1. Introduction
All species constitutively produce antimicrobial peptides (AMPs) to defend the host from undesired microbial invasion [
1]. Venoms, on the other hand, are toxic and complex secretions applied through the sting, teeth, fangs, or harpoons of venomous animals, such as insects, snakes, cone snails, and spiders [
2]. The primary use of venom is to fight aggressors and deter predators, in addition to paralyzing and/or killing and preserving prey. Interestingly, depending on the organism’s habitat, many venoms are evolutionarily optimized to also contain broad-spectrum antimicrobial peptides. Most of these short, amphiphilic, and often cationic AMPs tend to act on bacterial membranes and hence show an intrinsic tendency to destabilize the lipid bilayer. This property of AMPs has been exploited since their discovery in the 1990s, with the aim of optimizing their potential against microbial infections, evaluating their cytotoxicity toward cancer cells, and reducing their hemolytic effects against erythrocytes, amongst other potential applications. AMPs are essential components of the venom of many (but not all) spiders. From an estimated 150,000 species of spiders, only a quarter (approximately 43,000) of spider species are cataloged, and the venom components of only 174 spiders have been described [
3,
4,
5]. These data clearly indicate the dearth of information available on potentially promising molecules that are currently lacking attention and waiting to be optimized for a given application. By some estimate, spider venom has been predicted to contain over 10 million bioactive peptides [
6] with a wide range of activities, including analgesic, antiparasitic, cytolytic, antitumor, and anti-inflammatory activities. However, only a minuscule fraction of these peptides have been studied or biophysically characterized to be exploited for human benefit.
In this work, we focus on the membrane interactions of one such group of seven linear helical peptides called latarcins (
Table 1). Latarcins are the C-terminal fragments of larger precursor proteins isolated from the venom of the spider
Lachesana tarabaevi. They consist of a signal peptide, followed by an acidic fragment that is cleaved off post-translationally, resulting in one common group of seven AMPs. They were discovered, isolated and characterized by Kozlov et al. in 2006 [
7]. Latarcins were isolated from crude venom by HPLC fractionation, characterized by mass spectrometry, and sequenced using amino acid analysis.
Table 1 shows the primary structure of latarcins. It was found that Ltc3a, Ltc4a, and Ltc5a are amidated; the other four latarcins bear a free -COOH group at the C-terminus. Once the sequence identity had been established, the peptides were chemically synthesized. As with the majority of AMPs, latarcins were found to be unstructured in water, whereas, in 50% trifluoroethanol (TFE), they are able to fold into α-helices [
7]. It is well known that TFE promotes α-helical structures [
8,
9] and that a majority of membrane-active peptides tend to fold into helices in this solvent system. These helices, however, may or may not represent the state of the peptide when it interacts with the target biomembrane.
Latarcins have been demonstrated to exhibit high antimicrobial, cytolytic, and membrane perturbing activity, with the exception of Ltc6a and Ltc7 [
7]. While the exact role of these multifunctional peptides in the spider venom remains unclear, it appears that their major contribution is toward the paralyzing and cytolytic effects [
7]. Furthermore, based on the observed synergistic effects of the venom components with other neurotoxins, it is also proposed that latarcins may facilitate the passage of neurotoxins through cellular barriers and sterilize the prey, in addition to protecting the venom glands from external microbial infections [
10]. Most investigations in the past have restricted their focus to Ltc1 [
11] or Ltc2a [
12]. Based on circular dichroism spectroscopy (CD) and solution NMR data in SDS micelles and restrained Monte Carlo simulations, a membrane-bound structure has been presented for these two selected peptides. In an attempt to fine-tune the antimicrobial property of Ltc2a, it was further shown that the hydrophobic N-terminal part in Ltc2a triggers hemolytic side effects [
13]. The same has not been shown for other latarcins. Although
L. tarabaevi is reported to be a rich source of linear membrane-active compounds [
14], i.e., latarcins and other peptides, there has been no study thus far in which all members of the latarcin family have been systematically studied in the presence of membranes. Comparative studies on all members of the latarcin family include only a determination of their antimicrobial and hemolytic activities [
7,
10]. Biophysical studies have only been performed on Ltc1 and Ltc2a [
7,
11,
12].
All latarcins are positively charged (largely Lys rather than Arg) and contain several aromatic amino acids (
Table 1), but they do not show any sequence homology.
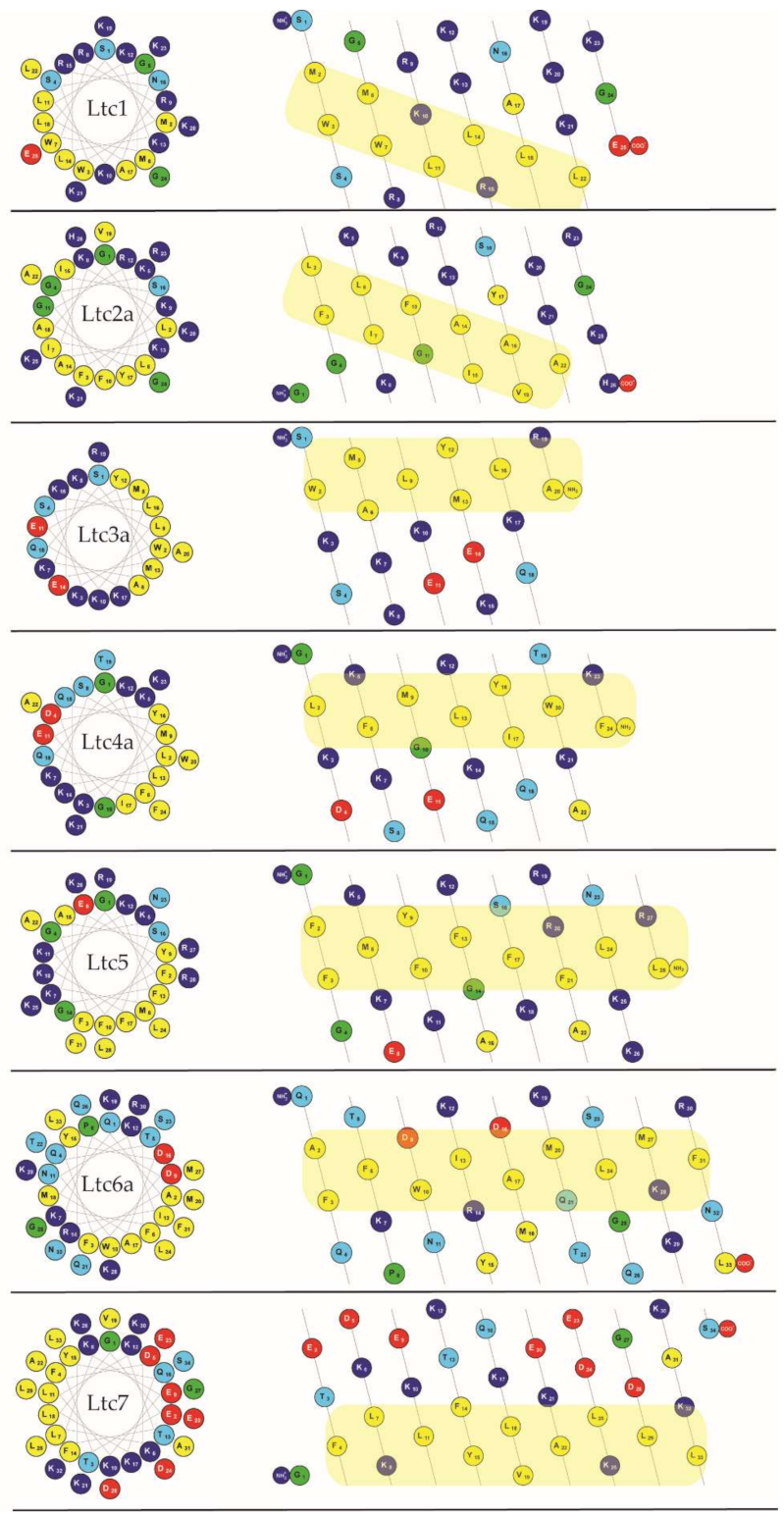
Figure 1 (left column) illustrates how all latarcins can form amphipathic structures when folded as an α-helix, as the hydrophobic residues are segregated on one face of the helical wheel and the charged (and polar) ones on the opposite face. This property implies that they should all bind favorably to lipid membranes in this conformation. However, there are some noticeable differences among this group of peptides. For example, the net charge varies from +2 to +10, and when considering the peptide lengths between 20 to 34 amino acids, the average net charge per residue varies from +0.06 to +0.40. According to the sequences and other properties listed in
Table 1 as well as the patterns shown in
Figure 1, we propose here and will proceed to group the latarcins into four types with related structural properties:
Group A peptides (Ltc1 and Ltc2a) consist of 25–26 residues and have a free C-terminus. They carry the highest net charge, resulting from 9 to 10 exclusively cationic groups without any anionic residues, besides 9–11 hydrophobic amino acids. In the helical mesh representation, it can be seen (better than in the helical wheel) that the strip of hydrophobic residues runs diagonally along/around the helix axis. Notably, in Ltc1, the hydrophobic strip is disrupted by two cationic residues (Lys10; Arg15), which is not the case in Ltc2a.
Group B peptides (Ltc3a and Ltc4a) are the shortest peptides with 20-24 residues and possess an amidated C-terminus. Their net charge of +6 results from 8 positively and 2 negatively charged groups, besides 8–9 hydrophobic amino acids. They display a typical amphiphilic profile, with a straight strip of hydrophobic residues that are flanked by cationic residues (nicely seen in the helical wheel representation).
Group C comprises only a single peptide (Ltc5) with 28 residues and an amidated C-terminus. It has a net charge of +10 and an unusually high number of 7 aromatic amino acids (mainly Phe) amongst 12 hydrophobic ones. The helical mesh representation indicates that the hydrophobic strip becomes less defined towards the C-terminus.
Group D (Ltc6a and Ltc7) are the longest peptides with 33–34 residues, possessing a free C-terminus. Ltc6a is rich in polar residues, while Ltc7 is particularly rich in charged residues (8 Glu/Asp and 10 Lys). The resulting net charge of +5 and +2, respectively, is relatively low. Some 12–13 hydrophobic residues contribute to a long and well-defined amphiphilic profile.
Given the similar properties, we expect that the peptides within each group might also display similar activities in the presence of membranes. For example, one can directly predict from the charge that peptides in Groups A and C (Ltc1, Ltc2a and Ltc5), which have the highest charge per residue of 0.36, should be favorably attracted to bacterial membranes that are generally anionic. Previous studies have shown some indication in this direction. In contrast, Group D peptides (Ltc6a and Ltc7), which have a lower net charge, are expected to be less active against bacteria, while Group B should be moderately active. From the helical wheels and mesh representations shown in
Figure 1, one can also predict that all latarcins should be able to fold into amphipathic helices and thereby interact with lipid membranes. Previous studies have only shown a helical fold in TFE, and there are no data on the entire set of latarcins to demonstrate their folding into helices upon binding to a membrane. With the current understanding of the physical properties of peptides, it is not possible, however, to predict how such helical peptides will behave when bound to a membrane, i.e., whether they will stay on the surface, or become tilted, or insert as oligomers, or have any tendency to aggregate non-productively. It is also not possible to predict whether they would form any transient or stable pores or cause any damage to the lipid bilayer that can be assessed by monitoring vesicle leakage.
Therefore, for the first time, we have performed a combined study of the full set of all seven latarcins using several biophysical techniques and biological assays. In particular, we have characterized the structure of these antimicrobial peptides in lipid bilayers using CD, oriented CD (OCD), and solid-state
15N-NMR methods to assess the predicted and previously proposed structures [
10,
11,
12] of the peptides per se and in the presence of lipid bilayers. Using solution CD, one can determine whether they are indeed helical in the presence of lipids, as implied in
Figure 1 and demonstrated in TFE [
7]. If they are helical, it is important to know how they integrate into the membrane. This information can provide important clues on the mechanism of action of these peptides on the target cell, i.e., whether they stay folded or rather aggregate, or assemble to form pores in the bilayer, or have any other targets inside the bacterial cell. For this purpose, we used OCD and solid-state
15N-NMR to determine the helix alignment in oriented membrane samples. Furthermore, we determined the antimicrobial and hemolytic properties of all peptides and performed vesicle leakage to assess their intrinsic toxicity toward model membranes of a given composition.
3. Discussion
In this work, we present a biophysical characterization of the full set of seven latarcin peptides in membranes, using CD, OCD, NMR, and vesicle leakage assay. Furthermore, we correlate these results with the antimicrobial and hemolytic activities of latarcins. These data are summarized in
Table 2. All latarcins used in this study were synthesized with a backbone
15N-label and purified before use. As described in the introduction, we divided the seven peptides into four groups based on their (genetically unrelated) sequences and properties. Here, we evaluate and discuss our results.
Similar to most membrane-active peptides, all latarcins remained essentially unfolded in aqueous phosphate buffer (
Supplementary Figure S2). However, in the presence of vesicles composed of DMPC/DMPG (7/3) at a P/L of 1/50, all of them were found to fold into the expected amphipathic α-helical structures, as illustrated in
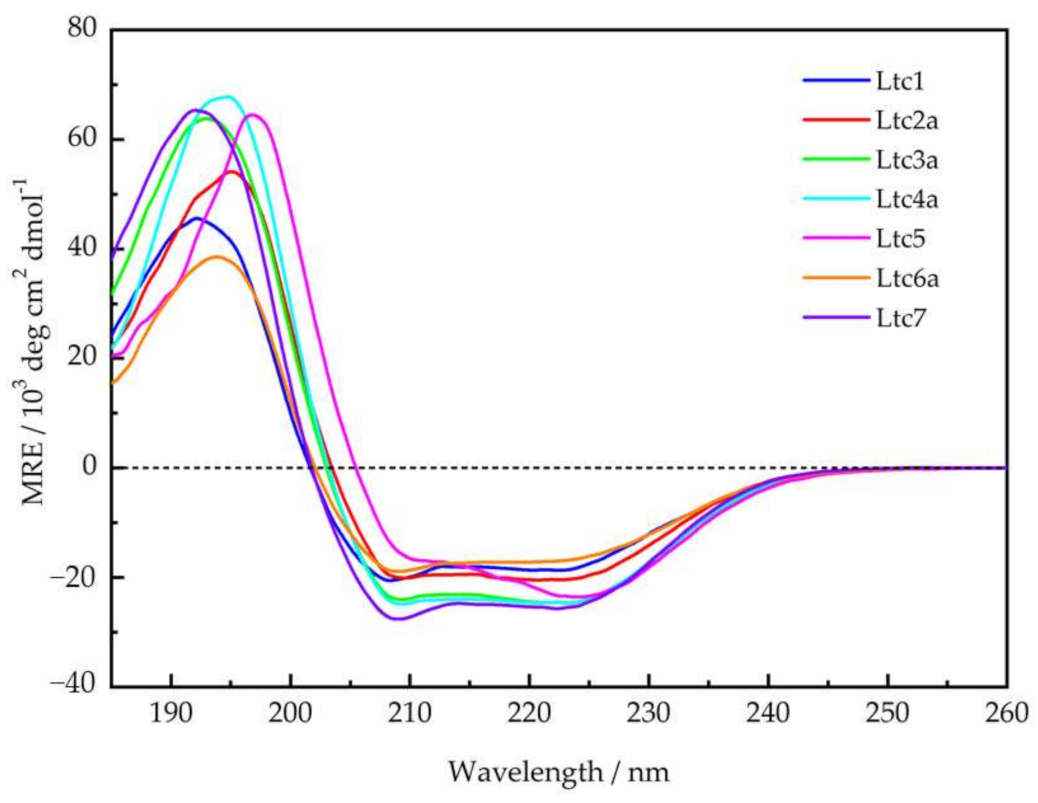
Figure 1. The CD line shapes (
Figure 2) were deconvoluted to estimate the secondary structure contributions (
Supplementary Table S1) using the CDSSTR, CONTIN-LL, and SELCON-3 algorithms provided on the DICHROWEB online server as described previously [
17]. In group A, Ltc1 and Ltc2a show nearly identical spectral features with bands centered around 208 nm and 224 nm except for the positive band, which shows some blueshift for Ltc1 towards lower wavelength with reduced intensity, indicating that some unordered part may be present in this peptide. This result is also reflected in the helix content, which is significantly reduced for Ltc1 (66%) compared to Ltc2a (81%).
Figure 1 shows that the two peptides have a similar hydrophobic sector, but Ltc2a has more distinct segregation between the polar and nonpolar residues, which favors better membrane binding and folding. In Ltc1, on the other hand, two charges (Lys10 and Lys21) are placed within the hydrophobic patch, as seen in the helical mesh representation. Group B peptides also have almost identical CD spectra and show the highest helix content of 87% to 94% (
Table 2). These two peptides have the most distinct segregation of polar and nonpolar residues in the folded form and are therefore expected to build ideal α-helices, as shown in
Figure 1. Group C (Ltc5) also shows similar hydrophobic and hydrophilic sectors, and this peptide is also expected to form an α-helix where most residues remain in the folded form. Despite the somewhat unusual line shape with a positive band shifted to the long wavelength and the rather intense negative band centered around 225 nm (in comparison to the band at 210 nm), Ltc5 indeed shows very high helicity (85%). The peculiar shift of the long-wavelength band can be attributed to the presence of a large number of aromatic residues (
Table 1) in Ltc5. The helical mesh presented in
Figure 1 shows that the hydrophobic sector is dominated by the presence of these aromatic residues. In peptides containing a high proportion of aromatic residues, such shifts of the long-wavelength band are well documented [
25,
26]. Group D peptides show rather large differences in their CD line shapes. Ltc6a has the lowest helicity (65%), whereas Ltc7 shows a pronounced line shape with a high helicity of 89% (
Table 2). Given the nearly identical length and number of hydrophobic residues, this appears to be a surprisingly large difference in the folded fraction for these two peptides. In comparison to Ltc6a, Ltc7 has a larger and better separation between hydrophobic and hydrophilic sectors. Ltc6a actually carries several bulky hydrophobic residues (Tyr15, Met18, and Leu33) in its polar sector, unlike Ltc7, where Ala31 may be regarded as a rather inconspicuous side chain. These residues may cause some unfolding or perturb the binding of Ltc6a, resulting in an overall lower helicity. To conclude, the CD experiments, performed under identical conditions, were important because: (a) they show for the first time that all latarcins are highly helical in the presence of a lipid membrane, and (b) the evidently helical state allows a subsequent determination of the helix tilt angle using oriented circular dichroism and solid-state NMR techniques.
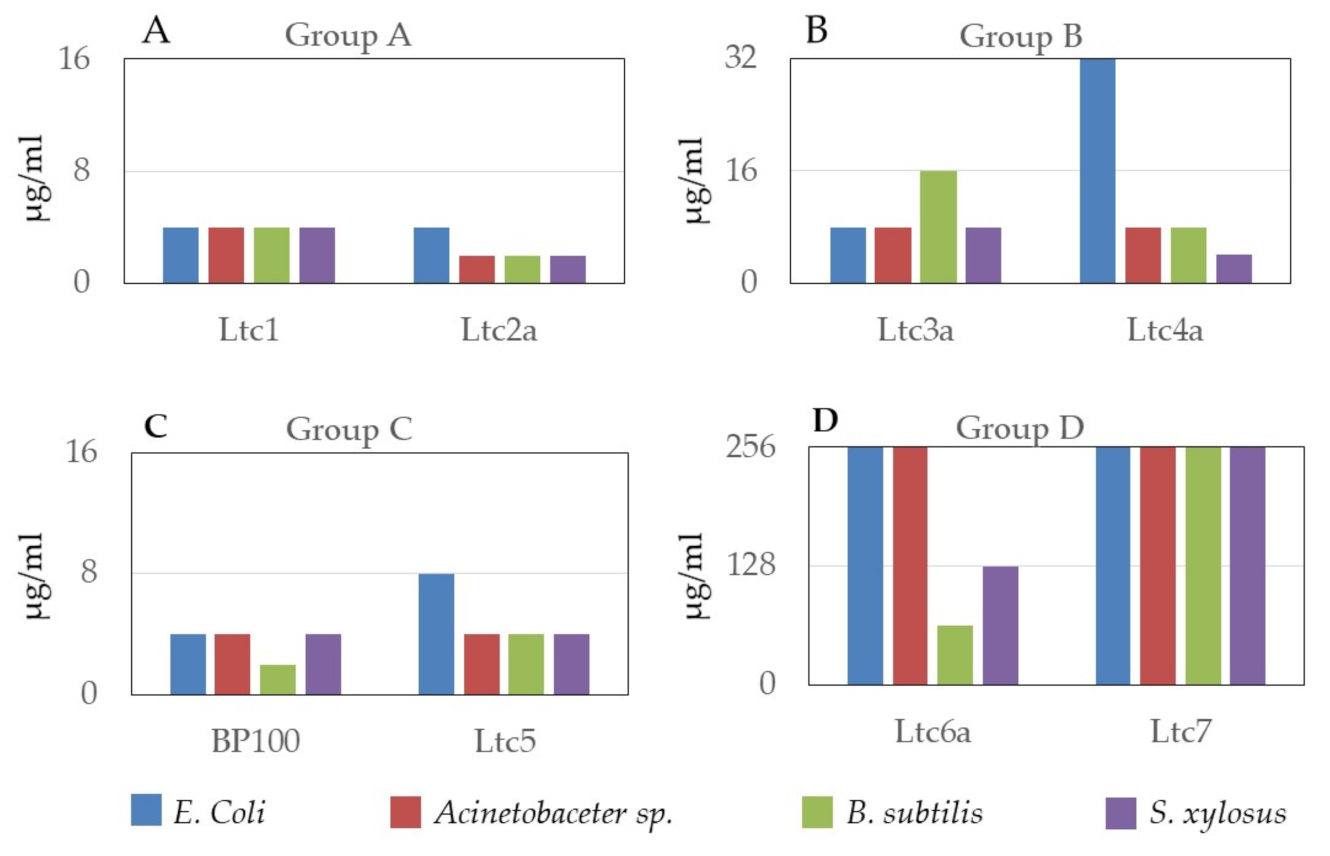
The antimicrobial and hemolytic activities of latarcins have been reported previously [
10]. In this work, we confirmed the antimicrobial action of latarcins on two Gram-positive and two Gram-negative bacterial strains. These results are presented in
Figure 3, together with BP100, a well-known multifunctional antimicrobial peptide [
27] that was used as a positive control. These data support the classification of latarcins into the suggested four groups. Ltc1 and Ltc2a, belonging to Group A, have the highest antimicrobial activity and closely resemble Ltc5 from Group C. This result may be expected due to the high charge per residue of these peptides (
Table 1). Here, the MIC lies universally below 4 µg/mL irrespective of the bacterial strain, demonstrating their broad spectrum and rather indiscriminatory mechanism of bacterial killing. Next, only marginally less active are the Group B peptides comprising of Ltc3a and Ltc4a, where antimicrobial activity is observed in the range of 8–32 µg/mL. Similar to Groups A and C, Group B is also universally active on all strains. The peptides in Group D are less active. Here, Ltc6a inhibits the growth of the Gram-positive strains only, whereas Ltc7 does not inhibit bacterial growth up to a peptide concentration of 256 µg/mL. Although Ltc7 was found to be ineffective against the selected strains that have been tested here, it is conceivable that it may be active against other bacterial strains found in the habitat of the spider
L. tarabaevi or that it may have some activity against other microorganisms or fungi.
Hemolysis assays were performed on human erythrocytes, in contrast to earlier assays performed on rabbit erythrocytes in the literature [
7].
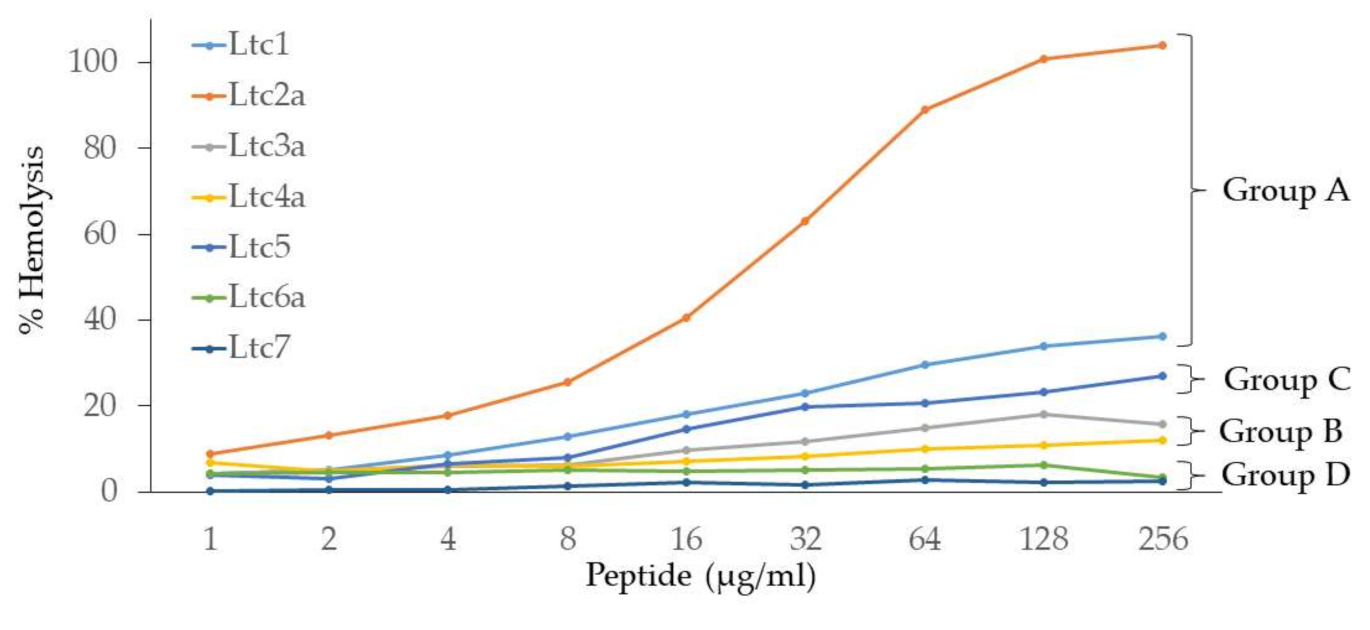
Figure 4 shows that the hemolytic action of latarcins varies widely. The Group A peptides (Ltc1 and Ltc2a) have the highest hemolytic activity, with Ltc2a being the most hemolytic of all. They are followed by the Group C peptide (Ltc5), which is less hemolytic than Group A peptides. Group B (Ltc3a and Ltc4a) shows rather low hemolysis, which is noteworthy given their high antimicrobial action. Finally, Group D (Ltc6a and Ltc7) showed no or marginal hemolysis. The order of the hemolytic activity of the seven latarcins thus follows the same order as of the antimicrobial activity (
Table 2). This finding suggests that the peptides have an indiscriminatory effect on the lipid membranes of bacteria or erythrocytes. Apparently, those peptides that are able to kill bacteria are also able to induce hemolysis of human erythrocytes, despite their fundamentally different lipid compositions, electrostatic properties, and transmembrane potentials.
We may, at this point, speculate about two hypothetical conclusions regarding the biological membrane activity of the latarcin peptides. The first observation is that latarcins do not seem to be able to discriminate between different target membranes, given the same relative activities against different types of biomembranes, i.e., of prokaryotic and eukaryotic cells. This unspecific target selection of latarcins appears quite plausible, considering the venom’s purpose to disinfect the venom gland, paralyze and sterilize the prey, and inflict pain and deter predators. The second hypothesis is that the peptides themselves have encoded distinctly different degrees of activity and may therefore display functional specificity toward lipid membranes in general. This means, for example, that Group D peptides (Ltc6a and Ltc7) do not show much activity in bursting (bacterial or erythrocyte) membranes but may display some rather different kinds of membrane activity, such as cell penetration or others (which have NOT been tested here). At the same time, peptides in Group A were found to be highly active against bacteria and erythrocytes, but they may not show a good ability for more subtle membrane interactions.
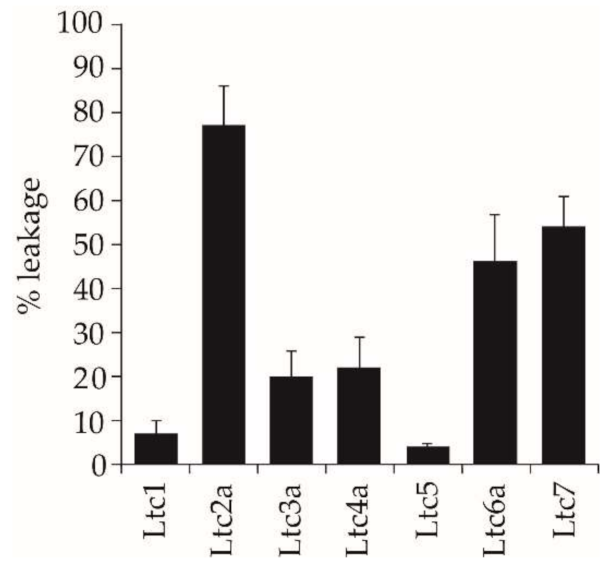
One way to test these hypotheses is to study the effects of the peptides against well-defined model membrane systems using a fluorescence leakage assay, which shows whether the peptides can directly induce rapid membrane permeabilization. Here, we used POPC/POPG (1/1) bilayers with a high proportion of anionic lipids. We found that our results support the concept of functional specificity. Ltc2a is extremely effective at inducing leakage in POPC/POPG vesicles, making this peptide an overall “membrane buster” that has an indiscriminatory response to bacterial membranes, eukaryotic membranes, and vesicles composed of synthetic lipids. The other Group A peptide, Ltc1, shows very low fluorescent leakage despite having similar properties as Ltc2a, and the same is true for Group C (Ltc5). It may have been expected that the high positive charge/residue ratio of these two peptides should provide sufficient electrostatic attraction to anionic vesicles and thus induce a high extent of leakage. Instead, we found that they induced the least leakage in vesicles made from POPC/POPG. From the CD and NMR results, it is clear that these peptides bind well but do not aggregate or perturb the bilayer, yet they are antimicrobial (
Table 2). Taken together, these results suggest that the two peptides, Ltc1 and Ltc5, might have intracellular targets in bacteria and can penetrate the lipid bilayer without much physical damage. The two Group B peptides (Ltc3a and Ltc4a) show a relatively low extent of leakage (20%), comparable to their low hemolysis, with moderate antimicrobial activity. Group D peptides (Ltc6a and Ltc7), on the other hand, show either low or no antimicrobial (or hemolytic) activity but induce considerable vesicle leakage of 50% (
Table 2). This means that they show specific action against the PC/PG model membranes but not against bacteria and erythrocytes. This finding suggests that the last two peptides, which showed no activity in our biological assays, may have other activities that are relevant for the spider.
Another possibility that we investigated here was the question of whether there could be any synergistic effects between the peptides. It is conceivable that some peptides do not show any impressive activity per se but turn out to be highly active in combination with another peptide partner. Such synergy is readily tested in a leakage assay because much smaller amounts of peptides are needed compared to antimicrobial and hemolytic experiments, and many samples can be prepared and measured effectively. All pairwise combinations of the seven latarcins were tested, but no synergy was found for any pair.
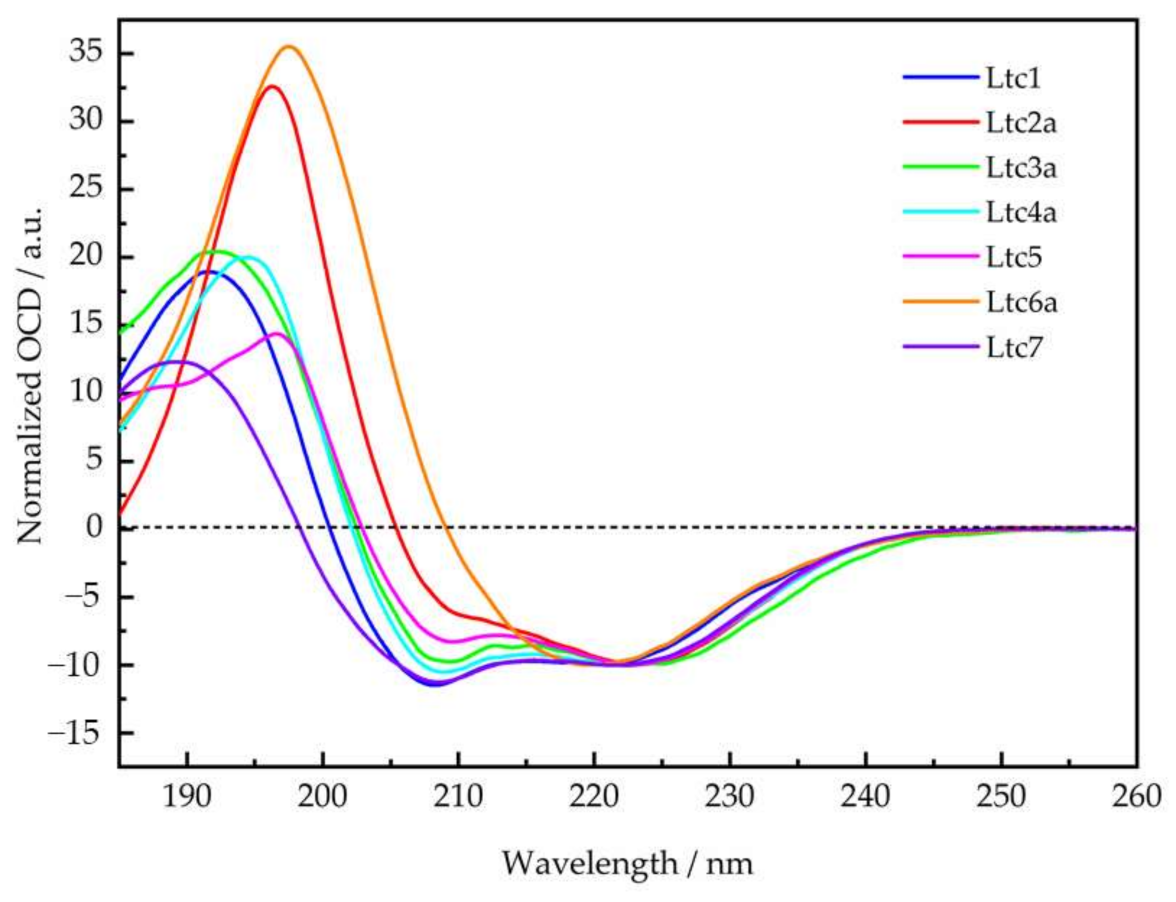
To obtain some molecular impressions of the underlying peptide-lipid interactions, the membrane alignment of the latarcins was examined using two complementary techniques, namely OCD and solid-state
15N-NMR. Both of these methods use identical sample preparation, where lipids are oriented on glass surfaces and well hydrated but without any excess water. Under these conditions, the peptides remain fully bound to the membrane and do not float away into regions of bulk water. The orientation of the α-helical peptides can be determined using these two methods, as seen in
Figure 6 and
Figure 7. OCD clearly shows that within Group A, the helix of Ltc1 is found to lie flat on the membrane surface. Ltc2a, on the other hand, is the most tilted peptide of the entire series. Interestingly, Ltc2a also has the highest membrane permeabilizing activity and the least selective effect on bilayers of different compositions. Group B peptides (Ltc3a and Ltc4a) show similar spectral profiles, indicating that both are surface-bound or very slightly tilted in the membrane. Ltc5 (Group C) and Ltc 7 in Group D were also found to lay flat on the membrane surface in the OCD experiments. Our results clearly indicate that when peptides bind to the membrane, they may not fold completely, even though they seemed to fold from the CD line shapes in TFE or in the presence of bulk water; thus, OCD is a unique way of determining the fold change of the peptides in the membranous environment. Such structural transitions have been observed previously, where otherwise helical peptides tend to unfold to aggregate as β-sheets in the 2D plane of the membrane [
28,
29]. In fact, Ltc6a loses its helical fold and aggregates in the oriented membrane samples prepared for OCD and
15N-NMR analysis; therefore, one cannot determine its alignment within the membrane.
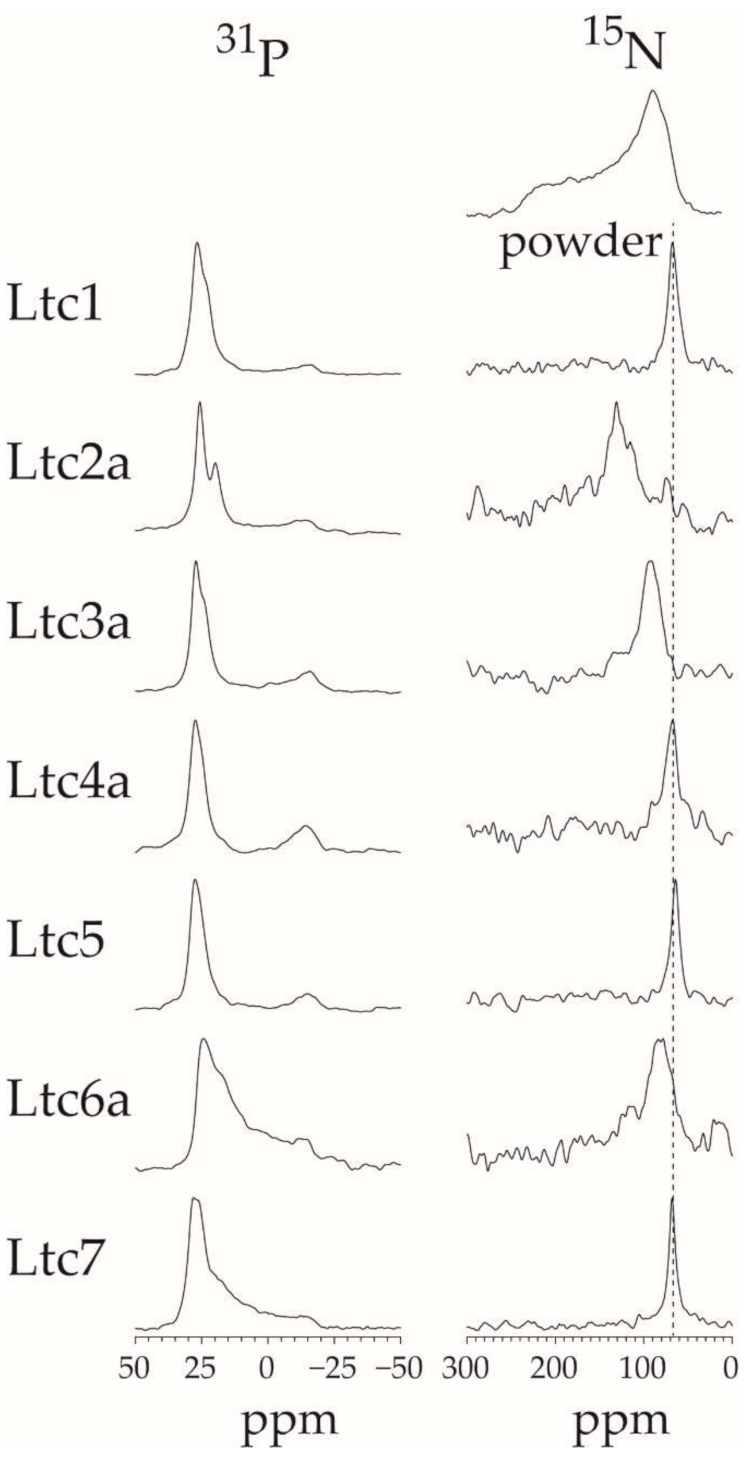
The solid-state
15N-NMR results (
Figure 7) nicely confirm the peptide orientation observed by OCD (
Table 2). In these experiments, Ltc2a was found to be the most tilted helix in the membrane, with a signal around 130 ppm. Ltc3a is tilted slightly in the membrane, with a peak around 93 ppm, in line with our OCD results. All other latarcins show a
15N-NMR signal around 64–68 ppm, indicating that these helices are surface-bound. (Additionally, Ltc5 shows a signal clearly indicative of a surface-bound state, which confirms our interpretation of the OCD data above, which had been challenged by the presence of numerous aromatic residues.)
The
31P-NMR data show that most peptides do not perturb the lamellar bilayer order. The Group D peptides are an exception in producing a high mosaic spread (i.e., orientational disorder) of the lipids, and indeed they also show high vesicle leakage. Perhaps the most striking observation here is the fact that Ltc2a is the only peptide to induce lipid segregation: besides the usual signal at 26 ppm, a second
31P-NMR peak is seen to have emerged around 21 ppm in
Figure 7. Such additional signal is sometimes observed in
31P-NMR of cationic amphiphilic peptides in a DMPC/DMPG matrix [
30,
31], especially in cases where lipid clustering has been demonstrated [
32]. We, therefore, attribute the
31P-NMR signature of Ltc2a to its ability to induce lipid segregation due to its high charge. Furthermore, a significant upfield shift of a
31P-NMR signal would be expected if the lipids are motionally averaged by diffusing through a highly curved environment. The latter effect would indeed be plausible if Ltc2a were to cause a dimple in the bilayer due to its high tilt angle of 30–40° away from the membrane surface according to
15N-NMR.
Compared to the other latarcins, it is likely that the very high charge density in Ltc2a (11 Lys amongst 26 amino acids, see
Table 1) and its pronounced amphiphilicity (11 hydrophobic residues) contribute to its particularly disruptive membrane interactions. The most unusual feature of Ltc2a, in our opinion, is the strip of hydrophobic amino acids that does not run in parallel along but rather diagonally around the helix axis, as seen in the helical mesh representation of
Figure 1. This amphiphilic profile is closely reminiscent of a coiled-coil motif—a pattern that is typically known to drive the assembly of two (or more) α-helices in an aqueous solution. In the case of Ltc2a, however, we suggest that this particular structural element is instead critically important in modulating its preferred mode of membrane binding. Given the 180° angle by which the hydrophobic strip is wrapped around the helix, the fully folded Ltc2a molecule is expected to be quite unable to bind straight onto the flat membrane surface, unlike most other known amphiphilic peptides. Instead, the Ltc2a helix will be forced to insert more steeply into and possibly across the lipid bilayer in a slanted manner in order to optimize its hydrophobic contacts with the lipids, as was indeed observed here by OCD and NMR (
Figure 6 and
Figure 7). This unusual kind of coiled-coil amphiphilicity is described and interpreted here for the first time with regard to a membrane-active peptide. It should be noted that Ltc1 possesses a similar coiled-coil pattern at first sight (see
Figure 1), but its hydrophobic strip is interrupted by two cationic residues (Lys10, Arg15). Therefore, it is much easier for Ltc1 to adjust to the flat membrane surface by kinking. Indeed, the helicity of Ltc1 is found to be lower than for Ltc2a in the membrane-bound state, according to CD (
Figure 2). The other members of the latarcin family display no coiled-coil features, as their amphiphilic profiles run parallel to the helix axis in all cases. With its peculiar and well-defined coiled-coil motif and its resultant vigorous membrane interactions, Ltc2a stands out distinctly from Ltc1 and the other latarcins and also in all of the functional assays that were studied here. Ltc2a is found to be the most aggressive and least selective membrane-permeabilizing agent, with an extremely high antimicrobial, hemolytic, and leakage-inducing activity.
In summary, our results show that the amphiphilic latarcin helices lie on the membrane surface or can tilt slightly into the lipid bilayer. It is tempting to speculate that, mechanistically, the entire group of seven peptides covers a wide spectrum of activities and specificities. Some act via lipid clustering and massive permeabilization (Ltc2a) or by disordering the lamellar bilayer (Ltc6a, Ltc7), while others might be able to translocate non-perturbingly towards intracellular targets (Ltc1, Ltc5). Some are highly antibiotic but show low hemolytic effects (Ltc3a, Ltc4a, Ltc5a), while others destroy membranes indiscriminately (Ltc2a), and yet, others are inefficient antibiotics but permeabilize lipid bilayers (Ltc6a, Ltc7). Given the absence of any pairwise synergy, these diverse features highlight the multiple purposes of spider venom.


 ,
, 







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}