
Endothelial Colony-Forming Cells Dysfunctions Are Associated with Arterial Hypertension in a Rat Model of Intrauterine Growth Restriction

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Results
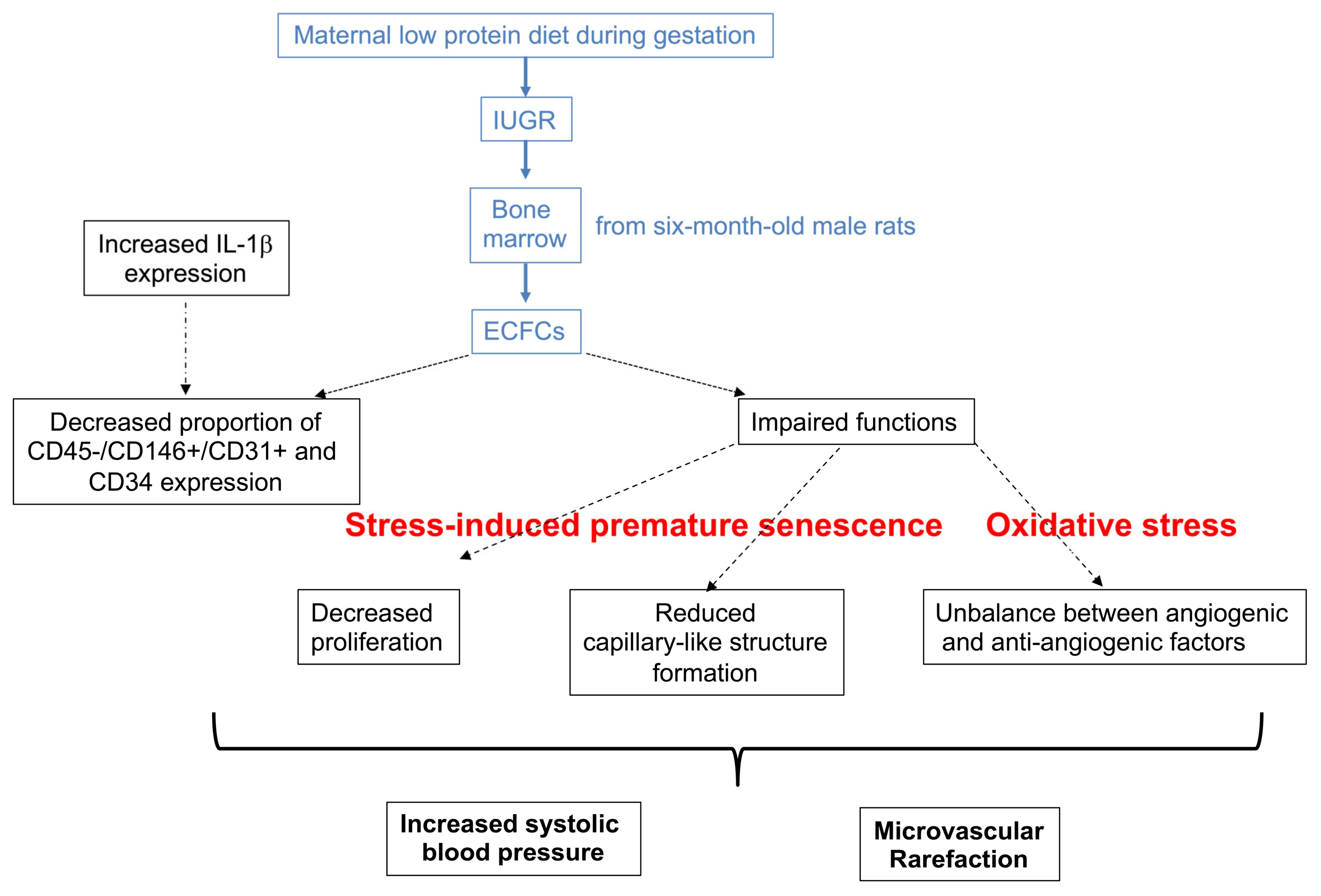
2.1. IUGR-Induced Lower Body Weight at Birth and at Six Months of Life
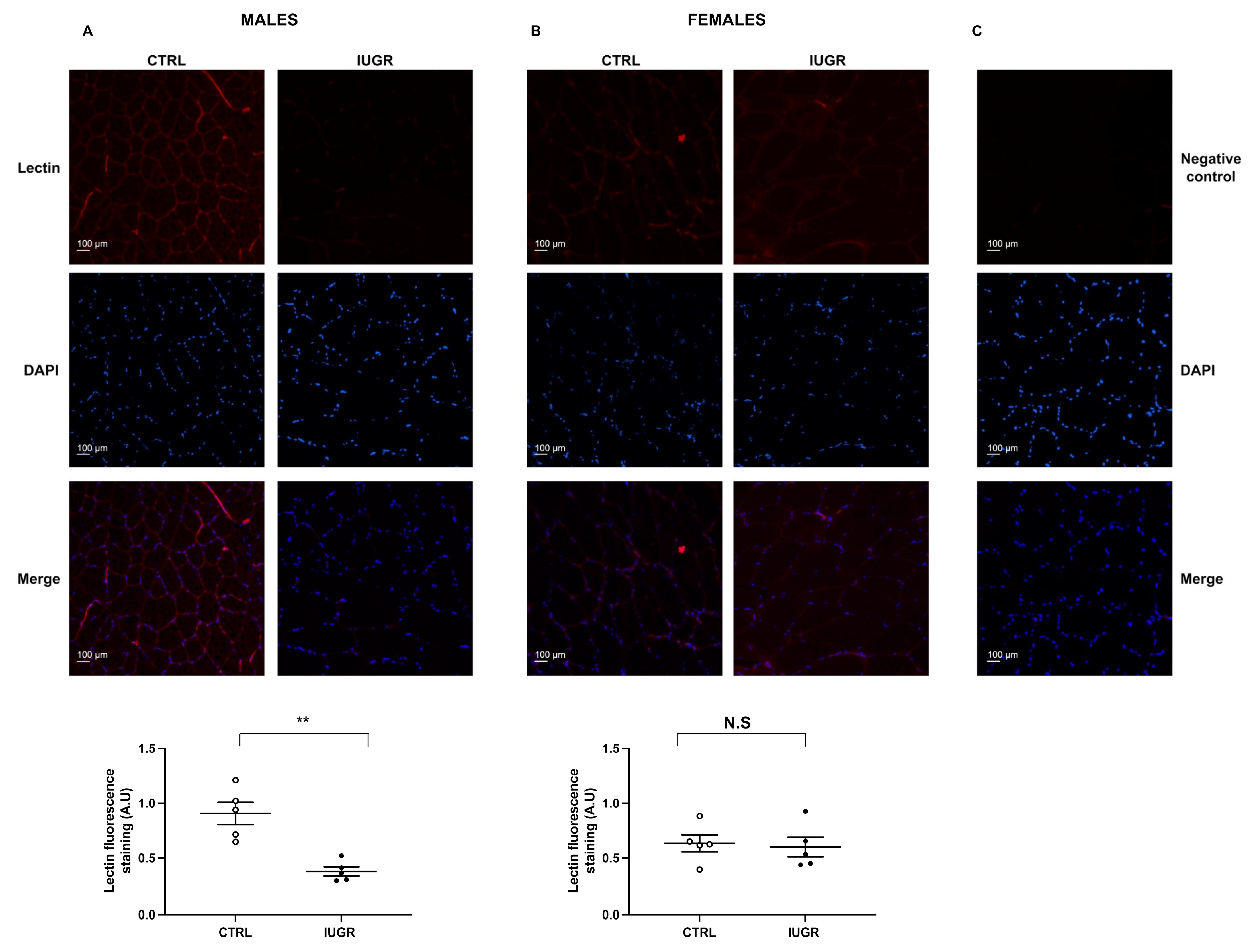
2.2. IUGR-Induced Increased Systolic Blood Pressure and Microvascular Rarefaction
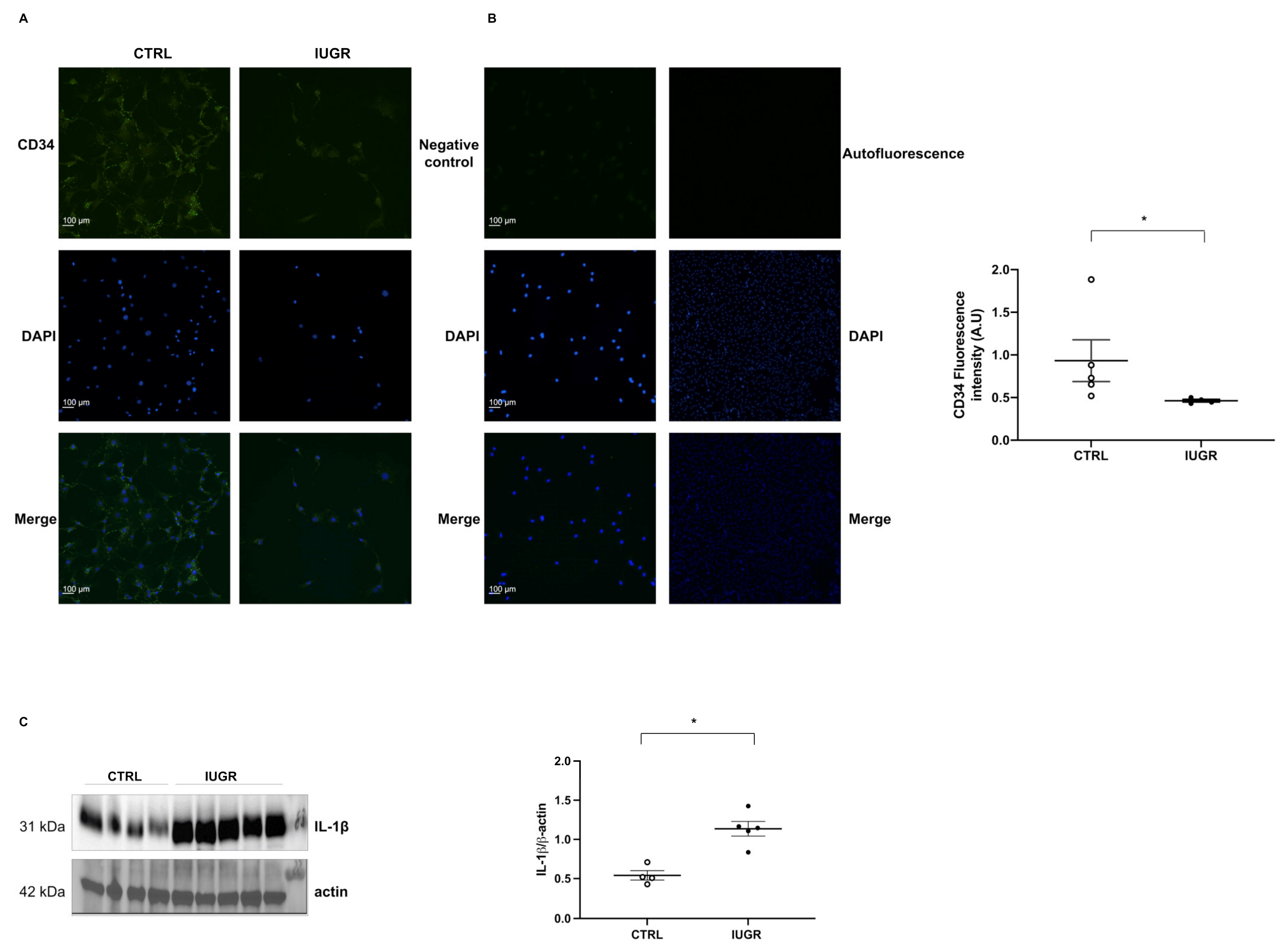
2.3. Decreased Number of IUGR-ECFCs
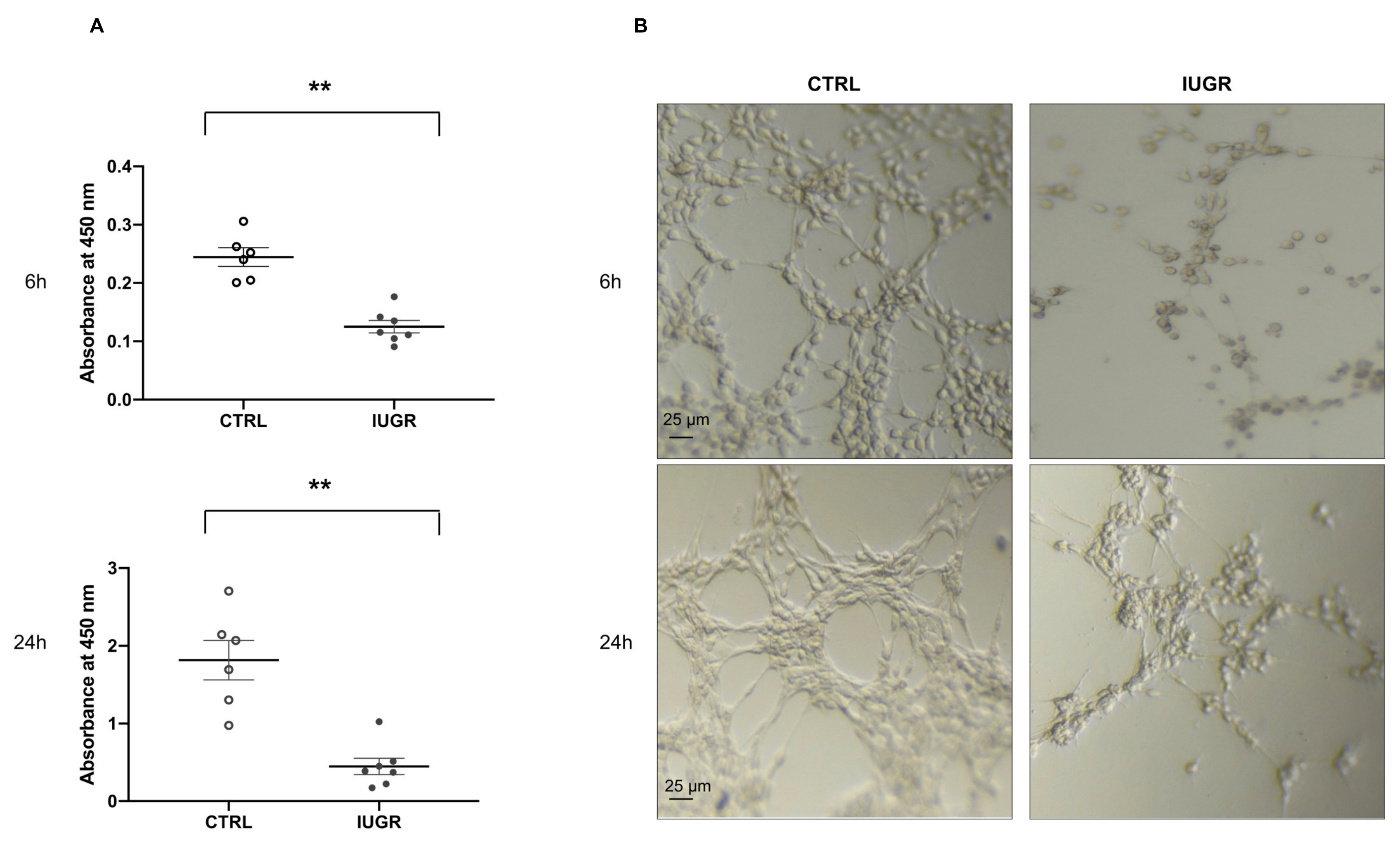
2.4. Altered Proliferation and Capillary-like Outgrowth Sprout Properties
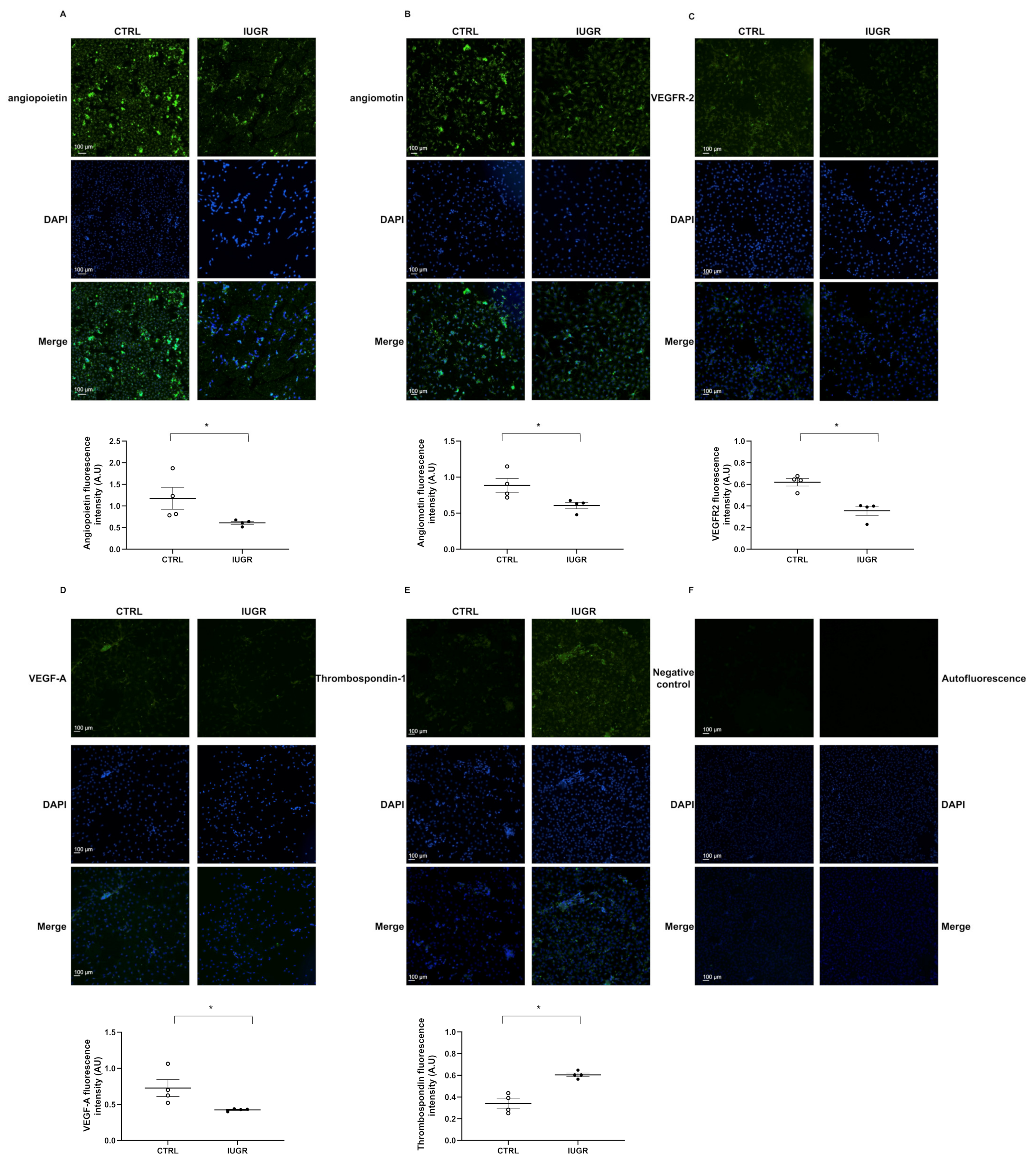
2.5. Impaired Angiogenic Capacity
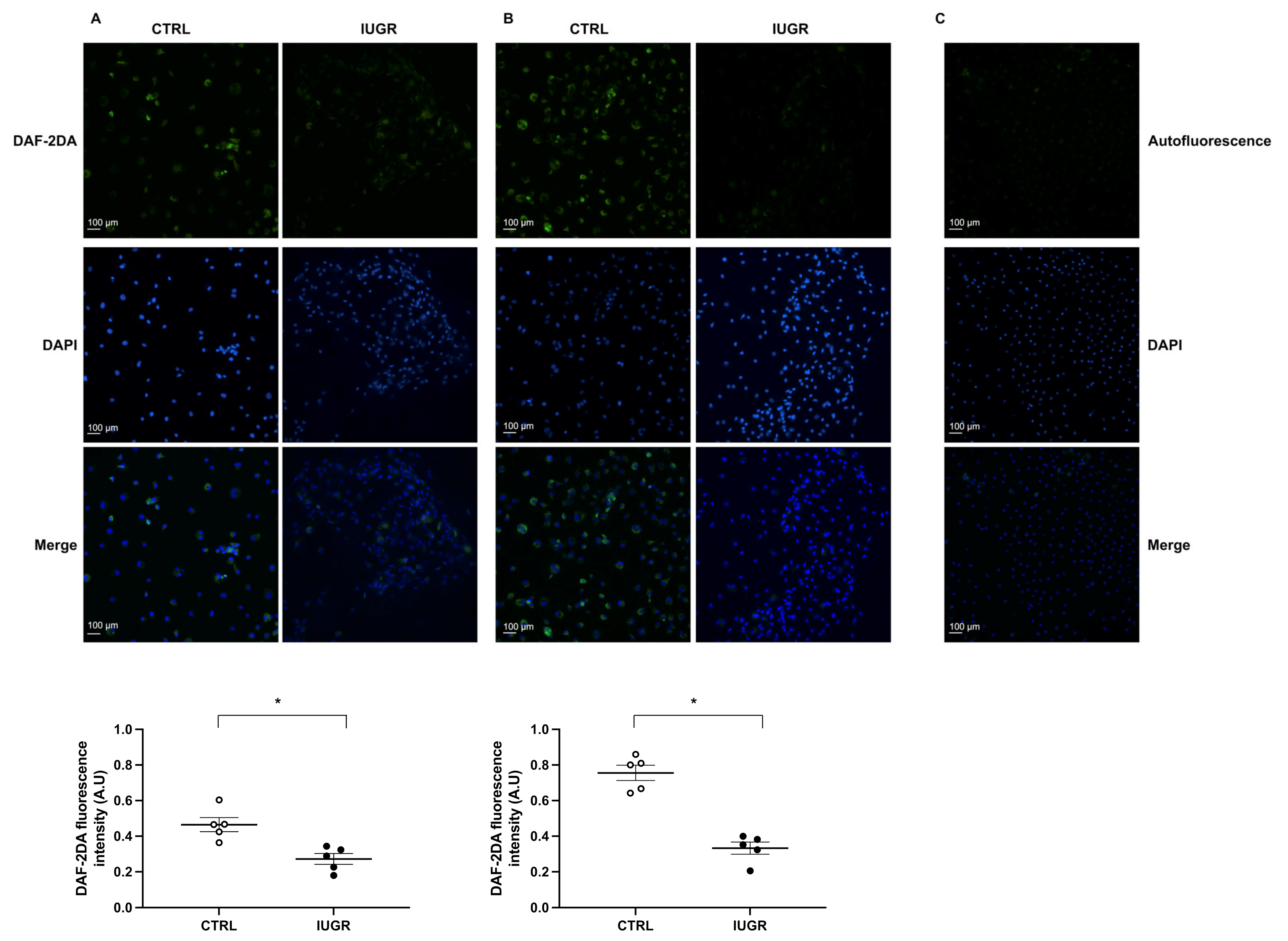
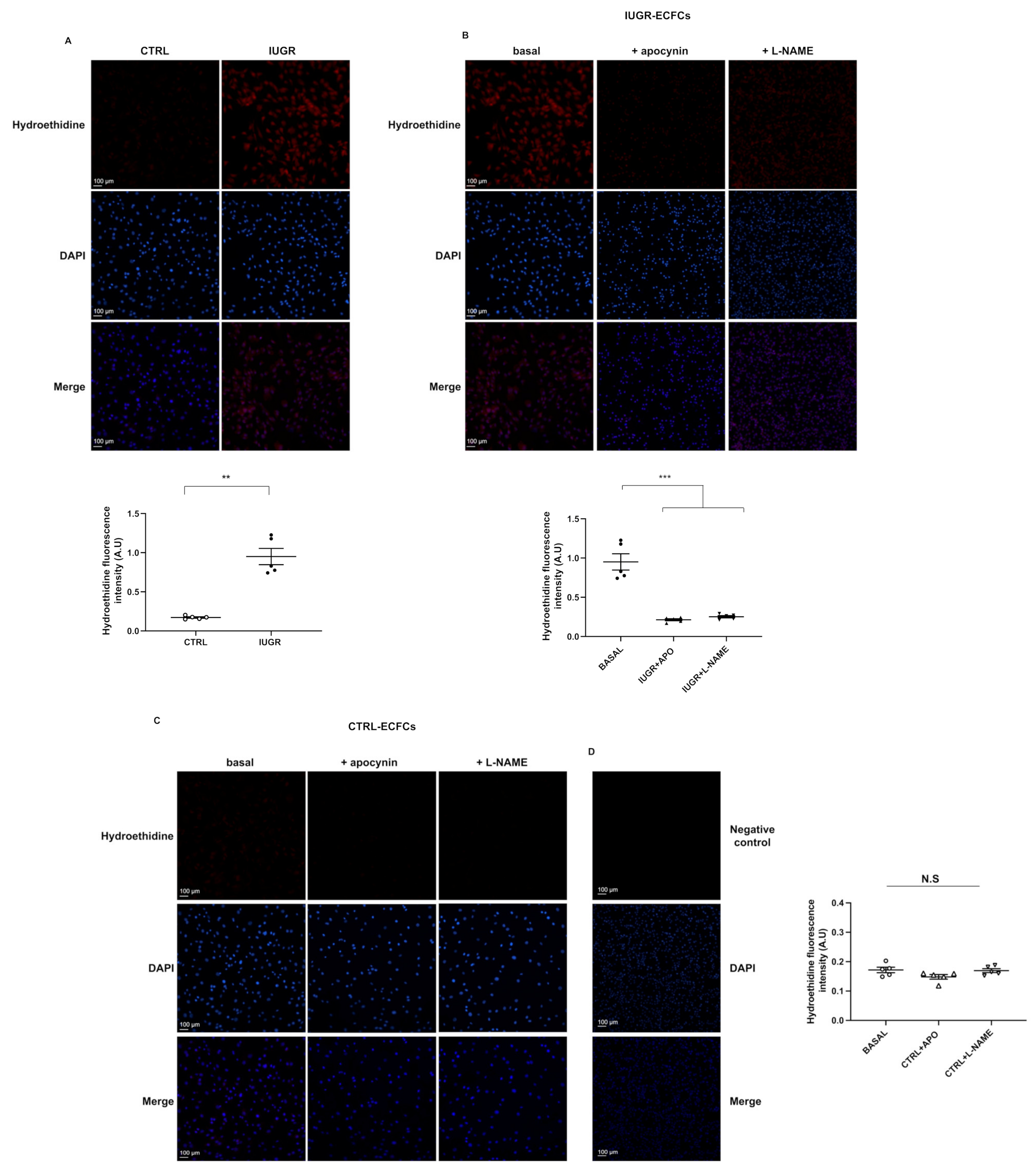
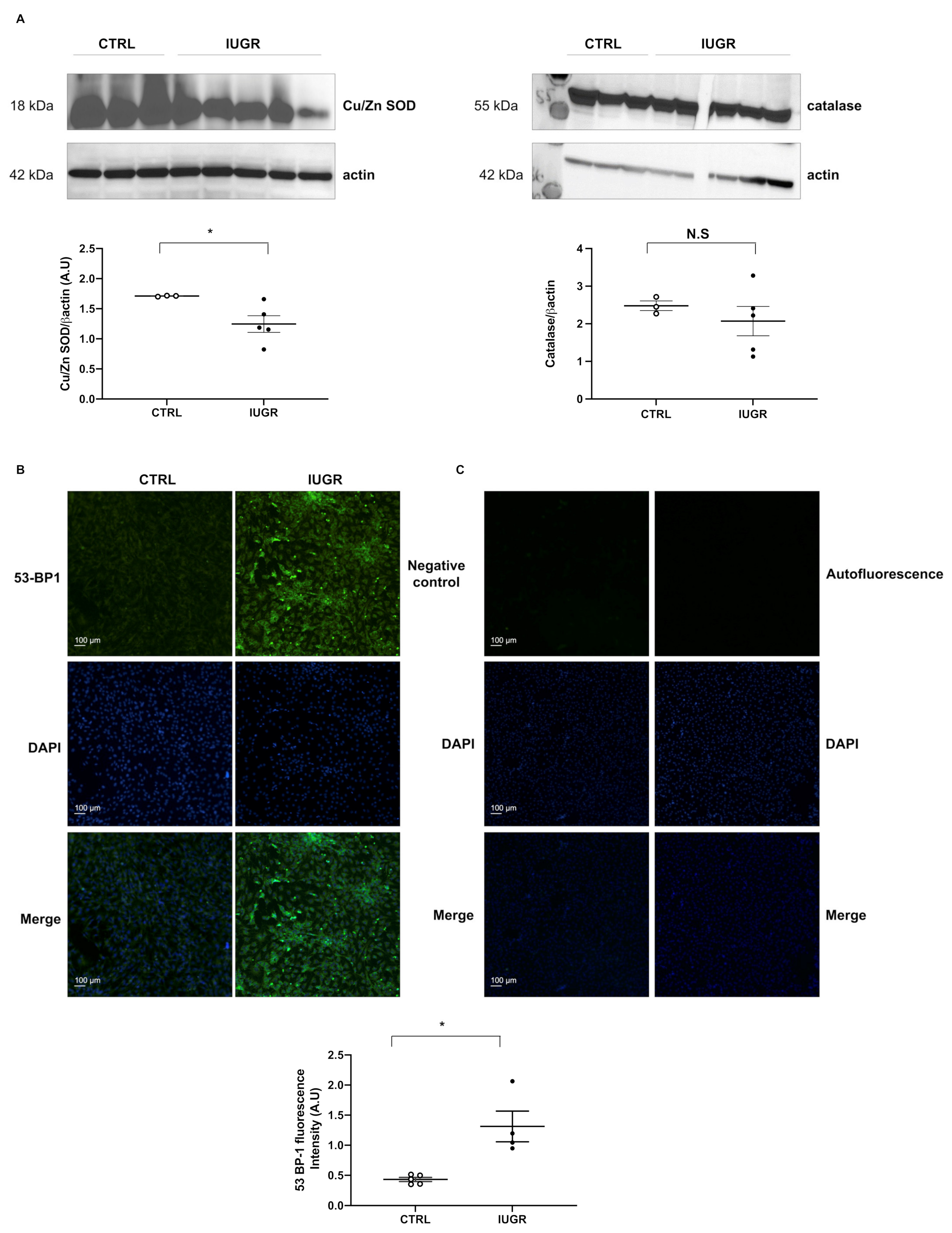
2.6. Oxidative Stress
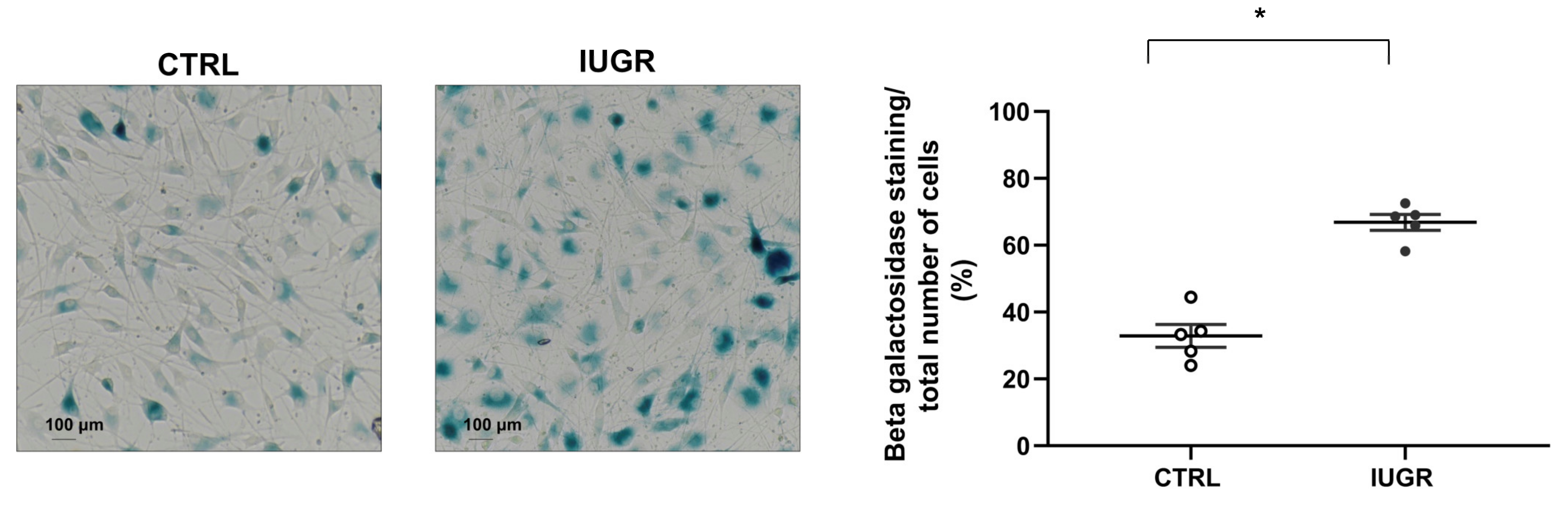
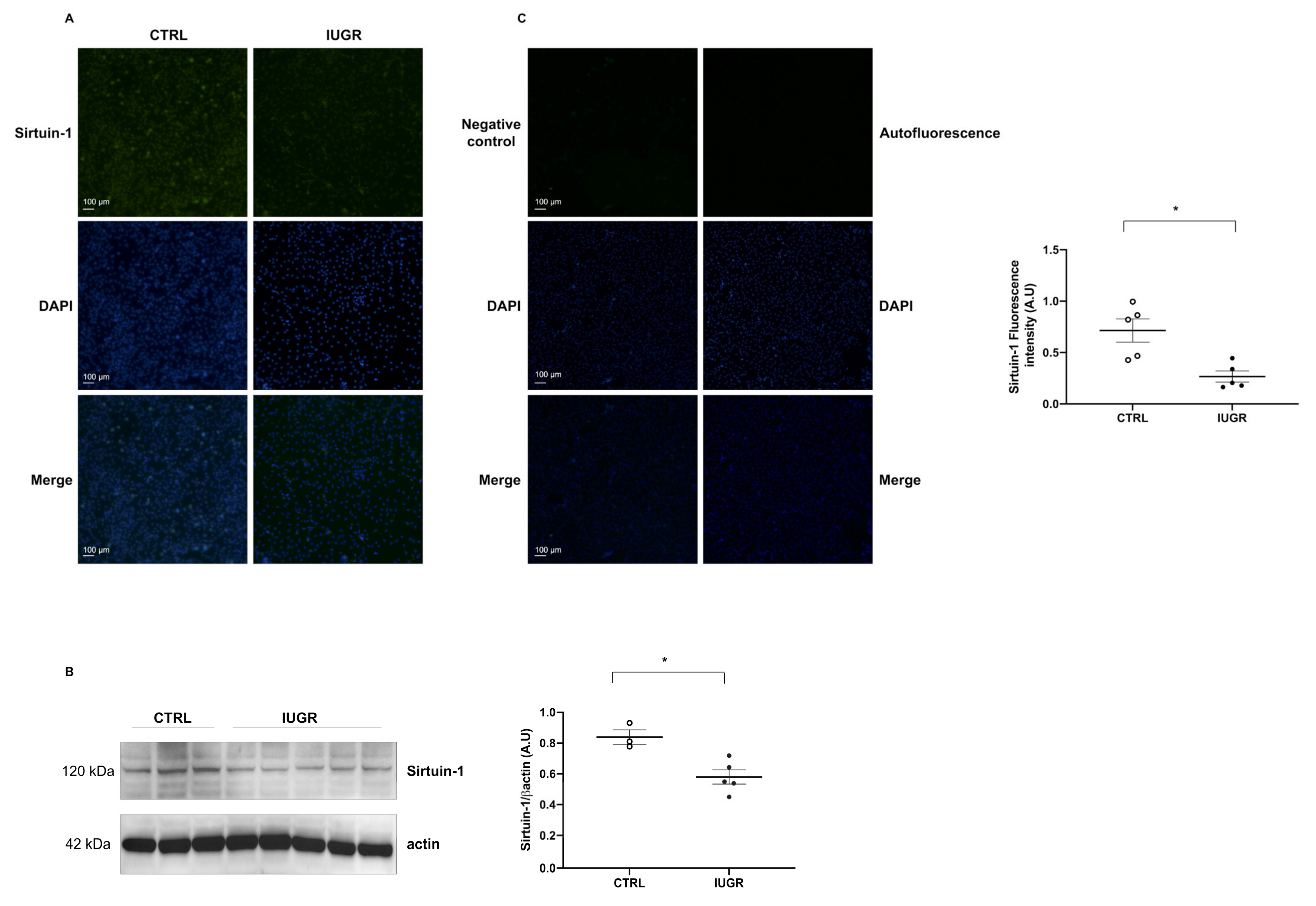
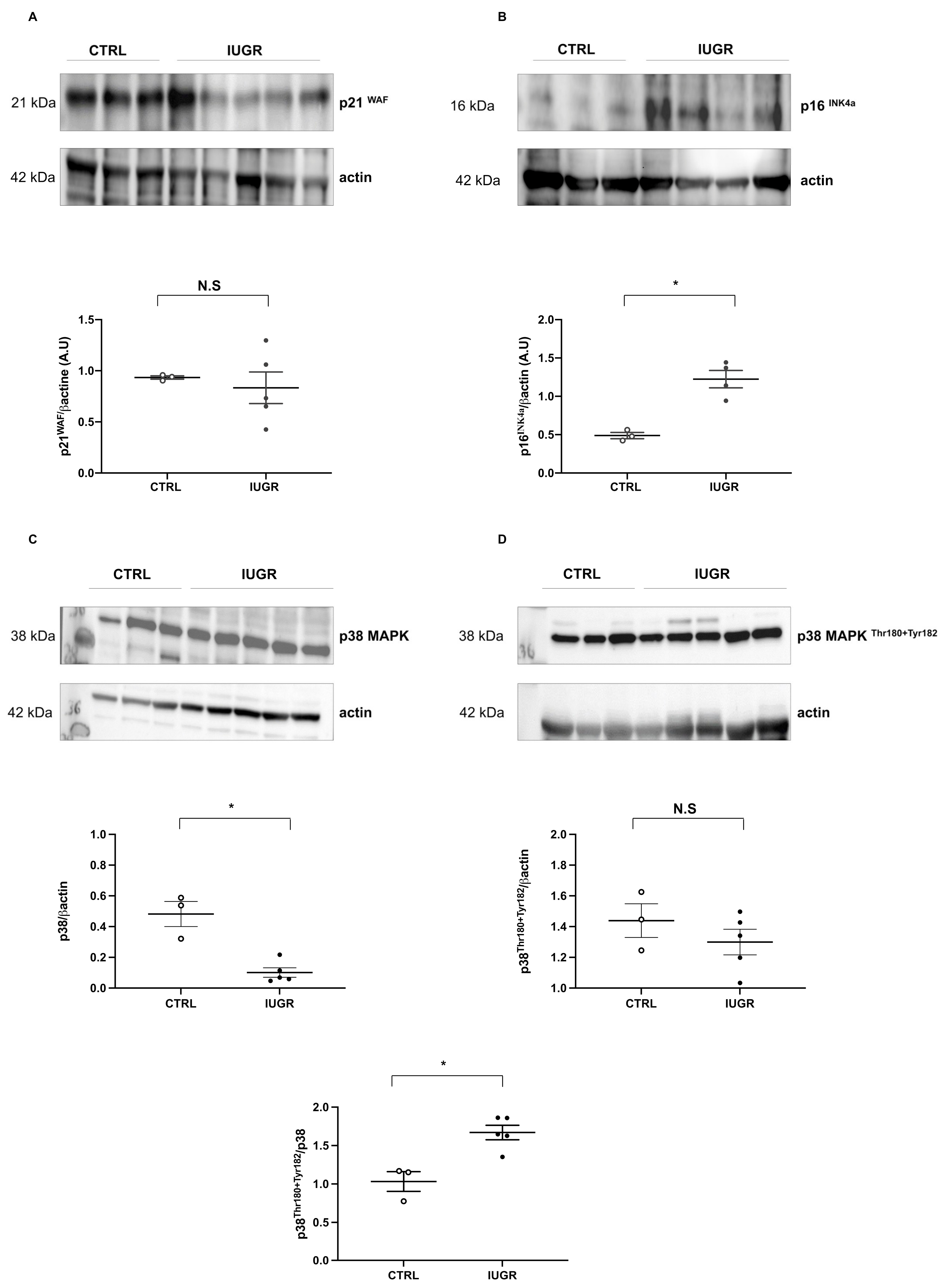
2.7. Cellular Senescence
3. Discussion
4. Materials and Methods
4.1. Body Weight Measurement
4.2. Animal Model
4.3. Systolic Blood Pressure Measurement
4.4. Microvascular Density Measurement
4.5. Endothelial Progenitor Cell Isolation
4.6. ECFC Quantification Using Flow Cytometry
4.7. ECFC Proliferation Test
4.8. ECFC Capillary-like Structure Formation
4.9. Measurement of NO Production by ECFCs
4.10. Measurement of Superoxide Anion Production by ECFCs
4.11. Senescence Detection in ECFCs
4.12. Immunofluorescence
4.13. Protein Expression Evaluation Using Western Blotting
4.14. Statistical Analyses
5. Conclusions
5.1. Conclusion
5.2. Limitations
5.3. Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fattal-Valevski, A.; Bernheim, J.; Leitner, Y.; Redianu, B.; Bassan, H.; Harel, S. Blood pressure values in children with intrauterine growth retardation. Isr. Med. Assoc. J. 2001, 3, 805–808. [Google Scholar]
- Rossi, P.; Tauzin, L.; Marchand, E.; Boussuges, A.; Gaudart, J.; Frances, Y. Respective roles of preterm birth and fetal growth restriction in blood pressure and arterial stiffness in adolescence. J. Adolesc. Health 2011, 48, 520–522. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, P.M.; Ostergren, P.O.; Nyberg, P.; Soderstrom, M.; Allebeck, P. Low birth weight is associated with elevated systolic blood pressure in adolescence: A prospective study of a birth cohort of 149378 Swedish boys. J. Hypertens. 1997, 15, 1627–1631. [Google Scholar] [CrossRef] [PubMed]
- Leon, D.A.; Johansson, M.; Rasmussen, F. Gestational age and growth rate of fetal mass are inversely associated with systolic blood pressure in young adults: An epidemiologic study of 165,136 Swedish men aged 18 years. Am. J. Epidemiol. 2000, 152, 8. [Google Scholar] [CrossRef]
- Curhan, G.C.; Chertow, G.M.; Willett, W.C.; Spiegelman, D.; Colditz, G.A.; Manson, J.E.; Speizer, F.E.; Stampfer, M.J. Birth weight and adult hypertension and obesity in women. Circulation 1996, 94, 1310–1315. [Google Scholar] [CrossRef]
- Law, C.M.; Shiell, A.W. Is blood pressure inversely related to birth weight? The strength of evidence from a systematic review of the literature. J. Hypertens. 1996, 14, 935–941. [Google Scholar] [CrossRef] [PubMed]
- Martyn, C.N.; Barker, D.J.; Jespersen, S.; Greenwald, S.; Osmond, C.; Berry, C. Growth in utero, adult blood pressure, and arterial compliance. Br. Heart J. 1995, 73, 6. [Google Scholar] [CrossRef] [Green Version]
- Yzydorczyk, C.; Armengaud, J.B.; Peyter, A.-C.; Chehade, H.; Cachat, F.; Juvet, C.; Siddeek, B.; Simoncini, S.; Sabatier, F.; Dignat-George, F.; et al. Endothelial dysfunction in individuals born after fetal growth restriction: Cardiovascular and renal consequences and preventive approaches. J. Dev. Orig. Health Dis. 2017, 8, 448–464. [Google Scholar] [CrossRef]
- Leeson, C.; Whincup, P.; Cook, D.; Donald, A.; Papacosta, O.; Lucas, A.; Deanfield, J. Flow-mediated dilation in 9-to 11-year-old children: The influence of intrauterine and childhood factors. Circulation 1997, 96, 2233–2238. [Google Scholar] [CrossRef]
- Martin, H.; Gazelius, B.; Norman, M. Impaired acetylcholine-induced vascular relaxation in low birth weight infants: Implications for adult hypertension? Pediatr. Res. 2000, 47, 457–462. [Google Scholar] [CrossRef] [Green Version]
- Martyn, C.N.; Greenwald, S.E. Impaired synthesis of elastin in walls of aorta and large conduit arteries during early development as an initiating event in pathogenesis of systemic hypertension. Lancet 1997, 350, 953–955. [Google Scholar] [CrossRef]
- Joo Turoni, C.; Chaila, Z.; Chahla, R.; Bazan de Casella, M.C.; Peral de Bruno, M. Vascular Function in Children with Low Birthweight and Its Relationship with Early Markers of Cardiovascular Risk. Horm. Res. Paediatr. 2016, 85, 396–405. [Google Scholar] [CrossRef]
- Medina, R.J.; Barber, C.L.; Sabatier, F.; Dignat-George, F.; Melero-Martin, J.M.; Khosrotehrani, K.; Ohneda, O.; Randi, A.M.; Chan, J.K.Y.; Yamaguchi, T.; et al. Endothelial Progenitors: A Consensus Statement on Nomenclature. Stem. Cells Transl. Med. 2017, 6, 1316–1320. [Google Scholar] [CrossRef] [PubMed]
- Mund, J.A.; Estes, M.L.; Yoder, M.C.; Ingram, D.A., Jr.; Case, J. Flow cytometric identification and functional characterization of immature and mature circulating endothelial cells. Arterioscler. Thromb. Vasc. Biol. 2012, 32, 1045–1053. [Google Scholar] [CrossRef] [Green Version]
- Estes, M.L.; Mund, J.A.; Ingram, D.A.; Case, J. Identification of Endothelial Cells and Progenitor Cell Subsets in Human Peripheral Blood. Curr. Protoc. Cytom. 2010, 52, 9.33.1–9.33.11. [Google Scholar] [CrossRef]
- Asahara, T.; Kawamoto, A.; Masuda, H. Concise review: Circulating endothelial progenitor cells for vascular medicine. Stem. Cells 2011, 29, 1650–1655. [Google Scholar] [CrossRef] [PubMed]
- Werner, N.; Wassmann, S.; Ahlers, P.; Schiegl, T.; Kosiol, S.; Link, A.; Walenta, K.; Nickenig, G. Endothelial progenitor cells correlate with endothelial function in patients with coronary artery disease. Basic Res. Cardiol. 2007, 102, 565–571. [Google Scholar] [CrossRef]
- Souza, L.V.; De Meneck, F.; Oliveira, V.; Higa, E.M.; Akamine, E.H.; Franco, M.D.C. Detrimental Impact of Low Birth Weight on Circulating Number and Functional Capacity of Endothelial Progenitor Cells in Healthy Children: Role of Angiogenic Factors. J. Pediatr. 2019, 206, 72–77.e1. [Google Scholar] [CrossRef]
- Kang, M.; Thebaud, B. Stem cell biology and regenerative medicine for neonatal lung diseases. Pediatr. Res. 2018, 83, 291–297. [Google Scholar] [CrossRef]
- Ligi, I.; Simoncini, S.; Tellier, E.; Grandvuillemin, I.; Marcelli, M.; Bikfalvi, A.; Buffat, C.; Dignat-George, F.; Anfosso, F.; Simeoni, U. Altered angiogenesis in low birth weight individuals: A role for anti-angiogenic circulating factors. J. Matern. Fetal Neonatal Med. 2014, 27, 233–238. [Google Scholar] [CrossRef]
- Ligi, I.; Simoncini, S.; Tellier, E.; Vassallo, P.F.; Sabatier, F.; Guillet, B.; Lamy, E.; Sarlon, G.; Quemener, C.; Bikfalvi, A.; et al. A switch toward angiostatic gene expression impairs the angiogenic properties of endothelial progenitor cells in low birth weight preterm infants. Blood 2011, 118, 1699–1709. [Google Scholar] [CrossRef]
- Imanishi, T.; Tsujioka, H.; Akasaka, T. Endothelial progenitor cells dysfunction and senescence: Contribution to oxidative stress. Curr. Cardiol. Rev. 2008, 4, 275–286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Satoh, M.; Ishikawa, Y.; Takahashi, Y.; Itoh, T.; Minami, Y.; Nakamura, M. Association between oxidative DNA damage and telomere shortening in circulating endothelial progenitor cells obtained from metabolic syndrome patients with coronary artery disease. Atherosclerosis 2008, 198, 347–353. [Google Scholar] [CrossRef] [PubMed]
- Thum, T.; Fraccarollo, D.; Galuppo, P.; Tsikas, D.; Frantz, S.; Ertl, G.; Bauersachs, J. Bone marrow molecular alterations after myocardial infarction: Impact on endothelial progenitor cells. Cardiovasc. Res. 2006, 70, 50–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imanishi, T.; Hano, T.; Nishio, I. Angiotensin II accelerates endothelial progenitor cell senescence through induction of oxidative stress. J. Hypertens. 2005, 23, 97–104. [Google Scholar] [CrossRef]
- Guvendag Guven, E.S.; Karcaaltincaba, D.; Kandemir, O.; Kiykac, S.; Mentese, A. Cord blood oxidative stress markers correlate with umbilical artery pulsatility in fetal growth restriction. J. Matern. Fetal Neonatal Med. 2013, 26, 576–580. [Google Scholar] [CrossRef]
- Mitchell, B.M.; Cook, L.G.; Danchuk, S.; Puschett, J.B. Uncoupled endothelial nitric oxide synthase and oxidative stress in a rat model of pregnancy-induced hypertension. Am. J. Hypertens. 2007, 20, 1297–1304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baker, C.D.; Ryan, S.L.; Ingram, D.A.; Seedorf, G.J.; Abman, S.H.; Balasubramaniam, V. Endothelial colony-forming cells from preterm infants are increased and more susceptible to hyperoxia. Am. J. Respir. Crit. Care Med. 2009, 180, 454–461. [Google Scholar] [CrossRef] [Green Version]
- Fujinaga, H.; Baker, C.D.; Ryan, S.L.; Markham, N.E.; Seedorf, G.J.; Balasubramaniam, V.; Abman, S.H. Hyperoxia disrupts vascular endothelial growth factor-nitric oxide signaling and decreases growth of endothelial colony-forming cells from preterm infants. Am. J. Physiol.-Lung Cell. Mol. Physiol. 2009, 297, L1160–L1169. [Google Scholar] [CrossRef] [Green Version]
- Karowicz-Bilinska, A.; Suzin, J.; Sieroszewski, P. Evaluation of oxidative stress indices during treatment in pregnant women with intrauterine growth retardation. Med. Sci. Monit. 2002, 8, CR211–CR216. [Google Scholar]
- Saker, M.; Soulimane Mokhtari, N.; Merzouk, S.A.; Merzouk, H.; Belarbi, B.; Narce, M. Oxidant and antioxidant status in mothers and their newborns according to birthweight. Eur. J. Obstet. Gynecol. Reprod. Biol. 2008, 141, 95–99. [Google Scholar] [CrossRef] [PubMed]
- Biri, A.; Bozkurt, N.; Turp, A.; Kavutcu, M.; Himmetoglu, O.; Durak, I. Role of oxidative stress in intrauterine growth restriction. Gynecol. Obst. Investig. 2007, 64, 187–192. [Google Scholar] [CrossRef] [PubMed]
- Gupta, P.; Narang, M.; Banerjee, B.D.; Basu, S. Oxidative stress in term small for gestational age neonates born to undernourished mothers: A case control study. BMC Pediatr. 2004, 4, 14. [Google Scholar] [CrossRef] [Green Version]
- Vassallo, P.F.; Simoncini, S.; Ligi, I.; Chateau, A.-L.; Bachelier, R.; Robert, S.; Morere, J.; Fernandez, S.; Guillet, B.; Marcelli, M.; et al. Accelerated senescence of cord blood endothelial progenitor cells in premature neonates is driven by SIRT1 decreased expression. Blood 2014, 123, 2116–2126. [Google Scholar] [CrossRef] [PubMed]
- Daniel, Z.; Swali, A.; Emes, R.; Langley-Evans, S.C. The effect of maternal undernutrition on the rat placental transcriptome: Protein restriction up-regulates cholesterol transport. Genes Nutr. 2016, 11, 27. [Google Scholar] [CrossRef] [Green Version]
- Yzydorczyk, C.; Gobeil, F., Jr.; Cambonie, G.; Lahaie, I.; Le, N.L.; Samarani, S.; Ahmad, A.; Lavoie, J.C.; Oligny, L.L.; Pladys, P.; et al. Exaggerated vasomotor response to ANG II in rats with fetal programming of hypertension associated with exposure to a low-protein diet during gestation. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2006, 291, R1060–R1068. [Google Scholar] [CrossRef] [PubMed]
- Ojeda, N.B.; Grigore, D.; Robertson, E.B.; Alexander, B.T. Estrogen protects against increased blood pressure in postpubertal female growth restricted offspring. Hypertension 2007, 50, 679–685. [Google Scholar] [CrossRef] [Green Version]
- Boudier, H.A. Arteriolar and capillary remodelling in hypertension. Drugs 1999, 58, 37–40. [Google Scholar]
- Nuyt, A.M. Mechanisms underlying developmental programming of elevated blood pressure and vascular dysfunction: Evidence from human studies and experimental animal models. Clin. Sci. 2008, 114, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Yzydorczyk, C.; Comte, B.; Cambonie, G.; Lavoie, J.C.; Germain, N.; Ting Shun, Y.; Wolff, J.; Deschepper, C.; Touyz, R.M.; Lelievre-Pegorier, M.; et al. Neonatal oxygen exposure in rats leads to cardiovascular and renal alterations in adulthood. Hypertension 2008, 52, 889–895. [Google Scholar] [CrossRef]
- Pladys, P.; Sennlaub, F.; Brault, S.; Checchin, D.; Lahaie, I.; Lê, N.L.O.; Bibeau, K.; Cambonie, G.; Abran, D.; Brochu, M.; et al. Microvascular rarefaction and decreased angiogenesis in rats with fetal programming of hypertension associated with exposure to a low-protein diet in utero. Am. J. Physiol. Integr. Comp. Physiol. 2005, 289, R1580–R1588. [Google Scholar] [CrossRef] [Green Version]
- Alphonse, R.S.; Vadivel, A.; Fung, M.; Shelley, W.C.; Critser, P.J.; Ionescu, L.; O’Reilly, M.; Ohls, R.K.; McConaghy, S.; Eaton, F.; et al. Existence, functional impairment, and lung repair potential of endothelial colony-forming cells in oxygen-induced arrested alveolar growth. Circulation 2014, 129, 2144–2157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ingram, D.A.; Mead, L.E.; Tanaka, H.; Meade, V.; Fenoglio, A.; Mortell, K.; Pollok, K.; Ferkowicz, M.J.; Gilley, D.; Yoder, M.C. Identification of a novel hierarchy of endothelial progenitor cells using human peripheral and umbilical cord blood. Blood 2004, 104, 2752–2760. [Google Scholar] [CrossRef] [PubMed]
- Tsukada, S.; Kwon, S.M.; Matsuda, T.; Jung, S.Y.; Lee, J.H.; Lee, S.H.; Masuda, H.; Asahara, T. Identification of mouse colony-forming endothelial progenitor cells for postnatal neovascularization: A novel insight highlighted by new mouse colony-forming assay. Stem. Cell Res. Ther. 2013, 4, 20. [Google Scholar] [CrossRef] [Green Version]
- Stella, C.C.; Cazzola, M.; De Fabritiis, P.; De Vincentiis, A.; Gianni, A.M.; Lanza, F.; Lauria, F.; Lemoli, R.M.; Tarella, C.; Zanon, P.; et al. CD34-positive cells: Biology and clinical relevance. Haematologica 1995, 80, 367–387. [Google Scholar]
- Lee, P.S.; Poh, K.K. Endothelial progenitor cells in cardiovascular diseases. World J. Stem. Cells 2014, 6, 355–366. [Google Scholar] [CrossRef] [PubMed]
- Shantsila, E.; Watson, T.; Lip, G.Y. Endothelial progenitor cells in cardiovascular disorders. J. Am. Coll. Cardiol. 2007, 49, 741–752. [Google Scholar] [CrossRef] [Green Version]
- Vasa, M.; Fichtlscherer, S.; Aicher, A.; Adler, K.; Urbich, C.; Martin, H.; Zeiher, A.M.; Dimmeler, S. Number and migratory activity of circulating endothelial progenitor cells inversely correlate with risk factors for coronary artery disease. Circ. Res. 2001, 89, E1–E7. [Google Scholar] [CrossRef] [Green Version]
- Peyter, A.C.; Armengaud, J.B.; Guillot, E.; Yzydorczyk, C. Endothelial Progenitor Cells Dysfunctions and Cardiometabolic Disorders: From Mechanisms to Therapeutic Approaches. Int. J. Mol. Sci. 2021, 22, 6667. [Google Scholar] [CrossRef]
- Borghesi, A.; Massa, M.; Campanelli, R.; Bollani, L.; Tzialla, C.; Figar, T.A.; Ferrari, G.; Bonetti, E.; Chiesa, G.; de Silvestri, A.; et al. Circulating endothelial progenitor cells in preterm infants with bronchopulmonary dysplasia. Am. J. Respir. Crit. Care Med. 2009, 180, 540–546. [Google Scholar] [CrossRef]
- Calcaterra, F.; Taddeo, A.; Colombo, E.; Cappelletti, M.; Martinelli, A.; Calabrese, S.; Mavilio, D.; Cetin, I.; Della Bella, S. Reduction of maternal circulating endothelial progenitor cells in human pregnancies with intrauterine growth restriction. Placenta 2014, 35, 431–436. [Google Scholar] [CrossRef] [PubMed]
- Meister, B.; Totsch, M.; Mayr, A.; Widschwendter, M.; Huter, O.; Sperl, W. Identification of CD34+ cord blood cells and their subpopulations in preterm and term neonates using three-color flow cytometry. Biol. Neonate 1994, 66, 272–279. [Google Scholar] [CrossRef] [PubMed]
- Tang, L.; He, G.; Liu, X.; Xu, W. Progress in the understanding of the etiology and predictability of fetal growth restriction. Reproduction 2017, 153, R227–R240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boeuf, P.; Aitken, E.H.; Chandrasiri, U.; Chua, C.L.; McInerney, B.; McQuade, L.; Duffy, M.; Molyneux, M.; Brown, G.; Glazier, J.; et al. Plasmodium falciparum malaria elicits inflammatory responses that dysregulate placental amino acid transport. PLoS Pathog. 2013, 9, e1003153. [Google Scholar] [CrossRef] [Green Version]
- Delia, D.; Lampugnani, M.G.; Resnati, M.; Dejana, E.; Aiello, A.; Fontanella, E.; Soligo, D.; Pierotti, M.A.; Greaves, M.F. CD34 expression is regulated reciprocally with adhesion molecules in vascular endothelial cells in vitro. Blood 1993, 81, 1001–1008. [Google Scholar] [CrossRef] [Green Version]
- Luque Contreras, D.; Vargas Robles, H.; Romo, E.; Rios, A.; Escalante, B. The role of nitric oxide in the post-ischemic revascularization process. Pharmacol. Ther. 2006, 112, 553–563. [Google Scholar] [CrossRef]
- Duda, D.G.; Fukumura, D.; Jain, R.K. Role of eNOS in neovascularization: NO for endothelial progenitor cells. Trends Mol. Med. 2004, 10, 143–145. [Google Scholar] [CrossRef]
- Babaei, S.; Stewart, D.J. Overexpression of endothelial NO synthase induces angiogenesis in a co-culture model. Cardiovasc. Res. 2002, 55, 190–200. [Google Scholar] [CrossRef] [Green Version]
- Grandvuillemin, I.; Buffat, C.; Boubred, F.; Lamy, E.; Fromonot, J.; Charpiot, P.; Simoncini, S.; Sabatier, F.; Dignat-George, F.; Peyter, A.C.; et al. Arginase up-regulation and eNOS uncoupling contribute to impaired endothelium-dependent vasodilation in a rat model of intrauterine growth restriction. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2018, 315, R509–R520. [Google Scholar] [CrossRef]
- Hoetzer, G.L.; Irmiger, H.M.; Keith, R.S.; Westbrook, K.M.; DeSouza, C.A. Endothelial nitric oxide synthase inhibition does not alter endothelial progenitor cell colony forming capacity or migratory activity. J. Cardiovasc. Pharmacol. 2005, 46, 387–389. [Google Scholar] [CrossRef]
- Prieto, C.P.; Krause, B.J.; Quezada, C.; San Martin, R.; Sobrevia, L.; Casanello, P. Hypoxia-reduced nitric oxide synthase activity is partially explained by higher arginase-2 activity and cellular redistribution in human umbilical vein endothelium. Placenta 2011, 32, 932–940. [Google Scholar] [CrossRef]
- Cogswell, M.E.; Yip, R. The influence of fetal and maternal factors on the distribution of birthweight. Semin Perinatol. 1995, 19, 222–240. [Google Scholar] [CrossRef]
- Kalka, C.; Masuda, H.; Takahashi, T.; Gordon, R.; Tepper, O.; Gravereaux, E.; Pieczek, A.; Iwaguro, H.; Hayashi, S.I.; Isner, J.M.; et al. Vascular endothelial growth factor(165) gene transfer augments circulating endothelial progenitor cells in human subjects. Circ. Res. 2000, 86, 1198–1202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Young, P.P.; Hofling, A.A.; Sands, M.S. VEGF increases engraftment of bone marrow-derived endothelial progenitor cells (EPCs) into vasculature of newborn murine recipients. Proc. Natl. Acad. Sci. USA 2002, 99, 11951–11956. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dulak, J.; Jozkowicz, A.; Frick, M.; Alber, H.F.; Dichtl, W.; Schwarzacher, S.P.; Pachinger, O.; Weidinger, F. Vascular endothelial growth factor: Angiogenesis, atherogenesis or both? J. Am. Coll. Cardiol. 2001, 38, 2137–2138. [Google Scholar] [CrossRef] [Green Version]
- Dulak, J.; Jozkowicz, A.; Dembinska-Kiec, A.; Guevara, I.; Zdzienicka, A.; Zmudzinska-Grochot, D.; Florek, I.; Wojtowicz, A.; Szuba, A.; Cooke, J.P. Nitric oxide induces the synthesis of vascular endothelial growth factor by rat vascular smooth muscle cells. Arterioscler. Thromb. Vasc. Biol. 2000, 20, 659–666. [Google Scholar] [CrossRef] [Green Version]
- Kimura, H.; Esumi, H. Reciprocal regulation between nitric oxide and vascular endothelial growth factor in angiogenesis. Acta Biochim. Pol. 2003, 50, 49–59. [Google Scholar] [CrossRef] [Green Version]
- Qin, W.; Xie, W.; Xia, N.; He, Q.; Sun, T. Silencing of Transient Receptor Potential Channel 4 Alleviates oxLDL-induced Angiogenesis in Human Coronary Artery Endothelial Cells by Inhibition of VEGF and NF-kappaB. Med. Sci. Monit. 2016, 22, 930–936. [Google Scholar] [CrossRef] [Green Version]
- Di, Y.; Zhang, D.; Hu, T.; Li, D. miR-23 regulate the pathogenesis of patients with coronary artery disease. Int. J. Clin. Exp. Med. 2015, 8, 11759–11769. [Google Scholar]
- Koch, S.; Claesson-Welsh, L. Signal transduction by vascular endothelial growth factor receptors. Cold Spring Harb. Perspect. Med. 2012, 2, a006502. [Google Scholar] [CrossRef]
- Dong, D.; Khoong, Y.; Ko, Y.; Zhang, Y. microRNA-646 inhibits angiogenesis of endothelial progenitor cells in pre-eclamptic pregnancy by targeting the VEGF-A/HIF-1alpha axis. Exp. Ther. Med. 2020, 20, 1879–1888. [Google Scholar] [CrossRef]
- Rozance, P.J.; Anderson, M.; Martinez, M.; Fahy, A.; Macko, A.R.; Kailey, J.; Seedorf, G.J.; Abman, S.H.; Hay, W.W., Jr.; Limesand, S.W. Placental insufficiency decreases pancreatic vascularity and disrupts hepatocyte growth factor signaling in the pancreatic islet endothelial cell in fetal sheep. Diabetes 2015, 64, 555–564. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Ji, Y.; Cai, S.; Ding, W. MiR-206 Suppresses the Progression of Coronary Artery Disease by Modulating Vascular Endothelial Growth Factor (VEGF) Expression. Med. Sci. Monit. 2016, 22, 5011–5020. [Google Scholar] [CrossRef] [Green Version]
- Ii, M.; Takenaka, H.; Asai, J.; Ibusuki, K.; Mizukami, Y.; Maruyama, K.; Yoon, Y.S.; Wecker, A.; Luedemann, C.; Eaton, E.; et al. Endothelial progenitor thrombospondin-1 mediates diabetes-induced delay in reendothelialization following arterial injury. Circ. Res. 2006, 98, 697–704. [Google Scholar] [CrossRef]
- Schurmann, C.; Schmidt, N.; Seitz, O.; Pfeilschifter, J.; Frank, S. Angiogenic response pattern during normal and impaired skin flap re-integration in mice: A comparative study. J. Craniomaxillofac. Surg. 2014, 42, 1710–1716. [Google Scholar] [CrossRef]
- Babaei, S.; Teichert-Kuliszewska, K.; Zhang, Q.; Jones, N.; Dumont, D.J.; Stewart, D.J. Angiogenic actions of angiopoietin-1 require endothelium-derived nitric oxide. Am. J. Pathol. 2003, 162, 1927–1936. [Google Scholar] [CrossRef] [Green Version]
- Balaji, S.; Han, N.; Moles, C.; Shaaban, A.F.; Bollyky, P.L.; Crombleholme, T.M.; Keswani, S.G. Angiopoietin-1 improves endothelial progenitor cell-dependent neovascularization in diabetic wounds. Surgery 2015, 158, 846–856. [Google Scholar] [CrossRef] [Green Version]
- Davis, S.; Aldrich, T.H.; Jones, P.F.; Acheson, A.; Compton, D.L.; Jain, V.; Ryan, T.E.; Bruno, J.; Radziejewski, C.; Maisonpierre, P.C.; et al. Isolation of angiopoietin-1, a ligand for the TIE2 receptor, by secretion-trap expression cloning. Cell 1996, 87, 1161–1169. [Google Scholar] [CrossRef] [Green Version]
- Troyanovsky, B.; Levchenko, T.; Mansson, G.; Matvijenko, O.; Holmgren, L. Angiomotin: An angiostatin binding protein that regulates endothelial cell migration and tube formation. J. Cell Biol. 2001, 152, 1247–1254. [Google Scholar] [CrossRef] [PubMed]
- Kappou, D.; Sifakis, S.; Androutsopoulos, V.; Konstantinidou, A.; Spandidos, D.A.; Papantoniou, N. Placental mRNA expression of angiopoietins (Ang)-1, Ang-2 and their receptor Tie-2 is altered in pregnancies complicated by preeclampsia. Placenta 2014, 35, 718–723. [Google Scholar] [CrossRef] [PubMed]
- Aase, K.; Ernkvist, M.; Ebarasi, L.; Jakobsson, L.; Majumdar, A.; Yi, C.; Birot, O.; Ming, Y.; Kvanta, A.; Edholm, D.; et al. Angiomotin regulates endothelial cell migration during embryonic angiogenesis. Genes Dev. 2007, 21, 2055–2068. [Google Scholar] [CrossRef] [Green Version]
- Griendling, K.K.; Sorescu, D.; Ushio-Fukai, M. NAD(P)H oxidase: Role in cardiovascular biology and disease. Circ. Res. 2000, 86, 494–501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zalba, G.; San Jose, G.; Moreno, M.U.; Fortuno, M.A.; Fortuno, A.; Beaumont, F.J.; Diez, J. Oxidative stress in arterial hypertension: Role of NAD(P)H oxidase. Hypertension 2001, 38, 1395–1399. [Google Scholar] [CrossRef] [PubMed]
- He, T.; Peterson, T.E.; Holmuhamedov, E.L.; Terzic, A.; Caplice, N.M.; Oberley, L.W.; Katusic, Z.S. Human endothelial progenitor cells tolerate oxidative stress due to intrinsically high expression of manganese superoxide dismutase. Arterioscler. Thromb. Vasc. Biol. 2004, 24, 2021–2027. [Google Scholar] [CrossRef] [Green Version]
- Ingram, D.A.; Krier, T.R.; Mead, L.E.; McGuire, C.; Prater, D.N.; Bhavsar, J.; Saadatzadeh, M.R.; Bijangi-Vishehsaraei, K.; Li, F.; Yoder, M.C.; et al. Clonogenic endothelial progenitor cells are sensitive to oxidative stress. Stem. Cells 2007, 25, 297–304. [Google Scholar] [CrossRef]
- Rodriguez-Iturbe, B.; Zhan, C.D.; Quiroz, Y.; Sindhu, R.K.; Vaziri, N.D. Antioxidant-rich diet relieves hypertension and reduces renal immune infiltration in spontaneously hypertensive rats. Hypertension 2003, 41, 341–346. [Google Scholar] [CrossRef] [Green Version]
- Tanito, M.; Nakamura, H.; Kwon, Y.W.; Teratani, A.; Masutani, H.; Shioji, K.; Kishimoto, C.; Ohira, A.; Horie, R.; Yodoi, J. Enhanced oxidative stress and impaired thioredoxin expression in spontaneously hypertensive rats. Antioxid. Redox. Signal. 2004, 6, 89–97. [Google Scholar] [CrossRef]
- Park, J.B.; Touyz, R.M.; Chen, X.; Schiffrin, E.L. Chronic treatment with a superoxide dismutase mimetic prevents vascular remodeling and progression of hypertension in salt-loaded stroke-prone spontaneously hypertensive rats. Am. J. Hypertens. 2002, 15, 78–84. [Google Scholar] [CrossRef]
- Yzydorczyk, C.; Li, N.; Chehade, H.; Mosig, D.; Bidho, M.; Keshavjee, B.; Armengaud, J.B.; Nardou, K.; Siddeek, B.; Benahmed, M.; et al. Transient postnatal overfeeding causes liver stress-induced premature senescence in adult mice. Sci. Rep. 2017, 7, 12911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Endtmann, C.; Ebrahimian, T.; Czech, T.; Arfa, O.; Laufs, U.; Fritz, M.; Wassmann, K.; Werner, N.; Petoumenos, V.; Nickenig, G.; et al. Angiotensin II impairs endothelial progenitor cell number and function in vitro and in vivo: Implications for vascular regeneration. Hypertension 2011, 58, 394–403. [Google Scholar] [CrossRef]
- Yao, E.H.; Fukuda, N.; Matsumoto, T.; Kobayashi, N.; Katakawa, M.; Yamamoto, C.; Tsunemi, A.; Suzuki, R.; Ueno, T.; Matsumoto, K. Losartan improves the impaired function of endothelial progenitor cells in hypertension via an antioxidant effect. Hypertens Res. 2007, 30, 1119–1128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minamino, T.; Komuro, I. Vascular cell senescence: Contribution to atherosclerosis. Circ. Res. 2007, 100, 15–26. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.Y.; Han, J.A.; Im, J.S.; Morrone, A.; Johung, K.; Goodwin, E.C.; Kleijer, W.J.; DiMaio, D.; Hwang, E.S. Senescence-associated beta-galactosidase is lysosomal beta-galactosidase. Aging Cell 2006, 5, 187–195. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, V.; de Souza, L.V.; Fernandes, T.; Junior, S.D.S.; de Carvalho, M.H.C.; Akamine, E.H.; Michelini, L.C.; de Oliveira, E.M.; Franco, M.D.C. Intrauterine growth restriction-induced deleterious adaptations in endothelial progenitor cells: Possible mechanism to impair endothelial function. J. Dev. Orig. Health Dis. 2017, 8, 665–673. [Google Scholar] [CrossRef] [PubMed]
- Re, R.N.; Cook, J.L. Senescence, apoptosis, and stem cell biology: The rationale for an expanded view of intracrine action. Am. J. Physiol. Heart Circ. Physiol. 2009, 297, H893–H901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ovadya, Y.; Krizhanovsky, V. Senescent cells: SASPected drivers of age-related pathologies. Biogerontology 2014, 15, 627–642. [Google Scholar] [CrossRef]
- Simoncini, S.; Chateau, A.L.; Robert, S.; Todorova, D.; Yzydorzick, C.; Lacroix, R.; Ligi, I.; Louis, L.; Bachelier, R.; Simeoni, U.; et al. Biogenesis of Pro-senescent Microparticles by Endothelial Colony Forming Cells from Premature Neonates is driven by SIRT1-Dependent Epigenetic Regulation of MKK6. Sci. Rep. 2017, 7, 8277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cuenda, A.; Rousseau, S. p38 MAP-kinases pathway regulation, function and role in human diseases. Biochim. Biophys. Acta 2007, 1773, 1358–1375. [Google Scholar] [CrossRef] [Green Version]
- Freund, A.; Patil, C.K.; Campisi, J. p38MAPK is a novel DNA damage response-independent regulator of the senescence-associated secretory phenotype. EMBO J. 2011, 30, 1536–1548. [Google Scholar] [CrossRef] [Green Version]
- Corre, I.; Paris, F.; Huot, J. The p38 pathway, a major pleiotropic cascade that transduces stress and metastatic signals in endothelial cells. Oncotarget 2017, 8, 55684–55714. [Google Scholar] [CrossRef]
- Shen, X.H.; Xu, S.J.; Jin, C.Y.; Ding, F.; Zhou, Y.C.; Fu, G.S. Interleukin-8 prevents oxidative stress-induced human endothelial cell senescence via telomerase activation. Int. Immunopharmacol. 2013, 16, 261–267. [Google Scholar] [CrossRef]
- Romero, A.; Dongil, P.; Valencia, I.; Vallejo, S.; San Hipólito-Luengo, Á.; Díaz-Araya, G.; Peiró, C. Pharmacological Blockade of NLRP3 Inflammasome/IL-1β-Positive Loop Mitigates Endothelial Cell Senescence and Dysfunction. Aging Dis. 2021, 13. [Google Scholar] [CrossRef]
- Armengaud, J.B.; Dennebouy, Z.; Labes, D.; Fumey, C.; Wilson, A.; Candotti, F.; Yzydorczyk, C.; Simeoni, U. Intrauterine Growth Restriction Induced by Maternal Low Protein Diet Causes Long-Term Alterations of Thymic Structure and Function in Adult Male Rat Offspring. Br. J. Nutr. 2020, 123, 892–900. [Google Scholar] [CrossRef]
- Piotrowska, H.; Kucinska, M.; Murias, M. Biological activity of piceatannol: Leaving the shadow of resveratrol. Mutat. Res. 2012, 750, 60–82. [Google Scholar] [CrossRef] [PubMed]
- Mei, Y.Z.; Liu, R.X.; Wang, D.P.; Wang, X.; Dai, C.C. Biocatalysis and biotransformation of resveratrol in microorganisms. Biotechnol. Lett. 2015, 37, 9–18. [Google Scholar] [CrossRef]
- Wang, X.B.; Huang, J.; Zou, J.G.; Su, E.B.; Shan, Q.J.; Yang, Z.J.; Cao, K.J. Effects of resveratrol on number and activity of endothelial progenitor cells from human peripheral blood. Clin. Exp. Pharmacol. Physiol. 2007, 34, 1109–1115. [Google Scholar] [CrossRef] [PubMed]
- Wallerath, T.; Deckert, G.; Ternes, T.; Anderson, H.; Li, H.; Witte, K.; Forstermann, U. Resveratrol, a polyphenolic phytoalexin present in red wine, enhances expression and activity of endothelial nitric oxide synthase. Circulation 2002, 106, 1652–1658. [Google Scholar] [CrossRef] [Green Version]
- Gracia-Sancho, J.; Villarreal, G., Jr.; Zhang, Y.; Garcia-Cardena, G. Activation of SIRT1 by resveratrol induces KLF2 expression conferring an endothelial vasoprotective phenotype. Cardiovasc. Res. 2010, 85, 514–519. [Google Scholar] [CrossRef] [Green Version]
- Xia, L.; Wang, X.X.; Hu, X.S.; Guo, X.G.; Shang, Y.P.; Chen, H.J.; Zeng, C.L.; Zhang, F.R.; Chen, J.Z. Resveratrol reduces endothelial progenitor cells senescence through augmentation of telomerase activity by Akt-dependent mechanisms. Br. J. Pharmacol. 2008, 155, 387–394. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.B.; Zhu, L.; Huang, J.; Yin, Y.G.; Kong, X.Q.; Rong, Q.F.; Shi, A.W.; Cao, K.J. Resveratrol-induced augmentation of telomerase activity delays senescence of endothelial progenitor cells. Chin. Med. J. 2011, 124, 4310–4315. [Google Scholar] [PubMed]
- Wu, H.; Li, G.N.; Xie, J.; Li, R.; Chen, Q.H.; Chen, J.Z.; Wei, Z.H.; Kang, L.N.; Xu, B. Resveratrol ameliorates myocardial fibrosis by inhibiting ROS/ERK/TGF-beta/periostin pathway in STZ-induced diabetic mice. BMC Cardiovasc. Disord. 2016, 16, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basile, D.P.; Yoder, M.C. Circulating and tissue resident endothelial progenitor cells. J. Cell Physiol. 2014, 229, 10–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoder, M.C. Endothelial progenitor cell: A blood cell by many other names may serve similar functions. J. Mol. Med. 2013, 91, 285–295. [Google Scholar] [CrossRef] [PubMed]
- Chong, M.S.; Ng, W.K.; Chan, J.K. Concise Review: Endothelial Progenitor Cells in Regenerative Medicine: Applications and Challenges. Stem. Cells Transl. Med. 2016, 5, 530–538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.X.; Zhang, F.R.; Shang, Y.P.; Zhu, J.H.; Xie, X.D.; Tao, Q.M.; Zhu, J.H.; Chen, J.Z. Transplantation of autologous endothelial progenitor cells may be beneficial in patients with idiopathic pulmonary arterial hypertension: A pilot randomized controlled trial. J. Am. Coll. Cardiol. 2007, 49, 1566–1571. [Google Scholar] [CrossRef] [Green Version]
- Ambasta, R.K.; Kohli, H.; Kumar, P. Multiple therapeutic effect of endothelial progenitor cell regulated by drugs in diabetes and diabetes related disorder. J. Transl. Med. 2017, 15, 185. [Google Scholar] [CrossRef] [PubMed]
- Girard-Bock, C.; de Araujo, C.C.; Bertagnolli, M.; Mai-Vo, T.A.; Vadivel, A.; Alphonse, R.S.; Zhong, S.; Cloutier, A.; Sutherland, M.R.; Thebaud, B.; et al. Endothelial colony-forming cell therapy for heart morphological changes after neonatal high oxygen exposure in rats, a model of complications of prematurity. Physiol. Rep. 2018, 6, e13922. [Google Scholar] [CrossRef] [Green Version]
- Munoz-Hernandez, R.; Miranda, M.L.; Stiefel, P.; Lin, R.Z.; Praena-Fernandez, J.M.; Dominguez-Simeon, M.J.; Villar, J.; Moreno-Luna, R.; Melero-Martin, J.M. Decreased level of cord blood circulating endothelial colony-forming cells in preeclampsia. Hypertension 2014, 64, 165–171. [Google Scholar] [CrossRef] [Green Version]
- Luppi, P.; Powers, R.W.; Verma, V.; Edmunds, L.; Plymire, D.; Hubel, C.A. Maternal circulating CD34+VEGFR-2+ and CD133+VEGFR-2+ progenitor cells increase during normal pregnancy but are reduced in women with preeclampsia. Reprod. Sci. 2010, 17, 643–652. [Google Scholar] [CrossRef] [Green Version]
- Monga, R.; Buck, S.; Sharma, P.; Thomas, R.; Chouthai, N.S. Effect of preeclampsia and intrauterine growth restriction on endothelial progenitor cells in human umbilical cord blood. J. Matern. Fetal. Neonatal Med. 2012, 25, 2385–2389. [Google Scholar] [CrossRef] [PubMed]
- Rana, S.; Karumanchi, S.A.; Lindheimer, M.D. Angiogenic factors in diagnosis, management, and research in preeclampsia. Hypertension 2014, 63, 198–202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karumanchi, S.A. Angiogenic Factors in Preeclampsia: From Diagnosis to Therapy. Hypertension 2016, 67, 1072–1079. [Google Scholar] [CrossRef] [PubMed]
- Smadja, D.M.; Cornet, A.; Emmerich, J.; Aiach, M.; Gaussem, P. Endothelial progenitor cells: Characterization, in vitro expansion, and prospects for autologous cell therapy. Cell Biol. Toxicol. 2007, 23, 223–239. [Google Scholar] [CrossRef] [PubMed]
- Essaadi, A.; Nollet, M.; Moyon, A.; Stalin, J.; Simoncini, S.; Balasse, L.; Bertaud, A.; Bachelier, R.; Leroyer, A.S.; Sarlon, G.; et al. Stem cell properties of peripheral blood endothelial progenitors are stimulated by soluble CD146 via miR-21: Potential use in autologous cell therapy. Sci. Rep. 2018, 8, 9387. [Google Scholar] [CrossRef] [Green Version]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Body Weight at Birth | CTRL (gram) | IUGR (gram) | SIGNIFICANCE |
| Males (n = 25; 5 litters) | 7.73 ± 1.03 | 5.23 ± 0.48 | p < 0.001 |
| Females (n = 25; 5 litters) | 7.15 ± 0.48 | 4.76 ± 0.40 | p < 0.001 |
| Body Weight at 6 Months | CTRL (gram) | IUGR (gram) | SIGNIFICANCE |
| Males (n = 25; 5 litters) | 751.81 ± 64.64 | 586.16 ± 45.43 | p < 0.001 |
| Females (n = 25; 5 litters) | 378.62 ± 35.95 | 319.78 ± 16.94 | p < 0.001 |
| SBP at 6 Months of Life | CTRL (mmHg) | IUGR (mmHg) | SIGNIFICANCE |
| Males (n = 5; 5 litters) | 125.72 ± 4.62 | 153.44 ± 2.51 | p < 0.01 |
| Females (n = 5; 5 litters) | 112.56 ± 7.73 | 113.8 ± 5.92 | p > 0.05 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Simoncini, S.; Coppola, H.; Rocca, A.; Bachmann, I.; Guillot, E.; Zippo, L.; Dignat-George, F.; Sabatier, F.; Bedel, R.; Wilson, A.; et al. Endothelial Colony-Forming Cells Dysfunctions Are Associated with Arterial Hypertension in a Rat Model of Intrauterine Growth Restriction. Int. J. Mol. Sci. 2021, 22, 10159. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms221810159
Simoncini S, Coppola H, Rocca A, Bachmann I, Guillot E, Zippo L, Dignat-George F, Sabatier F, Bedel R, Wilson A, et al. Endothelial Colony-Forming Cells Dysfunctions Are Associated with Arterial Hypertension in a Rat Model of Intrauterine Growth Restriction. International Journal of Molecular Sciences. 2021; 22(18):10159. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms221810159
Chicago/Turabian StyleSimoncini, Stephanie, Hanna Coppola, Angela Rocca, Isaline Bachmann, Estelle Guillot, Leila Zippo, Françoise Dignat-George, Florence Sabatier, Romain Bedel, Anne Wilson, and et al. 2021. "Endothelial Colony-Forming Cells Dysfunctions Are Associated with Arterial Hypertension in a Rat Model of Intrauterine Growth Restriction" International Journal of Molecular Sciences 22, no. 18: 10159. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms221810159