
Impact of Stilbenes as Epigenetic Modulators of Breast Cancer Risk and Associated Biomarkers


Abstract
:1. Introduction
2. Scope of Primary Prevention of Breast Cancer Using Phytochemicals Including Stilbenes
3. Scope of Primary Prevention in Breast Cancer from a Detailed Epigenetic Perspective
3.1. Genetic and Epigenetic Biomarker Landscape: Breast Cancer Risk Factors and Susceptibility
3.1.1. DNA Methylation as Biomarkers
3.1.2. Histone Modifications as Biomarkers
3.1.3. Non-Coding RNA as Biomarkers

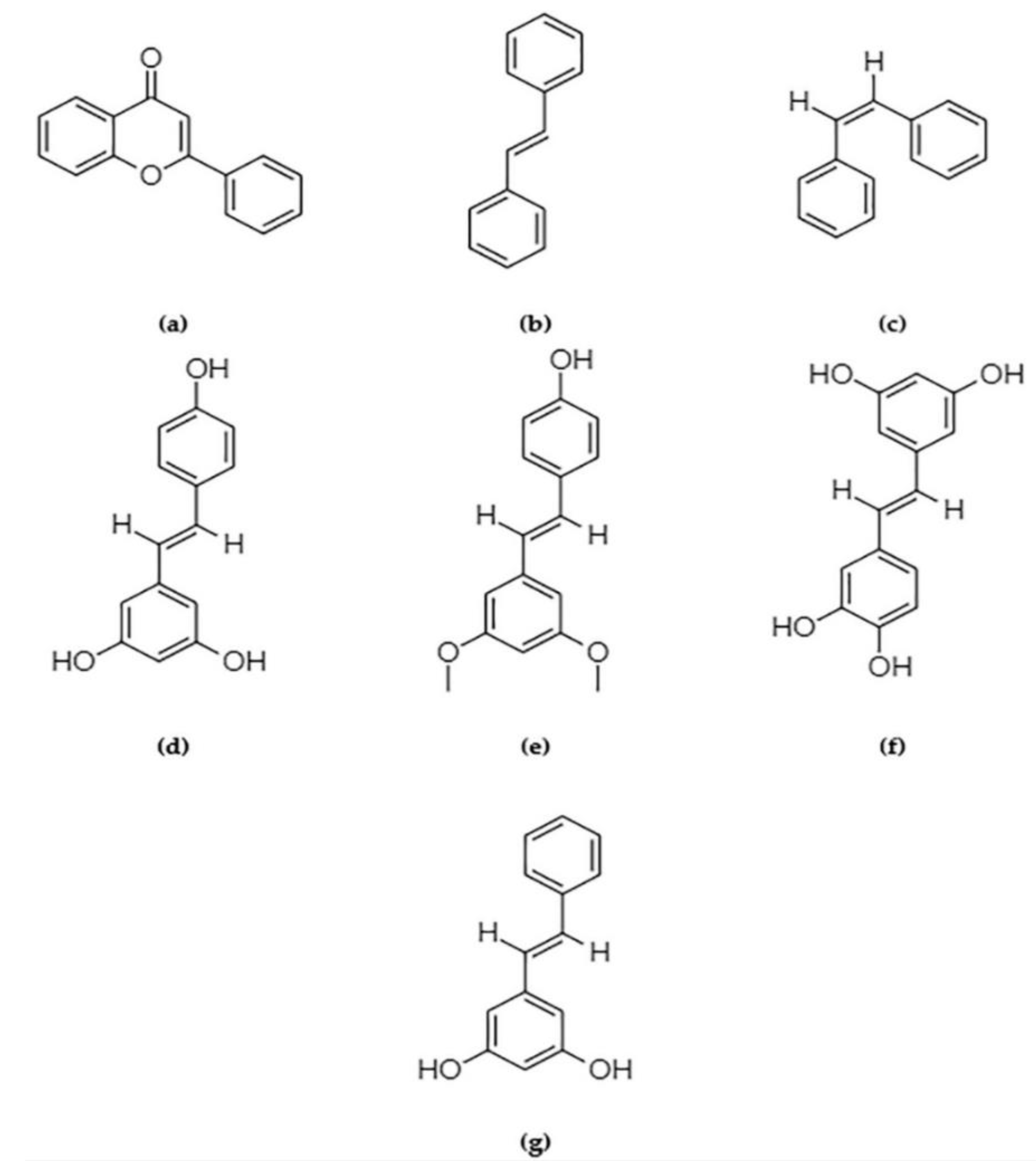{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ncRNA | Therapeutic Significance | Breast Cancer Subtype | Reference |
|---|---|---|---|
| miR-21-3p, miR-21-5p, and miR-99a-5p, miR-1246, miR-1307-3p, miR-4634, miR-6861-5p, and miR-6875-5p | Risk prediction and early detection and overall survival | Unspecified | [43] |
| Panel of 8 miRNAs (miR-139-5p, miR-10b-5p, miR-486-5p, miR-455-3p, miR-107, miR-146b-5p, miR-324-5p, and miR-20a-5p) | Predicting risk of relapse | Triple-negative breast cancer | [70] |
| Panel of 6 miRNAs (miR-3124-5p, miR-1184, miR-4423-3p, miR-4529-3p, miR-7855-5p, and miR-4446-3p) | Breast cancer risk prediction | Unspecified | [65] |
| miRNA-191, miRNA-484, miR-16, miR-25, miR-222, and miR-324-3p | Breast cancer risk predictor | Unspecified | [71] |
| Panel of 4 miRNAs (hsa-miR-21, hsa-miR-494, hsa-miR-494, and hsa-miR-183) | Metastatic risk prediction | Unspecified | [72] |
| Panel of 4 miRNAs (miR-1246, miR-206, miR-24, and miR-373) | Early diagnosis of breast cancer | Unspecified | [66] |
| lncRNAs like PVT1, MAPT-AS1, LINC00667, and LINC00938 | Predicting breast cancer survival | Unspecified | [73] |
| Panel of miR-127-3p, miR-148b, miR-376a, miR-376c, miR-409-3p, miR-652, and miR-801 | Distinguishing healthy women from women carrying benign or malignant breast tumors with more accuracy in younger individuals | Unspecified | [74] |
| miR-200a, miR-200b, miR-200c, miR-210, miR-215 and miR-486-5p | Metastasis onset predictor | Unspecified | [74] |
| mRNA-lncRNA conjugate (mRNA species for FCGR1A, RSAD2, CHRDL1, and the lncRNA species for HIF1A-AS2 and AK124454) | Predicting risk of relapse | Triple-negative breast cancer | [32] |
4. In-Practice Clinical Methods for Addressing Primary Prevention
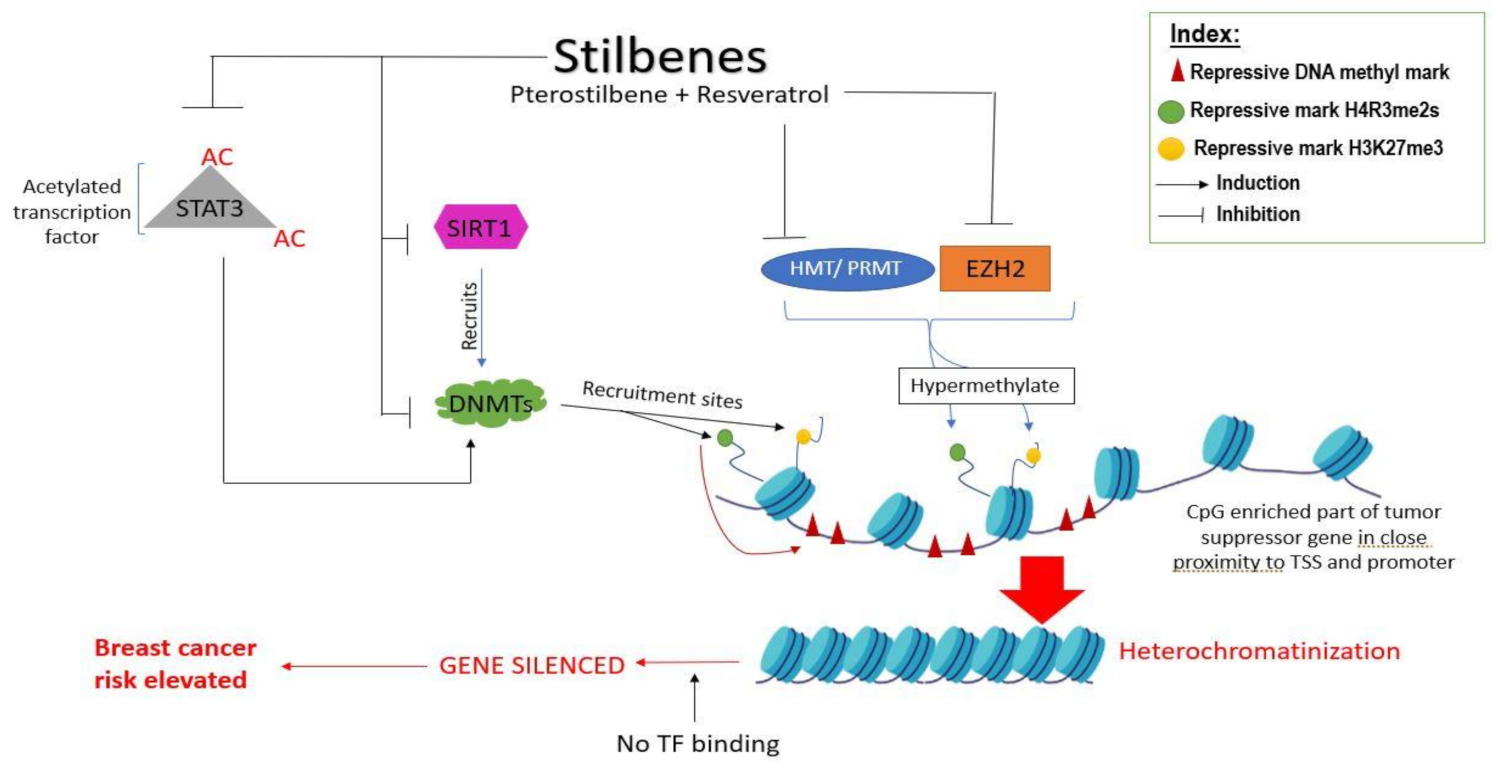
4.1. Role of Stilbenes in Differentially Modulating DNA Methylation of Genes and Gene Loci
4.2. Role of Stilbenes in Differentially Modifying Histones
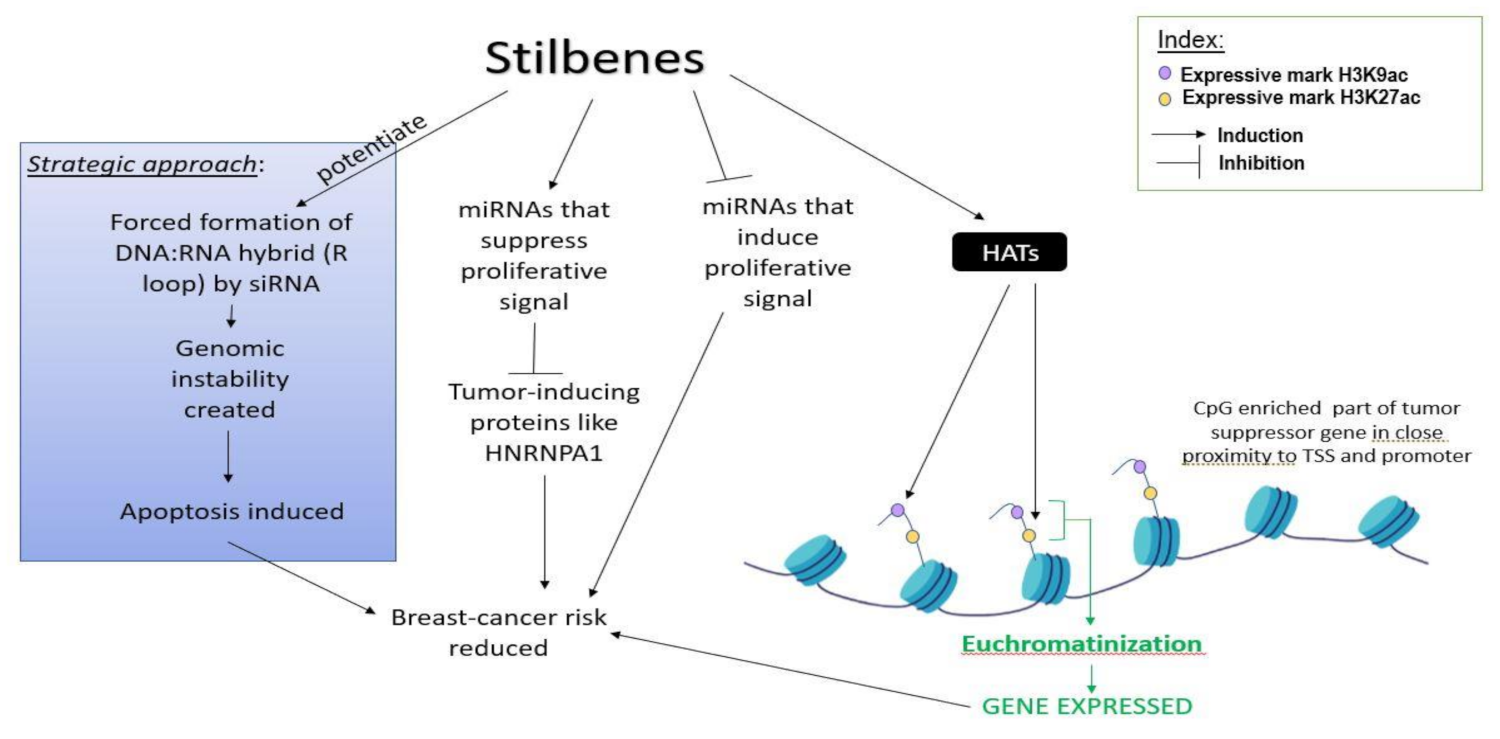
4.3. Role of Stilbenes in Differentially Modulating the Activity of Non-Coding RNAs and Preventing Breast Cancer Initiation
5. Epigenetic Effect of Stilbene on Genes and Proteins Related to Cell-Proliferation and Metastasis
6. Bioavailability of Stilbenes in Target Tissue and Limitations
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| Ais | Aromatase inhibitors |
| Akt | Protein kinase B |
| ARF | Adenosine 5’-diphosphate ribosylation factor |
| ATM | Serine/ Threonine kinase |
| Bax | Bcl-2-associated X protein |
| Bcl-2 | B-cell lymphoma 2 protein |
| BRCA1/2 | Breast cancer gene 1 and 2 |
| CD1 | Cluster of differentiation 1 |
| cIAP2 | Cellular Inhibitor Of Apoptosis 2 gene |
| c-Myc | Myelocytomatosis proto-oncogene |
| CNA-high | Copy number alterations high |
| CNA-quiet | Copy number alterations quiet |
| COX2 | Cyclooxygenase 2 gene |
| CpG | Regions of DNA with repetitive occurrence of cytosine and guanosine nucleotides |
| Cyth3 | Cytohesin-3 gene |
| DNA | Deoxyribonucleic acid |
| DNMT1 | DNA methyltransferase 1 |
| DNMT3A | DNA methyltransferase 3 Alpha |
| DNMT3B | DNA methyltransferase 3 Beta |
| DNMTs | DNA methyl transferases |
| EGCG | Epigallocatechin gallate |
| EMT | Epithelial mesenchymal transformation |
| ER- | Estrogen receptor negative |
| ER+ | Estrogen receptor positive |
| ERBB2 | Erythroblastic leukemia viral oncogene homologue 2 |
| ERK1/2 | Extracellular signal-regulated kinases |
| EZH2 | Enhancer of zeste homolog 2 protein |
| Git2 | Guanosine triphosphatase-activating protein gene |
| HATs | Histone acetyltransferases |
| HDACi | Histone deacetylase inhibitor |
| HDACs | Histone deacetylases |
| HMTs | Histone methyltransferases |
| HNRNPA1 | Heterogeneous nuclear ribonucleoprotein A1 |
| hTERT | Human telomerase reverse transcriptase |
| H3R2me2a | Asymmetrical dimethylation at arginine residue 2 of histone subunit 3 |
| HER2 | Human epidermal growth factor receptor 2 |
| H4R3me2s | Dimethylation at arginine residue 3 residue of histone subunit 4 |
| H3ac | Acetylation at histone subunit 3 |
| H4ac | Acetylation at histone subunit 4 |
| HAC6 | Histone deacetylase 6 |
| H3K4me3 | Trimethylation at histone subunit 3 at lysine 4 |
| H3R8me2a | Asymmetrical dimethylation at arginine residue 8 residue of histone subunit 3 |
| H3R8me2s | Dimethylation at arginine residue 8 residue of histone subunit 3 |
| H4K5 | Histone subunit 4 at lysine residue 5 |
| H3K9 | Histone subunit 3 at lysine residue 9 |
| H3K9me3 | Trimethylation at histone subunit 3 at lysine 9 |
| H4K8 | Histone subunit 4 at lysine residue 8 |
| H3K9ac | Acetylation at histone subunit 3 at lysine 9 |
| H4K12 | Histone subunit 4 at lysine residue 12 |
| H3K14 | Histone subunit 3 at lysine residue 14 |
| H4K16 | Histone subunit 4 at lysine residue 16 |
| H3K18 | Histone subunit 3 at lysine residue 18 |
| H3K23 | Histone subunit 3 at lysine residue 23 |
| H3K27me3 | Trimethylation at histone subunit 3 at lysine 27 |
| JAK | Janus kinase |
| Ki-67 | Kiel-67 antibody |
| Let-7b | MicroRNA Let-7b |
| LINEs | Long interspersed nuclear elements |
| lncRNA | Long non-coding RNA |
| MAPK | Mitogen-activated protein kinase |
| MBD2 | Methyl-CpG-binding domain protein 2 |
| MeCP2 | Methyl-CpG-binding protein 2 |
| MET | Mesenchymal-to-epithelial transformation |
| miRNA | MicroRNA |
| mTOR | Mammalian target of rapamycin |
| N terminal | Nitrogen terminal |
| ncRNA | Non-coding RNA |
| NF-κB | Nuclear factor kappa light chain enhancer of activated B cells |
| NSAIDs | Non-steroidal anti-inflammatory drugs |
| p300/CBP | CREB-binding protein and its homolog p300 |
| PALB2 | Partner and localizer of BRCA2 |
| PARP | Poly (ADP-ribose) polymerase |
| PEITC | Phenethyl isothiocyanate |
| PI3K | Phosphoinositide 3-Kinase |
| PI3Ki | Phosphoinositide 3-Kinase inhibitor |
| PIK3CA | Phosphatidylinositol-4,5-Bisphosphate 3-Kinase Catalytic Subunit Alpha |
| PR- | Progesterone receptor negative |
| PR+ | Progesterone receptor positive |
| PTEN | Phosphatase and Tensin homolog deleted on chromosome 10 |
| RAS | Rat sarcoma |
| RNA | Ribonucleic acid |
| RNAi | Interfering RNA |
| SAHA | Suberanilohydroxamic acid |
| SALL3 | Spalt Like Transcription Factor 3 coding gene |
| SEMA3A | Semaphorin 3A gene |
| SERMs | Selective estrogen receptor modulators |
| siRNA | Small interfering RNA |
| SIRT1 | Sirtuin 1 |
| SNPs | Single nucleotide polymorphism |
| STAT | Signal transducer and activator of transcription proteins |
| STK11 | Serine/threonine kinase 11 |
| TNBC | Triple-negative breast cancer |
| TP53 | Tumor protein 53 |
| TSGs | Tumor suppressor genes |
| Tyr355 | Tyrosine 355 residue |
References
- Ahmad, A. Breast Cancer Statistics: Recent Trends. Adv. Exp. Med. Biol. 2019, 1152, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA A Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Thorat, M.A.; Balasubramanian, R. Breast cancer prevention in high-risk women. Best Pract. Res. Clin. Obstet. Gynaecol. 2020, 65, 18–31. [Google Scholar] [CrossRef]
- Lee, C.I.; Chen, L.E.; Elmore, J.G. Risk-based Breast Cancer Screening: Implications of Breast Density. Med. Clin. N. Am. 2017, 101, 725–741. [Google Scholar] [CrossRef]
- Sun, Y.S.; Zhao, Z.; Yang, Z.N.; Xu, F.; Lu, H.J.; Zhu, Z.Y.; Shi, W.; Jiang, J.; Yao, P.P.; Zhu, H.P. Risk Factors and Preventions of Breast Cancer. Int. J. Biol. Sci. 2017, 13, 1387–1397. [Google Scholar] [CrossRef] [Green Version]
- Coughlin, S.S. Epidemiology of Breast Cancer in Women. Adv. Exp. Med. Biol. 2019, 1152, 9–29. [Google Scholar] [CrossRef] [PubMed]
- Nelson, H.D.; Fu, R.; Zakher, B.; Pappas, M.; McDonagh, M. Medication Use for the Risk Reduction of Primary Breast Cancer in Women: Updated Evidence Report and Systematic Review for the US Preventive Services Task Force. JAMA 2019, 322, 868–886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shieh, Y.; Tice, J.A. Medications for Primary Prevention of Breast Cancer. JAMA 2020, 324, 291–292. [Google Scholar] [CrossRef]
- Graham, D.; DiNome, M.L.; Ganz, P.A. Breast Cancer Risk–Reducing Medications. JAMA 2020, 324, 310. [Google Scholar] [CrossRef]
- Li, F.; Dou, J.; Wei, L.; Li, S.; Liu, J. The selective estrogen receptor modulators in breast cancer prevention. Cancer Chemother. Pharmacol. 2016, 77, 895–903. [Google Scholar] [CrossRef]
- Kasiotis, K.M.; Lambrinidis, G.; Fokialakis, N.; Tzanetou, E.N.; Mikros, E.; Haroutounian, S.A. Novel Carbonyl Analogs of Tamoxifen: Design, Synthesis, and Biological Evaluation. Front. Chem. 2017, 5, 71. [Google Scholar] [CrossRef] [Green Version]
- Kala, R.; Tollefsbol, T.O. A Novel Combinatorial Epigenetic Therapy Using Resveratrol and Pterostilbene for Restoring Estrogen Receptor-α (ERα) Expression in ERα-Negative Breast Cancer Cells. PLoS ONE 2016, 11, e0155057. [Google Scholar] [CrossRef]
- Lee, W.J.; Shim, J.-Y.; Zhu, B.T. Mechanisms for the Inhibition of DNA Methyltransferases by Tea Catechins and Bioflavonoids. Mol. Pharmacol. 2005, 68, 1018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kala, R.; Shah, H.N.; Martin, S.L.; Tollefsbol, T.O. Epigenetic-based combinatorial resveratrol and pterostilbene alters DNA damage response by affecting SIRT1 and DNMT enzyme expression, including SIRT1-dependent γ-H2AX and telomerase regulation in triple-negative breast cancer. BMC Cancer 2015, 15, 672. [Google Scholar] [CrossRef] [Green Version]
- Samavat, H.; Wu, A.H.; Ursin, G.; Torkelson, C.J.; Wang, R.; Yu, M.C.; Yee, D.; Kurzer, M.S.; Yuan, J.M. Green Tea Catechin Extract Supplementation Does Not Influence Circulating Sex Hormones and Insulin-Like Growth Factor Axis Proteins in a Randomized Controlled Trial of Postmenopausal Women at High Risk of Breast Cancer. J. Nutr. 2019, 149, 619–627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, X.; Qi, M.; Li, P.; Zhan, Y.; Shao, H. Apigenin in cancer therapy: Anti-cancer effects and mechanisms of action. Cell Biosci. 2017, 7, 50. [Google Scholar] [CrossRef] [Green Version]
- Imran, M.; Rauf, A.; Abu-Izneid, T.; Nadeem, M.; Shariati, M.A.; Khan, I.A.; Imran, A.; Orhan, I.E.; Rizwan, M.; Atif, M.; et al. Luteolin, a flavonoid, as an anticancer agent: A review. Biomed. Pharmacother. 2019, 112, 108612. [Google Scholar] [CrossRef]
- Selvakumar, P.; Badgeley, A.; Murphy, P.; Anwar, H.; Sharma, U.; Lawrence, K.; Lakshmikuttyamma, A. Flavonoids and Other Polyphenols Act as Epigenetic Modifiers in Breast Cancer. Nutrients 2020, 12, 761. [Google Scholar] [CrossRef] [Green Version]
- Steed, K.L.; Jordan, H.R.; Tollefsbol, T.O. SAHA and EGCG Promote Apoptosis in Triple-negative Breast Cancer Cells, Possibly Through the Modulation of cIAP2. Anticancer Res. 2020, 40, 9–26. [Google Scholar] [CrossRef]
- Li, B.; Lu, Y.; Yu, L.; Han, X.; Wang, H.; Mao, J.; Shen, J.; Wang, B.; Tang, J.; Li, C.; et al. miR-221/222 promote cancer stem-like cell properties and tumor growth of breast cancer via targeting PTEN and sustained Akt/NF-κB/COX-2 activation. Chem.-Biol. Interact. 2017, 277, 33–42. [Google Scholar] [CrossRef]
- Tuli, H.S.; Tuorkey, M.J.; Thakral, F.; Sak, K.; Kumar, M.; Sharma, A.K.; Sharma, U.; Jain, A.; Aggarwal, V.; Bishayee, A. Molecular Mechanisms of Action of Genistein in Cancer: Recent Advances. Front Pharm. 2019, 10, 1336. [Google Scholar] [CrossRef] [Green Version]
- Ziaei, S.; Halaby, R. Dietary Isoflavones and Breast Cancer Risk. Medicines 2017, 4, 18. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Chen, H.; Hardy, T.M.; Tollefsbol, T.O. Epigenetic Regulation of Multiple Tumor-Related Genes Leads to Suppression of Breast Tumorigenesis by Dietary Genistein. PLoS ONE 2013, 8, e54369. [Google Scholar] [CrossRef] [Green Version]
- Fabian, C.J.; Khan, S.A.; Garber, J.E.; Dooley, W.C.; Yee, L.D.; Klemp, J.R.; Nydegger, J.L.; Powers, K.R.; Kreutzjans, A.L.; Zalles, C.M.; et al. Randomized Phase IIB Trial of the Lignan Secoisolariciresinol Diglucoside in Premenopausal Women at Increased Risk for Development of Breast Cancer. Cancer Prev. Res. 2020, 13, 623. [Google Scholar] [CrossRef] [Green Version]
- Park, J.E.; Sun, Y.; Lim, S.K.; Tam, J.P.; Dekker, M.; Chen, H.; Sze, S.K. Dietary phytochemical PEITC restricts tumor development via modulation of epigenetic writers and erasers. Sci. Rep. 2017, 7, 40569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuran, D.; Pogorzelska, A.; Wiktorska, K. Breast Cancer Prevention-Is there a Future for Sulforaphane and Its Analogs? Nutrients 2020, 12, 1559. [Google Scholar] [CrossRef]
- Yang, F.; Wang, F.; Liu, Y.; Wang, S.; Li, X.; Huang, Y.; Xia, Y.; Cao, C. Sulforaphane induces autophagy by inhibition of HDAC6-mediated PTEN activation in triple negative breast cancer cells. Life Sci. 2018, 213, 149–157. [Google Scholar] [CrossRef] [PubMed]
- Meeran, S.M.; Patel, S.N.; Tollefsbol, T.O. Sulforaphane causes epigenetic repression of hTERT expression in human breast cancer cell lines. PLoS ONE 2010, 5, e11457. [Google Scholar] [CrossRef] [PubMed]
- Sirerol, J.A.; Rodríguez, M.L.; Mena, S.; Asensi, M.A.; Estrela, J.M.; Ortega, A.L. Role of Natural Stilbenes in the Prevention of Cancer. Oxidative Med. Cell. Longev. 2016, 2016, 3128951. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cottart, C.H.; Nivet-Antoine, V.; Laguillier-Morizot, C.; Beaudeux, J.L. Resveratrol bioavailability and toxicity in humans. Mol. Nutr. Food Res. 2010, 54, 7–16. [Google Scholar] [CrossRef]
- Sinha, D.; Sarkar, N.; Biswas, J.; Bishayee, A. Resveratrol for breast cancer prevention and therapy: Preclinical evidence and molecular mechanisms. Semin. Cancer Biol. 2016, 40–41, 209–232. [Google Scholar] [CrossRef]
- Jiang, Y.-Z.; Liu, Y.-R.; Xu, X.-E.; Jin, X.; Hu, X.; Yu, K.-D.; Shao, Z.-M. Transcriptome Analysis of Triple-Negative Breast Cancer Reveals an Integrated mRNA-lncRNA Signature with Predictive and Prognostic Value. Cancer Res. 2016, 76, 2105. [Google Scholar] [CrossRef] [Green Version]
- Dutra, L.A.; Heidenreich, D.; Silva, G.D.B.d.; Man Chin, C.; Knapp, S.; Santos, J.L.D. Dietary Compound Resveratrol Is a Pan-BET Bromodomain Inhibitor. Nutrients 2017, 9, 1172. [Google Scholar] [CrossRef] [Green Version]
- Shin, H.J.; Han, J.M.; Choi, Y.S.; Jung, H.J. Pterostilbene Suppresses both Cancer Cells and Cancer Stem-Like Cells in Cervical Cancer with Superior Bioavailability to Resveratrol. Molecules 2020, 25, 228. [Google Scholar] [CrossRef] [Green Version]
- Ko, H.S.; Lee, H.-J.; Kim, S.-H.; Lee, E.-O. Piceatannol Suppresses Breast Cancer Cell Invasion through the Inhibition of MMP-9: Involvement of PI3K/AKT and NF-κB Pathways. J. Agric. Food Chem. 2012, 60, 4083–4089. [Google Scholar] [CrossRef] [PubMed]
- Tsang, J.Y.S.; Tse, G.M. Molecular Classification of Breast Cancer. Adv. Anat. Pathol. 2020, 27, 27–35. [Google Scholar] [CrossRef]
- Santisteban, M.; Reynolds, C.; Barr Fritcher, E.G.; Frost, M.H.; Vierkant, R.A.; Anderson, S.S.; Degnim, A.C.; Visscher, D.W.; Pankratz, V.S.; Hartmann, L.C. Ki67: A time-varying biomarker of risk of breast cancer in atypical hyperplasia. Breast Cancer Res. Treat. 2010, 121, 431–437. [Google Scholar] [CrossRef] [PubMed]
- Provenzano, E.; Ulaner, G.A.; Chin, S.F. Molecular Classification of Breast Cancer. PET Clin. 2018, 13, 325–338. [Google Scholar] [CrossRef] [PubMed]
- Yeo, S.K.; Guan, J.L. Breast Cancer: Multiple Subtypes within a Tumor? Trends Cancer 2017, 3, 753–760. [Google Scholar] [CrossRef]
- Jackson, H.W.; Fischer, J.R.; Zanotelli, V.R.T.; Ali, H.R.; Mechera, R.; Soysal, S.D.; Moch, H.; Muenst, S.; Varga, Z.; Weber, W.P.; et al. The single-cell pathology landscape of breast cancer. Nature 2020, 578, 615–620. [Google Scholar] [CrossRef] [PubMed]
- Barnard, M.E.; Boeke, C.E.; Tamimi, R.M. Established breast cancer risk factors and risk of intrinsic tumor subtypes. Biochim. Biophys. Acta (BBA)-Rev. Cancer 2015, 1856, 73–85. [Google Scholar] [CrossRef]
- Yang, X.; Han, H.; De Carvalho, D.D.; Lay, F.D.; Jones, P.A.; Liang, G. Gene body methylation can alter gene expression and is a therapeutic target in cancer. Cancer Cell 2014, 26, 577–590. [Google Scholar] [CrossRef] [Green Version]
- Nassar, F.J.; Chamandi, G.; Tfaily, M.A.; Zgheib, N.K.; Nasr, R. Peripheral Blood-Based Biopsy for Breast Cancer Risk Prediction and Early Detection. Front. Med. 2020, 7, 28. [Google Scholar] [CrossRef]
- Pasha, H.A.; Rezk, N.A.; Riad, M.A. Circulating Cell Free Nuclear DNA, Mitochondrial DNA and Global DNA Methylation: Potential Noninvasive Biomarkers for Breast Cancer Diagnosis. Cancer Investig. 2019, 37, 432–439. [Google Scholar] [CrossRef] [PubMed]
- Park, S.Y.; Seo, A.N.; Jung, H.Y.; Gwak, J.M.; Jung, N.; Cho, N.Y.; Kang, G.H. Alu and LINE-1 hypomethylation is associated with HER2 enriched subtype of breast cancer. PLoS ONE 2014, 9, e100429. [Google Scholar] [CrossRef]
- Hansmann, T.; Pliushch, G.; Leubner, M.; Kroll, P.; Endt, D.; Gehrig, A.; Preisler-Adams, S.; Wieacker, P.; Haaf, T. Constitutive promoter methylation of BRCA1 and RAD51C in patients with familial ovarian cancer and early-onset sporadic breast cancer. Hum. Mol. Genet. 2012, 21, 4669–4679. [Google Scholar] [CrossRef] [PubMed]
- Ennour-Idrissi, K.; Dragic, D.; Durocher, F.; Diorio, C. Epigenome-wide DNA methylation and risk of breast cancer: A systematic review. BMC Cancer 2020, 20, 1048. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Wu, L.; Shu, X.-O.; Cai, Q.; Shu, X.; Li, B.; Guo, X.; Ye, F.; Michailidou, K.; Bolla, M.K.; et al. Genetically Predicted Levels of DNA Methylation Biomarkers and Breast Cancer Risk: Data From 228 951 Women of European Descent. J. Natl. Cancer Inst. 2020, 112, 295–304. [Google Scholar] [CrossRef]
- Shukla, S.; Penta, D.; Mondal, P.; Meeran, S.M. Epigenetics of Breast Cancer: Clinical Status of Epi-drugs and Phytochemicals. Adv. Exp. Med. Biol. 2019, 1152, 293–310. [Google Scholar] [CrossRef]
- Nindrea, R.D.; Harahap, W.A.; Aryandono, T.; Lazuardi, L. Association of BRCA1 Promoter Methylation with Breast Cancer in Asia: A Meta- Analysis. Asian Pac. J. Cancer Prev. 2018, 19, 885–889. [Google Scholar] [CrossRef]
- Iwamoto, T.; Yamamoto, N.; Taguchi, T.; Tamaki, Y.; Noguchi, S. BRCA1 promoter methylation in peripheral blood cells is associated with increased risk of breast cancer with BRCA1 promoter methylation. Breast Cancer Res. Treat. 2011, 129, 69–77. [Google Scholar] [CrossRef]
- Begam, N.; Jamil, K.; Raju, S.G. Promoter Hypermethylation of the ATM Gene as a Novel Biomarker for Breast Cancer. Asian Pac. J. Cancer Prev. 2017, 18, 3003–3009. [Google Scholar] [CrossRef] [PubMed]
- Potapova, A.; Hoffman, A.M.; Godwin, A.K.; Al-Saleem, T.; Cairns, P. Promoter hypermethylation of the PALB2 susceptibility gene in inherited and sporadic breast and ovarian cancer. Cancer Res. 2008, 68, 998–1002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masood, S.; El-Gabry, E.; Zhang, C.; Wang, Z. DNA Methylation of the hTERT Gene in Breast Cancer Revisited: Diagnostic and Clinical Implications. Lab. Med. 2016, 47, 293–299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ennour-Idrissi, K.; Dragic, D.; Issa, E.; Michaud, A.; Chang, S.-L.; Provencher, L.; Durocher, F.; Diorio, C. DNA Methylation and Breast Cancer Risk: An Epigenome-Wide Study of Normal Breast Tissue and Blood. Cancers 2020, 12, 3088. [Google Scholar] [CrossRef] [PubMed]
- Pasculli, B.; Barbano, R.; Parrella, P. Epigenetics of breast cancer: Biology and clinical implication in the era of precision medicine. Semin. Cancer Biol. 2018, 51, 22–35. [Google Scholar] [CrossRef]
- Marmorstein, R.; Zhou, M.M. Writers and readers of histone acetylation: Structure, mechanism, and inhibition. Cold Spring Harb. Perspect. Biol. 2014, 6, a018762. [Google Scholar] [CrossRef] [Green Version]
- Blanc, R.S.; Richard, S. Arginine Methylation: The Coming of Age. Mol. Cell 2017, 65, 8–24. [Google Scholar] [CrossRef] [Green Version]
- Daniel, M.; Tollefsbol, T.O. Epigenetic linkage of aging, cancer and nutrition. J. Exp. Biol. 2015, 218, 59–70. [Google Scholar] [CrossRef] [Green Version]
- Zhuang, J.; Huo, Q.; Yang, F.; Xie, N. Perspectives on the Role of Histone Modification in Breast Cancer Progression and the Advanced Technological Tools to Study Epigenetic Determinants of Metastasis. Front. Genet. 2020, 11, 603552. [Google Scholar] [CrossRef]
- Bhat, S.A.; Majid, S.; Wani, H.A.; Rashid, S. Diagnostic utility of epigenetics in breast cancer—A review. Cancer Treat. Res. Commun. 2019, 19, 100125. [Google Scholar] [CrossRef]
- Suvanto, M.; Beesley, J.; Blomqvist, C.; Chenevix-Trench, G.; Khan, S.; Nevanlinna, H. SNPs in lncRNA Regions and Breast Cancer Risk. Front. Genet. 2020, 11, 550. [Google Scholar] [CrossRef] [PubMed]
- Chandra Gupta, S.; Nandan Tripathi, Y. Potential of long non-coding RNAs in cancer patients: From biomarkers to therapeutic targets. Int. J. Cancer 2017, 140, 1955–1967. [Google Scholar] [CrossRef] [PubMed]
- Li, S.-Q.; Li, F.; Xiao, Y.; Wang, C.-M.; Tuo, L.; Hu, J.; Yang, X.-B.; Wang, J.-S.; Shi, W.-H.; Li, X.; et al. Comparison of long non-coding RNAs, microRNAs and messenger RNAs involved in initiation and progression of esophageal squamous cell carcinoma. Mol. Med. Rep. 2014, 10, 652–662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farina, N.H.; Ramsey, J.E.; Cuke, M.E.; Ahern, T.P.; Shirley, D.J.; Stein, J.L.; Stein, G.S.; Lian, J.B.; Wood, M.E. Development of a predictive miRNA signature for breast cancer risk among high-risk women. Oncotarget 2017, 8, 112170–112183. [Google Scholar] [CrossRef] [Green Version]
- Jang, J.Y.; Kim, Y.S.; Kang, K.N.; Kim, K.H.; Park, Y.J.; Kim, C.W. Multiple microRNAs as biomarkers for early breast cancer diagnosis. Mol. Clin. Oncol. 2021, 14, 31. [Google Scholar] [CrossRef]
- Zhang, C.; Wang, X.; Li, X.; Zhao, N.; Wang, Y.; Han, X.; Ci, C.; Zhang, J.; Li, M.; Zhang, Y. The landscape of DNA methylation-mediated regulation of long non-coding RNAs in breast cancer. Oncotarget 2017, 8, 51134–51150. [Google Scholar] [CrossRef] [Green Version]
- McFadden, E.J.; Hargrove, A.E. Biochemical Methods To Investigate lncRNA and the Influence of lncRNA:Protein Complexes on Chromatin. Biochemistry 2016, 55, 1615–1630. [Google Scholar] [CrossRef] [Green Version]
- Cheuk, I.W.Y.; Shin, V.Y.; Kwong, A. Detection of Methylated Circulating DNA as Noninvasive Biomarkers for Breast Cancer Diagnosis. J. Breast Cancer 2017, 20, 12–19. [Google Scholar] [CrossRef] [Green Version]
- Hong, H.C.; Chuang, C.H.; Huang, W.C.; Weng, S.L.; Chen, C.H.; Chang, K.H.; Liao, K.W.; Huang, H.D. A panel of eight microRNAs is a good predictive parameter for triple-negative breast cancer relapse. Theranostics 2020, 10, 8771–8789. [Google Scholar] [CrossRef]
- Hu, Z.; Dong, J.; Wang, L.-E.; Ma, H.; Liu, J.; Zhao, Y.; Tang, J.; Chen, X.; Dai, J.; Wei, Q.; et al. Serum microRNA profiling and breast cancer risk: The use of miR-484/191 as endogenous controls. Carcinogenesis 2012, 33, 828–834. [Google Scholar] [CrossRef] [Green Version]
- Marino, A.L.F.; Evangelista, A.F.; Vieira, R.A.C.; Macedo, T.; Kerr, L.M.; Abrahão-Machado, L.F.; Longatto-Filho, A.; Silveira, H.C.S.; Marques, M.M.C. MicroRNA expression as risk biomarker of breast cancer metastasis: A pilot retrospective case-cohort study. BMC Cancer 2014, 14, 739. [Google Scholar] [CrossRef] [Green Version]
- Zhu, M.; Lv, Q.; Huang, H.; Sun, C.; Pang, D.; Wu, J. Identification of a four-long non-coding RNA signature in predicting breast cancer survival. Oncol. Lett. 2020, 19, 221–228. [Google Scholar] [CrossRef] [Green Version]
- Zubor, P.; Kubatka, P.; Kajo, K.; Dankova, Z.; Polacek, H.; Bielik, T.; Kudela, E.; Samec, M.; Liskova, A.; Vlcakova, D.; et al. Why the Gold Standard Approach by Mammography Demands Extension by Multiomics? Application of Liquid Biopsy miRNA Profiles to Breast Cancer Disease Management. Int. J. Mol. Sci. 2019, 20, 2878. [Google Scholar] [CrossRef] [Green Version]
- Clarke, R.; Tyson, J.J.; Dixon, J.M. Endocrine resistance in breast cancer--An overview and update. Mol Cell Endocrinol 2015, 418 Pt 3, 220–234. [Google Scholar] [CrossRef] [Green Version]
- Ye, L.; Lin, C.; Wang, X.; Li, Q.; Li, Y.; Wang, M.; Zhao, Z.; Wu, X.; Shi, D.; Xiao, Y.; et al. Epigenetic silencing of SALL2 confers tamoxifen resistance in breast cancer. EMBO Mol. Med. 2019, 11, e10638. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Wang, J.; Ukai, M.; Sewon, K.; Chen, P.; Suzuki, Y.; Wang, H.; Aihara, K.; Okada-Hatakeyama, M.; Chen, L. Hunt for the tipping point during endocrine resistance process in breast cancer by dynamic network biomarkers. J. Mol. Cell Biol. 2019, 11, 649–664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mills, J.N.; Rutkovsky, A.C.; Giordano, A. Mechanisms of resistance in estrogen receptor positive breast cancer: Overcoming resistance to tamoxifen/aromatase inhibitors. Curr. Opin. Pharmacol. 2018, 41, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Shou, J.; Massarweh, S.; Osborne, C.K.; Wakeling, A.E.; Ali, S.; Weiss, H.; Schiff, R. Mechanisms of Tamoxifen Resistance: Increased Estrogen Receptor-HER2/neu Cross-Talk in ER/HER2–Positive Breast Cancer. J. Natl. Cancer Inst. 2004, 96, 926–935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merikhian, P.; Ghadirian, R.; Farahmand, L.; Mansouri, S.; Majidzadeh, A.K. MUC1 induces tamoxifen resistance in estrogen receptor-positive breast cancer. Expert Rev. Anticancer Ther. 2017, 17, 607–613. [Google Scholar] [CrossRef]
- Stone, A.; Valdés-Mora, F.; Gee, J.M.; Farrow, L.; McClelland, R.A.; Fiegl, H.; Dutkowski, C.; McCloy, R.A.; Sutherland, R.L.; Musgrove, E.A.; et al. Tamoxifen-induced epigenetic silencing of oestrogen-regulated genes in anti-hormone resistant breast cancer. PLoS ONE 2012, 7, e40466. [Google Scholar] [CrossRef] [Green Version]
- Sanchez-Spitman, A.B.; Swen, J.J.; Dezentje, V.O.; Moes, D.J.A.R.; Gelderblom, H.; Guchelaar, H.J. Clinical pharmacokinetics and pharmacogenetics of tamoxifen and endoxifen. Expert Rev. Clin. Pharmacol. 2019, 12, 523–536. [Google Scholar] [CrossRef] [PubMed]
- Pulliam, N.; Fang, F.; Ozes, A.R.; Tang, J.; Adewuyi, A.; Keer, H.; Lyons, J.; Baylin, S.B.; Matei, D.; Nakshatri, H.; et al. An Effective Epigenetic-PARP Inhibitor Combination Therapy for Breast and Ovarian Cancers Independent of BRCA Mutations. Clin. Cancer Res. 2018, 24, 3163–3175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boussios, S.; Abson, C.; Moschetta, M.; Rassy, E.; Karathanasi, A.; Bhat, T.; Ghumman, F.; Sheriff, M.; Pavlidis, N. Poly (ADP-Ribose) Polymerase Inhibitors: Talazoparib in Ovarian Cancer and Beyond. Drugs R&D 2020, 20, 55–73. [Google Scholar] [CrossRef] [Green Version]
- Hanahan, D.; Weinberg, R.A. Hallmarks of Cancer: The Next Generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [Green Version]
- Beetch, M.; Lubecka, K.; Shen, K.; Flower, K.; Harandi-Zadeh, S.; Suderman, M.; Flanagan, J.M.; Stefanska, B. Stilbenoid-Mediated Epigenetic Activation of Semaphorin 3A in Breast Cancer Cells Involves Changes in Dynamic Interactions of DNA with DNMT3A and NF1C Transcription Factor. Mol. Nutr. Food Res. 2019, 63, 1801386. [Google Scholar] [CrossRef]
- Zhu, W.; Qin, W.; Zhang, K.; Rottinghaus, G.E.; Chen, Y.C.; Kliethermes, B.; Sauter, E.R. Trans-resveratrol alters mammary promoter hypermethylation in women at increased risk for breast cancer. Nutr. Cancer 2012, 64, 393–400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aldawsari, F.S.; Aguayo-Ortiz, R.; Kapilashrami, K.; Yoo, J.; Luo, M.; Medina-Franco, J.L.; Velázquez-Martínez, C.A. Resveratrol-salicylate derivatives as selective DNMT3 inhibitors and anticancer agents. J. Enzym. Inhib. Med. Chem. 2016, 31, 695–703. [Google Scholar] [CrossRef]
- Chatterjee, B.; Ghosh, K.; Kanade, S.R. Resveratrol modulates epigenetic regulators of promoter histone methylation and acetylation that restores BRCA1, p53, p21CIP1 in human breast cancer cell lines. BioFactors 2019, 45, 818–829. [Google Scholar] [CrossRef]
- Carlos-Reyes, Á.; Lopez-Gonzalez, J.S.; Meneses-Flores, M.; Gallardo-Rincón, D.; Ruíz-García, E.; Marchat, L.; La Vega, H.A.-D.; De La Cruz, O.N.H.; López-Camarillo, C. Dietary Compounds as Epigenetic Modulating Agents in Cancer. Front. Genet. 2019, 10, 79. [Google Scholar] [CrossRef] [Green Version]
- Ren, Y.; Li, S.; Zhu, R.; Wan, C.; Song, D.; Zhu, J.; Cai, G.; Long, S.; Kong, L.; Yu, W. Discovery of STAT3 and Histone Deacetylase (HDAC) Dual-Pathway Inhibitors for the Treatment of Solid Cancer. J. Med. Chem. 2021, 64, 7468–7482. [Google Scholar] [CrossRef]
- Lee, H.; Zhang, P.; Herrmann, A.; Yang, C.; Xin, H.; Wang, Z.; Hoon, D.S.; Forman, S.J.; Jove, R.; Riggs, A.D.; et al. Acetylated STAT3 is crucial for methylation of tumor-suppressor gene promoters and inhibition by resveratrol results in demethylation. Proc. Natl. Acad. Sci. USA 2012, 109, 7765–7769. [Google Scholar] [CrossRef] [Green Version]
- Hagiwara, K.; Kosaka, N.; Yoshioka, Y.; Takahashi, R.-u.; Takeshita, F.; Ochiya, T. Stilbene derivatives promote Ago2-dependent tumour-suppressive microRNA activity. Sci. Rep. 2012, 2, 314. [Google Scholar] [CrossRef] [Green Version]
- Otsuka, K.; Yamamoto, Y.; Ochiya, T. Regulatory role of resveratrol, a microRNA-controlling compound, in HNRNPA1 expression, which is associated with poor prognosis in breast cancer. Oncotarget 2018, 9, 24718–24730. [Google Scholar] [CrossRef] [Green Version]
- Wells, J.P.; White, J.; Stirling, P.C. R Loops and Their Composite Cancer Connections. Trends Cancer 2019, 5, 619–631. [Google Scholar] [CrossRef]
- Krawczyk, H. The stilbene derivatives, nucleosides, and nucleosides modified by stilbene derivatives. Bioorg. Chem. 2019, 90, 103073. [Google Scholar] [CrossRef]
- Huang, Y.; Du, J.; Mi, Y.; Li, T.; Gong, Y.; Ouyang, H.; Hou, Y. Long Non-coding RNAs Contribute to the Inhibition of Proliferation and EMT by Pterostilbene in Human Breast Cancer. Front. Oncol. 2018, 8, 8. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Ye, X.L.; Xu, J.; Cao, M.G.; Fang, Z.Y.; Li, L.Y.; Guan, G.H.; Liu, Q.; Qian, Y.H.; Xie, D. The lncRNA H19 mediates breast cancer cell plasticity during EMT and MET plasticity by differentially sponging miR-200b/c and let-7b. Sci. Signal. 2017, 10, eaak9557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mukohara, T. PI3K mutations in breast cancer: Prognostic and therapeutic implications. Breast Cancer 2015, 7, 111–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Litton, J.K.; Burstein, H.J.; Turner, N.C. Molecular Testing in Breast Cancer. Am. Soc. Clin. Oncol. Educ. Book 2019, 39, e1–e7. [Google Scholar] [CrossRef]
- Spangle, J.M.; Roberts, T.M.; Zhao, J.J. The emerging role of PI3K/AKT-mediated epigenetic regulation in cancer. Biochim. Biophys. Acta Rev. Cancer 2017, 1868, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Toska, E.; Osmanbeyoglu, H.U.; Castel, P.; Chan, C.; Hendrickson, R.C.; Elkabets, M.; Dickler, M.N.; Scaltriti, M.; Leslie, C.S.; Armstrong, S.A.; et al. PI3K pathway regulates ER-dependent transcription in breast cancer through the epigenetic regulator KMT2D. Science 2017, 355, 1324–1330. [Google Scholar] [CrossRef] [Green Version]
- Fouqué, A.; Delalande, O.; Jean, M.; Castellano, R.; Josselin, E.; Malleter, M.; Shoji, K.F.; Hung, M.D.; Rampanarivo, H.; Collette, Y.; et al. A Novel Covalent mTOR Inhibitor, DHM25, Shows in Vivo Antitumor Activity against Triple-Negative Breast Cancer Cells. J. Med. Chem. 2015, 58, 6559–6573. [Google Scholar] [CrossRef] [PubMed]
- Mosele, F.; Stefanovska, B.; Lusque, A.; Tran Dien, A.; Garberis, I.; Droin, N.; Le Tourneau, C.; Sablin, M.P.; Lacroix, L.; Enrico, D.; et al. Outcome and molecular landscape of patients with PIK3CA-mutated metastatic breast cancer. Ann. Oncol. Off. J. Eur. Soc. Med. Oncol. 2020, 31, 377–386. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Nie, J.; Ma, X.; Wei, Y.; Peng, Y.; Wei, X. Targeting PI3K in cancer: Mechanisms and advances in clinical trials. Mol. Cancer 2019, 18, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rimawi, M.F.; De Angelis, C.; Contreras, A.; Pareja, F.; Geyer, F.C.; Burke, K.A.; Herrera, S.; Wang, T.; Mayer, I.A.; Forero, A.; et al. Low PTEN levels and PIK3CA mutations predict resistance to neoadjuvant lapatinib and trastuzumab without chemotherapy in patients with HER2 over-expressing breast cancer. Breast Cancer Res. Treat. 2018, 167, 731–740. [Google Scholar] [CrossRef]
- Hu, H.; Zhu, J.; Zhong, Y.; Geng, R.; Ji, Y.; Guan, Q.; Hong, C.; Wei, Y.; Min, N.; Qi, A.; et al. PIK3CA mutation confers resistance to chemotherapy in triple-negative breast cancer by inhibiting apoptosis and activating the PI3K/AKT/mTOR signaling pathway. Ann. Transl. Med. 2021, 9, 410. [Google Scholar] [CrossRef]
- Banik, K.; Ranaware, A.M.; Harsha, C.; Nitesh, T.; Girisa, S.; Deshpande, V.; Fan, L.; Nalawade, S.P.; Sethi, G.; Kunnumakkara, A.B. Piceatannol: A natural stilbene for the prevention and treatment of cancer. Pharmacol. Res. 2020, 153, 104635. [Google Scholar] [CrossRef]
- Bose, S.; Banerjee, S.; Mondal, A.; Chakraborty, U.; Pumarol, J.; Croley, C.R.; Bishayee, A. Targeting the JAK/STAT Signaling Pathway Using Phytocompounds for Cancer Prevention and Therapy. Cells 2020, 9, 1451. [Google Scholar] [CrossRef]
- Farrand, L.; Byun, S.; Kim, J.Y.; Im-Aram, A.; Lee, J.; Lim, S.; Lee, K.W.; Suh, J.-Y.; Lee, H.J.; Tsang, B.K. Piceatannol Enhances Cisplatin Sensitivity in Ovarian Cancer via Modulation of p53, X-linked Inhibitor of Apoptosis Protein (XIAP), and Mitochondrial Fission*. J. Biol. Chem. 2013, 288, 23740–23750. [Google Scholar] [CrossRef] [Green Version]
- Ragab, H.M.; Samy, N.; Afify, M.; El Maksoud, N.A.; Shaaban, H.M. Assessment of Ki-67 as a potential biomarker in patients with breast cancer. J. Genet. Eng. Biotechnol. 2018, 16, 479–484. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Kaufman, P.D. Ki-67: More than a proliferation marker. Chromosoma 2018, 127, 175–186. [Google Scholar] [CrossRef]
- Li, L.-T.; Wang, X.; Zhu, W.-T.; Qian, G.-W.; Pei, D.-S.; Zheng, J.-N. Reciprocal Role Of DNA Methylation And Sp1 Binding In Ki-67 Gene Transcription. Cancer Manag. Res. 2019, 11, 9749–9759. [Google Scholar] [CrossRef] [Green Version]
- Selmin, O.I.; Donovan, M.G.; Stillwater, B.J.; Neumayer, L.; Romagnolo, D.F. Epigenetic Regulation and Dietary Control of Triple Negative Breast Cancer. Front. Nutr. 2020, 7, 159. [Google Scholar] [CrossRef] [PubMed]
- Ávila-Gálvez, M.Á.; González-Sarrías, A.; Martínez-Díaz, F.; Abellán, B.; Martínez-Torrano, A.J.; Fernández-López, A.J.; Giménez-Bastida, J.A.; Espín, J.C. Disposition of Dietary Polyphenols in Breast Cancer Patients’ Tumors, and Their Associated Anticancer Activity: The Particular Case of Curcumin. Mol. Nutr. Food Res. 2021, 65, 2100163. [Google Scholar] [CrossRef]
- Ávila-Gálvez, M.Á.; Giménez-Bastida, J.A.; Espín, J.C.; González-Sarrías, A. Dietary Phenolics against Breast Cancer. A Critical Evidence-Based Review and Future Perspectives. Int. J. Mol. Sci. 2020, 21, 5718. [Google Scholar] [CrossRef] [PubMed]
- Chan, E.W.C.; Wong, C.W.; Tan, Y.H.; Foo, J.P.Y.; Wong, S.K.; Chan, H.T. Resveratrol and pterostilbene: A comparative overview of their chemistry, biosynthesis, plant sources and pharmacological properties. J. Appl. Pharm. Sci. 2019, 9, 124–129. [Google Scholar] [CrossRef] [Green Version]
- Ko, J.-H.; Sethi, G.; Um, J.-Y.; Shanmugam, M.K.; Arfuso, F.; Kumar, A.P.; Bishayee, A.; Ahn, K.S. The Role of Resveratrol in Cancer Therapy. Int. J. Mol. Sci. 2017, 18, 2589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El Khawand, T.; Courtois, A.; Valls, J.; Richard, T.; Krisa, S. A review of dietary stilbenes: Sources and bioavailability. Phytochem. Rev. 2018, 17, 1007–1029. [Google Scholar] [CrossRef]
- Sergides, C.; Chirilă, M.; Silvestro, L.; Pitta, D.; Pittas, A. Bioavailability and safety study of resveratrol 500 mg tablets in healthy male and female volunteers. Exp. Med 2016, 11, 164–170. [Google Scholar] [CrossRef] [Green Version]
- Ávila-Gálvez, M.Á.; García-Villalba, R.; Martínez-Díaz, F.; Ocaña-Castillo, B.; Monedero-Saiz, T.; Torrecillas-Sánchez, A.; Abellán, B.; González-Sarrías, A.; Espín, J.C. Metabolic Profiling of Dietary Polyphenols and Methylxanthines in Normal and Malignant Mammary Tissues from Breast Cancer Patients. Mol. Nutr. Food Res. 2019, 63, 1801239. [Google Scholar] [CrossRef]
- Peng, R.-M.; Lin, G.-R.; Ting, Y.; Hu, J.-Y. Oral delivery system enhanced the bioavailability of stilbenes: Resveratrol and pterostilbene. BioFactors 2018, 44, 5–15. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Parvathaneni, V.; Shukla, S.K.; Kanabar, D.D.; Muth, A.; Gupta, V. Cyclodextrin Complexation for Enhanced Stability and Non-invasive Pulmonary Delivery of Resveratrol-Applications in Non-small Cell Lung Cancer Treatment. AAPS PharmSciTech 2020, 21, 183. [Google Scholar] [CrossRef] [PubMed]
- Kershaw, J.; Kim, K.-H. The Therapeutic Potential of Piceatannol, a Natural Stilbene, in Metabolic Diseases: A Review. J. Med. Food 2017, 20, 427–438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papoutsis, A.J.; Selmin, O.I.; Borg, J.L.; Romagnolo, D.F. Gestational exposure to the AhR agonist 2,3,7,8-tetrachlorodibenzo-p-dioxin induces BRCA-1 promoter hypermethylation and reduces BRCA-1 expression in mammary tissue of rat offspring: Preventive effects of resveratrol. Mol. Carcinog. 2015, 54, 261–269. [Google Scholar] [CrossRef]
- Harris, R.E.; Casto, B.C.; Harris, Z.M. Cyclooxygenase-2 and the inflammogenesis of breast cancer. World J. Clin. Oncol. 2014, 5, 677–692. [Google Scholar] [CrossRef] [Green Version]
- Regulski, M.; Piotrowska, H.; Prukała, W.; Dutkiewicz, Z.; Regulska, K.; Stanisz, B.; Murias, M. Synthesis, in vitro and in silico evaluation of novel trans -stilbene analogues as potential COX-2 inhibitors. Bioorg. Med. Chem. 2018, 26, 141–151. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ganguly, S.; Arora, I.; Tollefsbol, T.O. Impact of Stilbenes as Epigenetic Modulators of Breast Cancer Risk and Associated Biomarkers. Int. J. Mol. Sci. 2021, 22, 10033. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms221810033
Ganguly S, Arora I, Tollefsbol TO. Impact of Stilbenes as Epigenetic Modulators of Breast Cancer Risk and Associated Biomarkers. International Journal of Molecular Sciences. 2021; 22(18):10033. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms221810033
Chicago/Turabian StyleGanguly, Sebanti, Itika Arora, and Trygve O. Tollefsbol. 2021. "Impact of Stilbenes as Epigenetic Modulators of Breast Cancer Risk and Associated Biomarkers" International Journal of Molecular Sciences 22, no. 18: 10033. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms221810033