
Whence CRIPTO: The Reemergence of an Oncofetal Factor in ‘Wounds’ That Fail to Heal

 , , , , and
, , , , and
Abstract
:1. Introduction
Epigraphs
“Elements which have retained their […] embryonal characteristics in the adult organism or have regained them through some chemico-physiologic deviation, represent […] the generative elements of every tumor variety and specifically those of a malignant nature. Such elements may remain enclosed within matured tissues for many years, giving no indication of their presence, until an irritation—a simple stimulus suffices—rekindles their vital cellular activities…”—Mueller, 1874 [1].
“Could we but learn from whence his sorrows grow, we would as willingly give cure as know.”—William Shakespeare.
2. Rise and Fall of CRIPTO
2.1. CRIPTO Regulates Undifferentiated Cellular Phenotypes during Development
2.2. CRIPTO Hides
2.3. CRIPTO Re-Emerges in Wound Healing
2.4. A Role for CRIPTO in Fibrosis
3. Re-Emergence of CRIPTO
3.1. Diverse Tumors Invoke CRIPTO
3.2. CRIPTO in Specialized Tumor Cells
3.3. CRIPTOs Remergence in Cancer May Reflect Tumor Vascularization
3.4. CRIPTO as a Wound Healing Hallmark and a Fibrotic Marker in Cancer
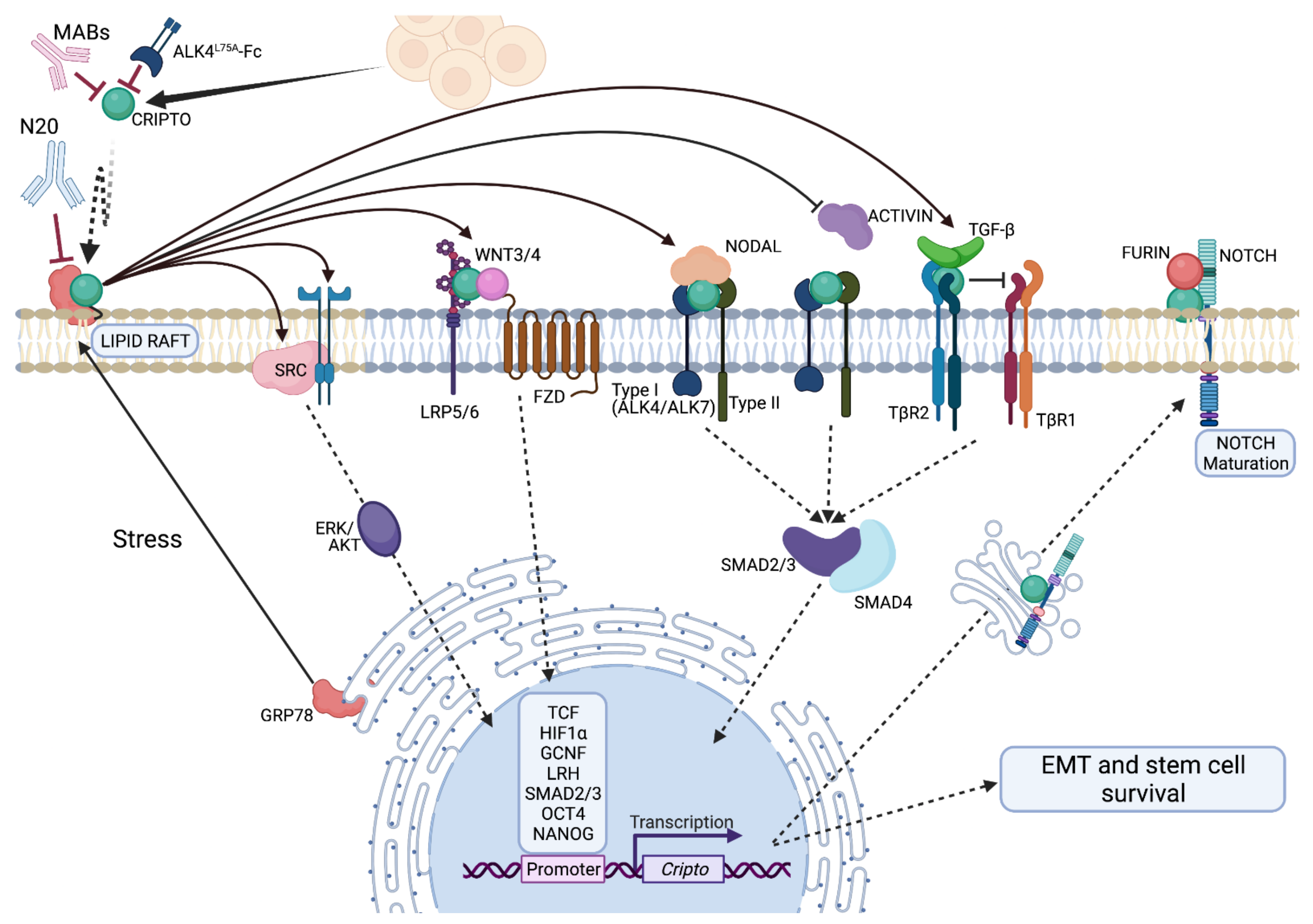
4. Classical and Emerging Mechanisms of Signaling
4.1. CRIPTO Integrates TGF-β and Growth Factor Related Signaling Pathways
4.2. Regulation of CRIPTO Signaling
4.3. CRIPTO Signaling in Trans
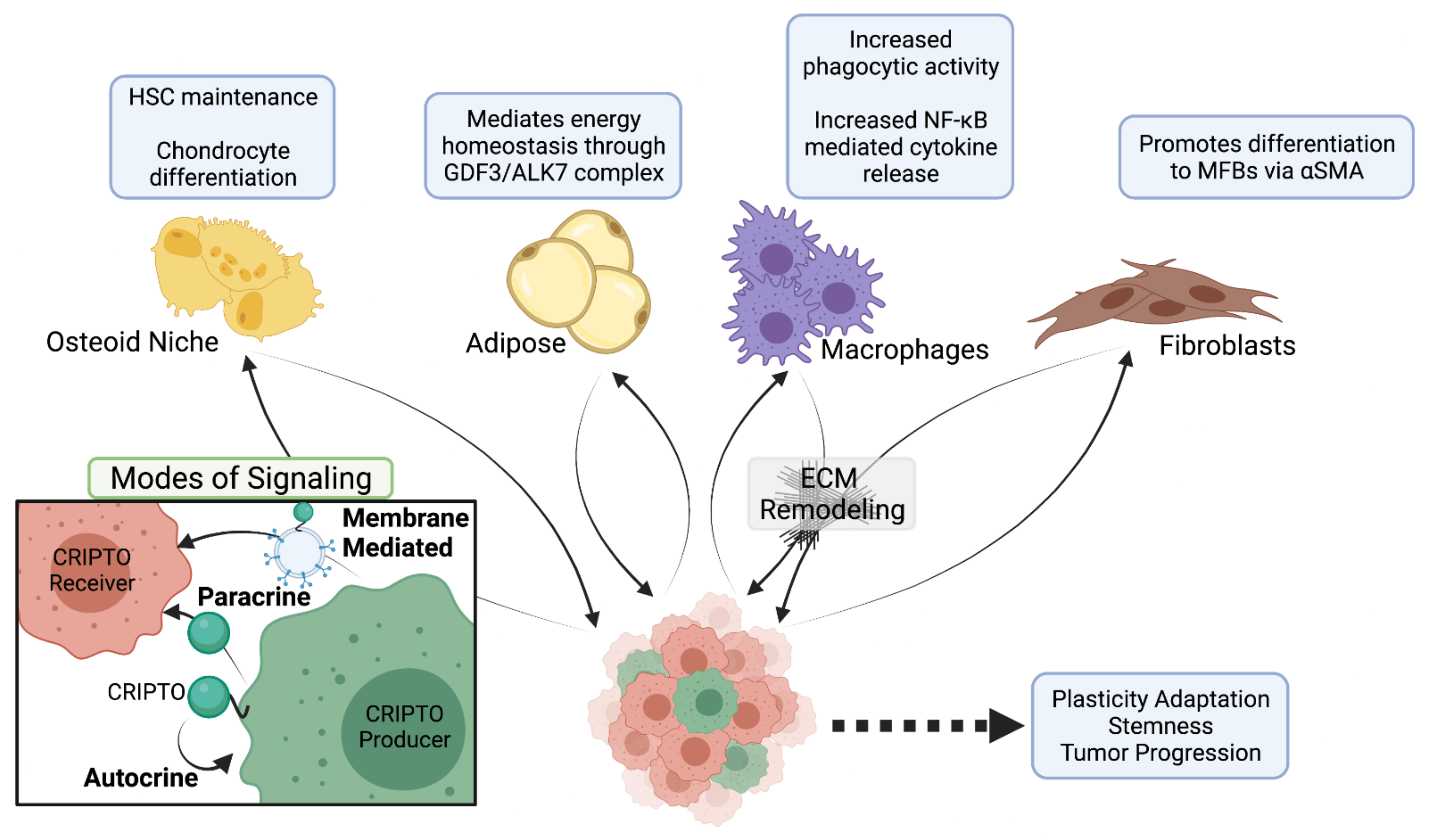
5. CRIPTO’s Role in Cancer
5.1. CRIPTO as a Driver of Tumor Progression and Metastasis
5.2. CRIPTO Action at a Distance
5.3. Promoting Metastatic Inroads
6. CRIPTO in the Clinic
6.1. CRIPTO as a Diagnostic Marker
6.2. Therapeutic Targeting of CRIPTO Signaling
7. Conclusions and Next Steps
7.1. Bridging the Gaps
7.2. Forging Ahead
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Spike, B.T. Chapter 5—Breast Cancer Stem Cells and the Move Toward High-Resolution Stem Cell Systems. In Cancer Stem Cells; Liu, H., Lathia, J.D., Eds.; Academic Press: Boston, MA, USA, 2016; pp. 121–148. [Google Scholar] [CrossRef]
- Brewer, B.G.; Mitchell, R.A.; Harandi, A.; Eaton, J.W. Embryonic vaccines against cancer: An early history. Exp. Mol. Pathol. 2009, 86, 192–197. [Google Scholar] [CrossRef]
- Klauzinska, M.; Castro, N.P.; Rangel, M.C.; Spike, B.T.; Gray, P.C.; Bertolette, D.; Cuttitta, F.; Salomon, D. The multifaceted role of the embryonic gene Cripto-1 in cancer, stem cells and epithelial-mesenchymal transition. Semin. Cancer Biol. 2014, 29, 51–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bianco, C.; Strizzi, L.; Normanno, N.; Khan, N.; Salomon, D.S. 3—Cripto-1: An Oncofetal Gene with Many Faces. In Current Topics in Developmental Biology; Schatten, G.P., Ed.; Academic Press: Cambridge, MA, USA, 2005; Volume 67, pp. 85–133. [Google Scholar]
- Rodrigues Sousa, E.; Zoni, E.; Karkampouna, S.; La Manna, F.; Gray, P.C.; De Menna, M.; Kruithof-de Julio, M. A Multidisciplinary Review of the Roles of Cripto in the Scientific Literature Through a Bibliometric Analysis of its Biological Roles. Cancers 2020, 12, 1480. [Google Scholar] [CrossRef]
- Bianco, C.; Rangel, M.C.; Castro, N.P.; Nagaoka, T.; Rollman, K.; Gonzales, M.; Salomon, D.S. Role of Cripto-1 in stem cell maintenance and malignant progression. Am. J. Pathol. 2010, 177, 532–540. [Google Scholar] [CrossRef]
- Sandomenico, A.; Ruvo, M. Targeting Nodal and Cripto-1: Perspectives Inside Dual Potential Theranostic Cancer Biomarkers. Curr. Med. Chem. 2019, 26, 1994–2050. [Google Scholar] [CrossRef] [PubMed]
- Minchiotti, G.; Parisi, S.; Liguori, G.; Signore, M.; Lania, G.; Adamson, E.D.; Lago, C.T.; Persico, M.G. Membrane-anchorage of Cripto protein by glycosylphosphatidylinositol and its distribution during early mouse development. Mech. Dev. 2000, 90, 133–142. [Google Scholar] [CrossRef]
- Zhang, J.; Talbot, W.S.; Schier, A.F. Positional cloning identifies zebrafish one-eyed pinhead as a permissive EGF-related ligand required during gastrulation. Cell 1998, 92, 241–251. [Google Scholar] [CrossRef] [Green Version]
- Saloman, D.S.; Bianco, C.; Ebert, A.D.; Khan, N.I.; De Santis, M.; Normanno, N.; Wechselberger, C.; Seno, M.; Williams, K.; Sanicola, M.; et al. The EGF-CFC family: Novel epidermal growth factor-related proteins in development and cancer. Endocr. Relat. Cancer 2000, 7, 199–226. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, K.; Nagaoka, T.; Strizzi, L.; Mancino, M.; Gonzales, M.; Bianco, C.; Salomon, D.S. Characterization of the glycosylphosphatidylinositol-anchor signal sequence of human Cryptic with a hydrophilic extension. Biochim. Biophys. Acta 2008, 1778, 2671–2681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dono, R.; Scalera, L.; Pacifico, F.; Acampora, D.; Persico, M.G.; Simeone, A. The murine cripto gene: Expression during mesoderm induction and early heart morphogenesis. Development 1993, 118, 1157–1168. [Google Scholar] [CrossRef] [PubMed]
- Kinoshita, N.; Minshull, J.; Kirschner, M.W. The identification of two novel ligands of the FGF receptor by a yeast screening method and their activity in Xenopus development. Cell 1995, 83, 621–630. [Google Scholar] [CrossRef] [Green Version]
- Ravisankar, V.; Singh, T.P.; Manoj, N. Molecular evolution of the EGF-CFC protein family. Gene 2011, 482, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Ding, J.; Yang, L.; Yan, Y.T.; Chen, A.; Desai, N.; Wynshaw-Boris, A.; Shen, M.M. Cripto is required for correct orientation of the anterior-posterior axis in the mouse embryo. Nature 1998, 395, 702–707. [Google Scholar] [CrossRef]
- Jin, J.Z.; Ding, J. Cripto is required for mesoderm and endoderm cell allocation during mouse gastrulation. Dev. Biol. 2013, 381, 170–178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reissmann, E.; Jornvall, H.; Blokzijl, A.; Andersson, O.; Chang, C.; Minchiotti, G.; Persico, M.G.; Ibanez, C.F.; Brivanlou, A.H. The orphan receptor ALK7 and the Activin receptor ALK4 mediate signaling by Nodal proteins during vertebrate development. Genes Dev. 2001, 15, 2010–2022. [Google Scholar] [CrossRef] [Green Version]
- Gray, P.C.; Vale, W. Cripto/GRP78 modulation of the TGF-beta pathway in development and oncogenesis. FEBS Lett. 2012, 586, 1836–1845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pawlak, J.B.; Blobe, G.C. TGF-beta superfamily co-receptors in cancer. Dev. Dyn. 2021. [Google Scholar] [CrossRef] [PubMed]
- Gray, P.C.; Shani, G.; Aung, K.; Kelber, J.; Vale, W. Cripto binds transforming growth factor beta (TGF-beta) and inhibits TGF-beta signaling. Mol. Cell Biol. 2006, 26, 9268–9278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adkins, H.B.; Bianco, C.; Schiffer, S.G.; Rayhorn, P.; Zafari, M.; Cheung, A.E.; Orozco, O.; Olson, D.; De Luca, A.; Chen, L.L.; et al. Antibody blockade of the Cripto CFC domain suppresses tumor cell growth in vivo. J. Clin. Investig. 2003, 112, 575–587. [Google Scholar] [CrossRef] [Green Version]
- Gray, P.C.; Harrison, C.A.; Vale, W. Cripto forms a complex with activin and type II activin receptors and can block activin signaling. Proc. Natl. Acad. Sci. USA 2003, 100, 5193–5198. [Google Scholar] [CrossRef] [Green Version]
- Assou, S.; Cerecedo, D.; Tondeur, S.; Pantesco, V.; Hovatta, O.; Klein, B.; Hamamah, S.; De Vos, J. A gene expression signature shared by human mature oocytes and embryonic stem cells. BMC Genom. 2009, 10, 10. [Google Scholar] [CrossRef] [Green Version]
- Mizuno, H.; Spike, B.T.; Wahl, G.M.; Levine, A.J. Inactivation of p53 in breast cancers correlates with stem cell transcriptional signatures. Proc. Natl. Acad. Sci. USA 2010, 107, 22745–22750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hough, S.R.; Laslett, A.L.; Grimmond, S.B.; Kolle, G.; Pera, M.F. A continuum of cell states spans pluripotency and lineage commitment in human embryonic stem cells. PLoS ONE 2009, 4, e7708. [Google Scholar] [CrossRef]
- Takahashi, K.; Yamanaka, S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell 2006, 126, 663–676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fiorenzano, A.; Pascale, E.; D’Aniello, C.; Acampora, D.; Bassalert, C.; Russo, F.; Andolfi, G.; Biffoni, M.; Francescangeli, F.; Zeuner, A.; et al. Cripto is essential to capture mouse epiblast stem cell and human embryonic stem cell pluripotency. Nat. Commun. 2016, 7, 12589. [Google Scholar] [CrossRef]
- Wei, C.L.; Miura, T.; Robson, P.; Lim, S.K.; Xu, X.Q.; Lee, M.Y.; Gupta, S.; Stanton, L.; Luo, Y.; Schmitt, J.; et al. Transcriptome profiling of human and murine ESCs identifies divergent paths required to maintain the stem cell state. Stem Cells 2005, 23, 166–185. [Google Scholar] [CrossRef]
- Loh, Y.H.; Wu, Q.; Chew, J.L.; Vega, V.B.; Zhang, W.; Chen, X.; Bourque, G.; George, J.; Leong, B.; Liu, J.; et al. The Oct4 and Nanog transcription network regulates pluripotency in mouse embryonic stem cells. Nat. Genet. 2006, 38, 431–440. [Google Scholar] [CrossRef] [PubMed]
- Gershon, E.; Hadas, R.; Elbaz, M.; Booker, E.; Muchnik, M.; Kleinjan-Elazary, A.; Karasenti, S.; Genin, O.; Cinnamon, Y.; Gray, P.C. Identification of Trophectoderm-Derived Cripto as an Essential Mediator of Embryo Implantation. Endocrinology 2018, 159, 1793–1807. [Google Scholar] [CrossRef]
- Bandeira, C.L.; Urban Borbely, A.; Pulcineli Vieira Francisco, R.; Schultz, R.; Zugaib, M.; Bevilacqua, E. Tumorigenic factor CRIPTO-1 is immunolocalized in extravillous cytotrophoblast in placenta creta. BioMed Res. Int. 2014, 2014, 892856. [Google Scholar] [CrossRef]
- Shafiei, S.; Dufort, D. Maternal Cripto is critical for proper development of the mouse placenta and the placental vasculature. Placenta 2021, 107, 13–23. [Google Scholar] [CrossRef]
- Miharada, K.; Karlsson, G.; Rehn, M.; Rorby, E.; Siva, K.; Cammenga, J.; Karlsson, S. Hematopoietic Stem Cells Viii. In Annals of the New York Academy of Sciences; Kanz, L., Fibbe, W.E., Lengerke, C., Dick, J.E., Eds.; Wiley: Hoboken, NJ, USA, 2012; Volume 1266, pp. 55–62. [Google Scholar]
- Papageorgiou, I.; Nicholls, P.K.; Wang, F.; Lackmann, M.; Makanji, Y.; Salamonsen, L.A.; Robertson, D.M.; Harrison, C.A. Expression of nodal signalling components in cycling human endometrium and in endometrial cancer. Reprod. Biol. Endocrinol. 2009, 7, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Karkampouna, S.; van der Helm, D.; van Hoek, B.; Verspaget, H.W.; Goumans, M.J.T.H.; Coenraad, M.; Kruithof, B.T.H.; de Kruithof, J.M. Oncofetal protein CRIPTO regulates wound healing and fibrogenesis in regenerating liver and is associated with the initial stages of cardiac fibrosis. bioRxiv 2021. [Google Scholar] [CrossRef]
- Zhang, Y.Q.; Cleary, M.M.; Si, Y.J.; Liu, G.X.; Eto, Y.; Kritzik, M.; Dabernat, S.; Kayali, A.G.; Sarvetnick, N. Inhibition of activin signaling induces pancreatic epithelial cell expansion and diminishes terminal differentiation of pancreatic beta-cells. Diabetes 2004, 53, 2024–2033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guardiola, O.; Lafuste, P.; Brunelli, S.; Iaconis, S.; Touvier, T.; Mourikis, P.; De Bock, K.; Lonardo, E.; Andolfi, G.; Bouche, A.; et al. Cripto regulates skeletal muscle regeneration and modulates satellite cell determination by antagonizing myostatin. Proc. Natl. Acad. Sci. USA 2012, 109, E3231–E3240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia de Vinuesa, A.; Sanchez-Duffhues, G.; Blaney-Davidson, E.; van Caam, A.; Lodder, K.; Ramos, Y.; Kloppenburg, M.; Meulenbelt, I.; van der Kraan, P.; Goumans, M.J.; et al. Cripto favors chondrocyte hypertrophy via TGF-beta SMAD1/5 signaling during development of osteoarthritis. J. Pathol. 2021. [Google Scholar] [CrossRef]
- Miharada, K.; Karlsson, G.; Rehn, M.; Rorby, E.; Siva, K.; Cammenga, J.; Karlsson, S. Cripto Regulates Hematopoietic Stem Cells as a Hypoxic-Niche-Related Factor through Cell Surface Receptor GRP78. Cell Stem Cell 2011, 9, 330–344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ciardiello, F.; Kim, N.; Saeki, T.; Dono, R.; Persico, M.G.; Plowman, G.D.; Garrigues, J.; Radke, S.; Todaro, G.J.; Salomon, D.S. Differential expression of epidermal growth factor-related proteins in human colorectal tumors. Proc. Natl. Acad. Sci. USA 1991, 88, 7792–7796. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.H.; Wei, W.; Xu, J.; Guo, Z.X.; Xiao, C.Z.; Zhang, Y.F.; Jian, P.E.; Wu, X.L.; Shi, M.; Guo, R.P. Elevated expression of Cripto-1 correlates with poor prognosis in hepatocellular carcinoma. Oncotarget 2015, 6, 35116–35128. [Google Scholar] [CrossRef] [Green Version]
- Spike, B.T.; Kelber, J.A.; Booker, E.; Kalathur, M.; Rodewald, R.; Lipianskaya, J.; La, J.; He, M.; Wright, T.; Klemke, R.; et al. CRIPTO/GRP78 signaling maintains fetal and adult mammary stem cells ex vivo. Stem Cell Rep. 2014, 2, 427–439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herrington, E.E.; Ram, T.G.; Salomon, D.S.; Johnson, G.R.; Gullick, W.J.; Kenney, N.; Hosick, H.L. Expression of epidermal growth factor-related proteins in the aged adult mouse mammary gland and their relationship to tumorigenesis. J. Cell Physiol. 1997, 170, 47–56. [Google Scholar] [CrossRef]
- Kenney, N.J.; Huang, R.P.; Johnson, G.R.; Wu, J.X.; Okamura, D.; Matheny, W.; Kordon, E.; Gullick, W.J.; Plowman, G.; Smith, G.H.; et al. Detection and location of amphiregulin and Cripto-1 expression in the developing postnatal mouse mammary gland. Mol. Reprod. Dev. 1995, 41, 277–286. [Google Scholar] [CrossRef]
- Panico, L.; D’Antonio, A.; Salvatore, G.; Mezza, E.; Tortora, G.; De Laurentiis, M.; De Placido, S.; Giordano, T.; Merino, M.; Salomon, D.S.; et al. Differential immunohistochemical detection of transforming growth factor alpha, amphiregulin and CRIPTO in human normal and malignant breast tissues. Int. J. Cancer 1996, 65, 51–56. [Google Scholar] [CrossRef]
- Jin, J.Z.; Tan, M.; Ding, J. Analysis of Cripto expression during mouse cardiac myocyte differentiation. Int. J. Dev. Biol. 2013, 57, 793–797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iavarone, F.; Guardiola, O.; Scagliola, A.; Andolfi, G.; Esposito, F.; Serrano, A.; Perdiguero, E.; Brunelli, S.; Munoz-Canoves, P.; Minchiotti, G. Cripto shapes macrophage plasticity and restricts EndMT in injured and diseased skeletal muscle. EMBO Rep. 2020, 21, e49075. [Google Scholar] [CrossRef] [PubMed]
- Stephens, E.B.; Jackson, M.; Cui, L.; Pacyniak, E.; Choudhuri, R.; Liverman, C.S.; Salomon, D.S.; Berman, N.E. Early dysregulation of cripto-1 and immunomodulatory genes in the cerebral cortex in a macaque model of neuroAIDS. Neurosci. Lett. 2006, 410, 94–99. [Google Scholar] [CrossRef] [PubMed]
- Pilgaard, L.; Mortensen, J.H.; Henriksen, M.; Olesen, P.; Sorensen, P.; Laursen, R.; Vyberg, M.; Agger, R.; Zachar, V.; Moos, T.; et al. Cripto-1 Expression in Glioblastoma Multiforme. Brain Pathol. 2014, 24, 360–370. [Google Scholar] [CrossRef]
- Friess, H.; Yamanaka, Y.; Buchler, M.; Kobrin, M.S.; Tahara, E.; Korc, M. Cripto, a member of the epidermal growth factor family, is over-expressed in human pancreatic cancer and chronic pancreatitis. Int. J. Cancer 1994, 56, 668–674. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wang, J.; Yang, T.; Liu, R.; Xu, Y. Overexpression levels of cripto-1 predict poor prognosis in patients with prostate cancer following radical prostatectomy. Oncol. Lett. 2019, 18, 2584–2591. [Google Scholar] [CrossRef] [Green Version]
- Terry, S.; El-Sayed, I.Y.; Destouches, D.; Maille, P.; Nicolaiew, N.; Ploussard, G.; Semprez, F.; Pimpie, C.; Beltran, H.; Londono-Vallejo, A.; et al. CRIPTO overexpression promotes mesenchymal differentiation in prostate carcinoma cells through parallel regulation of AKT and FGFR activities. Oncotarget 2015, 6, 11994–12008. [Google Scholar] [CrossRef] [Green Version]
- Zoni, E.; Chen, L.; Karkampouna, S.; Granchi, Z.; Verhoef, E.I.; La Manna, F.; Kelber, J.; Pelger, R.C.M.; Henry, M.D.; Snaar-Jagalska, E.; et al. CRIPTO and Its signaling partner GRP78 drive the metastatic phenotype in human osteotropic prostate cancer. Oncogene 2017, 36, 4739–4749. [Google Scholar] [CrossRef] [Green Version]
- Shukla, A.; Ho, Y.; Liu, X.; Ryscavage, A.; Glick, A.B. Cripto-1 alters keratinocyte differentiation via blockade of transforming growth factor-beta1 signaling: Role in skin carcinogenesis. Mol. Cancer Res. 2008, 6, 509–516. [Google Scholar] [CrossRef] [Green Version]
- De Luca, A.; Lamura, L.; Strizzi, L.; Roma, C.; D’Antonio, A.; Margaryan, N.; Pirozzi, G.; Hsu, M.Y.; Botti, G.; Mari, E.; et al. Expression and functional role of CRIPTO-1 in cutaneous melanoma. Br. J. Cancer 2011, 105, 1030–1038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xue, Y.-J.; Chen, S.-N.; Chen, W.-G.; Wu, G.-Q.; Liao, Y.-F.; Xu, J.-B.; Tang, H.; Yang, S.-H.; He, S.-Y.; Luo, Y.-F.; et al. Cripto-1 expression in patients with clear cell renal cell carcinoma is associated with poor disease outcome. J. Exp. Clin. Cancer Res. 2019, 38, 378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saeki, T.; Stromberg, K.; Qi, C.F.; Gullick, W.J.; Tahara, E.; Normanno, N.; Ciardiello, F.; Kenney, N.; Johnson, G.R.; Salomon, D.S. Differential immunohistochemical detection of amphiregulin and cripto in human normal colon and colorectal tumors. Cancer Res 1992, 52, 3467–3473. [Google Scholar] [PubMed]
- Saeki, T.; Salomon, D.; Gullick, W.; Mandai, K.; Yamagami, K.; Moriwaki, S.; Takashima, S.; Nishikawa, Y.; Tahara, E. Expression of cripto-1 in human colorectal adenomas and carcinomas is related to the degree of dysplasia. Int. J. Oncol. 1994, 5, 445–451. [Google Scholar] [CrossRef] [PubMed]
- Sato, J.; Karasawa, H.; Suzuki, T.; Nakayama, S.; Katagiri, M.; Maeda, S.; Ohnuma, S.; Motoi, F.; Naitoh, T.; Unno, M. The Function and Prognostic Significance of Cripto-1 in Colorectal Cancer. Cancer Investig. 2020, 38, 214–227. [Google Scholar] [CrossRef] [PubMed]
- Yoon, H.J.; Hong, J.S.; Shin, W.J.; Lee, Y.J.; Hong, K.O.; Lee, J.I.; Hong, S.P.; Hong, S.D. The role of Cripto-1 in the tumorigenesis and progression of oral squamous cell carcinoma. Oral Oncol. 2011, 47, 1023–1031. [Google Scholar] [CrossRef]
- Ertoy, D.; Ayhan, A.; Sarac, E.; Karaagaoglu, E.; Yasui, W.; Tahara, E.; Ayhan, A. Clinicopathological implication of cripto expression in early stage invasive cervical carcinomas. Eur. J. Cancer 2000, 36, 1002–1007. [Google Scholar] [CrossRef]
- Zhang, J.-G.; Zhao, J.; Xin, Y. Significance and relationship between Cripto-1 and p-STAT3 expression in gastric cancer and precancerous lesions. World J. Gastroenterol. 2010, 16, 571–577. [Google Scholar] [CrossRef] [PubMed]
- Saeki, T.; Salomon, D.; Normanno, N.; Johnson, G.; Gullick, W.; Mandai, K.; Moriwaki, S.; Takashima, S.; Kuniyasu, M.; Tahara, E.; et al. Immunohistochemical detection of cripto-1, amphiregulin and transforming growth-factor-alpha in human gastric carcinomas and intestinal metaplasias. Int. J. Oncol. 1994, 5, 215–223. [Google Scholar] [CrossRef]
- Wei, B.; Jin, W.; Ruan, J.; Xu, Z.; Zhou, Y.; Liang, J.; Cheng, H.; Jin, K.; Huang, X.; Lu, P.; et al. Cripto-1 expression and its prognostic value in human bladder cancer patients. Tumor Biol. 2015, 36, 1105–1113. [Google Scholar] [CrossRef]
- Xu, C.; Yuan, Q.; Hu, H.; Wang, W.; Zhang, Q.; Li, L.; Wang, J.; Yang, R. Expression of Cripto-1 predicts poor prognosis in stage I non-small cell lung cancer. J. Cell. Mol. Med. 2020, 24, 9705–9711. [Google Scholar] [CrossRef]
- Huang, C.; Chen, W.; Wang, X.; Zhao, J.; Li, Q.; Fu, Z. Cripto-1 Promotes the Epithelial-Mesenchymal Transition in Esophageal Squamous Cell Carcinoma Cells. Evid. Based Complement. Altern. Med. 2015, 2015, 421285. [Google Scholar] [CrossRef]
- Mahmoudian, R.A.; Abbaszadegan, M.R.; Forghanifard, M.M.; Moghbeli, M.; Moghbeli, F.; Chamani, J.; Gholamin, M. Biological and Clinicopathological Significance of Cripto-1 Expression in the Progression of Human ESCC. Rep. Biochem. Mol. Biol. 2017, 5, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Van der Flier, L.G.; Sabates-Bellver, J.; Oving, I.; Haegebarth, A.; De Palo, M.; Anti, M.; Van Gijn, M.E.; Suijkerbuijk, S.; Van de Wetering, M.; Marra, G.; et al. The Intestinal Wnt/TCF Signature. Gastroenterology 2007, 132, 628–632. [Google Scholar] [CrossRef]
- Francescangeli, F.; Contavalli, P.; De Angelis, M.L.; Baiocchi, M.; Gambara, G.; Pagliuca, A.; Fiorenzano, A.; Prezioso, C.; Boe, A.; Todaro, M.; et al. Dynamic regulation of the cancer stem cell compartment by Cripto-1 in colorectal cancer. Cell Death Differ. 2015, 22, 1700–1713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niemeyer, C.C.; Persico, M.G.; Adamson, E.D. Cripto: Roles in mammary cell growth, survival, differentiation and transformation. Cell Death Differ. 1998, 5, 440–449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klauzinska, M.; McCurdy, D.; Rangel, M.C.; Vaidyanath, A.; Castro, N.P.; Shen, M.M.; Gonzales, M.; Bertolette, D.; Bianco, C.; Callahan, R.; et al. Cripto-1 ablation disrupts alveolar development in the mouse mammary gland through a progesterone receptor-mediated pathway. Am. J. Pathol. 2015, 185, 2907–2922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Y.; Strizzi, L.; Raafat, A.; Hirota, M.; Bianco, C.; Feigenbaum, L.; Kenney, N.; Wechselberger, C.; Callahan, R.; Salomon, D.S. Overexpression of human Cripto-1 in transgenic mice delays mammary gland development and differentiation and induces mammary tumorigenesis. Am. J. Pathol. 2005, 167, 585–597. [Google Scholar] [CrossRef] [Green Version]
- Wechselberger, C.; Ebert, A.D.; Bianco, C.; Khan, N.I.; Sun, Y.P.; Wallace-Jones, B.; Montesano, R.; Salomon, D.S. Cripto-1 enhances migration and branching morphogenesis of mouse mammary epithelial cells. Exp. Cell Res. 2001, 266, 95–105. [Google Scholar] [CrossRef]
- Thomas, P.A.; Kirschmann, D.A.; Cerhan, J.R.; Folberg, R.; Seftor, E.A.; Sellers, T.A.; Hendrix, M.J. Association between keratin and vimentin expression, malignant phenotype, and survival in postmenopausal breast cancer patients. Clin. Cancer Res. 1999, 5, 2698–2703. [Google Scholar]
- Loh, K.M.; Lim, B. A precarious balance: Pluripotency factors as lineage specifiers. Cell Stem Cell 2011, 8, 363–369. [Google Scholar] [CrossRef] [Green Version]
- Spike, B.T.; Engle, D.D.; Lin, J.C.; Cheung, S.K.; La, J.; Wahl, G.M. A mammary stem cell population identified and characterized in late embryogenesis reveals similarities to human breast cancer. Cell Stem Cell 2012, 10, 183–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garland, M.A.; Sengupta, S.; Mathew, L.K.; Truong, L.; de Jong, E.; Piersma, A.H.; La Du, J.; Tanguay, R.L. Glucocorticoid receptor-dependent induction of cripto-1 (one-eyed pinhead) inhibits zebrafish caudal fin regeneration. Toxicol. Rep. 2019, 6, 529–537. [Google Scholar] [CrossRef]
- Prezioso, C.; Iaconis, S.; Andolfi, G.; Zentilin, L.; Iavarone, F.; Guardiola, O.; Minchiotti, G. Conditional Cripto overexpression in satellite cells promotes myogenic commitment and enhances early regeneration. Front. Cell Dev. Biol. 2015, 3, 31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoover, M. Identification of a Novel Cripto/Myosin II Interaction that Promotes Stem Cell Mediated Tissue Regeneration. Ph.D. Thesis, California State University, Long Beach, CA, USA, 2017. [Google Scholar]
- Hoover, M.; Runa, F.; Booker, E.; Diedrich, J.K.; Duell, E.; Williams, B.; Arellano-Garcia, C.; Uhlendorf, T.; La Kim, S.; Fischer, W.; et al. Identification of myosin II as a cripto binding protein and regulator of cripto function in stem cells and tissue regeneration. Biochem. Biophys. Res. Commun. 2019, 509, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Lonardo, E.; Parish, C.L.; Ponticelli, S.; Marasco, D.; Ribeiro, D.; Ruvo, M.; De Falco, S.; Arenas, E.; Minchiotti, G. A small synthetic cripto blocking Peptide improves neural induction, dopaminergic differentiation, and functional integration of mouse embryonic stem cells in a rat model of Parkinson’s disease. Stem Cells 2010, 28, 1326–1337. [Google Scholar] [CrossRef] [PubMed]
- Parish, C.L.; Parisi, S.; Persico, M.G.; Arenas, E.; Minchiotti, G. Cripto as a Target for Improving Embryonic Stem Cell–Based Therapy in Parkinson’s Disease. Stem Cells 2005, 23, 471–476. [Google Scholar] [CrossRef]
- Kim, S.; Yoon, Y.M.; Han, Y.S.; Lee, J.H.; Hur, J.; Lee, S.H. Administration of Cripto in GRP78 overexpressed human MSCs enhances stem cell viability and angiogenesis during human MSC transplantation therapy. Cell Prolif. 2018, 51, e12463. [Google Scholar] [CrossRef]
- Bianco, C.; Cotten, C.; Lonardo, E.; Strizzi, L.; Baraty, C.; Mancino, M.; Gonzales, M.; Watanabe, K.; Nagaoka, T.; Berry, C.; et al. Cripto-1 Is Required for Hypoxia to Induce Cardiac Differentiation of Mouse Embryonic Stem Cells. Am. J. Pathol. 2009, 175, 2146–2158. [Google Scholar] [CrossRef] [Green Version]
- Klauzinska, M.; Bertolette, D.; Tippireddy, S.; Strizzi, L.; Gray, P.C.; Gonzales, M.; Duroux, M.; Ruvo, M.; Wechselberger, C.; Castro, N.P.; et al. Cripto-1: An extracellular protein—Connecting the sequestered biological dots. Connect. Tissue Res. 2015, 56, 364–380. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.-M.; Bao, Y.-L.; Yu, C.-L.; Wang, Y.-M.; Song, Z.-B. Cripto-1 modulates macrophage cytokine secretion and phagocytic activity via NF-κB signaling. Immunol. Res. 2016, 64, 104–114. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.; Guo, Y.-Z.; Yue, X.; Zhang, G.-P.; Zhang, Y.; Kuang, M.; Peng, B.-G.; Li, S.-Q. Cripto-1 promotes tumor invasion and predicts poor outcomes in hepatocellular carcinoma. Carcinogenesis 2020, 41, 571–581. [Google Scholar] [CrossRef] [PubMed]
- Goumans, M.J.; Ten Dijke, P. TGF-beta Signaling in Control of Cardiovascular Function. Cold Spring Harb. Perspect. Biol. 2018, 10, a022210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pahnke, A.; Conant, G.; Huyer, L.D.; Zhao, Y.; Feric, N.; Radisic, M. The role of Wnt regulation in heart development, cardiac repair and disease: A tissue engineering perspective. Biochem. Biophys. Res. Commun. 2016, 473, 698–703. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Greene, S.B.; Bonilla-Claudio, M.; Tao, Y.; Zhang, J.; Bai, Y.; Huang, Z.; Black, B.L.; Wang, F.; Martin, J.F. Bmp signaling regulates myocardial differentiation from cardiac progenitors through a MicroRNA-mediated mechanism. Dev. Cell 2010, 19, 903–912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parisi, S.; D’Andrea, D.; Lago, C.T.; Adamson, E.D.; Persico, M.G.; Minchiotti, G. Nodal-dependent Cripto signaling promotes cardiomyogenesis and redirects the neural fate of embryonic stem cells. J. Cell Biol. 2003, 163, 303–314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Aniello, C.; Lonardo, E.; Iaconis, S.; Guardiola, O.; Liguoro, A.M.; Liguori, G.L.; Autiero, M.; Carmeliet, P.; Minchiotti, G. G Protein-Coupled Receptor APJ and Its Ligand Apelin Act Downstream of Cripto to Specify Embryonic Stem Cells Toward the Cardiac Lineage Through Extracellular Signal-Regulated Kinase/p70S6 Kinase Signaling Pathway. Circ. Res. 2009, 105, 231–238. [Google Scholar] [CrossRef] [Green Version]
- Eyries, M.; Siegfried, G.; Ciumas, M.; Montagne, K.; Agrapart, M.; Lebrin, F.; Soubrier, F. Hypoxia-induced apelin expression regulates endothelial cell proliferation and regenerative angiogenesis. Circ. Res. 2008, 103, 432–440. [Google Scholar] [CrossRef] [Green Version]
- Kirabo, A.; Ryzhov, S.; Gupte, M.; Sengsayadeth, S.; Gumina, R.J.; Sawyer, D.B.; Galindo, C.L. Neuregulin-1 beta induces proliferation, survival and paracrine signaling in normal human cardiac ventricular fibroblasts. J. Mol. Cell. Cardiol. 2017, 105, 59–69. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Xu, H.; Chi, X.; Fan, Y.; Shi, Y.; Niu, J. High level of serum Cripto-1 in hepatocellular carcinoma, especially with hepatitis B virus infection. Medicine 2018, 97, e11781. [Google Scholar] [CrossRef] [PubMed]
- Karkampouna, S.; Helm, D.V.D.; Gray, P.C.; Chen, L.; Klima, I.; Grosjean, J.; Burgmans, M.C.; Farina-Sarasqueta, A.; Snaar-Jagalska, E.B.; Stroka, D.M.; et al. CRIPTO promotes an aggressive tumour phenotype and resistance to treatment in hepatocellular carcinoma. J. Pathol. 2018, 245, 297–310. [Google Scholar] [CrossRef]
- Lo, R.C.-L.; Leung, C.O.-N.; Chan, K.K.-S.; Ho, D.W.-H.; Wong, C.-M.; Lee, T.K.-W.; Ng, I.O.-L. Cripto-1 contributes to stemness in hepatocellular carcinoma by stabilizing Dishevelled-3 and activating Wnt/β-catenin pathway. Cell Death Differ. 2018, 25, 1426–1441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, C.; Sun, L.; Jiang, K.; Gao, D.-M.; Kang, X.-N.; Wang, C.; Zhang, S.; Huang, S.; Qin, X.; Li, Y.; et al. NANOG promotes liver cancer cell invasion by inducing epithelial–mesenchymal transition through NODAL/SMAD3 signaling pathway. Int. J. Biochem. Cell Biol. 2013, 45, 1099–1108. [Google Scholar] [CrossRef] [PubMed]
- Wynn, T.A. Cellular and molecular mechanisms of fibrosis. J. Pathol. 2008, 214, 199–210. [Google Scholar] [CrossRef] [Green Version]
- Kelber, J.A.; Panopoulos, A.D.; Shani, G.; Booker, E.C.; Belmonte, J.C.; Vale, W.W.; Gray, P.C. Blockade of Cripto binding to cell surface GRP78 inhibits oncogenic Cripto signaling via MAPK/PI3K and Smad2/3 pathways. Oncogene 2009, 28, 2324–2336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castro, N.P.; Fedorova-Abrams, N.D.; Merchant, A.S.; Rangel, M.C.; Nagaoka, T.; Karasawa, H.; Klauzinska, M.; Hewitt, S.M.; Biswas, K.; Sharan, S.K.; et al. Cripto-1 as a novel therapeutic target for triple negative breast cancer. Oncotarget 2015, 6, 11910–11929. [Google Scholar] [CrossRef] [Green Version]
- Bianco, C.; Strizzi, L.; Mancino, M.; Rehman, A.; Hamada, S.; Watanabe, K.; De Luca, A.; Jones, B.; Balogh, G.; Russo, J.; et al. Identification of cripto-1 as a novel serologic marker for breast and colon cancer. Clin. Cancer Res. 2006, 12, 5158–5164. [Google Scholar] [CrossRef] [Green Version]
- Xu, C.H.; Wang, Y.; Qian, L.H.; Yu, L.K.; Zhang, X.W.; Wang, Q.B. Serum Cripto-1 is a novel biomarker for non-small cell lung cancer diagnosis and prognosis. Clin. Respir. J. 2017, 11, 765–771. [Google Scholar] [CrossRef]
- Wechselberger, C.; Strizzi, L.; Kenney, N.; Hirota, M.; Sun, Y.; Ebert, A.; Orozco, O.; Bianco, C.; Khan, N.I.; Wallace-Jones, B.; et al. Human Cripto-1 overexpression in the mouse mammary gland results in the development of hyperplasia and adenocarcinoma. Oncogene 2005, 24, 4094–4105. [Google Scholar] [CrossRef] [Green Version]
- Sun, C.; Orozco, O.; Olson, D.L.; Choi, E.; Garber, E.; Tizard, R.; Szak, S.; Sanicola, M.; Carulli, J.P. CRIPTO3, a presumed pseudogene, is expressed in cancer. Biochem. Biophys. Res. Commun. 2008, 377, 215–220. [Google Scholar] [CrossRef]
- Hentschke, M.; Kurth, I.; Borgmeyer, U.; Hubner, C.A. Germ cell nuclear factor is a repressor of CRIPTO-1 and CRIPTO-3. J. Biol. Chem. 2006, 281, 33497–33504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, W.; He, X.; Hoadley, K.A.; Parker, J.S.; Hayes, D.N.; Perou, C.M. Comparison of RNA-Seq by poly (A) capture, ribosomal RNA depletion, and DNA microarray for expression profiling. BMC Genom. 2014, 15, 419. [Google Scholar] [CrossRef] [Green Version]
- Baldassarre, G.; Tucci, M.; Lembo, G.; Pacifico, F.M.; Dono, R.; Lago, C.T.; Barra, A.; Bianco, C.; Viglietto, G.; Salomon, D.; et al. A Truncated Form of Teratocarcinoma-Derived Growth Factor-1 (Cripto-1) mRNA Expressed in Human Colon Carcinoma Cell Lines and Tumors. Tumor Biol. 2001, 22, 286–293. [Google Scholar] [CrossRef] [PubMed]
- Clevers, H. The cancer stem cell: Premises, promises and challenges. Nat. Med. 2011, 17, 313–319. [Google Scholar] [CrossRef] [PubMed]
- Batlle, E.; Clevers, H. Cancer stem cells revisited. Nat. Med. 2017, 23, 1124–1134. [Google Scholar] [CrossRef]
- Quintana, E.; Shackleton, M.; Sabel, M.S.; Fullen, D.R.; Johnson, T.M.; Morrison, S.J. Efficient tumour formation by single human melanoma cells. Nature 2008, 456, 593–598. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, K.; Meyer, M.J.; Strizzi, L.; Lee, J.M.; Gonzales, M.; Bianco, C.; Nagaoka, T.; Farid, S.S.; Margaryan, N.; Hendrix, M.J.; et al. Cripto-1 is a cell surface marker for a tumorigenic, undifferentiated subpopulation in human embryonal carcinoma cells. Stem Cells 2010, 28, 1303–1314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loying, P.; Manhas, J.; Sen, S.; Bose, B. Autoregulation and heterogeneity in expression of human Cripto-1. PLoS ONE 2015, 10, e0116748. [Google Scholar] [CrossRef] [Green Version]
- Cocciadiferro, L.; Miceli, V.; Kang, K.S.; Polito, L.M.; Trosko, J.E.; Carruba, G. Profiling cancer stem cells in androgen-responsive and refractory human prostate tumor cell lines. Ann. N. Y. Acad. Sci. 2009, 1155, 257–262. [Google Scholar] [CrossRef] [PubMed]
- Strizzi, L.; Mancino, M.; Bianco, C.; Raafat, A.; Gonzales, M.; Booth, B.W.; Watanabe, K.; Nagaoka, T.; Mack, D.L.; Howard, B.; et al. Netrin-1 can affect morphogenesis and differentiation of the mouse mammary gland. J. Cell Physiol. 2008, 216, 824–834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.; Boleslaw Olszewski, M.; Kruithof-de Julio, M.; Snaar-Jagalska, B.E. Zebrafish Microenvironment Elevates EMT and CSC-Like Phenotype of Engrafted Prostate Cancer Cells. Cells 2020, 9, 797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alam, M.J.; Takahashi, R.; Afify, S.M.; Oo, A.K.K.; Kumon, K.; Nawara, H.M.; Khayrani, A.C.; Du, J.; Zahra, M.H.; Seno, A.; et al. Exogenous Cripto-1 Suppresses Self-Renewal of Cancer Stem Cell Model. Int. J. Mol. Sci. 2018, 19, 3345. [Google Scholar] [CrossRef] [Green Version]
- Balcioglu, O.; Heinz, R.E.; Freeman, D.W.; Gates, B.L.; Hagos, B.M.; Booker, E.; Mirzaei Mehrabad, E.; Diesen, H.T.; Bhakta, K.; Ranganathan, S.; et al. CRIPTO antagonist ALK4(L75A)-Fc inhibits breast cancer cell plasticity and adaptation to stress. Breast Cancer Res. 2020, 22, 125. [Google Scholar] [CrossRef]
- Tsai, Y.L.; Ha, D.P.; Zhao, H.; Carlos, A.J.; Wei, S.; Pun, T.K.; Wu, K.; Zandi, E.; Kelly, K.; Lee, A.S. Endoplasmic reticulum stress activates SRC, relocating chaperones to the cell surface where GRP78/CD109 blocks TGF-beta signaling. Proc. Natl. Acad. Sci. USA 2018, 115, E4245–E4254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, A.S. GRP78 induction in cancer: Therapeutic and prognostic implications. Cancer Res. 2007, 67, 3496–3499. [Google Scholar] [CrossRef] [Green Version]
- Wahl, G.M.; Spike, B.T. Cell state plasticity, stem cells, EMT, and the generation of intra-tumoral heterogeneity. NPJ Breast Cancer 2017, 3, 14. [Google Scholar] [CrossRef] [Green Version]
- Bianco, C.; Strizzi, L.; Ebert, A.; Chang, C.; Rehman, A.; Normanno, N.; Guedez, L.; Salloum, R.; Ginsburg, E.; Sun, Y.; et al. Role of Human Cripto-1 in Tumor Angiogenesis. JNCI J. Natl. Cancer Inst. 2005, 97, 132–141. [Google Scholar] [CrossRef] [Green Version]
- Alowaidi, F.; Hashimi, S.M.; Alqurashi, N.; Wood, S.A.; Wei, M.Q. Cripto-1 overexpression in U87 glioblastoma cells activates MAPK, focal adhesion and ErbB pathways. Oncol. Lett. 2019, 18, 3399–3406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, D.; Shi, Z.; Xu, H.; Chen, R.; Xue, S.; Sun, X. Knockdown of Cripto-1 inhibits the proliferation, migration, invasion, and angiogenesis in prostate carcinoma cells. J. Biosci. 2017, 42, 405–416. [Google Scholar] [CrossRef]
- Corti, F.; Simons, M. Modulation of VEGF receptor 2 signaling by protein phosphatases. Pharm. Res 2017, 115, 107–123. [Google Scholar] [CrossRef] [Green Version]
- Dvorak, H.F. Tumors: Wounds that do not heal. Similarities between tumor stroma generation and wound healing. N. Engl. J. Med. 1986, 315, 1650–1659. [Google Scholar] [CrossRef]
- Dvorak, H.F. Tumors: Wounds that do not heal-redux. Cancer Immunol. Res. 2015, 3, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Yu, L.X.; Ling, Y.; Wang, H.Y. Role of nonresolving inflammation in hepatocellular carcinoma development and progression. NPJ Precis. Oncol. 2018, 2, 6. [Google Scholar] [CrossRef] [Green Version]
- Visser, L.L.; Elshof, L.E.; Schaapveld, M.; van de Vijver, K.; Groen, E.J.; Almekinders, M.M.; Bierman, C.; van Leeuwen, F.E.; Rutgers, E.J.; Schmidt, M.K.; et al. Clinicopathological Risk Factors for an Invasive Breast Cancer Recurrence after Ductal Carcinoma in Situ-A Nested Case-Control Study. Clin. Cancer Res. 2018, 24, 3593–3601. [Google Scholar] [CrossRef] [Green Version]
- Moretti, S.; Spallanzani, A.; Pinzi, C.; Prignano, F.; Fabbri, P. Fibrosis in regressing melanoma versus nonfibrosis in halo nevus upon melanocyte disappearance: Could it be related to a different cytokine microenvironment? J. Cutan. Pathol. 2007, 34, 301–308. [Google Scholar] [CrossRef]
- Paszek, M.J.; Weaver, V.M. The tension mounts: Mechanics meets morphogenesis and malignancy. J. Mammary Gland Biol. Neoplasia 2004, 9, 325–342. [Google Scholar] [CrossRef] [PubMed]
- Acerbi, I.; Cassereau, L.; Dean, I.; Shi, Q.; Au, A.; Park, C.; Chen, Y.Y.; Liphardt, J.; Hwang, E.S.; Weaver, V.M. Human breast cancer invasion and aggression correlates with ECM stiffening and immune cell infiltration. Integr. Biol. 2015, 7, 1120–1134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drifka, C.R.; Loeffler, A.G.; Mathewson, K.; Keikhosravi, A.; Eickhoff, J.C.; Liu, Y.; Weber, S.M.; Kao, W.J.; Eliceiri, K.W. Highly aligned stromal collagen is a negative prognostic factor following pancreatic ductal adenocarcinoma resection. Oncotarget 2016, 7, 76197–76213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayward, S.W.; Wang, Y.; Cao, M.; Hom, Y.K.; Zhang, B.; Grossfeld, G.D.; Sudilovsky, D.; Cunha, G.R. Malignant transformation in a nontumorigenic human prostatic epithelial cell line. Cancer Res. 2001, 61, 8135–8142. [Google Scholar] [PubMed]
- Li, Y.; Wang, R.; Xiong, S.; Wang, X.; Zhao, Z.; Bai, S.; Wang, Y.; Zhao, Y.; Cheng, B. Cancer-associated fibroblasts promote the stemness of CD24(+) liver cells via paracrine signaling. J. Mol. Med. 2019, 97, 243–255. [Google Scholar] [CrossRef] [PubMed]
- Cruz-Bermudez, A.; Laza-Briviesca, R.; Vicente-Blanco, R.J.; Garcia-Grande, A.; Coronado, M.J.; Laine-Menendez, S.; Alfaro, C.; Sanchez, J.C.; Franco, F.; Calvo, V.; et al. Cancer-associated fibroblasts modify lung cancer metabolism involving ROS and TGF-beta signaling. Free Radic. Biol. Med. 2019, 130, 163–173. [Google Scholar] [CrossRef]
- Vonlaufen, A.; Phillips, P.A.; Xu, Z.; Goldstein, D.; Pirola, R.C.; Wilson, J.S.; Apte, M.V. Pancreatic stellate cells and pancreatic cancer cells: An unholy alliance. Cancer Res. 2008, 68, 7707–7710. [Google Scholar] [CrossRef] [Green Version]
- Lonardo, E.; Frias-Aldeguer, J.; Hermann, P.C.; Heeschen, C. Pancreatic stellate cells form a niche for cancer stem cells and promote their self-renewal and invasiveness. Cell Cycle 2012, 11, 1282–1290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piersma, B.; Hayward, M.K.; Weaver, V.M. Fibrosis and cancer: A strained relationship. Biochim. Biophys. Acta Rev. Cancer 2020, 1873, 188356. [Google Scholar] [CrossRef] [PubMed]
- Costa, A.; Kieffer, Y.; Scholer-Dahirel, A.; Pelon, F.; Bourachot, B.; Cardon, M.; Sirven, P.; Magagna, I.; Fuhrmann, L.; Bernard, C.; et al. Fibroblast Heterogeneity and Immunosuppressive Environment in Human Breast Cancer. Cancer Cell 2018, 33, 463–479. [Google Scholar] [CrossRef] [Green Version]
- Shani, G.; Fischer, W.H.; Justice, N.J.; Kelber, J.A.; Vale, W.; Gray, P.C. GRP78 and Cripto form a complex at the cell surface and collaborate to inhibit transforming growth factor beta signaling and enhance cell growth. Mol. Cell Biol. 2008, 28, 666–677. [Google Scholar] [CrossRef] [Green Version]
- Morkel, M.; Huelsken, J.; Wakamiya, M.; Ding, J.; van de Wetering, M.; Clevers, H.; Taketo, M.M.; Behringer, R.R.; Shen, M.M.; Birchmeier, W. Beta-catenin regulates Cripto- and Wnt3-dependent gene expression programs in mouse axis and mesoderm formation. Development 2003, 130, 6283–6294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamada, S.; Watanabe, K.; Hirota, M.; Bianco, C.; Strizzi, L.; Mancino, M.; Gonzales, M.; Salomon, D.S. beta-Catenin/TCF/LEF regulate expression of the short form human Cripto-1. Biochem. Biophys. Res. Commun. 2007, 355, 240–244. [Google Scholar] [CrossRef] [Green Version]
- Nagaoka, T.; Karasawa, H.; Turbyville, T.; Rangel, M.C.; Castro, N.P.; Gonzales, M.; Baker, A.; Seno, M.; Lockett, S.; Greer, Y.E.; et al. Cripto-1 enhances the canonical Wnt/beta-catenin signaling pathway by binding to LRP5 and LRP6 co-receptors. Cell Signal. 2013, 25, 178–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Qin, Z.B.; Yang, K.; Liu, R.L.; Xu, Y. Cripto-1 promotes epithelial-mesenchymal transition in prostate cancer via Wnt/beta-catenin signaling. Oncol. Rep. 2017, 37, 1521–1528. [Google Scholar] [CrossRef] [Green Version]
- Bianco, C.; Castro, N.P.; Baraty, C.; Rollman, K.; Held, N.; Rangel, M.C.; Karasawa, H.; Gonzales, M.; Strizzi, L.; Salomon, D.S. Regulation of human Cripto-1 expression by nuclear receptors and DNA promoter methylation in human embryonal and breast cancer cells. J. Cell Physiol. 2013, 228, 1174–1188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Humphreys, R.C.; Lydon, J.; O’Malley, B.W.; Rosen, J.M. Mammary gland development is mediated by both stromal and epithelial progesterone receptors. Mol. Endocrinol. 1997, 11, 801–811. [Google Scholar] [CrossRef]
- Axlund, S.D.; Sartorius, C.A. Progesterone regulation of stem and progenitor cells in normal and malignant breast. Mol. Cell Endocrinol. 2012, 357, 71–79. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.J.; Chapman-Davis, E. Role of progesterone in endometrial cancer. Semin. Reprod. Med. 2010, 28, 81–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lange, C.A.; Sartorius, C.A.; Abdel-Hafiz, H.; Spillman, M.A.; Horwitz, K.B.; Jacobsen, B.M. Progesterone receptor action: Translating studies in breast cancer models to clinical insights. Adv. Exp. Med. Biol. 2008, 630, 94–111. [Google Scholar]
- Lawrence, M.G.; Margaryan, N.V.; Loessner, D.; Collins, A.; Kerr, K.M.; Turner, M.; Seftor, E.A.; Stephens, C.R.; Lai, J.; Postovit, L.-M.; et al. Reactivation of embryonic nodal signaling is associated with tumor progression and promotes the growth of prostate cancer cells. Prostate 2011, 71, 1198–1209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kach, J.; Conzen, S.D.; Szmulewitz, R.Z. Targeting the glucocorticoid receptor in breast and prostate cancers. Sci. Transl. Med. 2015, 7, 305ps319. [Google Scholar] [CrossRef] [Green Version]
- Zvibel, I.; Brill, S.; Halpern, Z.; Papa, M. Hepatocyte extracellular matrix modulates expression of growth factors and growth factor receptors in human colon cancer cells. Exp. Cell Res. 1998, 245, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Zvibel, I.; Halpern, Z.; Papa, M. Extracellular matrix modulates expression of growth factors and growth-factor receptors in liver-colonizing colon-cancer cell lines. Int. J. Cancer 1998, 77, 295–301. [Google Scholar] [CrossRef]
- Zvibel, I.; Brill, S.; Halpern, Z.; Papa, M. Amphiregulin and hepatocyte-derived extracellular matrix regulate proliferation and autocrine growth factor expression in colon cancer cell lines of varying liver-colonizing capability. J. Cell. Biochem. 2000, 76, 332–340. [Google Scholar] [CrossRef]
- Strizzi, L.; Bianco, C.; Normanno, N.; Seno, M.; Wechselberger, C.; Wallace-Jones, B.; Khan, N.I.; Hirota, M.; Sun, Y.P.; Sanicola, M.; et al. Epithelial mesenchymal transition is a characteristic of hyperplasias and tumors in mammary gland from MMTV-cripto-1 transgenic mice. J. Cell. Physiol. 2004, 201, 266–276. [Google Scholar] [CrossRef]
- Bianco, C.; Strizzi, L.; Rehman, A.; Normanno, N.; Wechselberger, C.; Sun, Y.; Khan, N.; Hirota, M.; Adkins, H.; Williams, K.; et al. A Nodal- and ALK4-independent signaling pathway activated by Cripto-1 through Glypican-1 and c-Src. Cancer Res. 2003, 63, 1192–1197. [Google Scholar]
- Sun, G.; Yan, S.S.; Shi, L.; Wan, Z.Q.; Jiang, N.; Fu, L.S.; Li, M.; Guo, J. MicroRNA-15b suppresses the growth and invasion of glioma cells through targeted inhibition of cripto-1 expression. Mol. Med. Rep. 2016, 13, 4897–4903. [Google Scholar] [CrossRef]
- Park, K.S.; Moon, Y.W.; Raffeld, M.; Lee, D.H.; Wang, Y.; Giaccone, G. High cripto-1 and low miR-205 expression levels as prognostic markers in early stage non-small cell lung cancer. Lung Cancer 2018, 116, 38–45. [Google Scholar] [CrossRef]
- Li, Y.; Luo, J.T.; Liu, Y.M.; Wei, W.B. miRNA-145/miRNA-205 inhibits proliferation and invasion of uveal melanoma cells by targeting NPR1/CDC42. Int. J. Ophthalmol. 2020, 13, 718–724. [Google Scholar] [CrossRef]
- Park, K.S.; Raffeld, M.; Moon, Y.W.; Xi, L.; Bianco, C.; Pham, T.; Lee, L.C.; Mitsudomi, T.; Yatabe, Y.; Okamoto, I.; et al. CRIPTO1 expression in EGFR-mutant NSCLC elicits intrinsic EGFR-inhibitor resistance. J. Clin. Investig. 2014, 124, 3003–3015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minchiotti, G.; Manco, G.; Parisi, S.; Lago, C.T.; Rosa, F.; Persico, M.G. Structure-function analysis of the EGF-CFC family member Cripto identifies residues essential for nodal signalling. Development 2001, 128, 4501–4510. [Google Scholar] [CrossRef] [PubMed]
- Blanchet, M.H.; Le Good, J.A.; Mesnard, D.; Oorschot, V.; Baflast, S.; Minchiotti, G.; Klumperman, J.; Constam, D.B. Cripto recruits Furin and PACE4 and controls Nodal trafficking during proteolytic maturation. EMBO J. 2008, 27, 2580–2591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cermola, F.; D’Aniello, C.; Tate, R.; De Cesare, D.; Martinez-Arias, A.; Minchiotti, G.; Patriarca, E.J. Gastruloid Development Competence Discriminates Different States of Pluripotency. Stem Cell Rep. 2021, 16, 354–369. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, K.; Bianco, C.; Strizzi, L.; Hamada, S.; Mancino, M.; Bailly, V.; Mo, W.; Wen, D.; Miatkowski, K.; Gonzales, M.; et al. Growth factor induction of Cripto-1 shedding by glycosylphosphatidylinositol-phospholipase D and enhancement of endothelial cell migration. J. Biol. Chem. 2007, 282, 31643–31655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, G.H.; Fujita, M.; Takaoka, K.; Murakami, Y.; Fujihara, Y.; Kanzawa, N.; Murakami, K.I.; Kajikawa, E.; Takada, Y.; Saito, K.; et al. A GPI processing phospholipase A2, PGAP6, modulates Nodal signaling in embryos by shedding CRIPTO. J. Cell Biol. 2016, 215, 705–718. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, K.; Hamada, S.; Bianco, C.; Mancino, M.; Nagaoka, T.; Gonzales, M.; Bailly, V.; Strizzi, L.; Salomon, D.S. Requirement of glycosylphosphatidylinositol anchor of Cripto-1 for trans activity as a Nodal co-receptor. J. Biol. Chem. 2007, 282, 35772–35786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watanabe, K.; Salomon, D.S. Intercellular transfer regulation of the paracrine activity of GPI-anchored Cripto-1 as a Nodal co-receptor. Biochem. Biophys. Res. Commun. 2010, 403, 108–113. [Google Scholar] [CrossRef] [Green Version]
- Blanchet, M.H.; Le Good, J.A.; Oorschot, V.; Baflast, S.; Minchiotti, G.; Klumperman, J.; Constam, D.B. Cripto localizes Nodal at the limiting membrane of early endosomes. Sci. Signal. 2008, 1, ra13. [Google Scholar] [CrossRef] [PubMed]
- Kowal, J.; Arras, G.; Colombo, M.; Jouve, M.; Morath, J.P.; Primdal-Bengtson, B.; Dingli, F.; Loew, D.; Tkach, M.; Thery, C. Proteomic comparison defines novel markers to characterize heterogeneous populations of extracellular vesicle subtypes. Proc. Natl. Acad. Sci. USA 2016, 113, E968–E977. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watanabe, K.; Nagaoka, T.; Lee, J.M.; Bianco, C.; Gonzales, M.; Castro, N.P.; Rangel, M.C.; Sakamoto, K.; Sun, Y.; Callahan, R.; et al. Enhancement of Notch receptor maturation and signaling sensitivity by Cripto-1. J. Cell Biol. 2009, 187, 343–353. [Google Scholar] [CrossRef] [PubMed]
- D’Andrea, D.; Liguori, G.L.; Le Good, J.A.; Lonardo, E.; Andersson, O.; Constam, D.B.; Persico, M.G.; Minchiotti, G. Cripto promotes A-P axis specification independently of its stimulatory effect on Nodal autoinduction. J. Cell Biol. 2008, 180, 597–605. [Google Scholar] [CrossRef] [Green Version]
- Witt, K.; Ligtenberg, M.A.; Conti, L.; Lanzardo, S.; Ruiu, R.; Wallmann, T.; Tufvesson-Stiller, H.; Chambers, B.J.; Rolny, C.; Lladser, A.; et al. Cripto-1 Plasmid DNA Vaccination Targets Metastasis and Cancer Stem Cells in Murine Mammary Carcinoma. Cancer Immunol. Res. 2018, 6, 1417–1425. [Google Scholar] [CrossRef] [Green Version]
- Ishii, H.; Afify, S.M.; Hassan, G.; Salomon, D.S.; Seno, M. Cripto-1 as a Potential Target of Cancer Stem Cells for Immunotherapy. Cancers 2021, 13, 2491. [Google Scholar] [CrossRef]
- Nagaoka, T.; Karasawa, H.; Castro, N.P.; Rangel, M.C.; Salomon, D.S.; Bianco, C. An evolving web of signaling networks regulated by Cripto-1. Growth Factors 2012, 30, 13–21. [Google Scholar] [CrossRef]
- Lonardo, E.; Hermann, P.C.; Mueller, M.T.; Huber, S.; Balic, A.; Miranda-Lorenzo, I.; Zagorac, S.; Alcala, S.; Rodriguez-Arabaolaza, I.; Ramirez, J.C.; et al. Nodal/Activin signaling drives self-renewal and tumorigenicity of pancreatic cancer stem cells and provides a target for combined drug therapy. Cell Stem Cell 2011, 9, 433–446. [Google Scholar] [CrossRef] [Green Version]
- Fan, H.; Atiya, H.I.; Wang, Y.; Pisanic, T.R.; Wang, T.H.; Shih, I.M.; Foy, K.K.; Frisbie, L.; Buckanovich, R.J.; Chomiak, A.A.; et al. Epigenomic Reprogramming toward Mesenchymal-Epithelial Transition in Ovarian-Cancer-Associated Mesenchymal Stem Cells Drives Metastasis. Cell Rep 2020, 33, 108473. [Google Scholar] [CrossRef] [PubMed]
- Kouznetsova, V.L.; Hu, H.; Teigen, K.; Zanetti, M.; Tsigelny, I.F. Cripto stabilizes GRP78 on the cell membrane. Protein Sci. 2018, 27, 653–661. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.L.; Chen, C.M.; Zhang, J.; Wang, J.; Yang, C.Z.; Lin, L.Z. Glucose-Regulated Protein 78 Signaling Regulates Hypoxia-Induced Epithelial-Mesenchymal Transition in A549 Cells. Front. Oncol. 2019, 9, 137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samavarchi-Tehrani, P.; Golipour, A.; David, L.; Sung, H.K.; Beyer, T.A.; Datti, A.; Woltjen, K.; Nagy, A.; Wrana, J.L. Functional genomics reveals a BMP-driven mesenchymal-to-epithelial transition in the initiation of somatic cell reprogramming. Cell Stem Cell 2010, 7, 64–77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, H.; Chakraborty, G.; Lee-Lim, A.P.; Mo, Q.; Decker, M.; Vonica, A.; Shen, R.; Brogi, E.; Brivanlou, A.H.; Giancotti, F.G. The BMP inhibitor Coco reactivates breast cancer cells at lung metastatic sites. Cell 2012, 150, 764–779. [Google Scholar] [CrossRef] [Green Version]
- Buijs, J.T.; Henriquez, N.V.; van Overveld, P.G.; van der Horst, G.; Que, I.; Schwaninger, R.; Rentsch, C.; Ten Dijke, P.; Cleton-Jansen, A.M.; Driouch, K.; et al. Bone morphogenetic protein 7 in the development and treatment of bone metastases from breast cancer. Cancer Res. 2007, 67, 8742–8751. [Google Scholar] [CrossRef] [Green Version]
- Buijs, J.T.; Rentsch, C.A.; van der Horst, G.; van Overveld, P.G.; Wetterwald, A.; Schwaninger, R.; Henriquez, N.V.; Ten Dijke, P.; Borovecki, F.; Markwalder, R.; et al. BMP7, a putative regulator of epithelial homeostasis in the human prostate, is a potent inhibitor of prostate cancer bone metastasis in vivo. Am. J. Pathol. 2007, 171, 1047–1057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mancino, M.; Strizzi, L.; Wechselberger, C.; Watanabe, K.; Gonzales, M.; Hamada, S.; Normanno, N.; Salomon, D.S.; Bianco, C. Regulation of human Cripto-1 gene expression by TGF-beta1 and BMP-4 in embryonal and colon cancer cells. J. Cell Physiol. 2008, 215, 192–203. [Google Scholar] [CrossRef]
- Ye, Q.; Li, J.; Wang, X.; Zhang, X.; Lin, J.; Huo, Y.; Sun, Z.; Xie, S.; Huang, Z. In vivo and in vitro study of co-expression of LMP1 and Cripto-1 in nasopharyngeal carcinoma☆. Braz. J. Otorhinolaryngol. 2020, 86, 617–625. [Google Scholar] [CrossRef]
- Peinado, H.; Zhang, H.; Matei, I.R.; Costa-Silva, B.; Hoshino, A.; Rodrigues, G.; Psaila, B.; Kaplan, R.N.; Bromberg, J.F.; Kang, Y.; et al. Pre-metastatic niches: Organ-specific homes for metastases. Nat. Rev. Cancer 2017, 17, 302–317. [Google Scholar] [CrossRef] [PubMed]
- Melo, S.A.; Luecke, L.B.; Kahlert, C.; Fernandez, A.F.; Gammon, S.T.; Kaye, J.; LeBleu, V.S.; Mittendorf, E.A.; Weitz, J.; Rahbari, N.; et al. Glypican-1 identifies cancer exosomes and detects early pancreatic cancer. Nature 2015, 523, 177–182. [Google Scholar] [CrossRef] [Green Version]
- Folkman, J. Endogenous angiogenesis inhibitors. APMIS 2004, 112, 496–507. [Google Scholar] [CrossRef]
- Aggarwal, B.B.; Vijayalekshmi, R.V.; Sung, B. Targeting inflammatory pathways for prevention and therapy of cancer: Short-term friend, long-term foe. Clin. Cancer Res. 2009, 15, 425–430. [Google Scholar] [CrossRef] [Green Version]
- Pein, M.; Insua-Rodriguez, J.; Hongu, T.; Riedel, A.; Meier, J.; Wiedmann, L.; Decker, K.; Essers, M.A.G.; Sinn, H.P.; Spaich, S.; et al. Metastasis-initiating cells induce and exploit a fibroblast niche to fuel malignant colonization of the lungs. Nat. Commun. 2020, 11, 1494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, F.T.; Sun, W.; Zhang, J.T.; Fan, Y.Z. Cancer-associated fibroblast regulation of tumor neo-angiogenesis as a therapeutic target in cancer. Oncol. Lett. 2019, 17, 3055–3065. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruggiero, D.; Nappo, S.; Nutile, T.; Sorice, R.; Talotta, F.; Giorgio, E.; Bellenguez, C.; Leutenegger, A.-L.; Liguori, G.L.; Ciullo, M. Genetic Variants Modulating CRIPTO Serum Levels Identified by Genome-Wide Association Study in Cilento Isolates. PLoS Genet. 2015, 11, e1004976. [Google Scholar] [CrossRef]
- Jain, A.; Mallupattu, S.; Thakur, R.; Ghoshal, S.; Pal, A. Cripto 1, a potential biomarker for oral squamous cell carcinoma. Oral. Oncol. 2019, 90, 153–154. [Google Scholar] [CrossRef] [PubMed]
- Chen, V.; Iwama, E.; Kim, I.K.; Giaccone, G. Serum CRIPTO does not confer drug resistance against osimertinib but is an indicator of tumor burden in non-small cell lung cancer. Lung Cancer 2020, 145, 48–57. [Google Scholar] [CrossRef] [PubMed]
- Beacon-Biomedical. Non-Clinical Verification and Clinical Validationo of BeScreened-CRC, a Blood-Based In Vitro Diagnostic Multivariate Index Assay for the Detection of Colorectal Cancer in Screening Non-Compliant Patients; [White Paper]; Beacon-Biomedical Inc.: Phoenix, AZ, USA, 2017. [Google Scholar]
- Zhong, X.Y.; Zhang, L.H.; Jia, S.Q.; Shi, T.; Niu, Z.J.; Du, H.; Zhang, G.G.; Hu, Y.; Lu, A.P.; Li, J.Y.; et al. Positive association of up-regulated Cripto-1 and down-regulated E-cadherin with tumour progression and poor prognosis in gastric cancer. Histopathology 2008, 52, 560–568. [Google Scholar] [CrossRef]
- Miyoshi, N.; Ishii, H.; Mimori, K.; Sekimoto, M.; Doki, Y.; Mori, M. TDGF1 is a novel predictive marker for metachronous metastasis of colorectal cancer. Int. J. Oncol. 2010, 36, 563–568. [Google Scholar] [CrossRef] [PubMed]
- Connolly, E.C.; Freimuth, J.; Akhurst, R.J. Complexities of TGF-beta targeted cancer therapy. Int. J. Biol. Sci. 2012, 8, 964–978. [Google Scholar] [CrossRef] [Green Version]
- Ligtenberg, M.A.; Witt, K.; Galvez-Cancino, F.; Sette, A.; Lundqvist, A.; Lladser, A.; Kiessling, R. Cripto-1 vaccination elicits protective immunity against metastatic melanoma. OncoImmunology 2016, 5, e1128613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kelly, R.K.; Olson, D.L.; Sun, Y.; Wen, D.; Wortham, K.A.; Antognetti, G.; Cheung, A.E.; Orozco, O.E.; Yang, L.; Bailly, V.; et al. An antibody-cytotoxic conjugate, BIIB015, is a new targeted therapy for Cripto positive tumours. Eur. J. Cancer 2011, 47, 1736–1746. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Bao, Y.L.; Wu, Y.; Yu, C.L.; Huang, Y.X.; Sun, Y.; Zheng, L.H.; Li, Y.X. Alantolactone inhibits cell proliferation by interrupting the interaction between Cripto-1 and activin receptor type II A in activin signaling pathway. J. Biomol. Screen 2011, 16, 525–535. [Google Scholar] [CrossRef] [Green Version]
- Castro, N.P.; Rangel, M.C.; Merchant, A.S.; MacKinnon, G.; Cuttitta, F.; Salomon, D.S.; Kim, Y.S. Sulforaphane Suppresses the Growth of Triple-negative Breast Cancer Stem-like Cells In vitro and In vivo. Cancer Prev. Res. 2019, 12, 147–158. [Google Scholar] [CrossRef] [Green Version]
- Ni, M.; Zhang, Y.; Lee, A.S. Beyond the endoplasmic reticulum: Atypical GRP78 in cell viability, signalling and therapeutic targeting. Biochem. J. 2011, 434, 181–188. [Google Scholar] [CrossRef]
- Jiang, B.; Zhang, R.; Zhang, J.; Hou, Y.; Chen, X.; Zhou, M.; Tian, X.; Hao, C.; Fan, K.; Yan, X. GRP78-targeted ferritin nanocaged ultra-high dose of doxorubicin for hepatocellular carcinoma therapy. Theranostics 2019, 9, 2167–2182. [Google Scholar] [CrossRef]
- Liu, R.; Li, X.; Gao, W.; Zhou, Y.; Wey, S.; Mitra, S.K.; Krasnoperov, V.; Dong, D.; Liu, S.; Li, D.; et al. Monoclonal antibody against cell surface GRP78 as a novel agent in suppressing PI3K/AKT signaling, tumor growth, and metastasis. Clin. Cancer Res. 2013, 19, 6802–6811. [Google Scholar] [CrossRef] [Green Version]
- Bian, T.; Tagmount, A.; Vulpe, C.; Vijendra, K.C.; Xing, C. CXL146, a Novel 4H-Chromene Derivative, Targets GRP78 to Selectively Eliminate Multidrug-Resistant Cancer Cells. Mol. Pharm. 2020, 97, 402–408. [Google Scholar] [CrossRef] [Green Version]
- Foca, G.; Iaccarino, E.; Foca, A.; Sanguigno, L.; Untiveros, G.; Cuevas-Nunez, M.; Strizzi, L.; Leonardi, A.; Ruvo, M.; Sandomenico, A. Development of conformational antibodies targeting Cripto-1 with neutralizing effects in vitro. Biochimie 2019, 158, 246–256. [Google Scholar] [CrossRef] [PubMed]
- Ishii, H.; Zahra, M.H.; Takayanagi, A.; Seno, M. A Novel Artificially Humanized Anti-Cripto-1 Antibody Suppressing Cancer Cell Growth. Int. J. Mol. Sci. 2021, 22, 1709. [Google Scholar] [CrossRef] [PubMed]
- Cohnheim, J. Congenitales, quergestreiftes Muskelsarkom der Nieren. Arch. Pathol. Anat. Physiol. Klin. Med. 1875, 65, 64–69. [Google Scholar] [CrossRef] [Green Version]
- Virchow, R. Editorial Archive fuer pathologische. Anat. Physiol. Fuer Klin. Med. 1855, 8, 23–54. [Google Scholar]



{kind=link}
{kind=link}
| Tissue Type | Expression in Normal Tissue | Expression in Cancer | Method of Detection |
|---|---|---|---|
| Blood | Detected in long-term HSCs [39] | - | RT-qPCR |
| Liver | Undetectable in healthy livers [35,40] Elevated expression in cirrhotic livers [35] | Overexpressed correlates with poor prognosis in HCC [41] | Northern blot, RT-qPCR |
| Mammary | Detectable in the surrounding fat pad of developing rudiments [42] Peak epithelial expression during alveogenesis [43,44] | Overexpressed in ~80% of invading breast cancers [45] | Immunohistochemistry RT-qPCR, Western |
| Muscle | Expressed in developing cardiac ventricles and outflow tracts [46] Upregulated in cardiac tissue post MI [35] Promotes skeletal muscle regeneration through myogenic differentiation of satellite cells and macrophage plasticity [37,47] | - | RNA in situ, Immunohistochemistry, FACS |
| CNS | Expressed throughout the CNS in adult non-human primates and upregulated during infection with simian human immunodeficiency virus [48]. | Serum CRIPTO correlates with poor GBM prognosis. Localizes to perivascular tumor cells and endothelial cells in GMB [49] | ELISA, immunohistochemistry, RT-PCR |
| Pancreas | Moderate and low expression in normal ductal and acinar cells respectively. Increased expression in both ductal and acinar cells in chronic pancreatitis [50]. | CRIPTO overexpression correlates with tumor stage [50] | Immunohistochemistry, RT-qPCR |
| Prostate | Low/undetectable level in healthy and BPH patient samples [51,52] | Metastatic lesions are characterized by CRIPTO overexpression [53] Overexpression in primary tumors predicts patient prognosis [51] | RT-qPCR, immunohistochemistry |
| Skin | Undetectable in healthy murine skin, upregulated in TPA treated murine skin and benign murine papillomas [54] | Overexpressed in a subset of cutaneous melanomas. Inversely correlates with saracatinib efficacy [55]. | RT-qPCR |
| Renal | Low levels in adjacent non-tumor tissue relative to adjacent neoplastic lesions [56] | CRIPTO expression correlates with tumor aggressiveness and poor survival [56] | RT-qPCR |
| Colon | Undetectable in normal adult mucosa [57,58] | Detectable and independently correlates with poor prognosis [59] | Immunohistochemistry |
| Oral Squamous | Present sporadically at low levels in healthy tissue and widely expressed in dysplastic epithelia [60] | Overexpressed relative to homeostatic tissue [60] | Immunohistochemistry |
| Cervix | Low, variable expression across multiple healthy patient samples [61] | - | Immunohistochemistry |
| Gastric | Detectable in a subset of healthy patients [62,63] | Overexpressed relative to homeostatic tissue [62] | Immunohistochemistry |
| Bladder | - | Overexpressed relative to homeostatic tissue and correlated with tumor size and grade [64] | Immunohistochemistry |
| Lung | - | Overexpression predicts outcome in NSCLC [65] | Immunohistochemistry |
| Esophageal | Undetectable in normal adult epithelia [66] | Overexpressed in esophageal squamous cell carcinoma. Positively correlates with tumor stage [67]. | Immunohistochemistry |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Freeman, D.W.; Rodrigues Sousa, E.; Karkampouna, S.; Zoni, E.; Gray, P.C.; Salomon, D.S.; Kruithof-de Julio, M.; Spike, B.T. Whence CRIPTO: The Reemergence of an Oncofetal Factor in ‘Wounds’ That Fail to Heal. Int. J. Mol. Sci. 2021, 22, 10164. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms221810164
Freeman DW, Rodrigues Sousa E, Karkampouna S, Zoni E, Gray PC, Salomon DS, Kruithof-de Julio M, Spike BT. Whence CRIPTO: The Reemergence of an Oncofetal Factor in ‘Wounds’ That Fail to Heal. International Journal of Molecular Sciences. 2021; 22(18):10164. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms221810164
Chicago/Turabian StyleFreeman, David W., Elisa Rodrigues Sousa, Sofia Karkampouna, Eugenio Zoni, Peter C. Gray, David S. Salomon, Marianna Kruithof-de Julio, and Benjamin T. Spike. 2021. "Whence CRIPTO: The Reemergence of an Oncofetal Factor in ‘Wounds’ That Fail to Heal" International Journal of Molecular Sciences 22, no. 18: 10164. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms221810164