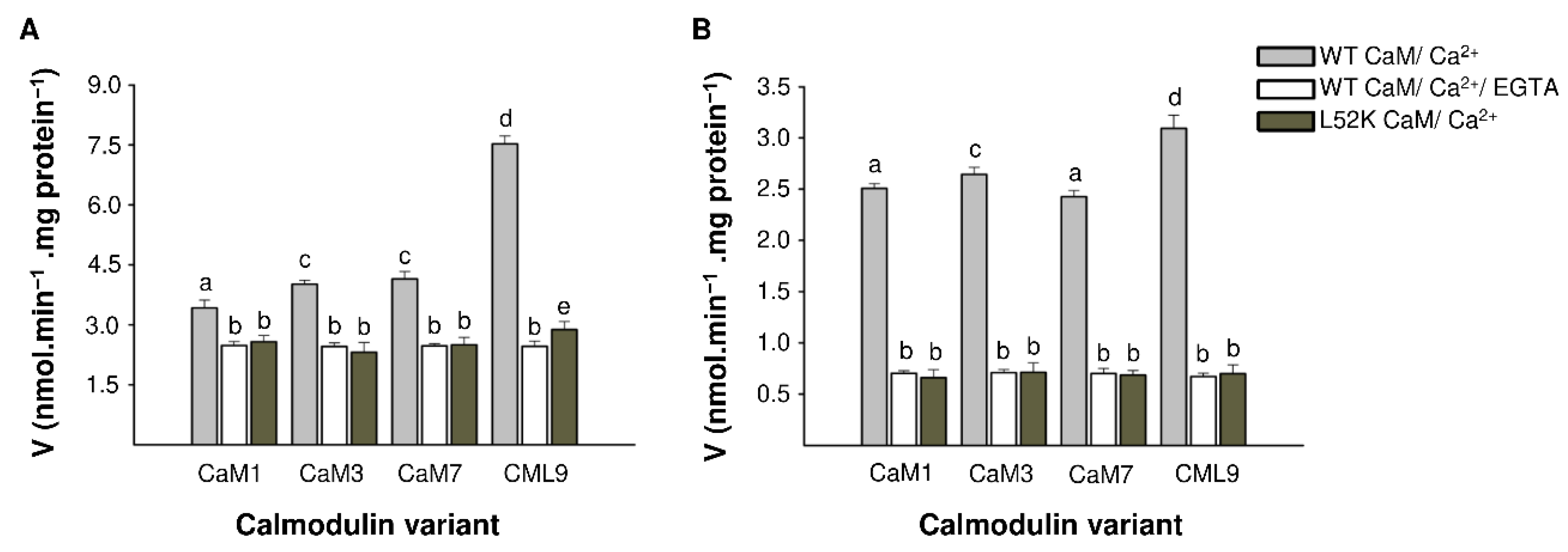
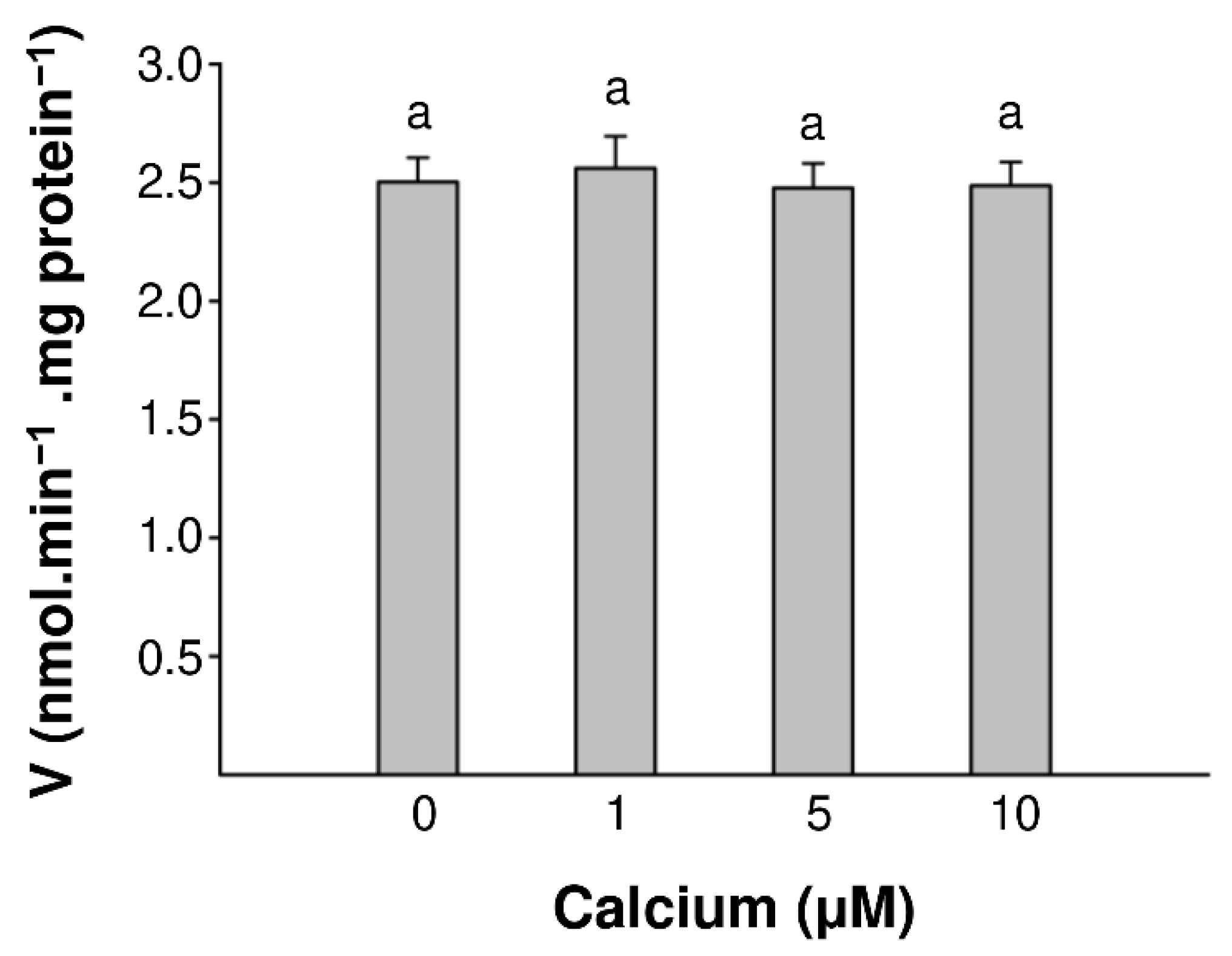
2.2. The Calmodulin/Ca2+ Complex Stimulates BDPDE1 Activity
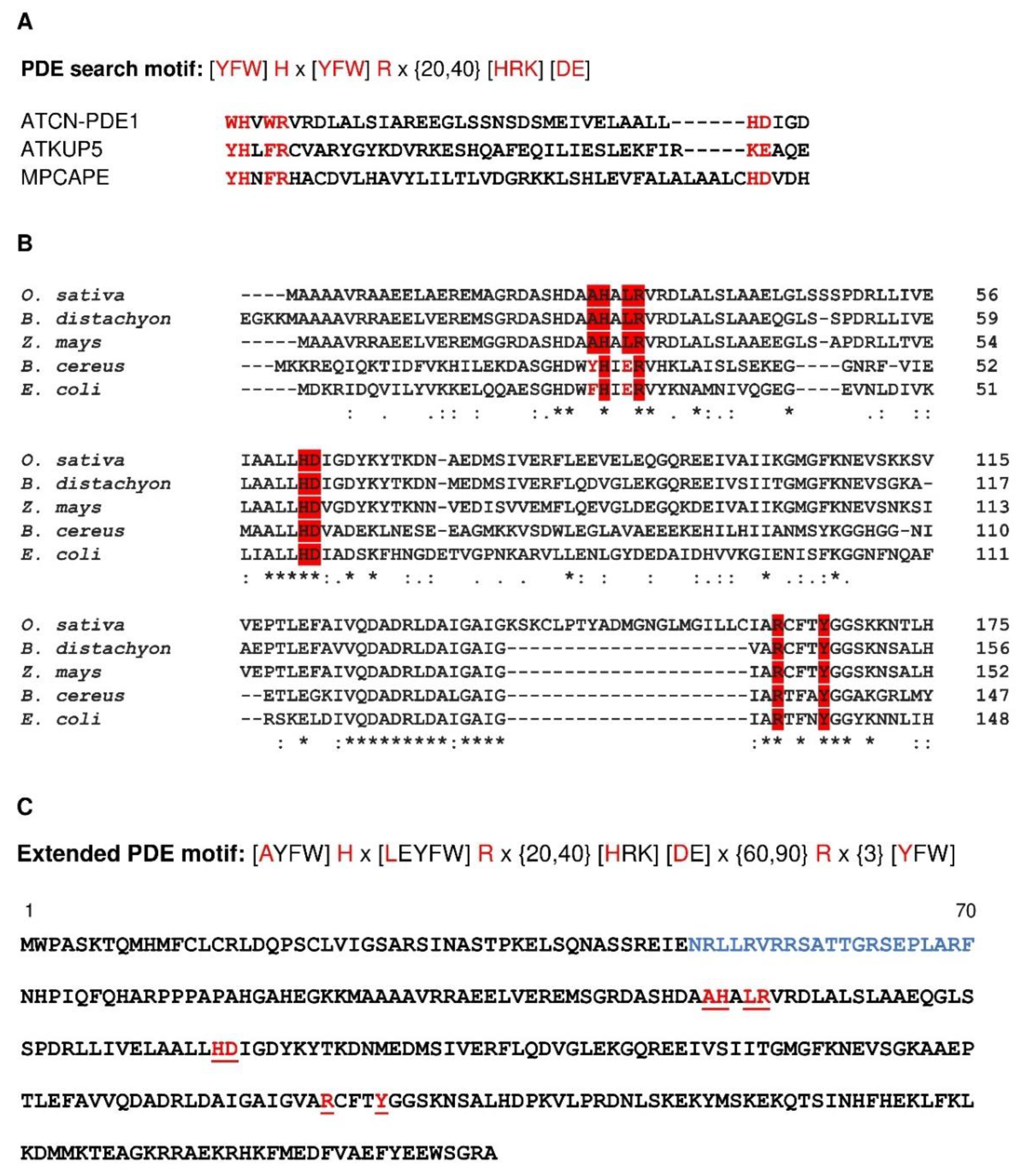
2.3. Probing the Catalytic Center of BDPDE1 and Construction of an Expanded PDE Search Motif
3. Materials and Methods
3.1. Expression Vector Construct
- -BDPDE1 (forward)
- 5′-GGATCCCCAGGAATTCCCATGTGGCCAGCATCCAAAACAC-3′
- -BDPDE1 (reverse)
- 5′-GATGCGGCCGCTCGAGAATCAAGCCCTGCCACTCCAC-3′
3.2. Site Directed Mutagenesis of BDPDE1
3.3. Expression and Purification of the Recombinant Protein
3.4. Structural Analysis of the PDE Center and CaM-Binding Site in BDPDE1
3.5. PDE Biochemical Assay and LC-MS/MS Analysis
3.6. Fluorescence Studies
3.7. Statistical Analyses
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Marondedze, C.; Wong, A.; Thomas, L.; Irving, H.; Gehring, C. Cyclic nucleotide monophosphates in plants and plant signaling. Handb. Exp. Pharmacol. 2017, 238, 87–103. [Google Scholar] [CrossRef]
- Gehring, C.; Turek, I.S. Cyclic nucleotide monophosphates and their cyclases in plant signaling. Front. Plant. Sci. 2017, 8, 1704. [Google Scholar] [CrossRef] [Green Version]
- Gehring, C. Adenyl cyclases and CAMP in plant signaling—Past and present. Cell Commun. Signal. 2010, 8, 15. [Google Scholar] [CrossRef] [Green Version]
- Lemtiri-Chlieh, F.; Thomas, L.; Marondedze, C.; Irving, H.; Gehring, C. Cyclic nucleotides and nucleotide cyclases in plant stress responses. In Abiotic Stress Response in Plants—Physiological, Biochemical and Genetic Perspectives; Shanker, A., Venkateswarlu, B., Eds.; IntechOpen Limited: London, UK, 2011; pp. 137–182. [Google Scholar] [CrossRef] [Green Version]
- Maathuis, F.J.M. CGMP modulates gene transcription and cation transport in Arabidopsis Roots. Plant. J. 2006, 45, 700–711. [Google Scholar] [CrossRef]
- Barnes, S.A.; Quaggio, R.B.; Chua, N.H. Phytochrome signal-transduction: Characterization of pathways and isolation of mutants. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1995, 350, 67–74. [Google Scholar] [CrossRef]
- Maathuis, F.J.M.; Sanders, D. Sodium uptake in Arabidopsis roots is regulated by cyclic nucleotides. Plant. Physiol. 2001, 127, 1617–1625. [Google Scholar] [CrossRef]
- Thomas, L.; Marondedze, C.; Ederli, L.; Pasqualini, S.; Gehring, C. Proteomic signatures implicate CAMP in light and temperature responses in Arabidopsis thaliana. J. Proteom. 2013, 83, 47–59. [Google Scholar] [CrossRef]
- Gross, I.; Durner, J. In search of enzymes with a role in 3′, 5′-Cyclic guanosine monophosphate metabolism in plants. Front. Plant. Sci. 2016, 7, 576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moutinho, A.; Hussey, P.J.; Trewavas, A.J.; Malhó, R. CAMP acts as a second messenger in pollen tube growth and reorientation. Proc. Natl. Acad. Sci. USA 2001, 98, 10481–10486. [Google Scholar] [CrossRef] [Green Version]
- Ruzvidzo, O.; Gehring, C.; Wong, A. New perspectives on plant Adenylyl Cyclases. Front. Plant. Sci. 2019, 6, 136. [Google Scholar] [CrossRef] [Green Version]
- Kwezi, L.; Ruzvidzo, O.; Wheeler, J.I.; Govender, K.; Iacuone, S.; Thompson, P.E.; Gehring, C.; Irving, H.R. The Phytosulfokine (PSK) receptor is capable of guanylate cyclase activity and enabling cyclic GMP-dependent signaling in plants. J. Biol. Chem. 2011, 286, 22580–22588. [Google Scholar] [CrossRef] [Green Version]
- Duszyn, M.; Świeżawska-Boniecka, B.; Wong, A.; Jaworski, K.; Szmidt-Jaworska, A. In vitro characterization of Guanylyl Cyclase BdPepR2 from Brachypodium distachyon identified through a motif-based approach. Int. J. Mol. Sci. 2021, 22, 6243. [Google Scholar] [CrossRef]
- Al-Younis, I.; Wong, A.; Gehring, C. The Arabidopsis thaliana K+-Uptake Permease 7 (AtKUP7) contains a functional cytosolic adenylate cyclase catalytic center. FEBS Lett. 2015, 589, 3848–3852. [Google Scholar] [CrossRef] [Green Version]
- Świeżawska, B.; Duszyn, M.; Kwiatkowski, M.; Jaworski, K.; Pawełek, A.; Szmidt-Jaworska, A. Brachypodium distachyon Triphosphate tunnel metalloenzyme 3 is both a triphosphatase and an adenylyl cyclase upregulated by mechanical wounding. FEBS Lett. 2019, 594, 1101–1111. [Google Scholar] [CrossRef]
- Isner, J.-C.; Olteanu, V.-A.; Hetherington, A.J.; Coupel-Ledru, A.; Sun, P.; Pridgeon, A.J.; Jones, G.S.; Oates, M.; Williams, T.A.; Maathuis, F.J.M.; et al. Short- and long-term effects of UVA on Arabidopsis are mediated by a novel CGMP phosphodiesterase. Curr. Biol. 2019, 29, 2580–2585.e4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwiatkowski, M.; Wong, A.; Kozakiewicz, A.; Gehring, C.; Jaworski, K. A tandem motif-based and structural approach can identify hidden functional phosphodiesterases. Comput. Struct. Biotechnol. J. 2021, 19, 970–975. [Google Scholar] [CrossRef] [PubMed]
- Kasahara, M.; Suetsugu, N.; Urano, Y.; Yamamoto, C.; Ohmori, M.; Takada, Y.; Okuda, S.; Nishiyama, T.; Sakayama, H.; Kohchi, T.; et al. An adenylyl cyclase with a phosphodiesterase domain in basal plants with a motile sperm system. Sci. Rep. 2016, 6, 39232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Świeżawska-Boniecka, B.; Duszyn, M.; Kwiatkowski, M.; Szmidt-Jaworska, A.; Jaworski, K. Cross talk between cyclic nucleotides and calcium signaling pathways in plants–achievements and prospects. Front. Plant. Sci. 2021, 12, 643560. [Google Scholar] [CrossRef] [PubMed]
- Muleya, V.; Wheeler, J.I.; Ruzvidzo, O.; Freihat, L.; Manallack, D.T.; Gehring, C.; Irving, H.R. Calcium is the switch in the moonlighting dual function of the ligand-activated receptor kinase phytosulfokine receptor 1. Cell Commun. Signal. 2014, 12, 60. [Google Scholar] [CrossRef]
- Halls, M.L.; Cooper, D.M.F. Regulation by Ca2+-signaling pathways of adenylyl cyclases. Cold Spring Harb. Perspect. Biol. 2011, 3, a004143. [Google Scholar] [CrossRef]
- Yu, J. Identification and characterisation of a human calmodulin-stimulated phosphodiesterase PDE1B1. Cell Signal. 1997, 9, 519–529. [Google Scholar] [CrossRef]
- Yakunin, A.F.; Proudfoot, M.; Kuznetsova, E.; Savchenko, A.; Brown, G.; Arrowsmith, C.H.; Edwards, A.M. The HD domain of the Escherichia coli TRNA nucleotidyltransferase has 2′,3′-cyclic phosphodiesterase, 2′-nucleotidase, and phosphatase activities. J. Biol. Chem. 2004, 279, 36819–36827. [Google Scholar] [CrossRef] [Green Version]
- Jeon, Y.J.; Park, S.C.; Song, W.S.; Kim, O.-H.; Oh, B.-C.; Yoon, S. Structural and biochemical characterization of bacterial YpgQ protein reveals a metal-dependent nucleotide pyrophosphohydrolase. J. Struct. Biol. 2016, 195, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Zimmerman, M.D.; Proudfoot, M.; Yakunin, A.; Minor, W. Structural insight into the mechanism of substrate specificity and catalytic activity of an HD-domain phosphohydrolase: The 5′-deoxyribonucleotidase YfbR from Escherichia coli. J. Mol. Biol. 2008, 378, 215–226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wada, H.; Osborne, J.C.; Manganiello, V.C. Effects of temperature on allosteric and catalytic properties of the CGMPStimulated cyclic nucleotide phosphodiesterase from calf liver. J. Biol. Chem. 1987, 262, 5139–5144. [Google Scholar] [CrossRef]
- Xu, R.X. Atomic structure of PDE4: Insights into phosphodiesterase mechanism and specificity. Science 2000, 288, 1822–1825. [Google Scholar] [CrossRef] [PubMed]
- Conti, M.; Beavo, J. Biochemistry and physiology of cyclic nucleotide phosphodiesterases: Essential components in cyclic nucleotide signaling. Ann. Rev. Biochem. 2007, 76, 481–511. [Google Scholar] [CrossRef]
- Beavo, J.A.; Hardman, J.G.; Sutherland, E.W. Stimulation of adenosine 3′,5′-monophosphate hydrolysis by guanosine 3′,5′-monophosphate. J. Biol. Chem. 1971, 246, 3841–3846. [Google Scholar] [CrossRef]
- Lee, D.I.; Kass, D.A. Phosphodiesterases and cyclic GMP regulation in heart muscle. Physiology 2012, 27, 248–258. [Google Scholar] [CrossRef] [Green Version]
- Snedden, W.A.; Fromm, H. Calmodulin, calmodulin-related proteins and plant responses to the environment. Trends Plant. Sci. 1998, 3, 299–304. [Google Scholar] [CrossRef]
- Charpenteau, M.; Jaworski, K.; Ramirez, B.C.; Tretyn, A.; Ranjeva, R.; Ranty, B. A receptor-like kinase from Arabidopsis thaliana is a calmodulin-binding protein. Biochem. J. 2004, 379, 841–848. [Google Scholar] [CrossRef]
- Jaworski, K.; Pawełek, A.; Kopcewicz, J.; Szmidt-Jaworska, A. The calcium-dependent protein kinase (PnCDPK1) is involved in Pharbitis nil flowering. J. Plant Physiol. 2012, 169, 1578–1585. [Google Scholar] [CrossRef]
- Jaworski, K.; Szmidt-Jaworska, A.; Kopcewicz, J. Two calcium dependent protein kinases are differently regulated by light and have different activity patterns during seedling growth in Pharbitis nil. Plant. Growth Regul. 2011, 65, 369–379. [Google Scholar] [CrossRef]
- Kakkar, R.; Raju, R.V.S.; Sharma, R.K. Calmodulin-dependent cyclic nucleotide phosphodiesterase (PDE1). Cell Mol. Life Sci. 1999, 55, 1164–1186. [Google Scholar] [CrossRef] [PubMed]
- Van de Weert, M.; Stella, L. Fluorescence quenching and ligand binding: A critical discussion of a popular methodology. J. Mol. Struct. 2011, 998, 144–150. [Google Scholar] [CrossRef]
- Calmodulin Target Database. Available online: http://calcium.uhnres.utoronto.ca/ctdb/ctdb/sequence.html (accessed on 9 June 2021).
- Harmon, A.C. Calcium-regulated protein kinases of plants. Gravit. Space Biol. Bull. 2003, 16, 83–90. [Google Scholar] [PubMed]
- Šali, A.; Blundell, T.L. Comparative protein modelling by satisfaction of spatial restraints. J. Mol. Biol. 1993, 234, 779–815. [Google Scholar] [CrossRef]
- Trott, O.; Olson, A.J. Software news and update AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF chimera—A visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [Green Version]
- Bers, D.M.; Patton, C.W.; Nuccitelli, R. A practical guide to the preparation of Ca(2+) buffers. Methods Cell Biol. 2010, 99, 1–26. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kwiatkowski, M.; Wong, A.; Kozakiewicz-Piekarz, A.; Gehring, C.; Jaworski, K. In Search of Monocot Phosphodiesterases: Identification of a Calmodulin Stimulated Phosphodiesterase from Brachypodium distachyon. Int. J. Mol. Sci. 2021, 22, 9654. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22179654
Kwiatkowski M, Wong A, Kozakiewicz-Piekarz A, Gehring C, Jaworski K. In Search of Monocot Phosphodiesterases: Identification of a Calmodulin Stimulated Phosphodiesterase from Brachypodium distachyon. International Journal of Molecular Sciences. 2021; 22(17):9654. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22179654
Chicago/Turabian StyleKwiatkowski, Mateusz, Aloysius Wong, Anna Kozakiewicz-Piekarz, Christoph Gehring, and Krzysztof Jaworski. 2021. "In Search of Monocot Phosphodiesterases: Identification of a Calmodulin Stimulated Phosphodiesterase from Brachypodium distachyon" International Journal of Molecular Sciences 22, no. 17: 9654. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22179654