
Immunomodulatory Properties of Umbilical Cord Blood-Derived Small Extracellular Vesicles and Their Therapeutic Potential for Inflammatory Skin Disorders

 ,
,  , , ,
, , ,
Abstract
:1. Introduction
2. Results
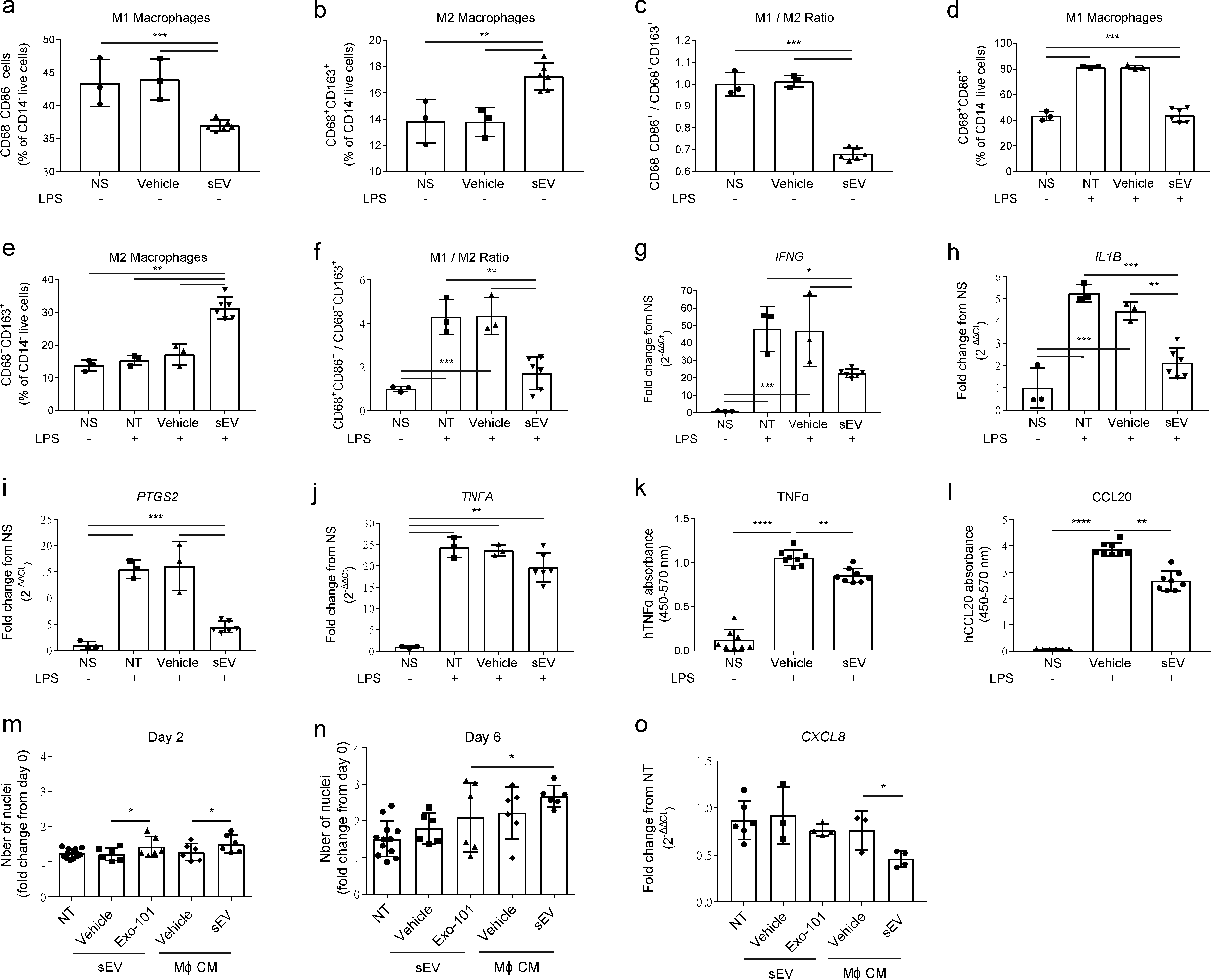
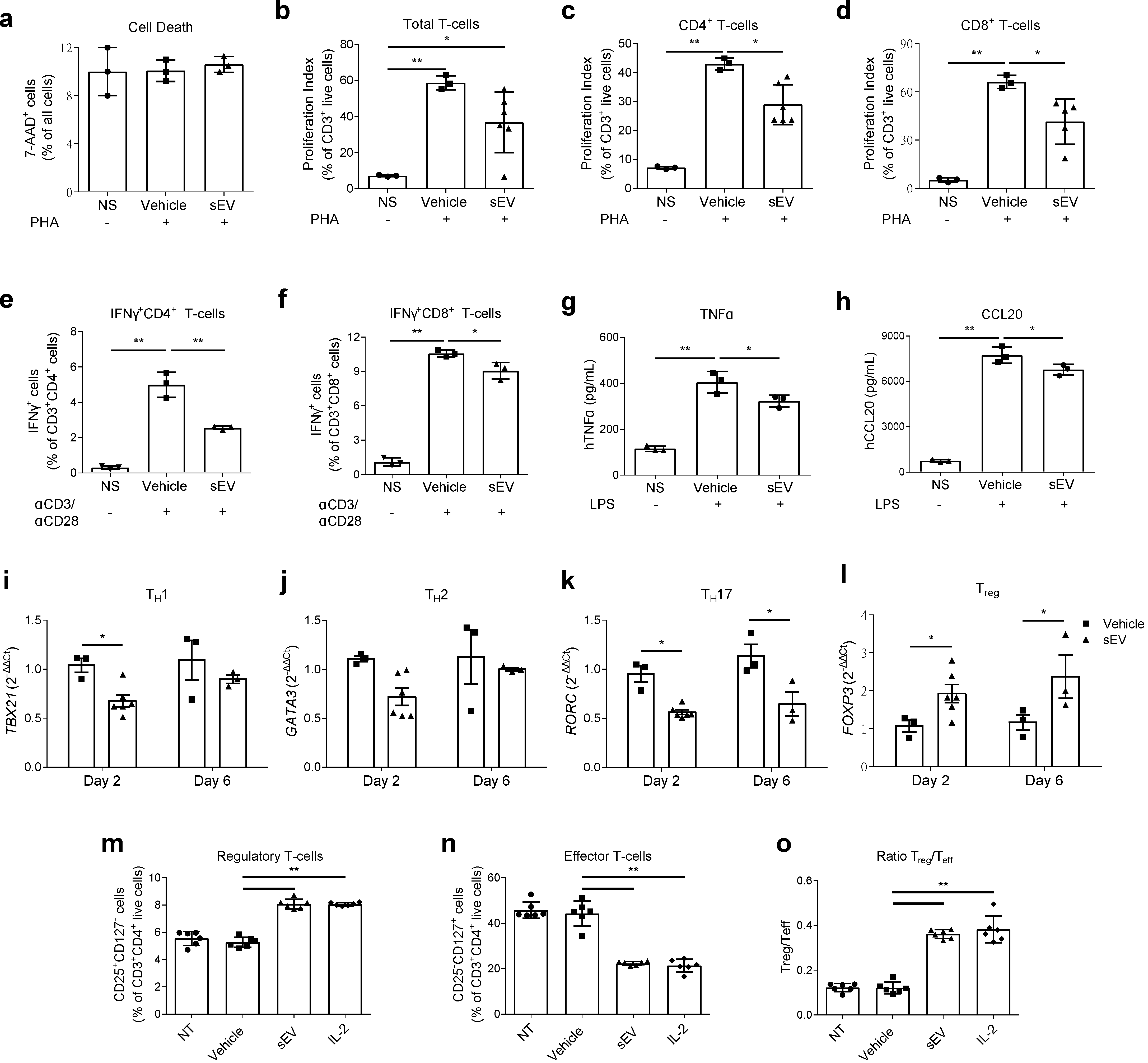
2.1. UCB-MNC-sEV Have Anti-Inflammatory and Tolerogenic Effects, Modulating Different Immune Players Directly and Indirectly
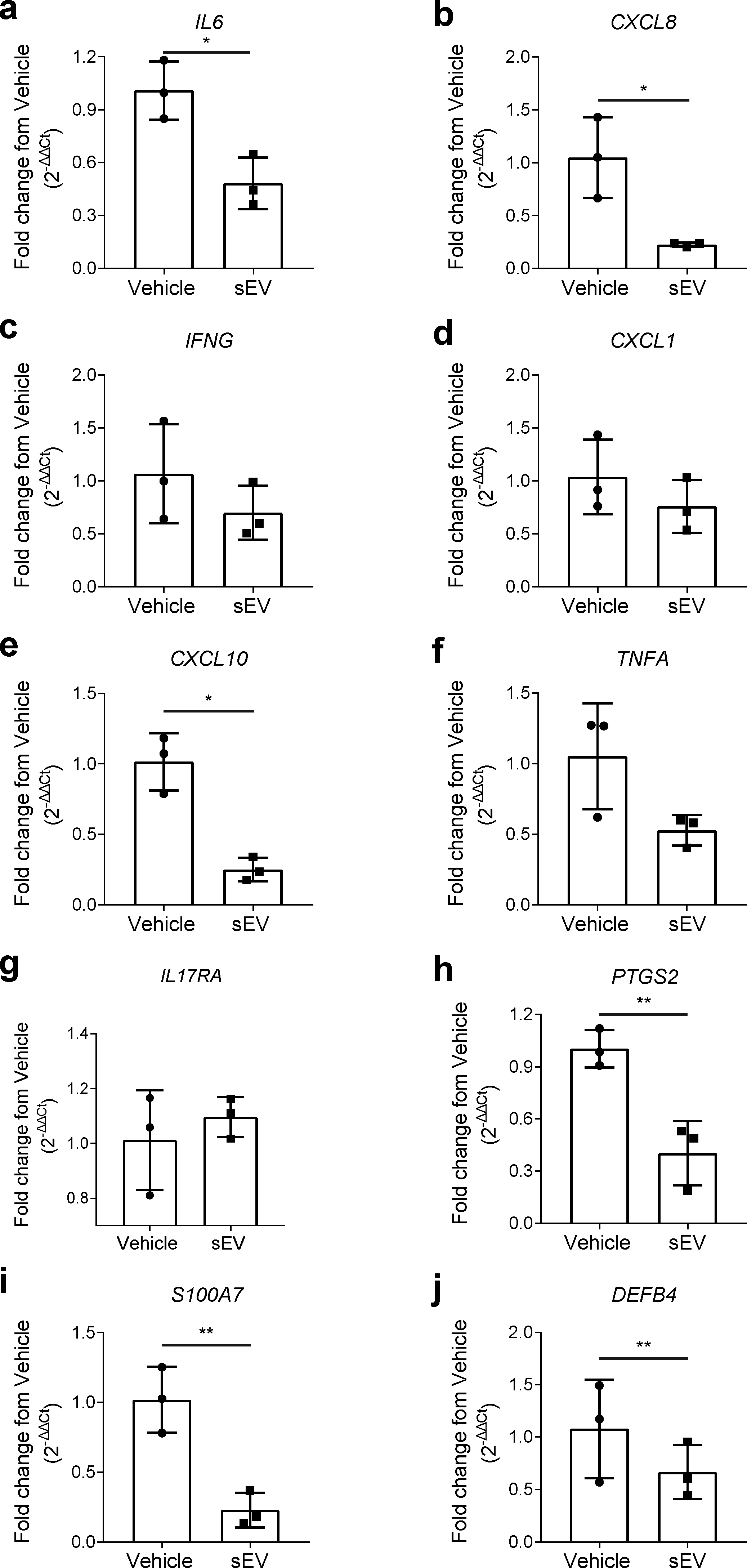
2.2. UCB-MNC-sEV Reduce the Expression of Psoriasis Markers, in an In Vitro 3D Model
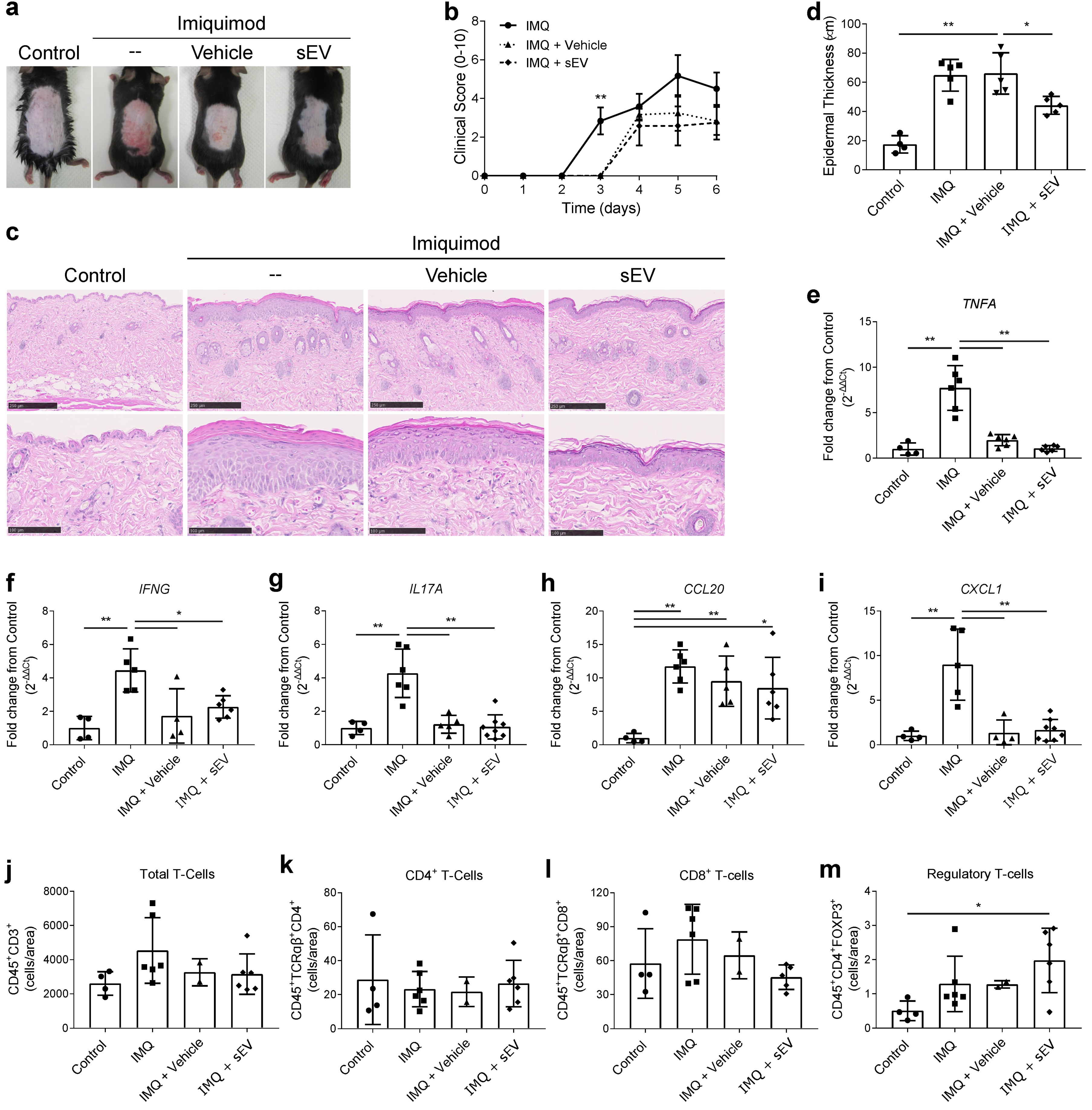
2.3. UCB-MNC-sEV Show a Modest Effect in Imiquimod-Induced Psoriasis, Regulating Keratinocyte Proliferation and T-Cell Homeostasis
3. Discussion
4. Materials and Methods
4.1. UCB Collection, Testing, and Data Protection
4.2. Cell Culture
4.2.1. UCB-MNC
4.2.2. THP-1-Derived Macrophages
4.2.3. Dermal Fibroblasts
4.2.4. PBMC
4.3. UCB-MNC-sEV Isolation
4.4. Gene Expression
4.5. Protein Quantification and Flow Cytometry (Human)
4.6. Reconstructed Psoriatic Human Epidermis
4.7. UCB-MNC-sEV Formulation for In Vivo Topical Use
4.8. Imiquimod-Induced Psoriasis
4.9. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bai, L.; Shao, H.; Wang, H.; Zhang, Z.; Su, C.; Dong, L.; Yu, B.; Chen, X.; Li, X.; Zhang, X. Effects of Mesenchymal Stem Cell-Derived Exosomes on Experimental Autoimmune Uveitis. Sci. Rep. 2017, 7, 4323. [Google Scholar] [CrossRef] [Green Version]
- Boehncke, W.-H.; Brembilla, N.C. Unmet Needs in the Field of Psoriasis: Pathogenesis and Treatment. Clin. Rev. Allergy Immunol. 2018, 55, 295–311. [Google Scholar] [CrossRef] [PubMed]
- Cao, L.; Xu, H.; Wang, G.; Liu, M.; Tian, D.; Yuan, Z. Extracellular vesicles derived from bone marrow mesenchymal stem cells attenuate dextran sodium sulfate-induced ulcerative colitis by promoting M2 macrophage polarization. Int. Immunopharmacol. 2019, 72, 264–274. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, R.M.S.; Rodrigues, S.C.; Gomes, C.F.; Duarte, F.V.; Romao, M.; Leal, E.C.; Freire, P.C.; Neves, R.; Simões-Correia, J. Development of an optimized and scalable method for isolation of umbilical cord blood-derived small extracellular vesicles for future clinical use. Stem Cells Transl. Med. 2021, 10, 910–921. [Google Scholar] [CrossRef]
- Del Fattore, A.; Luciano, R.; Pascucci, L.; Goffredo, B.M.; Giorda, E.; Scapaticci, M.; Fierabracci, A.; Muraca, M. Immunoregulatory Effects of Mesenchymal Stem Cell-Derived Extracellular Vesicles on T Lymphocytes. Cell Transplant. 2015, 24, 2615–2627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Der Fits, L.; Mourits, S.; Voerman, J.S.A.; Kant, M.; Boon, L.; Laman, J.D.; Cornelissen, F.; Mus, A.-M.; Florencia, E.; Prens, E.; et al. Imiquimod-Induced Psoriasis-Like Skin Inflammation in Mice Is Mediated via the IL-23/IL-17 Axis. J. Immunol. 2009, 182, 5836–5845. [Google Scholar] [CrossRef] [PubMed]
- Godfrey, W.R.; Spoden, D.J.; Ge, Y.G.; Baker, S.R.; Liu, B.; Levine, B.L.; June, C.H.; Blazar, B.R.; Porter, S.B. Cord blood CD4(+)CD25(+)-derived T regulatory cell lines express FoxP3 protein and manifest potent suppressor function. Blood 2005, 105, 750–758. [Google Scholar] [CrossRef]
- Gordon, K.B.; Strober, B.; Lebwohl, M.; Augustin, M.; Blauvelt, A.; Poulin, Y.; Papp, K.A.; Sofen, H.; Puig, L.; Foley, P.; et al. Efficacy and safety of risankizumab in moderate-to-severe plaque psoriasis (UltIMMa-1 and UltIMMa-2): Results from two double-blind, randomised, placebo-controlled and ustekinumab-controlled phase 3 trials. Lancet 2018, 392, 650–661. [Google Scholar] [CrossRef]
- Griffiths, C.E.M.; Armstrong, A.W.; Gudjonsson, J.E.; Barker, J.N.W.N. Psoriasis. Lancet 2021, 397, 1301–1315. [Google Scholar] [CrossRef]
- Harris, D.T.; Schumacher, M.J.; Locascio, J.; Besencon, F.J.; Olson, G.B.; DeLuca, D.; Shenker, L.; Bard, J.; Boyse, E.A. Phenotypic and functional immaturity of human umbilical cord blood T lymphocytes. Proc. Natl. Acad. Sci. USA 1992, 89, 10006–10010. [Google Scholar] [CrossRef] [Green Version]
- Hartwig, T.; Zwicky, P.; Schreiner, B.; Yawalkar, N.; Cheng, P.; Navarini, A.; Dummer, R.; Flatz, L.; Conrad, C.; Schlapbach, C.; et al. Regulatory T Cells Restrain Pathogenic T Helper Cells during Skin Inflammation. Cell Rep. 2018, 25, 3564–3572. [Google Scholar] [CrossRef] [Green Version]
- Henriques-Antunes, H.; Cardoso, R.; Zonari, A.; Correia, J.S.; Leal, E.; Jiménez-Balsa, A.; Lino, M.M.; Barradas, A.; Kostic, I.; Gomes, C.; et al. The Kinetics of Small Extracellular Vesicle Delivery Impacts Skin Tissue Regeneration. ACS Nano 2019, 13, 8694–8707. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.; Song, X.; Yu, H.; Sun, J.; Zhao, Y. Released Exosomes Contribute to the Immune Modulation of Cord Blood-Derived Stem Cells. Front. Immunol. 2020, 11, 165. [Google Scholar] [CrossRef] [PubMed]
- Karimkhani, C.; Dellavalle, R.P.; Coffeng, L.E.; Flohr, C.; Hay, R.J.; Langan, S.; Nsoesie, E.O.; Ferrari, A.; Erskine, H.E.; Silverberg, J.I.; et al. Global Skin Disease Morbidity and Mortality: An Update From the Global Burden of Disease Study 2013. JAMA Dermatol. 2017, 153, 406–412. [Google Scholar] [CrossRef]
- Kim, S.-H.; Lechman, E.; Bianco, N.; Menon, R.; Keravala, A.; Nash, J.; Mi, Z.; Watkins, S.; Gambotto, A.; Robbins, P.D. Exosomes derived from IL-10-treated dendritic cells can suppress inflammation and collagen-induced arthritis. J. Immunol. 2005, 174, 6440–6448. [Google Scholar] [CrossRef] [Green Version]
- Krueger, J.G.; Wharton, K.A.; Schlitt, T.; Suprun, M.; Torene, R.I.; Jiang, X.; Wang, C.Q.; Fuentes-Duculan, J.; Hartmann, N.; Peters, T.; et al. IL-17A inhibition by secukinumab induces early clinical, histopathologic, and molecular resolution of psoriasis. J. Allergy Clin. Immunol. 2019, 144, 750–763. [Google Scholar] [CrossRef] [Green Version]
- Lai, P.; Weng, J.; Guo, L.; Chen, X.; Du, X. Novel insights into MSC-EVs therapy for immune diseases. Biomark. Res. 2019, 7, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, H.W.; Collins, S.A.; Resneck, J.S.; Bolognia, J.L.; Hodge, J.A.; Rohrer, T.A.; Van Beek, M.J.; Margolis, D.J.; Sober, A.J.; Weinstock, M.A.; et al. The burden of skin disease in the United States. J. Am. Acad. Dermatol. 2017, 76, 958–972. [Google Scholar] [CrossRef] [Green Version]
- Ma, D.; Xu, K.; Zhang, G.; Liu, Y.; Gao, J.; Tian, M.; Wei, C.; Li, J.; Zhang, L. Immunomodulatory effect of human umbilical cord mesenchymal stem cells on T lymphocytes in rheumatoid arthritis. Int. Immunopharmacol. 2019, 74, 105687. [Google Scholar] [CrossRef]
- McBride, J.D.; Rodriguez-Menocal, L.; Badiavas, E.V. Extracellular Vesicles as Biomarkers and Therapeutics in Dermatology: A Focus on Exosomes. J. Investig. Dermatol. 2017, 137, 1622–1629. [Google Scholar] [CrossRef]
- Van Niel, G.; D’Angelo, G.; Raposo, G. Shedding light on the cell biology of extracellular vesicles. Nat. Reviews. Mol. Cell Biol. 2018, 19, 213–228. [Google Scholar] [CrossRef]
- Papp, K.A.; Merola, J.F.; Gottlieb, A.B.; Griffiths, C.E.; Cross, N.; Peterson, L.; Cioffi, C.; Blauvelt, A. Dual neutralization of both interleukin 17A and interleukin 17F with bimekizumab in patients with psoriasis: Results from BE ABLE 1, a 12-week randomized, double-blinded, placebo-controlled phase 2b trial. J. Am. Acad. Dermatol. 2018, 79, 277–286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phinney, D.G.; Pittenger, M.F. Concise Review: MSC-Derived Exosomes for Cell-Free Therapy. Stem Cells 2017, 35, 851–858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raposo, G.; Stoorvogel, W. Extracellular vesicles: Exosomes, microvesicles, and friends. J. Cell Biol. 2013, 200, 373–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodrigues, S.C.; Cardoso, R.M.S.; Gomes, C.F.; Duarte, F.V.; Freire, P.C.; Neves, R.; Simoes-Correia, J. Toxicological Profile of Umbilical Cord Blood-Derived Small Extracellular Vesicles. Membranes 2021, 11, 647. [Google Scholar] [CrossRef]
- Sugiyama, H.; Gyulai, R.; Toichi, E.; Garaczi, E.; Shimada, S.; Stevens, S.R.; McCormick, T.S.; Cooper, K. Dysfunctional blood and target tissue CD4+CD25high regulatory T cells in psoriasis: Mechanism underlying unrestrained pathogenic effector T cell proliferation. J. Immunol. 2005, 174, 164–173. [Google Scholar] [CrossRef] [Green Version]
- Di Trapani, M.; Bassi, G.; Midolo, M.; Gatti, A.; Kamga, P.T.; Cassaro, A.; Carusone, R.; Adamo, A.; Krampera, M. Differential and transferable modulatory effects of mesenchymal stromal cell-derived extracellular vesicles on T, B and NK cell functions. Sci. Rep. 2016, 6, 24120. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.M.; Wu, C.; Jin, H.Z. Exosomes in chronic inflammatory skin diseases and skin tumors. Exp. Dermatol. 2019, 28, 213–218. [Google Scholar] [CrossRef]
- Wang, Z.-Y.; Yan, B.-X.; Zhou, Y.; Chen, X.-Y.; Zhang, J.; Cai, S.-Q.; Zheng, M.; Man, X.-Y. miRNA Profiling of Extracellular Vesicles Reveals Biomarkers for Psoriasis. J. Investig. Dermatol. 2021, 141, 185–189. [Google Scholar] [CrossRef]
- Yuan, L.; Liu, Y.; Qu, Y.; Liu, L.; Li, H. Exosomes Derived From MicroRNA-148b-3p-Overexpressing Human Umbilical Cord Mesenchymal Stem Cells Restrain Breast Cancer Progression. Front. Oncol. 2019, 9, 1076. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.; Lee, H.; Wang, X.; Rai, A.; Groot, M.; Jin, Y. Exosome-Mediated Small RNA Delivery: A Novel Therapeutic Approach for Inflammatory Lung Responses. Mol. Ther. 2018, 26, 2119–2130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Y.-G.; Feng, X.-M.; Abbott, J.; Fang, X.-H.; Hao, Q.; Monsel, A.; Qu, J.-M.; Matthay, M.A.; Lee, J.W. Human mesenchymal stem cell microvesicles for treatment of Escherichia coli endotoxin-induced acute lung injury in mice. Stem Cells 2014, 32, 116–125. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target | Clone | Manufacturer |
|---|---|---|
| Anti-human antibodies | ||
| CD14 | M5E2 | BD Biosciences, San Jose, CA, USA |
| CD163 | GHI/61 | BD Biosciences, San Jose, CA, USA |
| CD28 | CD28.2 | BioLegend, San Diego, CA, USA |
| CD3 | OKT3 | BioLegend, San Diego, CA, USA |
| CD68 | Y1/82A | BD Biosciences, San Jose, CA, USA |
| CD86 | FUN-1 | BD Biosciences, San Jose, CA, USA |
| IL-2 | MQ1-17H12 | BioLegend, San Diego, CA, USA |
| Anti-mouse antibodies | ||
| CD11b | M1/70 | BD Biosciences, San Jose, CA, USA |
| CD11c | HL3 | BD Biosciences, San Jose, CA, USA |
| CD16/CD32 | 2.4G2 | Tonbo Biosciences, San Diego, CA, USA |
| CD25 | PC61.5 | eBioscience, San Diego, CA, USA |
| CD3e | 145-2C11 | BD Biosciences, San Jose, CA, USA |
| CD4 | RM4-5 | BD Biosciences, San Jose, CA, USA |
| CD45.2 | 104 | eBioscience, San Diego, CA, USA |
| CD62L | MEL-14 | BD Biosciences, San Jose, CA, USA |
| CD8 | 53-6.7 | BD Biosciences, San Jose, CA, USA |
| Ly6C | AL-21 | BD Biosciences, San Jose, CA, USA |
| Ly6G | 1A8 | BD Biosciences, San Jose, CA, USA |
| TCR γ/δ | GL3 | BioLegend, San Diego, CA, USA |
| TCR α/β | GL2 | BioLegend, San Diego, CA, USA |
| Target Gene | Forward Sequence | Reverse Sequence |
|---|---|---|
| Human | ||
| CXCL1 | AGGGAATTCACCCCAAGAAC | ACTATGGGGGATGCAGGATT |
| CXCL10 | TTCAAGGAGTACCTCTCTCTAG | CTGGATTCAGACATCTCTTCTC |
| DEFB4 | ATCAGCCATGAGGGTCTTGT | GAGACCACAGGTGCCAATTT |
| FOXP3 | GCTTCATCTGTGGCATCATC | TGGAGGAACTCTGGGAATGT |
| GATA3 | CGCCTGCGGGCTCTATC | CCTTCGCTTGGGCTTAATGA |
| IFNG | GGTAACTGACTTGAATGTCC | TTTTCGCTTCCCTGTTTTAG |
| IL1B | CTAAACAGATGAAGTGCTCC | GGTCATTCTCCTGGAAGG |
| IL6 | GGTACATCCTCGACGGCATCT | GT GCCTCTTTGCTGCTTTCAC |
| IL8 | GTTTTTGAAGAGGGCTGAG | TTTGCTTGAAGTTTCACTGG |
| PTGS2 | ATCTACCCTCCTCAAGTCCC | TACCAGAAGGGCAGGATACAG |
| RORC | TGGACCACCCCCTGCTGAGAA | CTTCAATTTGTGTTCTCATGACT |
| S100A7 | CCAAACACACACATCTCACTCA | TCAGCTTGAGTGTTGCTCATC |
| TBX21 | GATGTTTGTGGACGTGGTCTTG | CTTTCCACACTGCACCCACTT |
| TNFA | AGGCAGTCAGATCATCTTC | TTATCTCTCAGCTCCACG |
| Mouse | ||
| CCL20 | ACTGTTGCCTCTCGTACATACA | GAGGAGGTTCACAGCCCTTTT |
| CXCL1 | GCTTGAAGGTGTTGCCCTCAG | AGAAGCCAGCGTTCACCAGAC |
| FOXP3 | CACCCAGGAAAGACAGCAACC | GCAAGAGCTCTTGTCCATTGA |
| IFNG | CGGCACAGTCATTGAAAGCCTA | GTTGCTGATGGCCTGATTGTC |
| IL17A | TTTAACTCCCTTGGCGCAAAA | CTTTCCCTCCGCATTGACAC |
| TNFA | GTTCTATGGCCCAGACCCTCAC | GGCACCACTAGTTGGTTGTCTTTG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodrigues, S.C.; Cardoso, R.M.S.; Freire, P.C.; Gomes, C.F.; Duarte, F.V.; Neves, R.P.d.; Simões-Correia, J. Immunomodulatory Properties of Umbilical Cord Blood-Derived Small Extracellular Vesicles and Their Therapeutic Potential for Inflammatory Skin Disorders. Int. J. Mol. Sci. 2021, 22, 9797. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22189797
Rodrigues SC, Cardoso RMS, Freire PC, Gomes CF, Duarte FV, Neves RPd, Simões-Correia J. Immunomodulatory Properties of Umbilical Cord Blood-Derived Small Extracellular Vesicles and Their Therapeutic Potential for Inflammatory Skin Disorders. International Journal of Molecular Sciences. 2021; 22(18):9797. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22189797
Chicago/Turabian StyleRodrigues, Sílvia C., Renato M. S. Cardoso, Patricia C. Freire, Cláudia F. Gomes, Filipe V. Duarte, Ricardo Pires das Neves, and Joana Simões-Correia. 2021. "Immunomodulatory Properties of Umbilical Cord Blood-Derived Small Extracellular Vesicles and Their Therapeutic Potential for Inflammatory Skin Disorders" International Journal of Molecular Sciences 22, no. 18: 9797. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22189797