
RNA-Sequencing Based microRNA Expression Signature of Colorectal Cancer: The Impact of Oncogenic Targets Regulated by miR-490-3p

 , ,
, ,
Abstract
:1. Introduction
2. Results
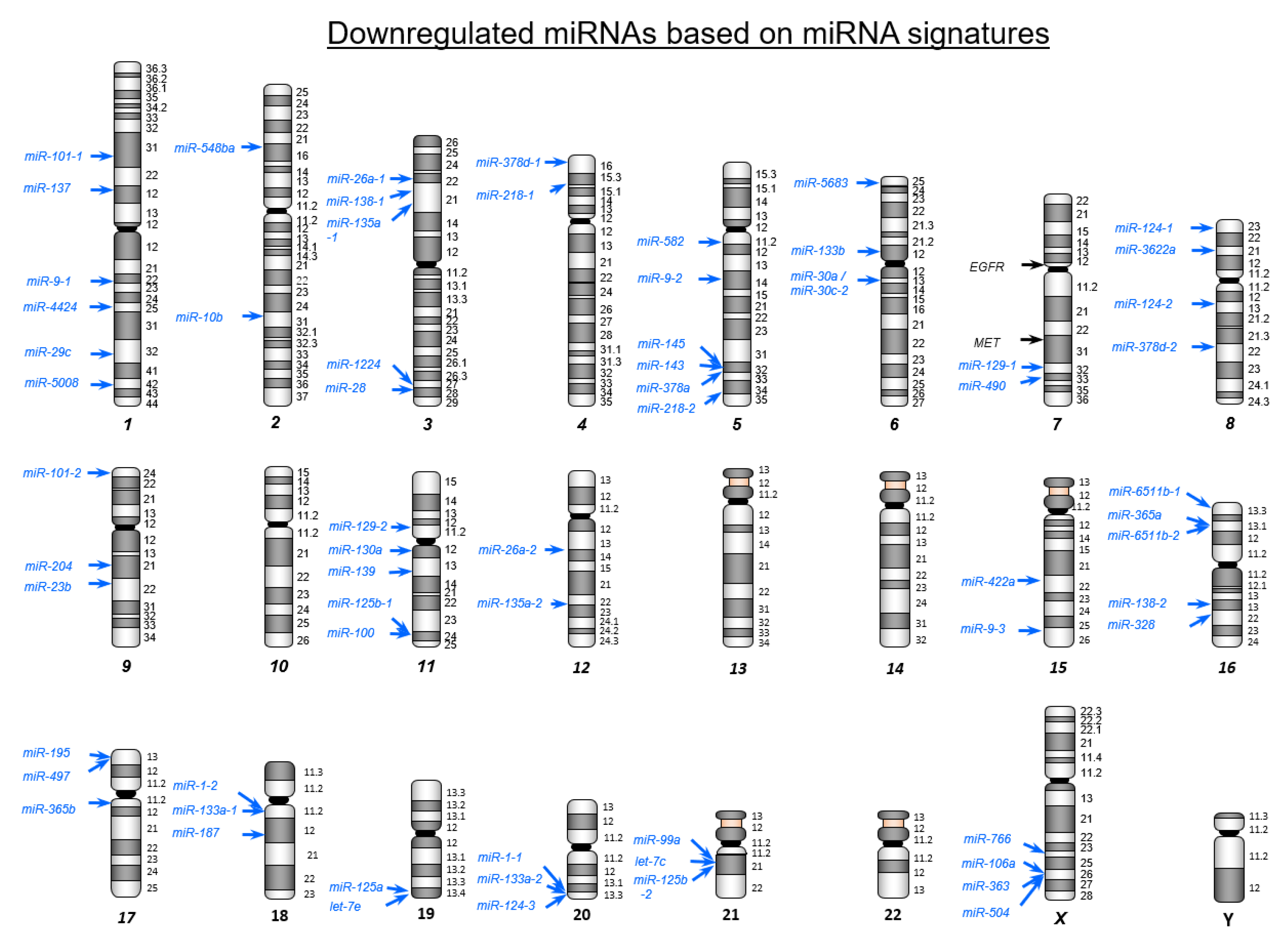
2.1. Creating miRNA Expression Signature in CRC
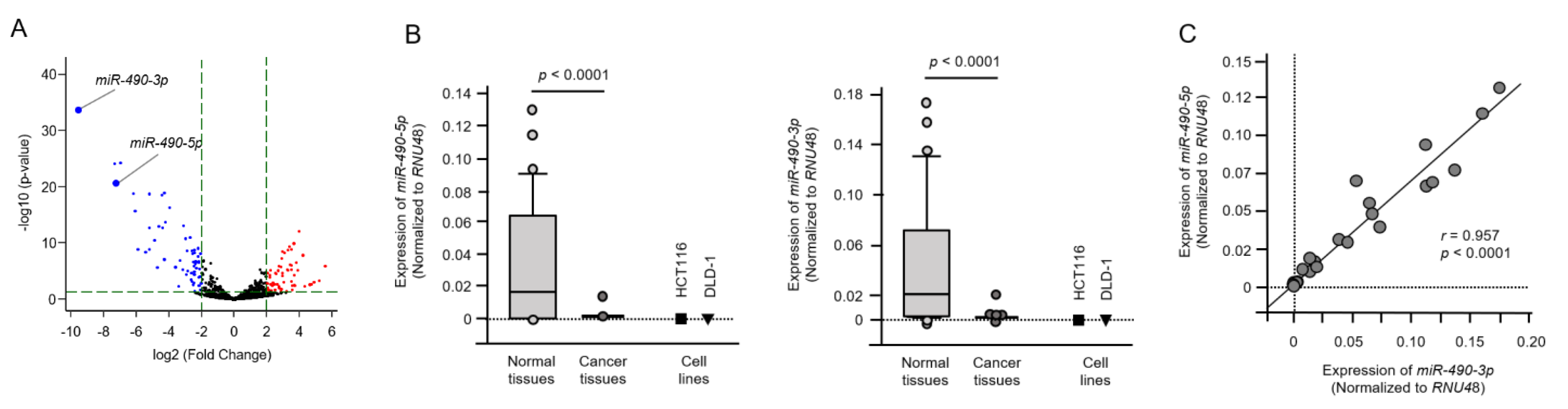
2.2. Tumor-Suppressive Roles of miR-490-5p and miR-490-3p in CRC Cells
2.3. MiR-490-3p and miR-490-5p Was Incorporated into the RNA-Induced Silencing Complex (RISC) in CRC Cells
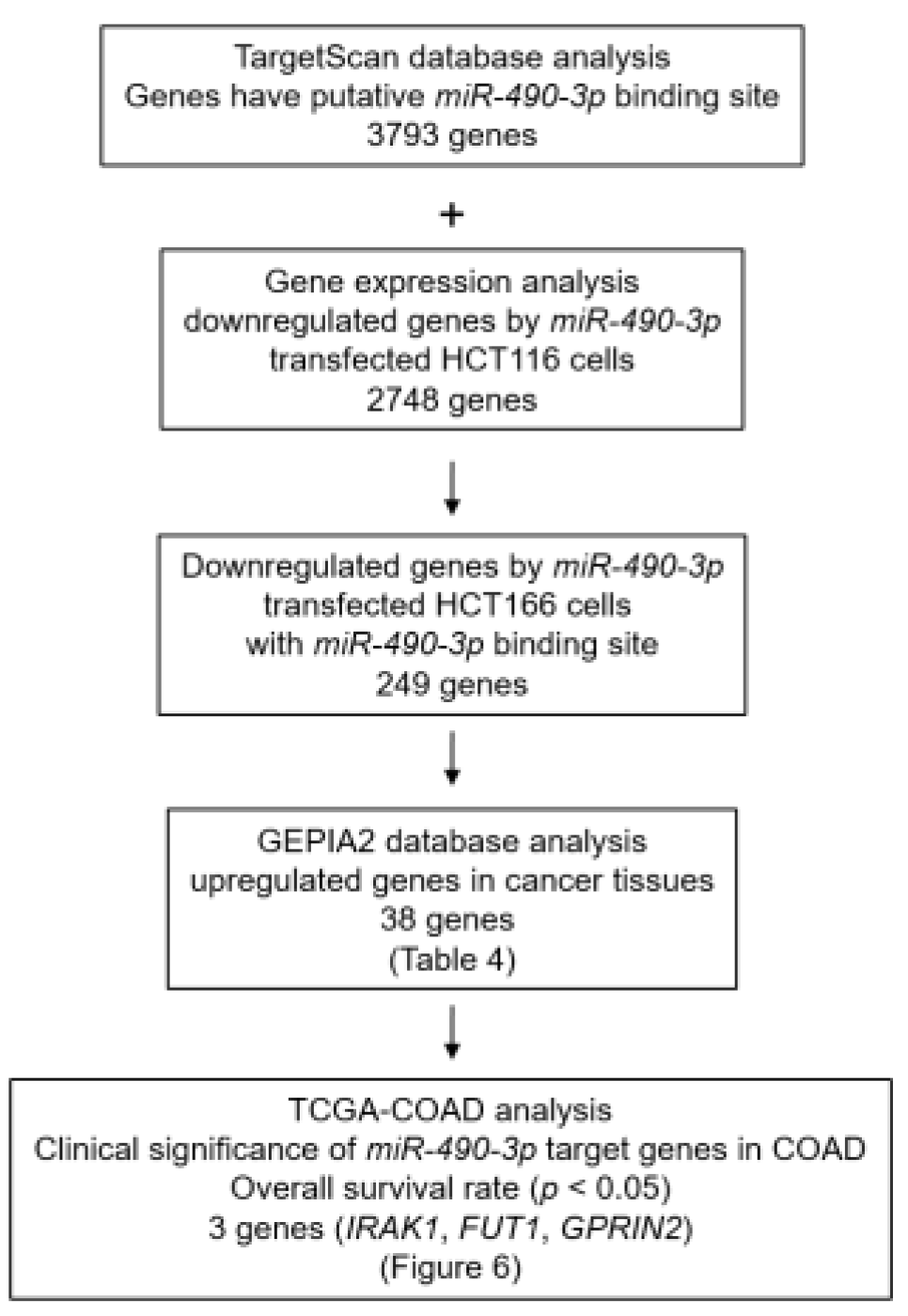
2.4. Screening of Putative Oncogenic Targets by miR-490-3p Regulation in CRC Cells
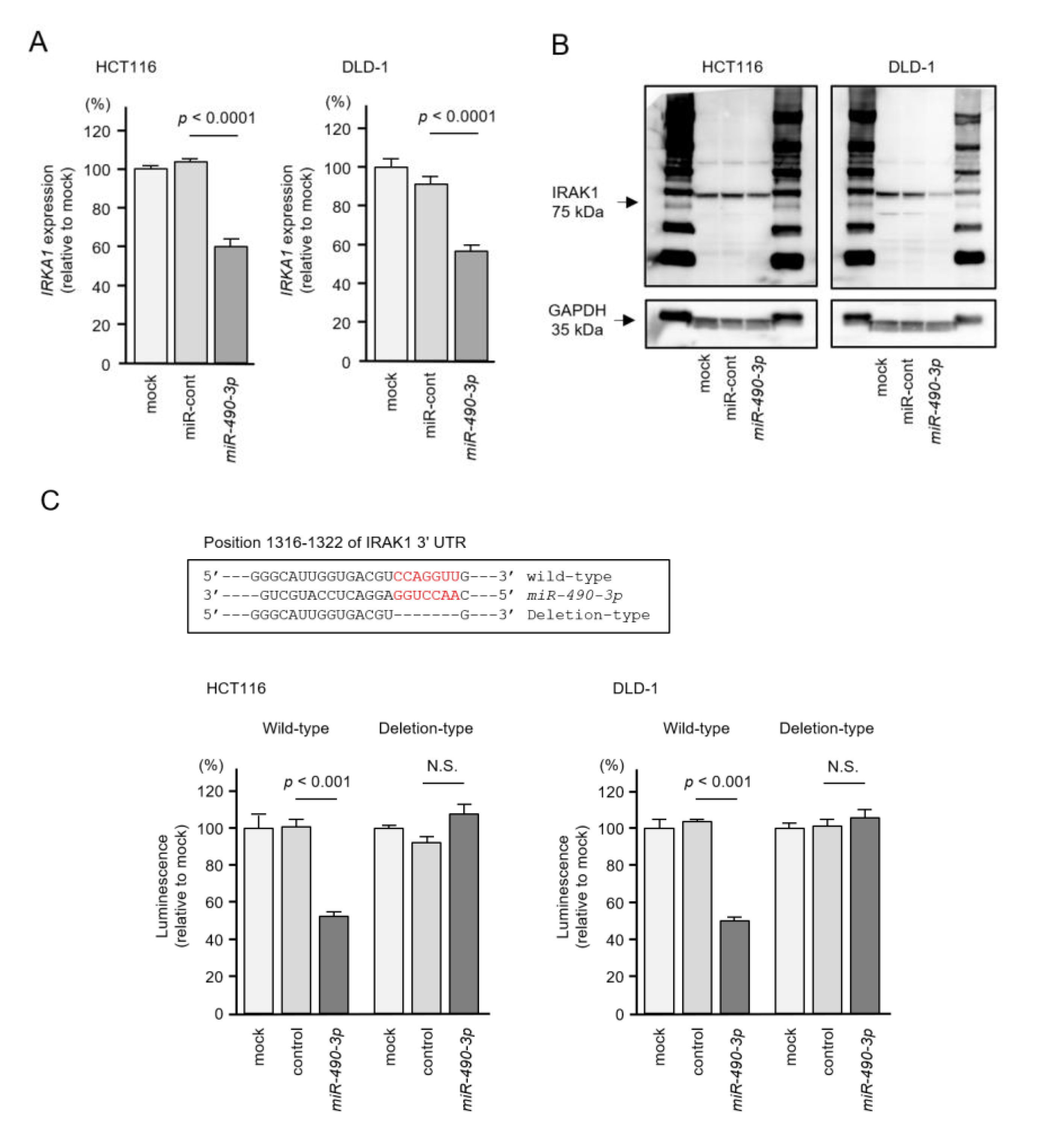
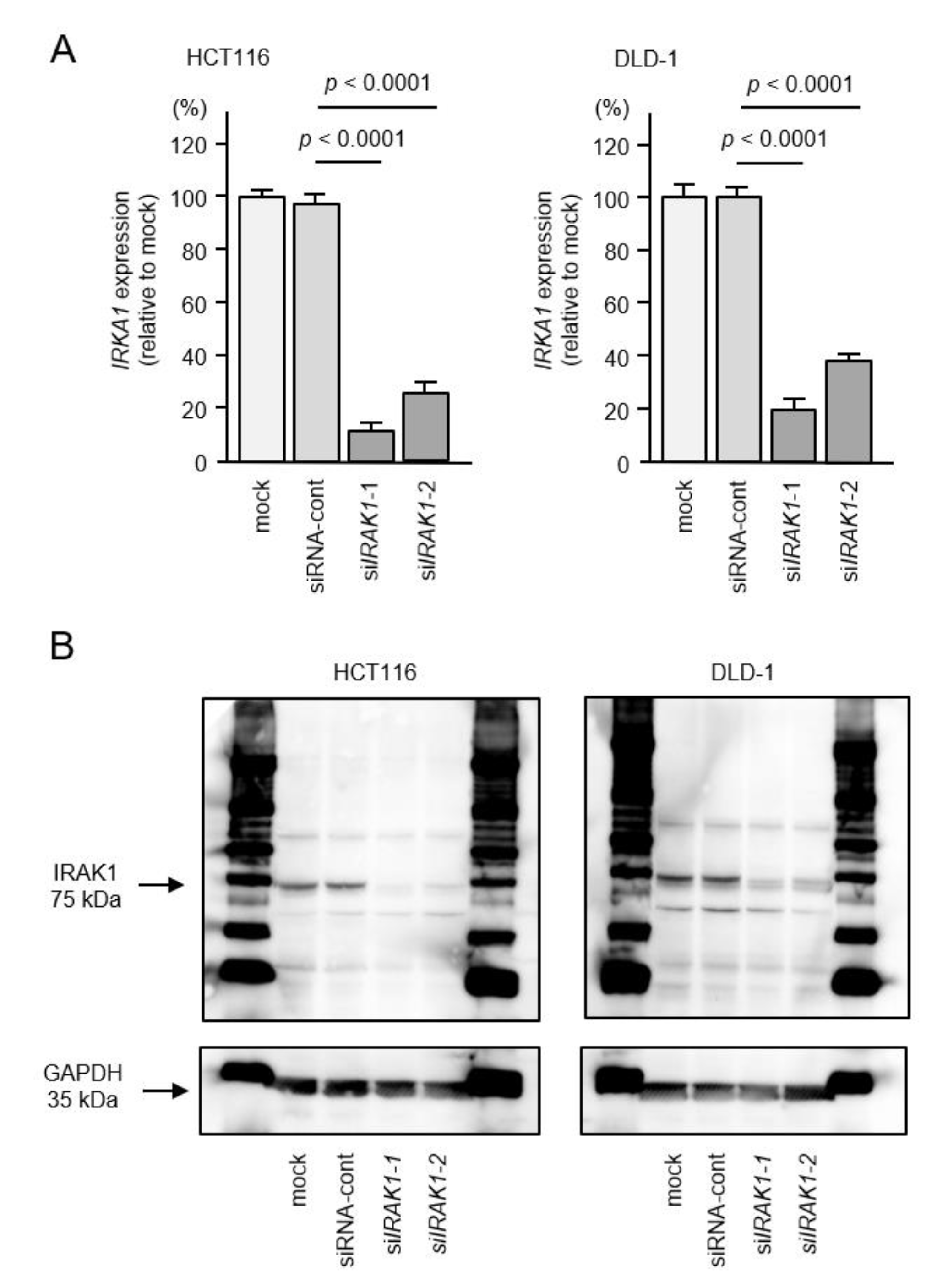
2.5. Direct Regulation of IRAK1 by miR-490-3p in CRC Cells
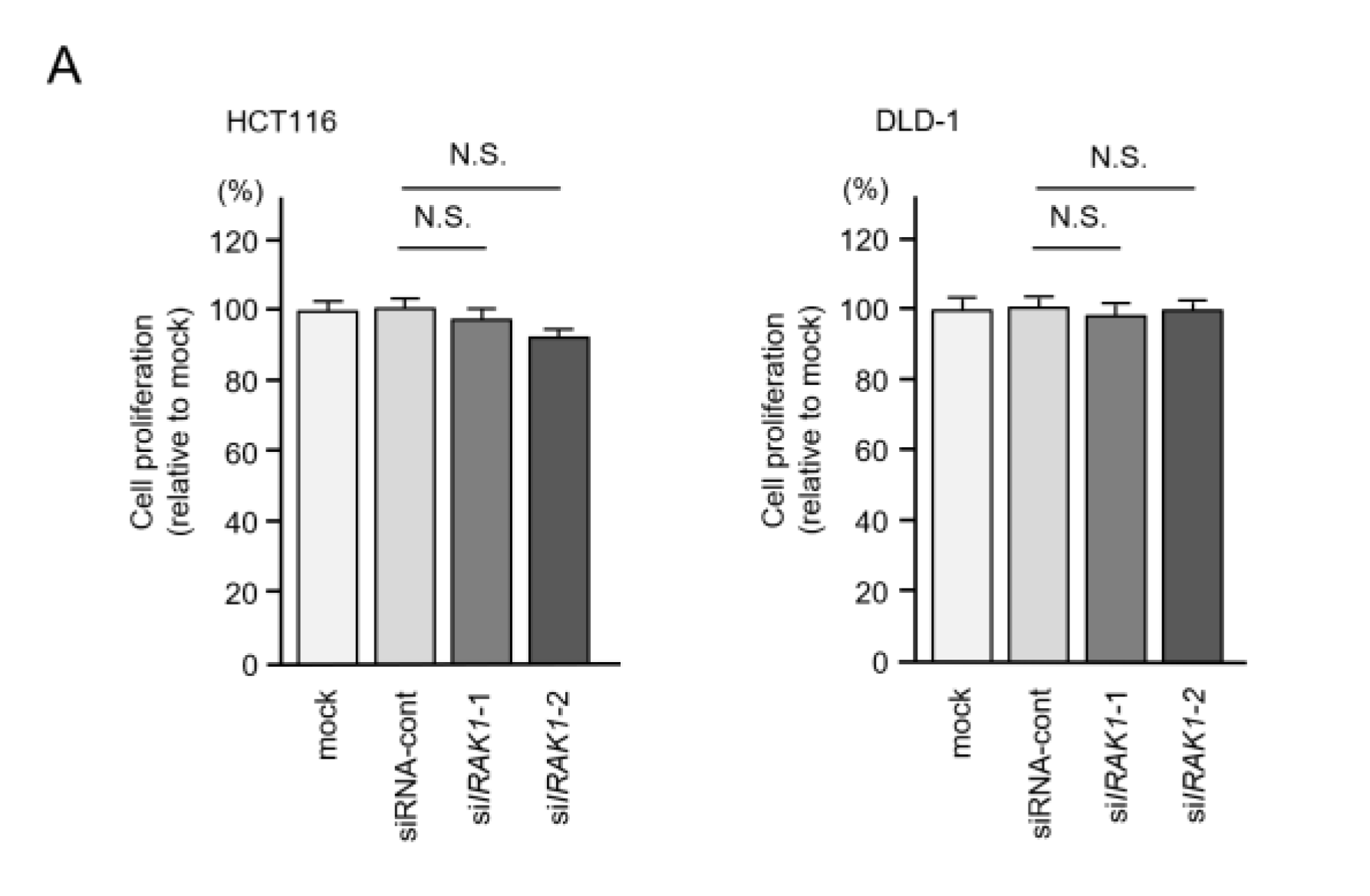
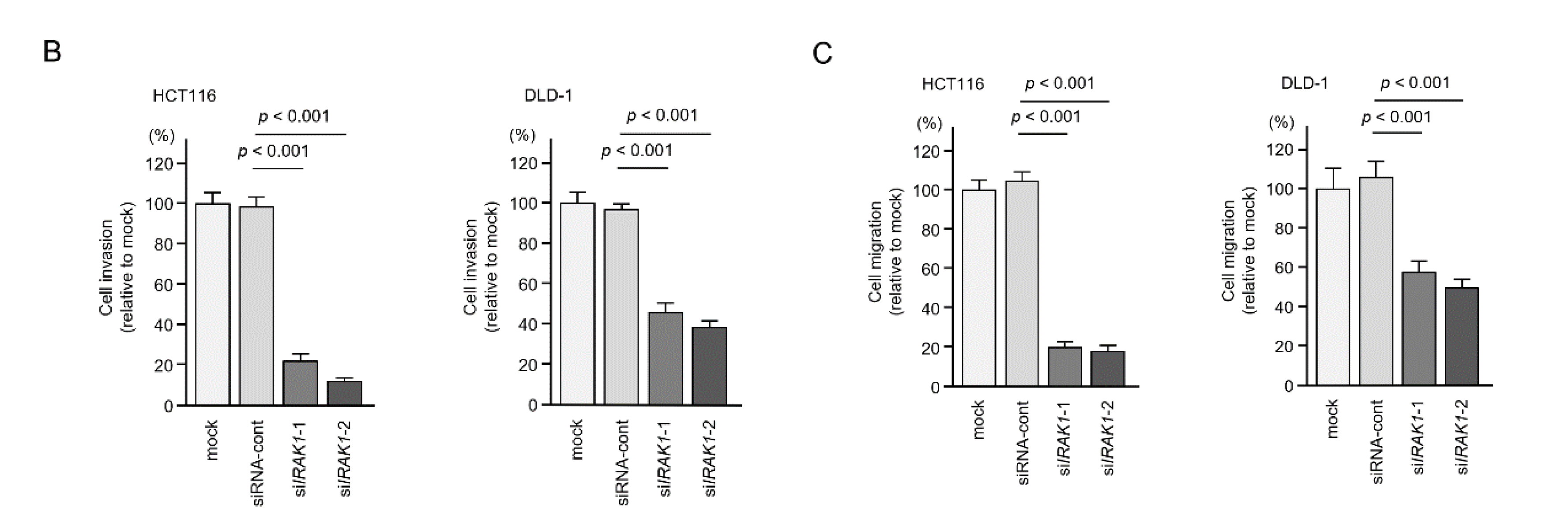
2.6. Effects of IRAK1 Knockdown in CRC Cells
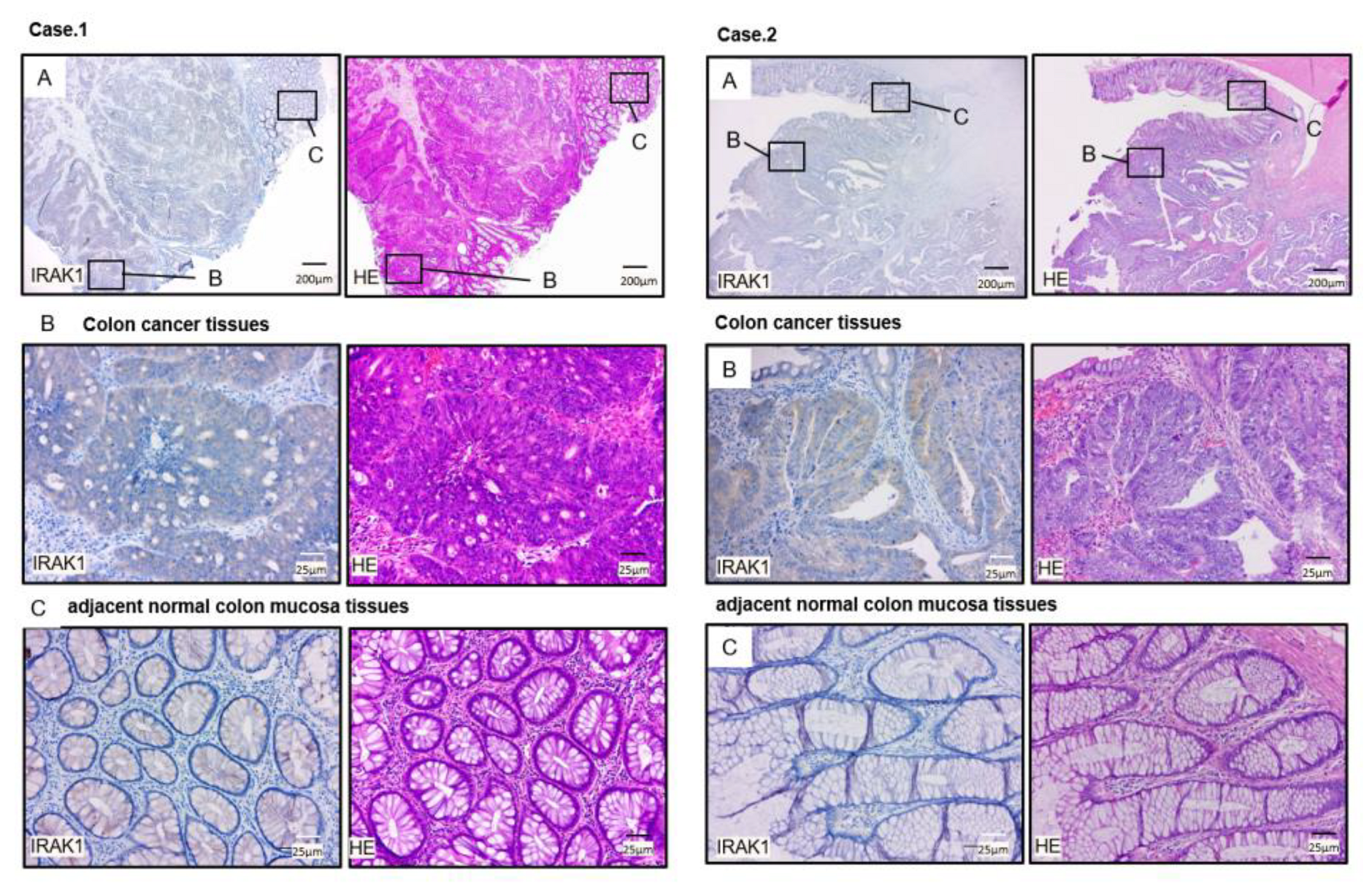
2.7. Expression of IRAK1 in CRC Clinical Specimens
3. Discussion
4. Materials and Methods
4.1. Patient Samples
4.2. CRC Cell Lines and Cell Culture
4.3. Small RNA Sequencing
4.4. RNA Extraction and qPCR
4.5. Transfection of Mature miRNAs, Small-Interfering RNAs, and Plasmid Vectors into CRC Cells
4.6. Cell Proliferation, Migration and Invasion Assays in CRC Cells
4.7. Assay of miR-490-3p Incorporation into the RNA-Induced Silencing Complex (RISC)
4.8. Candidate Target Genes Controlled by miR-490-3p in CRC Cells
4.9. Dual-Luciferase Reporter Assays
4.10. Western Blotting and Immunohistochemistry
4.11. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: Globocan estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [Green Version]
- Dekker, E.; Tanis, P.J.; Vleugels, J.L.A.; Kasi, P.M.; Wallace, M.B. Colorectal cancer. Lancet 2019, 394, 1467–1480. [Google Scholar] [CrossRef]
- Siegel, R.L.; Miller, K.D.; Goding Sauer, A.; Fedewa, S.A.; Butterly, L.F.; Anderson, J.C.; Cercek, A.; Smith, R.A.; Jemal, A. Colorectal cancer statistics, 2020. CA Cancer J. Clin. 2020, 70, 145–164. [Google Scholar] [CrossRef] [Green Version]
- Carlomagno, C.; De Stefano, A.; Rosanova, M.; De Falco, S.; Attademo, L.; Fiore, G.; De Placido, S. Multiple treatment lines and prognosis in metastatic colorectal cancer patients. Cancer Metastasis Rev. 2019, 38, 307–313. [Google Scholar] [CrossRef] [PubMed]
- Leslie, A.; Carey, F.A.; Pratt, N.R.; Steele, R.J. The colorectal adenoma–carcinoma sequence. BJS 2002, 89, 845–860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grady, W.M.; Markowitz, S.D. The Molecular Pathogenesis of Colorectal Cancer and Its Potential Application to Colorectal Cancer Screening. Dig. Dis. Sci. 2015, 60, 762–772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- The Cancer Genome Atlas Network. Comprehensive molecular characterization of human colon and rectal cancer. Nature 2012, 487, 330–337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bala, P.; Singh, A.K.; Kavadipula, P.; Kotapalli, V.; Sabarinathan, R.; Bashyam, M.D. Exome sequencing identifies ARID2 as a novel tumor suppressor in early-onset sporadic rectal cancer. Oncogene 2021, 40, 863–874. [Google Scholar] [CrossRef]
- Anfossi, S.; Babayan, A.; Pantel, K.; Calin, G.A. Clinical utility of circulating non-coding RNAs—An update. Nat. Rev. Clin. Oncol. 2018, 15, 541–563. [Google Scholar] [CrossRef]
- Esteller, M. Non-coding RNAs in human disease. Nat. Rev. Genet. 2011, 12, 861–874. [Google Scholar] [CrossRef] [PubMed]
- Panni, S.; Lovering, R.C.; Porras, P.; Orchard, S. Non-coding RNA regulatory networks. Biochim. Biophys. Acta Bioenerg. 2020, 1863, 194417. [Google Scholar] [CrossRef]
- Anastasiadou, E.; Jacob, L.S.; Slack, F.J. Non-coding RNA networks in cancer. Nat. Rev. Cancer 2018, 18, 5–18. [Google Scholar] [CrossRef]
- Liu, X.; Hao, L.; Li, D.; Zhu, L.; Hu, S. Long Non-coding RNAs and Their Biological Roles in Plants. Genom. Proteom. Bioinform. 2015, 13, 137–147. [Google Scholar] [CrossRef] [Green Version]
- Ha, M.; Kim, V.N. Regulation of microRNA biogenesis. Nat. Rev. Mol. Cell Biol. 2014, 15, 509–524. [Google Scholar] [CrossRef]
- He, L.; Hannon, G.J. MicroRNAs: Small RNAs with a big role in gene regulation. Nat. Rev. Genet. 2004, 5, 522–531. [Google Scholar] [CrossRef]
- Idichi, T.; Seki, N.; Kurahara, H.; Fukuhisa, H.; Toda, H.; Shimonosono, M.; Yamada, Y.; Arai, T.; Kita, Y.; Kijima, Y.; et al. Involvement of anti-tumor miR-124-3p and its targets in the pathogenesis of pancreatic ductal adenocarcinoma: Direct regulation of ITGA3 and ITGB1 by miR-124-3p. Oncotarget 2018, 9, 28849–28865. [Google Scholar] [CrossRef]
- Goto, Y.; Kojima, S.; Nishikawa, R.; Enokida, H.; Chiyomaru, T.; Kinoshita, T.; Nakagawa, M.; Naya, Y.; Ichikawa, T.; Seki, N. The microRNA-23b/27b/24-1 cluster is a disease progression marker and tumor suppressor in prostate cancer. Oncotarget 2014, 5, 7748–7759. [Google Scholar] [CrossRef] [Green Version]
- Xie, N.; Meng, Q.; Zhang, Y.; Luo, Z.; Xue, F.; Liu, S.; Li, Y.; Huang, Y. MicroRNA-142-3p suppresses cell proliferation, invasion and epithelial-to-mesenchymal transition via RAC1-ERK1/2 signaling in colorectal cancer. Mol. Med. Rep. 2021, 24, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Wada, M.; Goto, Y.; Tanaka, T.; Okada, R.; Moriya, S.; Idichi, T.; Noda, M.; Sasaki, K.; Kita, Y.; Kurahara, H.; et al. RNA sequencing-based microRNA expression signature in esophageal squamous cell carcinoma: Oncogenic targets by antitumor miR-143-5p and miR-143-3p regulation. J. Hum. Genet. 2020, 65, 1019–1034. [Google Scholar] [CrossRef] [PubMed]
- Toda, H.; Kurozumi, S.; Kijima, Y.; Idichi, T.; Shinden, Y.; Yamada, Y.; Arai, T.; Maemura, K.; Fujii, T.; Horiguchi, J.; et al. Molecular pathogenesis of triple-negative breast cancer based on microRNA expression signatures: Antitumor miR-204-5p targets AP1S3. J. Hum. Genet. 2018, 63, 1197–1210. [Google Scholar] [CrossRef] [PubMed]
- Yonemori, K.; Seki, N.; Idichi, T.; Kurahara, H.; Osako, Y.; Koshizuka, K.; Arai, T.; Okato, A.; Kita, Y.; Arigami, T.; et al. The microRNA expression signature of pancreatic ductal adenocarcinoma by RNA sequencing: Anti-tumour functions of the microRNA-216 cluster. Oncotarget 2017, 8, 70097–70115. [Google Scholar] [CrossRef] [Green Version]
- Pantaleão, L.C.; Ozanne, S. Small RNA Sequencing: A Technique for miRNA Profiling. Methods Mol. Biol. 2018, 1735, 321–330. [Google Scholar] [CrossRef] [PubMed]
- Xing, X.L.; Yao, Z.Y.; Zhang, T.; Zhu, N.; Liu, Y.W.; Peng, J. MicroRNA-Related Prognosis Biomarkers from High-Throughput Sequencing Data of Colorectal Cancer. Biomed. Res. Int. 2020, 2020, 7905380. [Google Scholar] [CrossRef]
- Neerincx, M.; Sie, D.; Buffart, T.E.; Verheul, H.M.W.; Van Der Wiel, M.A.; Van Grieken, N.C.T.; Burggraaf, J.D.; Dekker, H.; Eijk, P.P.; Ylstra, B.; et al. MiR expression profiles of paired primary colorectal cancer and metastases by next-generation sequencing. Oncogenesis 2015, 4, e170. [Google Scholar] [CrossRef] [Green Version]
- Hamfjord, J.; Stangeland, A.M.; Hughes, T.; Skrede, M.L.; Tveit, K.M.; Ikdahl, T.; Kure, E.H. Differential Expression of miRNAs in Colorectal Cancer: Comparison of Paired Tumor Tissue and Adjacent Normal Mucosa Using High-Throughput Sequencing. PLoS ONE 2012, 7, e34150. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Okada, R.; Hozaka, Y.; Wada, M.; Moriya, S.; Satake, S.; Idichi, T.; Kurahara, H.; Ohtsuka, T.; Seki, N. Molecular Pathogenesis of Pancreatic Ductal Adenocarcinoma: Impact of miR-30c-5p and miR-30c-2-3p Regulation on Oncogenic Genes. Cancers 2020, 12, 2731. [Google Scholar] [CrossRef]
- Kawagoe, K.; Wada, M.; Idichi, T.; Okada, R.; Yamada, Y.; Moriya, S.; Okubo, K.; Matsushita, D.; Arigami, T.; Kurahara, H.; et al. Regulation of aberrantly expressed SERPINH1 by antitumor miR-148a-5p inhibits cancer cell aggressiveness in gastric cancer. J. Hum. Genet. 2020, 65, 647–656. [Google Scholar] [CrossRef] [PubMed]
- Del Vecchio, F.; Mastroiaco, V.; Di Marco, A.; Compagnoni, C.; Capece, D.; Zazzeroni, F.; Capalbo, C.; Alesse, E.; Tessitore, A. Next-generation sequencing: Recent applications to the analysis of colorectal cancer. J. Transl. Med. 2017, 15, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Röhr, C.; Kerick, M.; Fischer, A.; Kühn, A.; Kashofer, K.; Timmermann, B.; Daskalaki, A.; Meinel, T.; Drichel, D.; Börno, S.T.; et al. High-Throughput miRNA and mRNA Sequencing of Paired Colorectal Normal, Tumor and Metastasis Tissues and Bioinformatic Modeling of miRNA-1 Therapeutic Applications. PLoS ONE 2013, 8, e67461. [Google Scholar] [CrossRef]
- Lee, J.; Hong, H.K.; Peng, S.B.; Kim, T.W.; Lee, W.Y.; Yun, S.H.; Kim, H.C.; Liu, J.; Ebert, P.J.; Aggarwal, A.; et al. Identifying metastasis-initiating miRNA-target regulations of colorectal cancer from expressional changes in primary tumors. Sci. Rep. 2020, 10, 14919. [Google Scholar] [CrossRef]
- Bandrés, E.; Cubedo, E.; Agirre, X.; Malumbres, R.; Zárate, R.; Ramirez, N.; Abajo, A.; Navarro, A.; Moreno, I.; Monzó, M.; et al. Identification by Real-time PCR of 13 mature microRNAs differentially expressed in colorectal cancer and non-tumoral tissues. Mol. Cancer 2006, 5, 29. [Google Scholar] [CrossRef] [Green Version]
- Sarver, A.L.; French, A.J.; Borralho, P.M.; Thayanithy, V.; Oberg, A.L.; Silverstein, K.A.; Morlan, B.W.; Riska, S.M.; Boardman, L.A.; Cunningham, J.M.; et al. Human colon cancer profiles show differential microRNA expression depending on mismatch repair status and are characteristic of undifferentiated proliferative states. BMC Cancer 2009, 9, 401. [Google Scholar] [CrossRef] [Green Version]
- Schetter, A.J.; Leung, S.Y.; Sohn, J.J.; Zanetti, K.A.; Bowman, E.D.; Yanaihara, N.; Yuen, S.T.; Chan, T.L.; Kwong, D.L.W.; Au, G.K.H.; et al. MicroRNA Expression Profiles Associated with Prognosis and Therapeutic Outcome in Colon Adenocarcinoma. JAMA 2008, 299, 425–436. [Google Scholar] [CrossRef] [Green Version]
- Cummins, J.M.; He, Y.; Leary, R.J.; Pagliarini, R.; Diaz, L.; Sjoblom, T.; Barad, O.; Bentwich, Z.; Szafranska, A.E.; Labourier, E.; et al. The colorectal microRNAome. Proc. Natl. Acad. Sci. USA 2006, 103, 3687–3692. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Mo, R.; Zheng, L. MicroRNA-490-3p inhibits migration and chemoresistance of colorectal cancer cells via targeting TNKS2. World J. Surg. Oncol. 2021, 19, 117. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; He, B.; Xu, T.; Pan, Y.; Hu, X.; Chen, X.; Wang, S. MiR-490-3p Functions as a Tumor Suppressor by Inhibiting Oncogene VDAC1 Expression in Colorectal Cancer. J. Cancer 2018, 9, 1218–1230. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Guo, J.; Zhou, J.; Wang, D.; Kang, X.; Zhou, L. NF-κB maintains the stemness of colon cancer cells by downregulating miR-195-5p/497-5p and upregulating MCM2. J. Exp. Clin. Cancer Res. 2020, 39, 225. [Google Scholar] [CrossRef]
- Koduru, S.V.; Tiwari, A.K.; Hazard, S.W.; Mahajan, M.; Ravnic, D.J. Exploration of small RNA-seq data for small non-coding RNAs in Human Colorectal Cancer. Int. J. Genom. 2017, 5, 16–31. [Google Scholar] [CrossRef] [Green Version]
- Yu, D.; Liu, H.; Qin, J.; Huangfu, M.; Guan, X.; Li, X.; Zhou, L.; Dou, T.; Liu, Y.; Wang, L.; et al. Curcumol inhibits the viability and invasion of colorectal cancer cells via miR-30a-5p and Hippo signaling pathway. Oncol. Lett. 2021, 21, 1–13. [Google Scholar] [CrossRef]
- Mitra, R.; Adams, C.M.; Jiang, W.; Greenawalt, E.; Eischen, C.M. Pan-cancer analysis reveals cooperativity of both strands of microRNA that regulate tumorigenesis and patient survival. Nat. Commun. 2020, 11, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Idichi, T.; Seki, N.; Kurahara, H.; Fukuhisa, H.; Toda, H.; Shimonosono, M.; Okato, A.; Arai, T.; Kita, Y.; Mataki, Y.; et al. Molecular pathogenesis of pancreatic ductal adenocarcinoma: Impact of passenger strand of pre-miR-148a on gene regulation. Cancer Sci. 2018, 109, 2013–2026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimomura, H.; Okada, R.; Tanaka, T.; Hozaka, Y.; Wada, M.; Moriya, S.; Idichi, T.; Kita, Y.; Kurahara, H.; Ohtsuka, T.; et al. Role of miR-30a-3p Regulation of Oncogenic Targets in Pancreatic Ductal Adenocarcinoma Pathogenesis. Int. J. Mol. Sci. 2020, 21, 6459. [Google Scholar] [CrossRef]
- Li, Y.; Tian, D.; Chen, H.; Cai, Y.; Chen, S.; Duan, S. MicroRNA-490-3p and -490-5p in carcinogenesis: Separate or the same goal? Oncol. Lett. 2021, 22, 678. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Zhao, L. MicroRNA-490 inhibits tumorigenesis and progression in breast cancer. Oncol. Targets Ther. 2016, 9, 4505–4516. [Google Scholar] [CrossRef] [Green Version]
- Jia, Z.; Liu, Y.; Gao, Q.; Han, Y.; Zhang, G.; Xu, S.; Cheng, K.; Zou, W. miR-490-3p inhibits the growth and invasiveness in triple-negative breast cancer by repressing the expression of TNKS2. Gene 2016, 593, 41–47. [Google Scholar] [CrossRef]
- Li, Z.; Jiang, D.; Yang, S. MiR-490-3p Inhibits the Malignant Progression of Lung Adenocarcinoma. Cancer Manag. Res. 2020, 12, 10975–10984. [Google Scholar] [CrossRef]
- Xu, X.; Chen, R.; Li, Z.; Huang, N.; Wu, X.; Li, S.; Li, Y.; Wu, S. MicroRNA-490-3p inhibits colorectal cancer metastasis by targeting TGFβR1. BMC Cancer 2015, 15, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.; Yin, M.; Cheng, C.; Jiang, H.; Jiang, K.; Shen, Z.; Ye, Y.; Wang, S. Decreased expression of miR-490-3p in colorectal cancer predicts poor prognosis and promotes cell proliferation and invasion by targeting RAB14. Int. J. Oncol. 2018, 53, 1247–1256. [Google Scholar] [CrossRef]
- Flannery, S.; Bowie, A.G. The interleukin-1 receptor-associated kinases: Critical regulators of innate immune signalling. Biochem. Pharmacol. 2010, 80, 1981–1991. [Google Scholar] [CrossRef]
- Rhyasen, G.W.; Starczynowski, D.T. IRAK signalling in cancer. Br. J. Cancer 2015, 112, 232–237. [Google Scholar] [CrossRef] [Green Version]
- Wee, Z.N.; Yatim, S.M.; Kohlbauer, V.K.; Feng, M.; Goh, J.Y.; Bao, Y.; Lee, P.L.; Zhang, S.; Wang, P.P.; Lim, E.; et al. IRAK1 is a therapeutic target that drives breast cancer metastasis and resistance to paclitaxel. Nat. Commun. 2015, 6, 8746. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, Y.; Duan, X.; Wang, Y.; Zhang, Z. Interleukin-1 receptor-associated kinase 1 correlates with metastasis and invasion in endometrial carcinoma. J. Cell Biochem. 2018, 119, 2545–2555. [Google Scholar] [CrossRef]
- Adams, A.K.; Bolanos, L.C.; Dexheimer, P.J.; Karns, R.A.; Aronow, B.J.; Komurov, K.; Jegga, A.G.; Casper, K.A.; Patil, Y.J.; Wilson, K.M.; et al. IRAK1 is a novel DEK transcriptional target and is essential for head and neck cancer cell survival. Oncotarget 2015, 6, 43395–43407. [Google Scholar] [CrossRef] [Green Version]
- Singer, J.W.; Fleischman, A.; Al-Fayoumi, S.; Mascarenhas, J.O.; Yu, Q.; Agarwal, A. Inhibition of interleukin-1 receptor-associated kinase 1 (IRAK1) as a therapeutic strategy. Oncotarget 2018, 9, 33416–33439. [Google Scholar] [CrossRef] [Green Version]
- Feng, Z.; Duan, Z.; Shi, G.; Wang, Q.; Zhou, J.; Chen, Y. Pharmacological inhibition of IRAK1 attenuates colitis-induced tumorigenesis in mice by inhibiting the inflammatory response and epithelial–mesenchymal transition. J. Biochem. Mol. Toxicol. 2021, e22838. [Google Scholar] [CrossRef]
- Toda, H.; Seki, N.; Kurozumi, S.; Shinden, Y.; Yamada, Y.; Nohata, N.; Moriya, S.; Idichi, T.; Maemura, K.; Fujii, T.; et al. RNA-sequence-based microRNA expression signature in breast cancer: Tumor-suppressive miR-101-5p regulates molecular pathogenesis. Mol. Oncol. 2020, 14, 426–446. [Google Scholar] [CrossRef] [Green Version]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Age (Years) | Sex | Location | Differentiation | T | N | M | Stage | ly | v | Recurrence |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 66 | Male | Rectum | Moderate | 3 | 1a | 0 | IIIB | 0 | 1 | — |
| 2 | 66 | Male | Colon(S) | Moderate | 3 | 1a | 0 | IIIB | 1 | 1 | — |
| 3 | 79 | Male | Rectum | Moderate | 3 | 2a | 0 | IIIB | 1 | 1 | — |
| 4 | 78 | Female | Colon(S) | Moderate | 3 | 0 | 0 | IIA | 0 | 1 | — |
| 5 | 83 | Female | Colon(S) | Moderate | 3 | 1a | 0 | IIIB | 1 | 0 | — |
| Entrez Gene ID | Gene Symbol | Gene Name | Binding Sites | HCT116 miR-490-3p Transfectant log2 FC ≤ 1 | OncoLnc 5 yrs (p-Value *) |
|---|---|---|---|---|---|
| 9721 | GPRIN2 | G protein regulated inducer of neurite outgrowth 2 | 1 | −2.3130598 | 0.008 |
| 3654 | IRAK1 | interleukin-1 receptor-associated kinase 1 | 1 | −1.0598125 | 0.0427 |
| 2523 | FUT1 | fucosyltransferase 1 (galactoside 2-alpha-L-fucosyltransferase, H blood group) | 1 | −1.5235968 | 0.0468 |
| 135112 | NCOA7 | nuclear receptor coactivator 7 | 1 | −1.177208 | 0.0585 |
| 84061 | MAGT1 | magnesium transporter 1 | 1 | −1.5050192 | 0.0799 |
| 6382 | SDC1 | syndecan 1 | 1 | −1.0336791 | 0.088 |
| 3635 | INPP5D | inositol polyphosphate-5-phosphatase, 145 kDa | 1 | −1.2042127 | 0.1059 |
| 3178 | HNRNPA1 | heterogeneous nuclear ribonucleoprotein A1 | 3 | −1.6466646 | 0.1496 |
| 84152 | PPP1R1B | protein phosphatase 1, regulatory (inhibitor) subunit 1B | 1 | −2.459818 | 0.163 |
| 56886 | UGGT1 | UDP-glucose glycoprotein glucosyltransferase 1 | 1 | −1.0872145 | 0.2326 |
| 8529 | CYP4F2 | cytochrome P450, family 4, subfamily F, polypeptide 2 | 1 | −2.5992675 | 0.2469 |
| 647024 | C6orf132 | chromosome 6 open reading frame 132 | 2 | −1.2631998 | 0.3329 |
| 10525 | HYOU1 | hypoxia up-regulated 1 | 2 | −1.6224588 | 0.3420 |
| 2444 | FRK | fyn-related kinase | 2 | −1.2127504 | 0.3546 |
| 154796 | AMOT | angiomotin | 1 | −1.0376037 | 0.3709 |
| 23446 | SLC44A1 | solute carrier family 44 (choline transporter), member 1 | 2 | −1.1456499 | 0.4167 |
| 440145 | MZT1 | mitotic spindle organizing protein 1 | 1 | −1.0596924 | 0.4636 |
| 28985 | MCTS1 | malignant T cell amplified sequence 1 | 1 | −1.0754105 | 0.4927 |
| 4642 | MYO1D | myosin ID | 1 | −1.1291242 | 0.5085 |
| 84187 | TMEM164 | transmembrane protein 164 | 1 | −1.0723546 | 0.5172 |
| 80223 | RAB11FIP1 | RAB11 family interacting protein 1 (class I) | 2 | −1.374427 | 0.5477 |
| 8635 | RNASET2 | ribonuclease T2 | 1 | −1.4826338 | 0.5618 |
| 3146 | HMGB1 | high mobility group box 1 | 1 | −1.8401318 | 0.5719 |
| 340706 | VWA2 | von Willebrand factor A domain containing 2 | 1 | −1.3371019 | 0.6107 |
| 51561 | IL23A | interleukin 23, alpha subunit p19 | 1 | −1.1564418 | 0.6474 |
| 6850 | SYK | spleen tyrosine kinase | 1 | −1.065637 | 0.6535 |
| 2232 | FDXR | ferredoxin reductase | 1 | −1.0080034 | 0.7008 |
| 2525 | FUT3 | fucosyltransferase 3 (galactoside 3(4)-L-fucosyltransferase, Lewis blood group) | 1 | −2.2499368 | 0.7117 |
| 7416 | VDAC1 | voltage-dependent anion channel 1 | 2 | −1.3866558 | 0.7136 |
| 80201 | HKDC1 | hexokinase domain containing 1 | 1 | −2.3432722 | 0.7136 |
| 196264 | MPZL3 | myelin protein zero-like 3 | 1 | −1.0834669 | 0.7250 |
| 3703 | STT3A | STT3A, subunit of the oligosaccharyltransferase complex (catalytic) | 1 | −1.6853191 | 0.7465 |
| 7039 | TGFA | transforming growth factor, alpha | 2 | −1.0281087 | 0.7569 |
| 4494 | MT1F | metallothionein 1F | 1 | −1.7403115 | 0.7756 |
| 6523 | SLC5A1 | solute carrier family 5 (sodium/glucose cotransporter), member 1 | 1 | −2.3984282 | 0.8507 |
| 6653 | SORL1 | sortilin-related receptor, L (DLR class) A repeats containing | 1 | −1.0742446 | 0.853 |
| 201595 | STT3B | STT3B, subunit of the oligosaccharyltransferase complex (catalytic) | 1 | −2.0069218 | 0.0046 |
| 345079 | SOWAHB | sosondowah ankyrin repeat domain family member B | 1 | −1.8673139 | 0.0519 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hozaka, Y.; Kita, Y.; Yasudome, R.; Tanaka, T.; Wada, M.; Idichi, T.; Tanabe, K.; Asai, S.; Moriya, S.; Toda, H.; et al. RNA-Sequencing Based microRNA Expression Signature of Colorectal Cancer: The Impact of Oncogenic Targets Regulated by miR-490-3p. Int. J. Mol. Sci. 2021, 22, 9876. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22189876
Hozaka Y, Kita Y, Yasudome R, Tanaka T, Wada M, Idichi T, Tanabe K, Asai S, Moriya S, Toda H, et al. RNA-Sequencing Based microRNA Expression Signature of Colorectal Cancer: The Impact of Oncogenic Targets Regulated by miR-490-3p. International Journal of Molecular Sciences. 2021; 22(18):9876. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22189876
Chicago/Turabian StyleHozaka, Yuto, Yoshiaki Kita, Ryutaro Yasudome, Takako Tanaka, Masumi Wada, Tetsuya Idichi, Kan Tanabe, Shunichi Asai, Shogo Moriya, Hiroko Toda, and et al. 2021. "RNA-Sequencing Based microRNA Expression Signature of Colorectal Cancer: The Impact of Oncogenic Targets Regulated by miR-490-3p" International Journal of Molecular Sciences 22, no. 18: 9876. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22189876