
Analysis of Molecular Networks in the Cerebellum in Chronic Schizophrenia: Modulation by Early Postnatal Life Stressors in Murine Models

 , , and
, , and
Abstract
:1. Introduction
2. Results
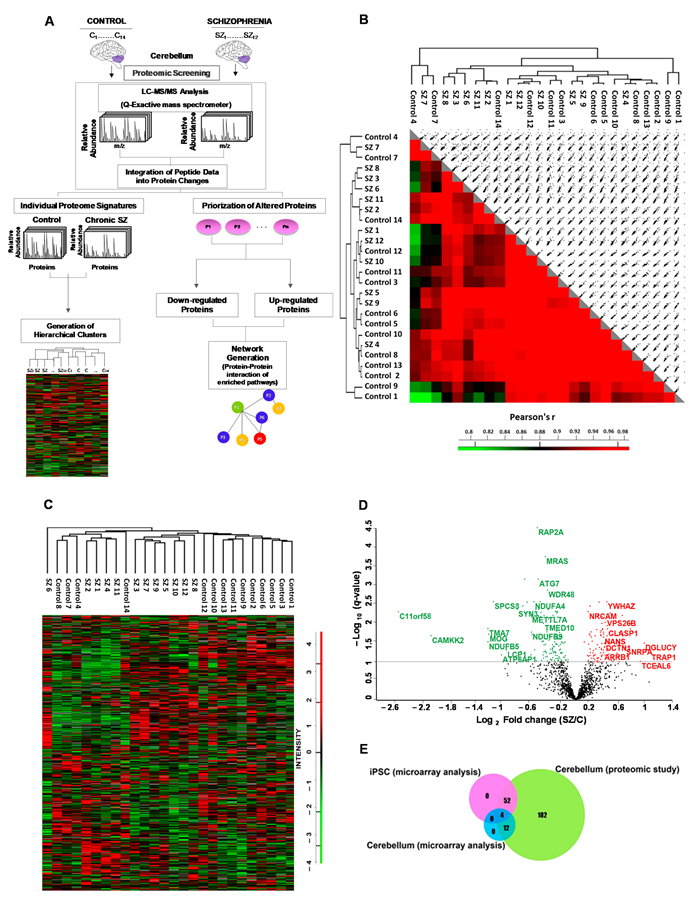
2.1. Quantitative Proteomic Analyses in Cerebellum in Chronic Schizophrenia
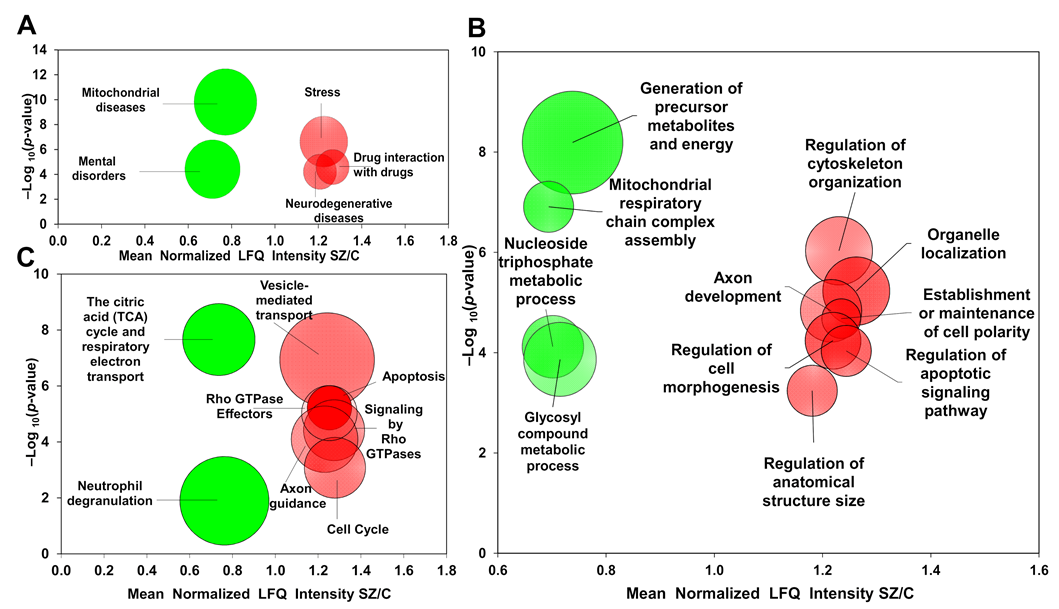
2.2. Gene Ontology Enrichment Analysis
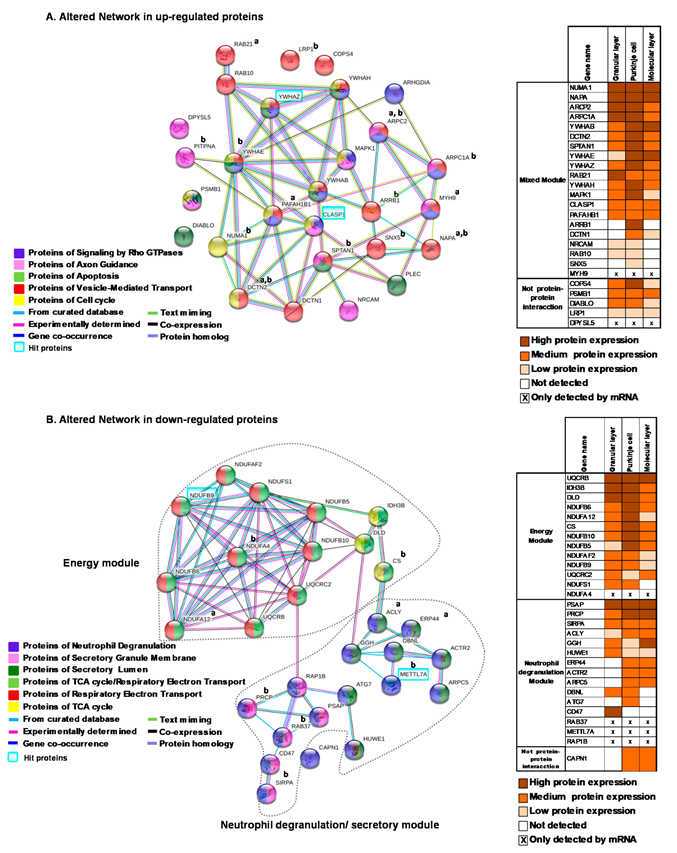
2.3. Network Generation from Enriched Pathways in the Cerebellum
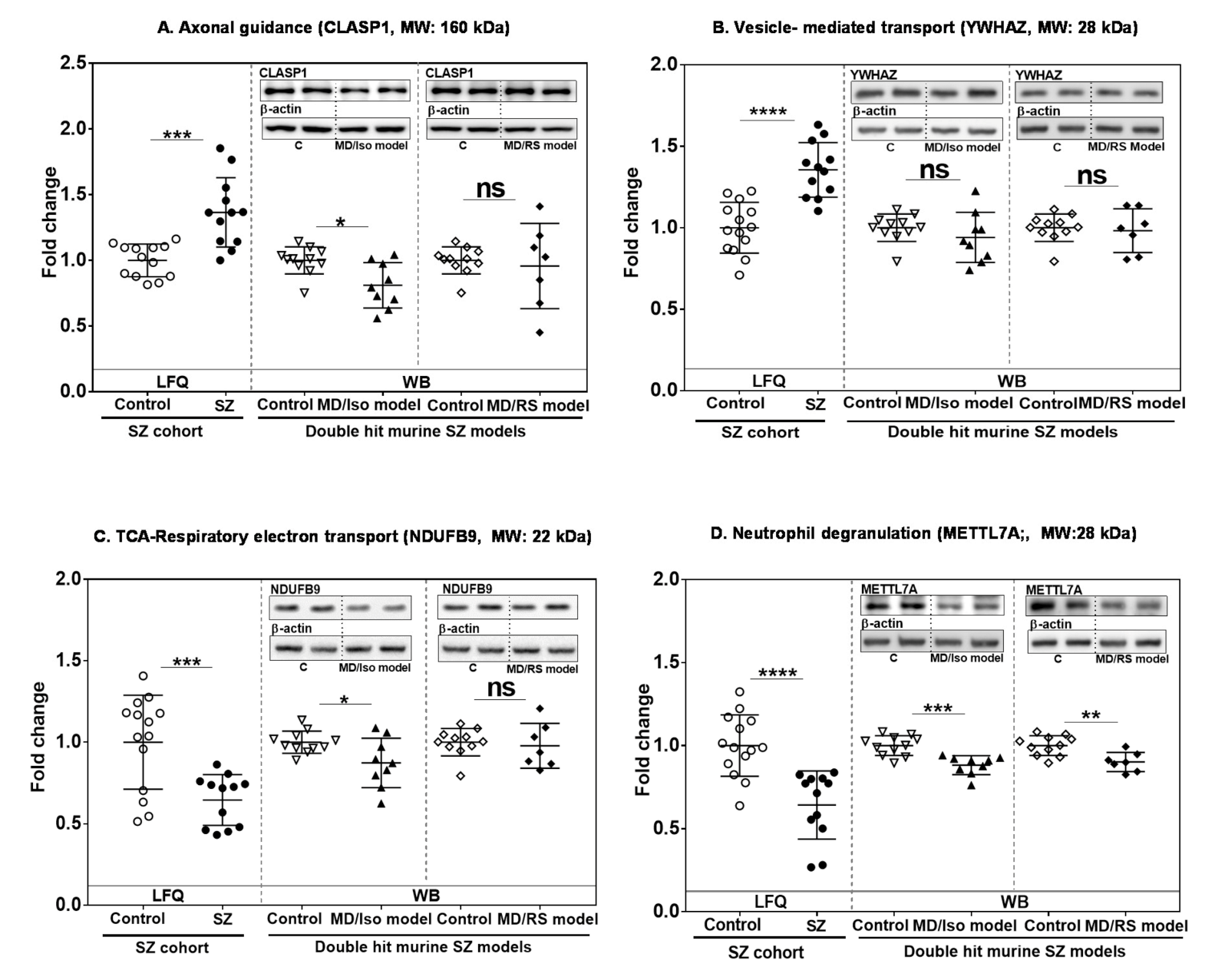
2.4. Analysis of Altered Robust Candidates in Double-Hit Stress Murine Models
3. Discussion
3.1. Up-Regulated Proteins Related to Stress and Axonal Functions
3.1.1. Vesicle-Mediated Transport
3.1.2. Axon Guidance
3.2. Down-Regulated Proteins Related to Mitochondrial Disease, Energy Functions and Immune Response
3.2.1. Energy Metabolism Module
3.2.2. Neutrophil Degranulation Module
4. Methods and Materials
4.1. Postmortem Human Brain Tissue
4.2. Label Free Quantification Proteomic Analysis Mass Spectrometry
4.3. Bioinformatic Analysis
4.4. Stress Murine Models
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| SZ | Schizophrenia |
| LC–MS/MS | Liquid chromatography–tandem mass spectrometry |
| LFQ | Label free quantitative |
| DSM-IV | Diagnostic and Statistical Manual of Mental Disorders |
| PMD | Postmortem delay |
| FDR | False discovery rate |
| GO | Gene ontology |
| GLAD4U | Gene List Automatically Derived for You |
| iPSC | induced pluripotent stem cells |
| EVs | Extracellular vesicles |
| TCA | The citric acid |
| PD | Postnatal day |
| MD/Iso | Maternal deprivation with isolation |
| MD/RS | Maternal deprivation with restraint stress |
| PharmGKB | PharmacoGenetics Knowledge Base |
| METTL7A | Methyltransferase-like 7A |
| NDUFB9 | NADH dehydrogenase [ubiquinone] 1 beta subcomplex subunit 9 |
| CLASP1 | CLIP-associating protein 1 |
| YWHAZ | 14-3-3 protein zeta/delta |
| COMT | Catechol O-methyltransferase |
| WB | Western Blot |
References
- Rosato, M.; Stringer, S.; Gebuis, T.; Paliukhovich, I.; Li, K.W.; Posthuma, D.; Sullivan, P.F.; Smit, A.B.; van Kesteren, R.E. Combined cellomics and proteomics analysis reveals shared neuronal morphology and molecular pathway phenotypes for multiple schizophrenia risk genes. Mol. Psychiatry 2021, 26, 784–799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGrath, J.; Saha, S.; Chant, D.; Welham, J. Schizophrenia: A concise overview of incidence, prevalence, and mortality. Epidemiol Rev. 2008, 30, 67–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hilker, R.; Helenius, D.; Fagerlund, B.; Skytthe, A.; Christensen, K.; Werge, T.M.; Nordentoft, M.; Glenthøj, B. Heritability of Schizophrenia and Schizophrenia Spectrum Based on the Nationwide Danish Twin Register. Biol. Psychiatry 2018, 83, 492–498. [Google Scholar] [CrossRef] [Green Version]
- Davis, J.; Eyre, H.; Jacka, F.N.; Dodd, S.; Dean, O.; McEwen, S.; Debnath, M.; McGrath, J.; Maes, M.; Amminger, G.P.; et al. A review of vulnerability and risks for schizophrenia: Beyond the two hit hypothesis. Neurosci. Biobehav. Rev. 2016, 65, 185–194. [Google Scholar] [CrossRef] [Green Version]
- Fatemi, S.H.; Folsom, T.D. The neurodevelopmental hypothesis of Schizophrenia, revisited. Schizophr. Bull. 2009, 35, 528–548. [Google Scholar] [CrossRef]
- Maynard, T.; Sikich, L.; Lieberman, J.A.; LaMantia, A.-S. Neural development, cell-cell signaling, and the “two-hit” hypothesis of schizophrenia. Schizophr. Bull. 2001, 27, 457–476. [Google Scholar] [CrossRef] [Green Version]
- Weinberger, D.R. Implications of Normal Brain Development for the Pathogenesis of Schizophrenia. Arch. Gen. Psychiatry 1988, 45, 1055. [Google Scholar] [CrossRef]
- Holtzman, C.; Trotman, H.; Goulding, S.; Ryan, A.; MacDonald, A.; Shapiro, D.; Brasfield, J.; Walker, E. Stress and neurodevelopmental processes in the emergence of psychosis. Neuroscience 2013, 249, 172–191. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Téllez, R.I.; Hernández-Torres, E.; Gamboa, C.; Flores, G. Prenatal stress alters spine density and dendritic length of nucleus accumbens and hippocampus neurons in rat offspring. Synapse 2009, 63, 794–804. [Google Scholar] [CrossRef]
- Shalev, U.; Weiner, I. Gender-dependent differences in latent inhibition following prenatal stress and corticosterone administration. Behav. Brain Res. 2001, 126, 57–63. [Google Scholar] [CrossRef]
- Lemaire, V.; Koehl, M.; Le Moal, M.; Abrous, D.N. Prenatal stress produces learning deficits associated with an inhibition of neurogenesis in the hippocampus. Proc. Natl. Acad. Sci. USA 2000, 97, 11032–11037. [Google Scholar] [CrossRef] [Green Version]
- Son, G.H.; Geum, D.; Chung, S.; Kim, E.J.; Jo, J.-H.; Kim, C.-M.; Lee, K.H.; Kim, H.T.; Choi, S.; Lee, C.J.; et al. Maternal stress produces learning deficits associated with impairment of NMDA receptor-mediated synaptic plasticity. J. Neurosci. 2006, 26, 3309–3318. [Google Scholar] [CrossRef]
- Weiss, I.C.; Pryce, C.R.; Jongen-Rêlo, A.L.; Nanz-Bahr, N.I.; Feldon, J. Effect of social isolation on stress-related behavioural and neuroendocrine state in the rat. Behav. Brain Res. 2004, 152, 279–295. [Google Scholar] [CrossRef]
- Giovanoli, S.; Engler, H.; Engler, A.; Richetto, J.; Feldon, J.; A Riva, M.; Schedlowski, M.; Meyer, U. Preventive effects of minocycline in a neurodevelopmental two-hit model with relevance to schizophrenia. Transl. Psychiatry 2016, 6, e772. [Google Scholar] [CrossRef] [Green Version]
- Monte, A.S.; Mello, B.S.F.; Borella, V.C.M.; da Silva Araujo, T.D.S.; Da Silva, F.E.R.; De Sousa, F.C.F.; De Oliveira, A.C.P.; Gama, C.S.; Seeman, M.V.; Vasconcelos, S.M.M.; et al. Two-hit model of schizophrenia induced by neonatal immune activation and peripubertal stress in rats: Study of sex differences and brain oxidative alterations. Behav. Brain Res 2017, 331, 30–37. [Google Scholar] [CrossRef]
- Weinberger, D.R. On the plausibility of “the neurodevelopmental hypothesis” of schizophrenia. Neuropsychopharmacology 1996, 14, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Marco, E.M.; Llorente, R.; Gallardo, M.L.; Mela, V.; Llorente-Berzal, Á.; Prada, C.; Viveros, M.-P. The maternal deprivation animal model revisited. Neurosci. Biobehav. Rev. 2015, 51, 151–163. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Rius, J. The cerebellum under stress. Front. Neuroendocrinol 2019, 54, 100774. [Google Scholar] [CrossRef] [PubMed]
- De Quervain, D.J.F.; Henke, K.; Aerni, A.; Treyer, V.; McGaugh, J.L.; Berthold, T.; Nitsch, R.M.; Buck, A.; Roozendaal, B.; Hock, C. Glucocorticoid-induced impairment of declarative memory retrieval is associated with reduced blood flow in the medial temporal lobe. Eur. J. Neurosci. 2003, 17, 1296–1302. [Google Scholar] [CrossRef]
- Hare, S.M.; Chiappelli, J.; Savransky, A.; Adhikari, B.M.; Wisner, K.; Kvarta, M.; Goldwaser, E.; Du, X.; Chen, S.; Rowland, L.M.; et al. The Role of Hippocampal Functional Connectivity on Multisystem Subclinical Abnormalities in Schizophrenia. Psychosom. Med. 2020, 82, 623–630. [Google Scholar] [CrossRef]
- van Leeuwen, J.M.; Vinkers, C.H.; Vink, M.; Kahn, R.S.; Joëls, M.; Hermans, E.J. Disrupted upregulation of salience network connectivity during acute stress in siblings of schizophrenia patients. Psychol. Med. 2021, 51, 1038–1048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quidé, Y.; Girshkin, L.; Watkeys, O.J.; Carr, V.J.; Green, M.J. The relationship between cortisol reactivity and emotional brain function is differently moderated by childhood trauma, in bipolar disorder, schizophrenia and healthy individuals. Eur. Arch. Psychiatry Clin. Neurosci. 2020, 271, 1089–1109. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.-J.; Huang, J.; Lui, S.S.Y.; Cheung, E.F.C.; Madsen, K.H.; Chan, R.C.K. The effect of effort-reward imbalance on brain structure and resting-state functional connectivity in individuals with high levels of schizotypal traits. Cogn. Neuropsychiatry 2021, 26, 166–182. [Google Scholar] [CrossRef] [PubMed]
- Altman, J. Postnatal development of the cerebellar cortex in the rat. II. Phases in the maturation of Purkinje cells and of the molecular layer. J. Comp. Neurol. 1972, 145, 399–463. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Cheng, L.; Grennan, K.; Pibiri, F.; Zhang, C.; A Badner, J.; Gershon, E.S.; Liu, C. Two gene co-expression modules differentiate psychotics and controls. Mol. Psychiatry 2013, 18, 1308–1314. [Google Scholar] [CrossRef] [Green Version]
- Vidal-Domènech, F.; Riquelme, G.; Pinacho, R.; Rodriguez-Mias, R.; Vera, A.; Monje, A.; Ferrer, I.; Callado, L.F.; Meana, J.J.; Villén, J.; et al. Calcium-binding proteins are altered in the cerebellum in schizophrenia. PLoS ONE 2020, 15, e0230400. [Google Scholar] [CrossRef]
- Reis-De-Oliveira, G.; Zuccoli, G.; Fioramonte, M.; Schimitt, A.; Falkai, P.; Almeida, V.; Martins-De-Souza, D. Digging deeper in the proteome of different regions from schizophrenia brains. J. Proteom. 2020, 223, 103814. [Google Scholar] [CrossRef]
- Brennand, K.; Simone, A.; Jou, J.; Gelboin-Burkhart, C.; Tran, N.; Sangar, S.; Li, Y.; Mu, Y.; Chen, G.; Yu, D.; et al. Modelling schizophrenia using human induced pluripotent stem cells. Nature 2011, 473, 221–225. [Google Scholar] [CrossRef]
- Ramaker, R.C.; Bowling, K.M.; Lasseigne, B.N.; Hagenauer, M.H.; Hardigan, A.A.; Davis, N.S.; Gertz, J.; Cartagena, P.M.; Walsh, D.M.; Vawter, M.P.; et al. Post-mortem molecular profiling of three psychiatric disorders. Genome Med. 2017, 9, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Owen, M.J.; Williams, N.M.; O’Donovan, M. The molecular genetics of schizophrenia: New findings promise new insights. Mol. Psychiatry 2004, 9, 14–27. [Google Scholar] [CrossRef] [Green Version]
- Armbruster, D.; Mueller, A.; Strobel, A.; Lesch, K.-P.; Brocke, B.; Kirschbaum, C. Children under stress-COMT genotype and stressful life events predict cortisol increase in an acute social stress paradigm. Int. J. Neuropsychopharmacol. 2012, 15, 1229–1239. [Google Scholar] [CrossRef] [Green Version]
- Walder, D.J.; Trotman, H.D.; Cubells, J.F.; Brasfield, J.; Tang, Y.-L.; Walker, E.F. Catechol-O-methyltransferase modulation of cortisol secretion in psychiatrically at-risk and healthy adolescents. Psychiatr. Genet. 2010, 20, 166. Available online: https://journals.lww.com/psychgenetics/Fulltext/2010/08000/Catechol_O_methyltransferase_modulation_of.5.aspx (accessed on 22 June 2021). [CrossRef] [Green Version]
- Heinzel, S.; Riemer, T.G.; Schulte, S.; Onken, J.; Heinz, A.; Rapp, M.A. Catechol-O-methyltransferase (COMT) genotype affects age-related changes in plasticity in working memory: A pilot study. Biomed. Res. Int. 2014, 2014, 1–7. [Google Scholar] [CrossRef]
- Egan, M.F.; Goldberg, T.E.; Kolachana, B.S.; Callicott, J.H.; Mazzanti, C.M.; Straub, R.E.; Goldman, D.; Weinberger, D.R. Effect of COMT Val108/158 Met genotype on frontal lobe function and risk for schizophrenia. Proc. Natl. Acad. Sci. USA 2001, 98, 6917–6922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mizuno, Y.; Jung, M.; Fujisawa, T.X.; Takiguchi, S.; Shimada, K.; Saito, D.N.; Kosaka, H.; Tomoda, A. Catechol-O-methyltransferase polymorphism is associated with the cortico-cerebellar functional connectivity of executive function in children with attention-deficit/hyperactivity disorder. Sci. Rep. 2017, 7, 1–8. [Google Scholar]
- Barlan, K.; Rossow, M.J.; Gelfand, V.I. The journey of the organelle: Teamwork and regulation in intracellular transport. Curr. Opin. Cell Biol. 2013, 25, 483–488. [Google Scholar] [CrossRef] [Green Version]
- Maher, B.J.; LoTurco, J.J. Disrupted-in-schizophrenia (DISC1) functions presynaptically at glutamatergic synapses. PLoS ONE 2012, 7, e34053. [Google Scholar] [CrossRef] [PubMed]
- Mirnics, K.; Middleton, F.; Marquez, A.; Lewis, D.; Levitt, P. Molecular Characterization of Schizophrenia Viewed by Microarray Analysis of Gene Expression in Prefrontal Cortex. Neuron 2000, 28, 53–67. Available online: https://www.cell.com/neuron/fulltext/S08966273(00)000854?_returnURL=https%3A%2F%2Flinkinghub.elsevier.com%2Fretrieve%2Fpii%2FS0896627300000854%3Fshowall%3Dtrue# (accessed on 7 December 2020). [CrossRef] [Green Version]
- Camargo, L.M.; Collura, V.; Rain, J.-C.; Mizuguchi, K.; Hermjakob, H.; Kerrien, S.; Bonnert, T.P.; Whiting, P.; Brandon, N. Disrupted in Schizophrenia 1 interactome: Evidence for the close connectivity of risk genes and a potential synaptic basis for schizophrenia. Mol. Psychiatry 2007, 12, 74–86. [Google Scholar] [CrossRef] [PubMed]
- Honer, W.G.; Falkai, P.; Bayer, T.A.; Xie, J.; Hu, L.; Li, H.-Y.; Arango, V.; Mann, J.J.; Dwork, A.J.; Trimble, W.S. Abnormalities of SNARE Mechanism Proteins in Anterior Frontal Cortex in Severe Mental Illness. Cereb. Cortex 2002, 12, 349–356. Available online: https://0-academic-oup-com.brum.beds.ac.uk/cercor/article-lookup/doi/10.1093/cercor/12.4.349 (accessed on 30 November 2020). [CrossRef] [Green Version]
- Mukaetova-Ladinska, E.; Hurt, J.; Honer, W.; Harrington, C.; Wischik, C. Loss of synaptic but not cytoskeletal proteins in the cerebellum of chronic schizophrenics. Neurosci. Lett. 2002, 317, 161–165. Available online: http://0-www-sciencedirect-com.brum.beds.ac.uk/science/article/pii/S0304394001024582 (accessed on 7 December 2020). [CrossRef]
- Martins-De-Souza, D.; Maccarrone, G.; Wobrock, T.; Zerr, I.; Gormanns, P.; Reckow, S.; Falkai, P.; Schmitt, A.; Turck, C.W. Proteome analysis of the thalamus and cerebrospinal fluid reveals glycolysis dysfunction and potential biomarkers candidates for schizophrenia. J. Psychiatr. Res. 2010, 44, 1176–1189. [Google Scholar] [CrossRef]
- Saia-Cereda, V.M.; Cassoli, J.S.; Schmitt, A.; Falkai, P.; Nascimento, J.M.; Martins-De-Souza, D. Proteomics of the corpus callosum unravel pivotal players in the dysfunction of cell signaling, structure, and myelination in schizophrenia brains. Eur. Arch. Psychiatry Clin. Neurosci. 2015, 65, 601–612. [Google Scholar] [CrossRef] [PubMed]
- Drago, F.; Lombardi, M.; Prada, I.; Gabrielli, M.; Joshi, P.; Cojoc, D.; Franck, J.; Fournier, I.; Vizioli, J.; Verderio, C. ATP modifies the proteome of extracellular vesicles released by microglia and influences their action on astrocytes. Front. Pharmacol. 2017, 8, 910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, S.-Y.; Huang, P.-H.; Cheng, H.-J. Disrupted-in-Schizophrenia 1-mediated axon guidance involves TRIO-RAC-PAK small GTPase pathway signaling. Proc. Natl. Acad. Sci. USA 2011, 108, 5861–5866. [Google Scholar] [CrossRef] [Green Version]
- Friston, K.; Brown, H.R.; Siemerkus, J.; Stephan, K.E. The dysconnection hypothesis. Schizophr. Res. 2016, 176, 83–94. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Li, P.; Wu, T.; Zhu, S.; Deng, L.; Cui, G. Axon guidance pathway genes are associated with schizophrenia risk. Exp. Ther. Med. 2018, 16, 4519–4526. [Google Scholar] [CrossRef] [Green Version]
- Sayas, C.L.; Basu, S.; Van Der Reijden, M.; Bustos-Morán, E.; Liz, M.; Sousa, M.; van Ijcken, W.; Avila, J.; Galjart, N. Distinct Functions for Mammalian CLASP1 and -2 During Neurite and Axon Elongation. Front. Cell. Neurosci. 2019, 13, 1–17. [Google Scholar] [CrossRef]
- Rajasekaran, A.; Venkatasubramanian, G.; Berk, M.; Debnath, M. Mitochondrial dysfunction in schizophrenia: Pathways, mechanisms and implications. Neurosci. Biobehav. Rev. 2015, 48, 10–21. [Google Scholar] [CrossRef] [PubMed]
- Semendeferi, K.; Damasio, H. The brain and its main anatomical subdivisions in living hominoids using magnetic resonance imaging. J. Hum. Evol. 2000, 38, 317–332. Available online: https://0-www-sciencedirect-com.brum.beds.ac.uk/science/article/pii/S0047248499903810 (accessed on 3 March 2021). [CrossRef]
- Howarth, C.; Gleeson, P.; Attwell, D. Updated energy budgets for neural computation in the neocortex and cerebellum. J. Cereb. Blood Flow Metab. 2012, 32, 1222–1232. [Google Scholar] [CrossRef] [PubMed]
- Nopoulos, P.; Ceilley, J.W.; Gailis, E.A.; Andreasen, N.C. An MRI study of cerebellar vermis morphology in patients with schizophrenia: Evidence in support of the cognitive dysmetria concept. Biol. Psychiatry 1999, 46, 703–711. [Google Scholar] [CrossRef]
- Koziol, L.F.; Budding, D.; Andreasen, N.; D’Arrigo, S.; Bulgheroni, S.; Imamizu, H.; Ito, M.; Manto, M.; Marvel, C.; Parker, K.; et al. Consensus paper: The cerebellum’s role in movement and cognition. Cerebellum 2014, 13, 151–177. [Google Scholar] [CrossRef]
- Verge, B.; Alonso, Y.; Valero, J.; Miralles, C.; Vilella, E.; Martorell, L. Mitochondrial DNA (mtDNA) and schizophrenia. Eur. Psychiatry 2011, 26, 45–56. [Google Scholar] [CrossRef] [PubMed]
- Prabakaran, S.; Swatton, J.E.; Ryan, M.M.; Huffaker, S.J.; Huang, J.; Griffin, J.L.; Wayland, M.; Freeman, T.; Dudbridge, F.; Lilley, K.S.; et al. Mitochondrial dysfunction in schizophrenia: Evidence for compromised brain metabolism and oxidative stress. Mol. psychiatry 2004, 9, 684–697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maurer, I.; Zierz, S.; Möller, H.J. Evidence for a mitochondrial oxidative phosphorylation defect in brains from patients with schizophrenia. Schizophr. Res. 2001, 48, 125–136. [Google Scholar] [CrossRef]
- Roberts, R.C. Postmortem studies on mitochondria in schizophrenia. Schizophr. Res. 2017, 187, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Zuccoli, G.S.; Saia-Cereda, V.M.; Nascimento, J.; Martins-De-Souza, D. The energy metabolism dysfunction in psychiatric disorders postmortem brains: Focus on proteomic evidence. Front. Neurosci. 2017, 11, 493. [Google Scholar] [CrossRef]
- Martins-De-Souza, D.; Gattaz, W.F.; Schmitt, A.; Novello, J.C.; Marangoni, S.; Turck, C.W.; Dias-Neto, E. Proteome analysis of schizophrenia patients Wernicke’s area reveals an energy metabolism dysregulation. BMC Psychiatry 2009, 9, 17. [Google Scholar] [CrossRef] [Green Version]
- English, J.A.; Pennington, K.; Dunn, M.J.; Cotter, D.R. The Neuroproteomics of Schizophrenia. Biol. Psychiatry 2011, 69, 163–172. [Google Scholar] [CrossRef]
- Bubber, P.; Hartounian, V.; Gibson, G.; Blass, J. Abnormalities in the tricarboxylic acid (TCA) cycle in the brains of schizophrenia patients. Eur. Neuropsychopharmacol. 2011, 21, 254–260. [Google Scholar] [CrossRef] [Green Version]
- Martins-De-Souza, D.; Harris, L.; Guest, P.; Bahn, S. The role of energy metabolism dysfunction and oxidative stress in schizophrenia revealed by proteomics. Antioxid. Redox Signal. 2011, 15, 2067–2079. [Google Scholar] [CrossRef] [PubMed]
- Rollins, B.L.; Morgan, L.; Hjelm, B.E.; Sequeira, A.; Schatzberg, A.F.; Barchas, J.D.; Lee, F.S.; Myers, R.M.; Watson, S.J.; Akil, H.; et al. Mitochondrial Complex I Deficiency in Schizophrenia and Bipolar Disorder and Medication Influence. Mol. Neuropsychiatry 2017, 3, 157–169. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, S.; Rosowski, E.E.; Huttenlocher, A. Neutrophil migration in infection and wound repair: Going forward in reverse. Nat. Rev. Immunol. 2016, 16, 378–391. [Google Scholar] [CrossRef]
- Kinney, D.K.; Hintz, K.; Shearer, E.M.; Barch, D.H.; Riffin, C.; Whitley, K.; Butler, R. A unifying hypothesis of schizophrenia: Abnormal immune system development may help explain roles of prenatal hazards, post-pubertal onset, stress, genes, climate, infections, and brain dysfunction. Med. Hypotheses 2010, 74, 555–563. [Google Scholar] [CrossRef] [PubMed]
- McKinnon, C.M.; Mellor, H. The tumor suppressor RhoBTB1 controls Golgi integrity and breast cancer cell invasion through METTL7B. BMC Cancer. 2017, 17, 145. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.; Choi, K.H.; Baykiz, A.F.; Gershenfeld, H.K. Suicide candidate genes associated with bipolar disorder and schizophrenia: An exploratory gene expression profiling analysis of post-mortem prefrontal cortex. BMC Genom. 2007, 8, 413. [Google Scholar] [CrossRef] [Green Version]
- Föcking, M.; Lopez, L.; A English, J.; Dicker, P.; Wolff, A.; Brindley, E.; Wynne, K.; Cagney, G.; Cotter, D. Proteomic and genomic evidence implicates the postsynaptic density in schizophrenia. Mol. Psychiatry 2015, 20, 424–432. [Google Scholar] [CrossRef]
- Scarr, E.; Udawela, M.; A Thomas, E.; Dean, B. Changed gene expression in subjects with schizophrenia and low cortical muscarinic M1 receptors predicts disrupted upstream pathways interacting with that receptor. Mol. Psychiatry 2018, 23, 295–303. [Google Scholar] [CrossRef] [Green Version]
- Gardner, D.M.; Murphy, A.L.; O’Donnell, H.; Centorrino, F.; Baldessarini, R.J. International consensus study of antipsychotic dosing. Am. J. Psychiatry 2010, 167, 686–693. [Google Scholar] [CrossRef] [Green Version]
- Pinacho, R.; Villalmanzo, N.; Lalonde, J.; Haro, J.M.; Meana, J.J.; Gill, G.; Ramos, B. The transcription factor SP4 is reduced in postmortem cerebellum of bipolar disorder subjects: Control by depolarization and lithium. Bipolar Disord. 2011, 13, 474–485. [Google Scholar] [CrossRef] [Green Version]
- Yosef, B.Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B 1995, 57, 289–300. [Google Scholar]
- Tyanova, S.; Temu, T.; Sinitcyn, P.; Carlson, A.; Hein, M.Y.; Geiger, T.; Mann, M.; Cox, J. The Perseus computational platform for comprehensive analysis of (prote)omics data. Nat. Methods 2016, 13, 731–740. [Google Scholar] [CrossRef]
- Wu, Y.; Yao, Y.-G.; Luo, X.-J. SZDB: A Database for Schizophrenia Genetic Research. Schizophr. Bull. 2017, 43, 459–471. [Google Scholar] [CrossRef] [Green Version]
- Pathan, M.; Keerthikumar, S.; Ang, C.-S.; Gangoda, L.; Quek, C.; Williamson, N.; Mouradov, D.; Sieber, O.; Simpson, R.; Salim, A.; et al. FunRich: An open access standalone functional enrichment and interaction network analysis tool. Proteomics 2015, 15, 2597–2601. [Google Scholar] [CrossRef]
- Hewett, M.; Oliver, D.E.; Rubin, D.L.; Easton, K.L.; Stuart, J.M.; Altman, R.B.; Altman, R.B.; Klein, T.E. BiGCaT Bioinformatics\rPharmGKB: The Pharmacogenetics Knowledge Base. Nucleic Acids Res. 2002, 30, 163–165. Available online: http://www.bigcat.nl (accessed on 7 March 2021). [CrossRef]
- Jourquin, J.; Duncan, D.; Shi, Z.; Zhang, B. GLAD4U: Deriving and prioritizing gene lists from PubMed literature. BMC Genom. 2012, 13 (Suppl. 8), S20. Available online: http://0-www-biomedcentral-com.brum.beds.ac.uk/1471-2164/13/S8/S20 (accessed on 20 December 2020). [CrossRef] [PubMed] [Green Version]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, 607–613. [Google Scholar] [CrossRef] [Green Version]
- Uhlén, M.; Fagerberg, L.; Hallström, B.M.; Lindskog, C.; Oksvold, P.; Mardinoglu, A.; Sivertsson, Å.; Kampf, C.; Sjöstedt, E.; Asplund, A.; et al. Tissue-based map of the human proteome. Science 2015, 347, 1260419. [Google Scholar]
- Bailoo, J.D.; Varholick, J.A.; Garza, X.J.; Jordan, R.L.; Hintze, S. Maternal separation followed by isolation-housing differentially affects prepulse inhibition of the acoustic startle response in C57BL/6 mice. Dev. Psychobiol. 2016, 58, 937–944. [Google Scholar] [CrossRef] [PubMed]
- Van Zyl, P.J.; Dimatelis, J.J.; Russell, V.A. Behavioural and biochemical changes in maternally separated Sprague—Dawley rats exposed to restraint stress. Metab. Brain Dis. 2016, 31, 121–133. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CLASSIFICATION | CATEGORY | PROTEIN OVERLAP IN CATEGORY | Total Number | Observed Number | E | p-Value | FDR |
|---|---|---|---|---|---|---|---|
| Up-regulated proteins | |||||||
| DISEASES | Stress | SYCRIP; COMT; TXN2; PDIA3 b; HSPA9; MYH9 a; NACA b; MAPK1; RPS3; SOD1 b; YWHAE b; YWHAZ; USP7 b | 592 | 13 | 2.20 | 2.30 × 10–7 | 6.02 × 10–4 |
| Drug interaction with drug | HSPA9; LRP1 b; ARRB1 b; PPIA; MAPK1; DIABLO; TPT1; YWHAB; YWHAZ | 423 | 9 | 1.57 | 2.56 × 10–5 | 3.36 × 10–2 | |
| Neurodegenerative diseases | DCTN1; HSPA9; LRP1 b; MAP1B; RTN1 b; SNCG; SNRPD1; SOD1 b; UCHL1 | 473 | 9 | 1.76 | 6.11 × 10–5 | 5.33 × 10–2 | |
| BIOLOGICAL PROCESSES | Regulation of cytoskeleton organization | ARPC2 b; ARPC1A b; DCTN1; CLASP1; ARHGDIA; LRP1 b; PAFAH1B1 a; MAPK1; RPS3; SPTAN1 b; PM1; CAPZA2 | 414 | 12 | 2.08 | 9.29 × 10–7 | 7.08 × 10–4 |
| Organelle localization | DCTN2 b; DCTN1; CLASP1; MAP1B; MYH9 a; NUMA1 b; PAFAH1B1 a; UCHL1; YWHAZ; CADPS; NAPA a,b; LIN7A b | 495 | 12 | 2.49 | 5.91 × 10–6 | 2.25 × 10–3 | |
| Axon development | RAB10; RAB21 a; ARHGDIA; MAP1B; NRCAM; PAFAH1B1 a; PITPNA b; MAPK1; DPYSL5; SPTAN1 b; UCHL1 | 452 | 11 | 2.27 | 1.46 × 10–5 | 3.71 × 10–5 | |
| Establishment or maintenance of cell polarity | RAB10; CLASP1; MAP1B; MYH9 a; NUMA1 b; PAFAH1B1 a; LIN7A b | 168 | 7 | 0.85 | 2.10 × 10–5 | 4.0 × 10–3 | |
| Regulation of cell morphogenesis | ARPC2 b; RAB21 a; ARHGDIA; MAP1B; MYH9 a; NRCAM; PAFAH1B1 a; C1QBP; TPM1; YWHAH | 433 | 10 | 2.18 | 5.78 × 10–5 | 6.29 × 10–3 | |
| Regulation of apoptotic Signaling pathway | PDIA3b; HNRNPK; RPS3; SOD1b; TPT1; YWHAB; YWHAE b; YWHAH; YWHAZ | 369 | 9 | 1.86 | 9.24 × 10–5 | 8.80 × 10–3 | |
| Multicellular organismal signaling | NRCAM; ATP2B1; ATP2B3; PAFAH1B1 a; SOD1 b; YWHAE b | 472 | 9 | 2.38 | 5.74 × 10–3 | 3.36 × 10–2 | |
| Regulation of anatomical structure size | ARPC2 b; ARPC1A b; RAB21 a; MAP1B; NRCAM; PAFAH1B1 a; SOD1 b; SPTAN1 b; CAPZA2 | 472 | 9 | 2.36 | 5.74 × 10–4 | 3.36 × 10–2 | |
| Cytosolic transport | VPS26B; DCTN1; RAB21 a; SNX5 b; MAPK1 | 138 | 5 | 0.69 | 6.46 × 10–4 | 3.51 v 10–2 | |
| Microtubule-basedmovement | DCTN1; RAB21 a; MAP1B; PAFAH1B1 a; SOD1 b; UCHL1 | 226 | 6 | 1.14 | 9.50 × 10–4 | 4.83 × 10–2 | |
| PATHWAYS | Vesicle-mediated transport | ARPC2 b; DCTN2 b; ARPC1A b; RAB10; DCTN1; RAB21 a; SNX5 b; LRP1 b; ARRB1 b; PAFAH1B1 a; COPS4; SPTAN1 b; YWHAB; YWHAE b; YWHAH; YWHAZ; NAPA a,b | 670 | 17 | 3.84 | 1.17 × 10–7 | 1.87 × 10–4 |
| Apoptosis | PLEC; DIABLO; PSMB1; SPTAN1 b; YWHAB; YWHAE b; YWHAH; YWHAZ | 174 | 8 | 1 | 6.07 × 10–3 | 1.57 × 10–3 | |
| Rho GTPase Effectors | ARPC2 b; ARPC1A b; CLASP1; MYH9 a; PAFAH1B1 a; MAPK1; YWHAB; YWHAE b; YWHAH; YWHAZ | 311 | 10 | 1.78 | 9.32 × 10–6 | 1.86 × 10–3 | |
| Signaling by Rho GTPases | ARPC2 b; ARPC1A b; CLASP1; ARHGDIA; MYH9 a; PAFAH1B1 a; MAPK1; YWHAB; YWHAE b; YWHAH; YWHAZ | 446 | 11 | 2.56 | 3.82 × 10–5 | 5.10 × 10–3 | |
| Axon guidance | ARPC2 b; ARPC1A b; CLASP1; ARRB1 b; MYH9 a; NRCAM; PITPNA b; MAPK1; PSMB1; DPYSL5; SPTAN1 b; YWHAB | 573 | 12 | 3.28 | 8.00 × 10–5 | 8.00 × 10–3 | |
| Cell Cycle | DCTN2 b; DCTN1; CLASP1; NUMA1 b; PAFAH1B1 a; MAPK1; PSMB1; YWHAB; YWHAE b; YWHAH; YWHAZ | 635 | 11 | 3.64 | 8.40 × 10–4 | 5.60 × 10–2 | |
| Down-regulated proteins | |||||||
| DISEASES | Mitochondrial diseases | DLD; SLC25A4b; SLC25A5; HSPD1; ACADVLb; NDUFA4b; NDUFB5; NDUFB6; NDUFB9; NDUFS1; NDUFA12a; SOD2; UQCRB; SLC25A22; CAPN1; SLC25A12b; NDUFAF2 | 353 | 17 | 2.32 | 1.53 × 10–10 | 4.0 × 10–7 |
| Mental disorders | ADH5; GABRA6 b; PCLO b; ANK3 b; MARK1; MOG; ATP1A3 b; PIP4K2A b; SLC17A7; PTPRD b; SMS; SYP b; SYN3 b; PICALM b; SLC25A12 b | 679 | 15 | 4.45 | 3.83 × 10–5 | 2.01 × 10–2 | |
| BIOLOGICAL PROCESSES | Generation of precursor metabolites and energy | COX17; CS b; DLD; GPD1; SLC25A4 b; IDH3B; ACADVL b; NDUFA4 b; NDUFB5; NDUFB6; NDUFB9; NDUFB10; NDUFS1; NDUFA12 a; UQCRB; UQCRC2; SLC25A12 b; NDUFAF2 | 365 | 18 | 3.36 | 6.47 × 10–9 | 4.93 × 10–6 |
| Mitochondrial respiratory chain complex assembly | COX17; NDUFB5; NDUFB6; NDUFB9; NDUFB10; NDUFS1; NDUFA12 a; UQCRB; NDUFA2 | 290 | 11 | 2.67 | 7.61 × 10–5 | 9.67 × 10–3 | |
| Nucleoside triphosphate metabolic process | DLD; GPD1; NDUFA4b; NDUFB5; NDUFB6; NDUFB9; NDUFB10; NDUFS1; NDUFA12a; UQCRB; UQCRC2 | 421 | 13 | 3.88 | 1.36 × 10–4 | 1.48 × 10–2 | |
| Glycosyl compound metabolic process | DLD; AHCY a; GPD1; NDUFA4b; NDUFB5; NDUFB6; NDUFB9; NDUFB10; NDUFS1; GMPR2; NDUFA12a; UQCRB; UQCRC2 | 34 | 4 | 3.13 | 2.59 × 10–4 | 2.45 × 10–2 | |
| ATP hydrolysis-coupled transmembrane transport | ATP1A1 b; ATP1A3 b; ATP6V1C1; ATP6AP1 b | 37 | 4 | 0.34 | 3.59 × 10–4 | 3.04 × 10–2 | |
| Tricarboxylic acid metabolic process | CS b; DLD; IDH3B; ACLY a | 37 | 4 | 0.34 | 3.59 × 10–5 | 3.04 × 10–3 | |
| PATHWAYS | The citric acid (TCA) cycle and respiratory electron transport | CS b; DLD; IDH3B; NDUFA4 b; NDUFB5; NDUFB6; NDUFB9; NDUFB10; NDUFS1; NDUFA12 a; UQCRB; UQCRC2; NDUFAF2 | 171 | 13 | 1.79 | 2.19 × 10–8 | 3.50 × 10–5 |
| Neutrophil degranulation | HUWE1; ARPC5; ACTR2; ATG7; SIRPA b; ERP44 a,b; METTL7A b; DBNL; RAB37 b; ACLY a; PRCP b; PSAP; RAP1B; CAPN1; GGH; CD47 | 487 | 16 | 5.07 | 4.0 × 10–5 | 1.28 × 10–2 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vera-Montecinos, A.; Rodríguez-Mias, R.; MacDowell, K.S.; García-Bueno, B.; Bris, Á.G.; Caso, J.R.; Villén, J.; Ramos, B. Analysis of Molecular Networks in the Cerebellum in Chronic Schizophrenia: Modulation by Early Postnatal Life Stressors in Murine Models. Int. J. Mol. Sci. 2021, 22, 10076. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms221810076
Vera-Montecinos A, Rodríguez-Mias R, MacDowell KS, García-Bueno B, Bris ÁG, Caso JR, Villén J, Ramos B. Analysis of Molecular Networks in the Cerebellum in Chronic Schizophrenia: Modulation by Early Postnatal Life Stressors in Murine Models. International Journal of Molecular Sciences. 2021; 22(18):10076. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms221810076
Chicago/Turabian StyleVera-Montecinos, América, Ricard Rodríguez-Mias, Karina S. MacDowell, Borja García-Bueno, Álvaro G. Bris, Javier R. Caso, Judit Villén, and Belén Ramos. 2021. "Analysis of Molecular Networks in the Cerebellum in Chronic Schizophrenia: Modulation by Early Postnatal Life Stressors in Murine Models" International Journal of Molecular Sciences 22, no. 18: 10076. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms221810076