
Rab11-FIP1 and Rab11-FIP5 Regulate pIgR/pIgA Transcytosis through TRIM21-Mediated Polyubiquitination


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
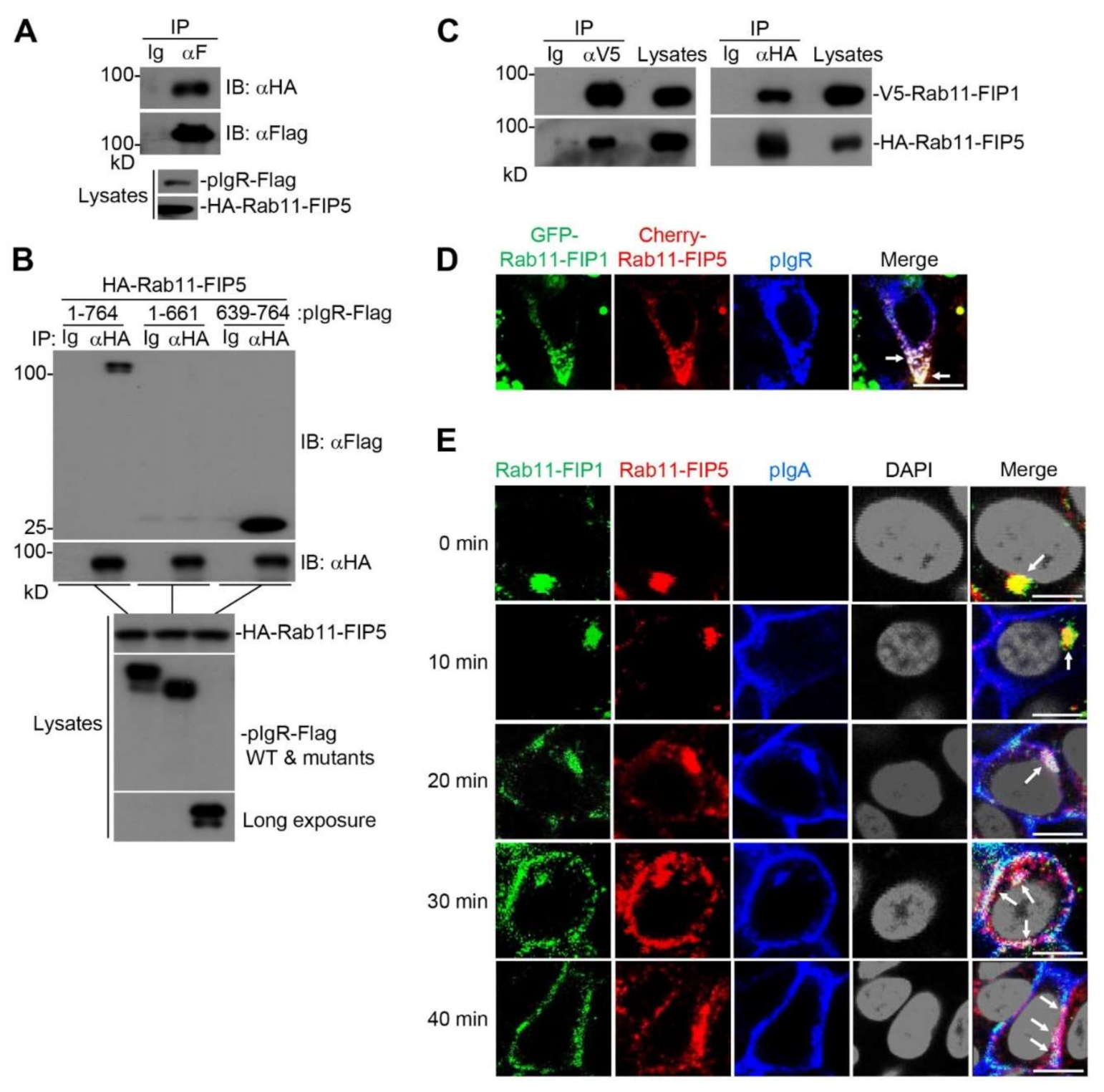
2.1. Rab11-FIP1 Interacts with pIgR
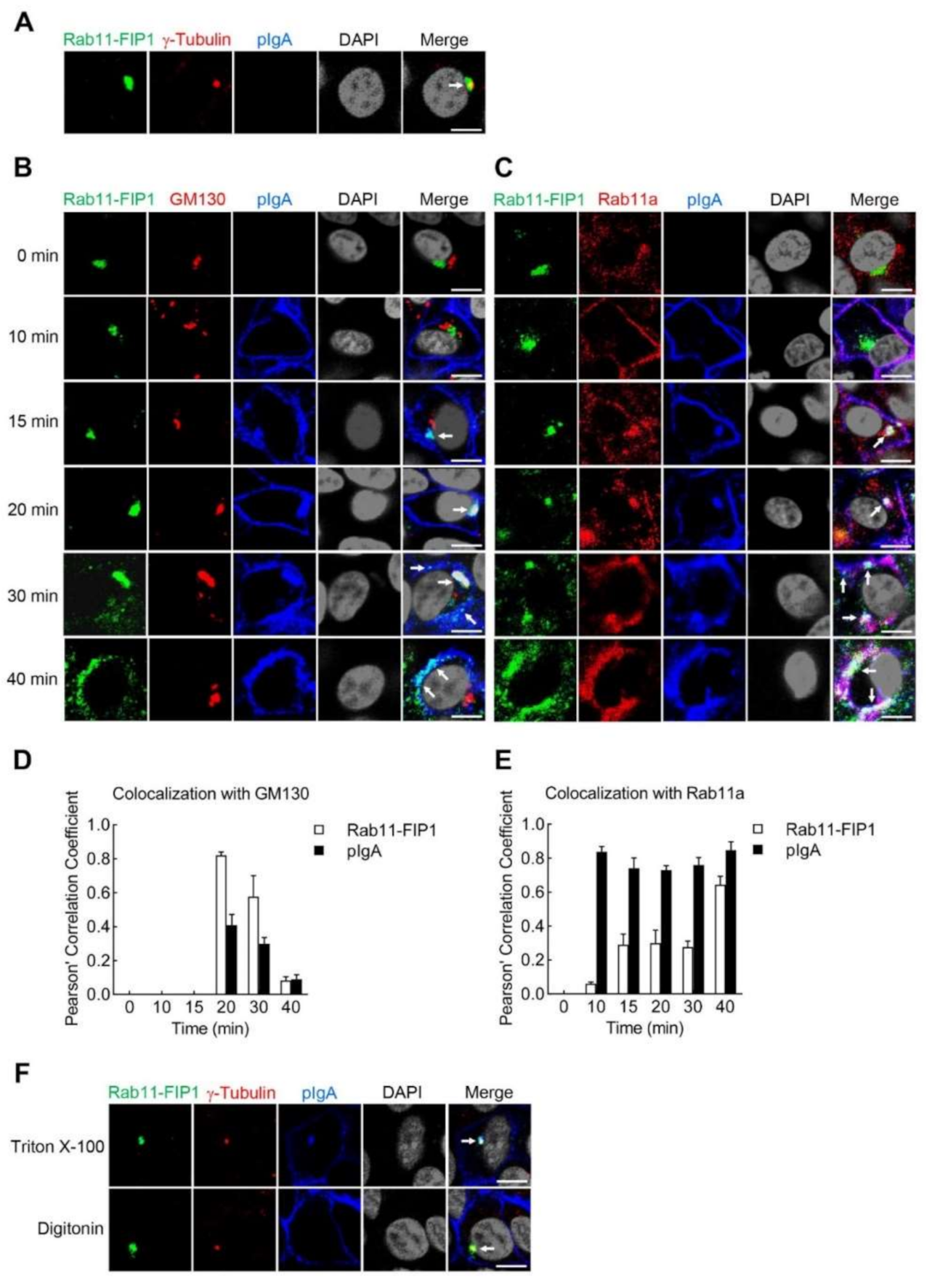
2.2. Rab11-FIP1 Colocalizes with pIgA during pIgA Transcytosis
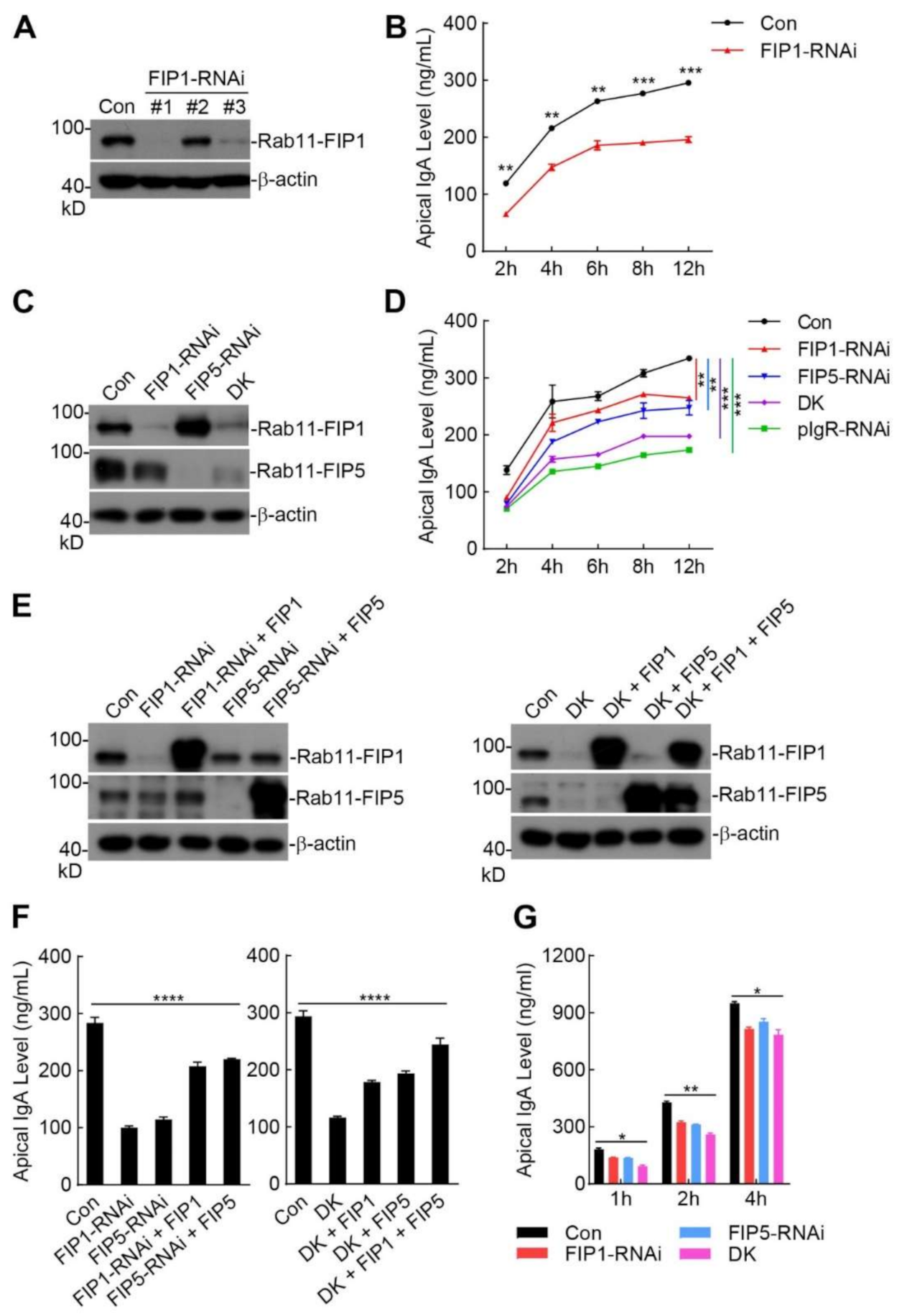
2.3. Rab11-FIP1 and Rab11-FIP5 Additively Promote Extracellular Secretion of pIgA
2.4. Rab11-FIP1, Rab11-FIP5 and pIgR Form a Complex during pIgA Transcytosis
2.5. Rab11-FIP1 Colocalizes with the Transcytosed pIgA from the Vicinity of Centrosome via Golgi Apparatus to Apical Plasma Membrane in Incompletely Polarized Vero-pIgR Cells
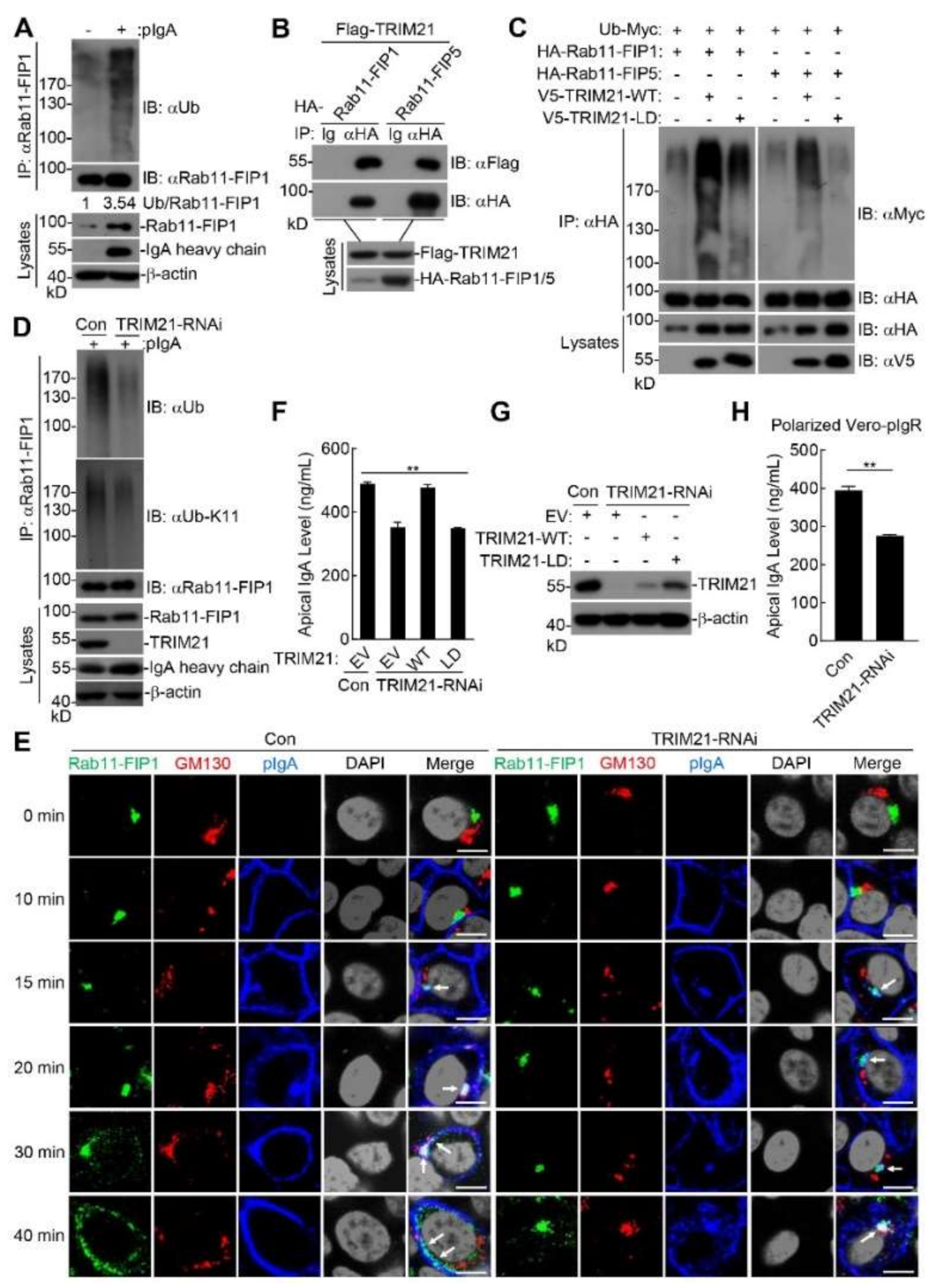
2.6. Polyubiquitinated Rab11-FIP1 and Rab11-FIP5 Mediated by TRIM21 Regulate pIgA Transcytosis
3. Discussion
4. Materials and Methods
4.1. Reagents, Antibodies and Cells
4.2. Constructs
4.3. Affinity Purification of Biotinylated Proteins
4.4. Coimmunoprecipitation and Immunoblotting Analyses
4.5. ELISA
4.6. Confocal Microscopy
4.7. Transepithelial Resistance (TER) Measurements
4.8. Establishment of Stable Cell Lines
4.9. Ubiquitination Assays
4.10. In Vitro Ubiquitination Assay
4.11. Quantitative Real-Time PCR
4.12. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Woof, J.M.; Kerr, M.A. The function of immunoglobulin A in immunity. J. Pathol. 2006, 208, 270–282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mantis, N.J.; Rol, N.; Corthesy, B. Secretory IgA′s complex roles in immunity and mucosal homeostasis in the gut. Mucosal. Immunol. 2011, 4, 603–611. [Google Scholar] [CrossRef] [PubMed]
- Casanova, J.E.; Breitfeld, P.P.; Ross, S.A.; Mostov, K.E. Phosphorylation of the polymeric immunoglobulin receptor required for its efficient transcytosis. Science 1990, 248, 742–745. [Google Scholar] [CrossRef]
- Mostov, K.E.; Deitcher, D.L. Polymeric immunoglobulin receptor expressed in MDCK cells transcytoses IgA. Cell 1986, 46, 613–621. [Google Scholar] [CrossRef]
- Breitfeld, P.P.; Harris, J.M.; Mostov, K.E. Postendocytotic sorting of the ligand for the polymeric immunoglobulin receptor in Madin-Darby canine kidney cells. J. Cell Biol. 1989, 109, 475–486. [Google Scholar] [CrossRef]
- Bidgood, S.R.; Tam, J.C.; McEwan, W.A.; Mallery, D.L.; James, L.C. Translocalized IgA mediates neutralization and stimulates innate immunity inside infected cells. Proc. Natl. Acad. Sci. USA 2014, 111, 13463–13468. [Google Scholar] [CrossRef] [Green Version]
- Kaetzel, C.S. The polymeric immunoglobulin receptor: Bridging innate and adaptive immune responses at mucosal surfaces. Immunol. Rev. 2005, 206, 83–99. [Google Scholar] [CrossRef]
- Asano, M.; Komiyama, K. Polymeric immunoglobulin receptor. J. Oral. Sci. 2011, 53, 147–156. [Google Scholar] [CrossRef] [Green Version]
- Rojas, R.; Apodaca, G. Immunoglobulin transport across polarized epithelial cells. Nat. Rev. Mol. Cell Biol. 2002, 3, 944–955. [Google Scholar] [CrossRef]
- Homma, Y.; Hiragi, S.; Fukuda, M. Rab family of small GTPases: An updated view on their regulation and functions. FEBS J. 2020, 288, 36–45. [Google Scholar] [CrossRef]
- Zerial, M.; McBride, H. Rab proteins as membrane organizers. Nat. Rev. Mol. Cell Biol. 2001, 2, 107–117. [Google Scholar] [CrossRef]
- Lakadamyali, M.; Rust, M.J.; Zhuang, X. Ligands for clathrin-mediated endocytosis are differentially sorted into distinct populations of early endosomes. Cell 2006, 124, 997–1009. [Google Scholar] [CrossRef] [Green Version]
- Sonnichsen, B.; De Renzis, S.; Nielsen, E.; Rietdorf, J.; Zerial, M. Distinct membrane domains on endosomes in the recycling pathway visualized by multicolor imaging of Rab4, Rab5, and Rab11. J. Cell Biol. 2000, 149, 901–914. [Google Scholar] [CrossRef]
- Van Ijzendoorn, S.C. Recycling endosomes. J. Cell Sci. 2006, 119, 1679–1681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakaguchi, A.; Sato, M.; Sato, K.; Gengyo-Ando, K.; Yorimitsu, T.; Nakai, J.; Hara, T.; Sato, K.; Sato, K. REI-1 Is a Guanine Nucleotide Exchange Factor Regulating RAB-11 Localization and Function in C. elegans Embryos. Dev. Cell 2015, 35, 211–221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Kumar, R.; Navarre, J.; Casanova, J.E.; Goldenring, J.R. Regulation of vesicle trafficking in madin-darby canine kidney cells by Rab11a and Rab25. J. Biol. Chem. 2000, 275, 29138–29146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horgan, C.P.; McCaffrey, M.W. The dynamic Rab11-FIPs. Biochem Soc. Trans. 2009, 37, 1032–1036. [Google Scholar] [CrossRef]
- Machesky, L.M. Rab11FIP proteins link endocytic recycling vesicles for cytoskeletal transport and tethering. Biosci. Rep. 2019, 39, BSR20182219. [Google Scholar] [CrossRef]
- Lindsay, A.J.; Hendrick, A.G.; Cantalupo, G.; Senic-Matuglia, F.; Goud, B.; Bucci, C.; McCaffrey, M.W. Rab coupling protein (RCP), a novel Rab4 and Rab11 effector protein. J. Biol. Chem. 2002, 277, 12190–12199. [Google Scholar] [CrossRef] [Green Version]
- Su, T.; Bryant, D.M.; Luton, F.; Verges, M.; Ulrich, S.M.; Hansen, K.C.; Datta, A.; Eastburn, D.J.; Burlingame, A.L.; Shokat, K.M.; et al. A kinase cascade leading to Rab11-FIP5 controls transcytosis of the polymeric immunoglobulin receptor. Nat. Cell Biol. 2010, 12, 1143–1153. [Google Scholar] [CrossRef] [Green Version]
- McRae, R.; Lapierre, L.A.; Manning, E.H.; Goldenring, J.R. Rab11-FIP1 phosphorylation by MARK2 regulates polarity in MDCK cells. Cell Logist. 2017, 7, e1271498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gundry, C.; Marco, S.; Rainero, E.; Miller, B.; Dornier, E.; Mitchell, L.; Caswell, P.T.; Campbell, A.D.; Hogeweg, A.; Sansom, O.J.; et al. Phosphorylation of Rab-coupling protein by LMTK3 controls Rab14-dependent EphA2 trafficking to promote cell:cell repulsion. Nat. Commun. 2017, 8, 14646. [Google Scholar] [CrossRef]
- Caswell, P.T.; Chan, M.; Lindsay, A.J.; McCaffrey, M.W.; Boettiger, D.; Norman, J.C. Rab-coupling protein coordinates recycling of alpha5beta1 integrin and EGFR1 to promote cell migration in 3D microenvironments. J. Cell Biol. 2008, 183, 143–155. [Google Scholar] [CrossRef] [Green Version]
- Foss, S.; Watkinson, R.; Sandlie, I.; James, L.C.; Andersen, J.T. TRIM21: A cytosolic Fc receptor with broad antibody isotype specificity. Immunol. Rev. 2015, 268, 328–339. [Google Scholar] [CrossRef] [Green Version]
- Xue, B.; Li, H.; Guo, M.; Wang, J.; Xu, Y.; Zou, X.; Deng, R.; Li, G.; Zhu, H. TRIM21 Promotes Innate Immune Response to RNA Viral Infection through Lys27-Linked Polyubiquitination of MAVS. J. Virol. 2018, 92, e00321-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watkinson, R.E.; McEwan, W.A.; James, L.C. Intracellular antibody immunity. J. Clin. Immunol. 2014, 34, S30–S34. [Google Scholar] [CrossRef] [PubMed]
- Hauler, F.; Mallery, D.L.; McEwan, W.A.; Bidgood, S.R.; James, L.C. AAA ATPase p97/VCP is essential for TRIM21-mediated virus neutralization. Proc. Natl. Acad. Sci. USA 2012, 109, 19733–19738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roux, K.J.; Kim, D.I.; Raida, M.; Burke, B. A promiscuous biotin ligase fusion protein identifies proximal and interacting proteins in mammalian cells. J. Cell Biol. 2012, 196, 801–810. [Google Scholar] [CrossRef] [Green Version]
- Su, T.; Chapin, S.J.; Bryant, D.M.; Shewan, A.M.; Young, K.; Mostov, K.E. Reduced immunoglobulin A transcytosis associated with immunoglobulin A nephropathy and nasopharyngeal carcinoma. J. Biol. Chem. 2011, 286, 44921–44925. [Google Scholar] [CrossRef] [Green Version]
- Oztan, A.; Rondanino, C.; Apodaca, G. Transcytosis of polymeric immunoglobulin a in polarized Madin-Darby canine kidney cells. Methods Mol. Biol. 2008, 440, 157–170. [Google Scholar] [CrossRef] [Green Version]
- Man, W.C.; Miyazaki, M.; Chu, K.; Ntambi, J.M. Membrane topology of mouse stearoyl-CoA desaturase 1. J. Biol. Chem. 2006, 281, 1251–1260. [Google Scholar] [CrossRef] [Green Version]
- Tang, B.L.; Tan, A.E.; Lim, L.K.; Lee, S.S.; Low, D.Y.; Hong, W. Syntaxin 12, a member of the syntaxin family localized to the endosome. J. Biol. Chem. 1998, 273, 6944–6950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hicke, L.; Dunn, R. Regulation of membrane protein transport by ubiquitin and ubiquitin-binding proteins. Annu. Rev. Cell Dev. Biol. 2003, 19, 141–172. [Google Scholar] [CrossRef]
- Piper, R.C.; Dikic, I.; Lukacs, G.L. Ubiquitin-dependent sorting in endocytosis. Cold Spring Harb. Perspect Biol. 2014, 6. [Google Scholar] [CrossRef]
- Benhra, N.; Vignaux, F.; Dussert, A.; Schweisguth, F.; Le Borgne, R. Neuralized promotes basal to apical transcytosis of delta in epithelial cells. Mol. Biol. Cell 2010, 21, 2078–2086. [Google Scholar] [CrossRef] [Green Version]
- Pan, J.A.; Sun, Y.; Jiang, Y.P.; Bott, A.J.; Jaber, N.; Dou, Z.; Yang, B.; Chen, J.S.; Catanzaro, J.M.; Du, C.; et al. TRIM21 Ubiquitylates SQSTM1/p62 and Suppresses Protein Sequestration to Regulate Redox Homeostasis. Mol. Cell 2016, 61, 720–733. [Google Scholar] [CrossRef] [Green Version]
- Wada, K.; Kamitani, T. Autoantigen Ro52 is an E3 ubiquitin ligase. Biochem. Biophys Res. Commun. 2006, 339, 415–421. [Google Scholar] [CrossRef]
- Macpherson, A.J.; McCoy, K.D.; Johansen, F.E.; Brandtzaeg, P. The immune geography of IgA induction and function. Mucosal. Immunol. 2008, 1, 11–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaetzel, C.S.; Robinson, J.K.; Chintalacharuvu, K.R.; Vaerman, J.P.; Lamm, M.E. The polymeric immunoglobulin receptor (secretory component) mediates transport of immune complexes across epithelial cells: A local defense function for IgA. Proc. Natl. Acad. Sci. USA 1991, 88, 8796–8800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mostov, K.; Su, T.; ter Beest, M. Polarized epithelial membrane traffic: Conservation and plasticity. Nat. Cell Biol. 2003, 5, 287–293. [Google Scholar] [CrossRef] [PubMed]
- Hansen, S.H.; Olsson, A.; Casanova, J.E. Wortmannin, an inhibitor of phosphoinositide 3-kinase, inhibits transcytosis in polarized epithelial cells. J. Biol. Chem. 1995, 270, 28425–28432. [Google Scholar] [CrossRef] [Green Version]
- Verges, M.; Luton, F.; Gruber, C.; Tiemann, F.; Reinders, L.G.; Huang, L.; Burlingame, A.L.; Haft, C.R.; Mostov, K.E. The mammalian retromer regulates transcytosis of the polymeric immunoglobulin receptor. Nat. Cell Biol. 2004, 6, 763–769. [Google Scholar] [CrossRef] [PubMed]
- Verges, M.; Sebastian, I.; Mostov, K.E. Phosphoinositide 3-kinase regulates the role of retromer in transcytosis of the polymeric immunoglobulin receptor. Exp. Cell Res. 2007, 313, 707–718. [Google Scholar] [CrossRef] [Green Version]
- Luton, F.; Hexham, M.J.; Zhang, M.; Mostov, K.E. Identification of a cytoplasmic signal for apical transcytosis. Traffic 2009, 10, 1128–1142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gallo, L.I.; Liao, Y.; Ruiz, W.G.; Clayton, D.R.; Li, M.; Liu, Y.J.; Jiang, Y.; Fukuda, M.; Apodaca, G.; Yin, X.M. TBC1D9B functions as a GTPase-activating protein for Rab11a in polarized MDCK cells. Mol. Biol. Cell 2014, 25, 3779–3797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ducharme, N.A.; Williams, J.A.; Oztan, A.; Apodaca, G.; Lapierre, L.A.; Goldenring, J.R. Rab11-FIP2 regulates differentiable steps in transcytosis. Am. J. Physiol. Cell Physiol. 2007, 293, 1059–1072. [Google Scholar] [CrossRef] [PubMed]
- Geuze, H.J.; Slot, J.W.; Strous, G.J.; Peppard, J.; von Figura, K.; Hasilik, A.; Schwartz, A.L. Intracellular receptor sorting during endocytosis: Comparative immunoelectron microscopy of multiple receptors in rat liver. Cell 1984, 37, 195–204. [Google Scholar] [CrossRef]
- Crifo, S.; Russo, M. IgA transport mechanism through the human nasal mucosa: An immunoenzymatic ultrastrutural study. Acta Otolaryngol. 1980, 89, 214–221. [Google Scholar] [CrossRef]
- In, J.G.; Tuma, P.L. MAL2 selectively regulates polymeric IgA receptor delivery from the Golgi to the plasma membrane in WIF-B cells. Traffic 2010, 11, 1056–1066. [Google Scholar] [CrossRef] [Green Version]
- Roohi, J.; Tegay, D.H.; Pomeroy, J.C.; Burkett, S.; Stone, G.; Stanyon, R.; Hatchwell, E. A de novo apparently balanced translocation [46,XY,t(2;9)(p13;p24)] interrupting RAB11FIP5 identifies a potential candidate gene for autism spectrum disorder. Am. J. Med. Genet. B Neuropsychiatr Genet. 2008, 147B, 411–417. [Google Scholar] [CrossRef] [Green Version]
- Matsunami, N.; Hensel, C.H.; Baird, L.; Stevens, J.; Otterud, B.; Leppert, T.; Varvil, T.; Hadley, D.; Glessner, J.T.; Pellegrino, R.; et al. Identification of rare DNA sequence variants in high-risk autism families and their prevalence in a large case/control population. Mol. Autism. 2014, 5, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, W.; Qin, G.; Shen, R. Rab11-FIP2 promotes the metastasis of gastric cancer cells. Int. J. Cancer 2016, 138, 1680–1688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, M.H.; Cho, K.H.; Jeong, K.J.; Park, Y.Y.; Kim, J.M.; Yu, S.L.; Park, C.G.; Mills, G.B.; Lee, H.Y. RCP induces Slug expression and cancer cell invasion by stabilizing beta1 integrin. Oncogene 2017, 36, 1102–1111. [Google Scholar] [CrossRef]
- Royo, M.; Gutierrez, Y.; Fernandez-Monreal, M.; Gutierrez-Eisman, S.; Jimenez, R.; Jurado, S.; Esteban, J.A. A retention-release mechanism based on RAB11FIP2 for AMPA receptor synaptic delivery during long-term potentiation. J. Cell Sci. 2019, 132. [Google Scholar] [CrossRef]
- Yoon, H.Y.; Kales, S.C.; Luo, R.; Lipkowitz, S.; Randazzo, P.A. ARAP1 association with CIN85 affects epidermal growth factor receptor endocytic trafficking. Biol. Cell 2011, 103, 171–184. [Google Scholar] [CrossRef]
- Yoon, H.Y.; Lee, J.S.; Randazzo, P.A. ARAP1 regulates endocytosis of EGFR. Traffic 2008, 9, 2236–2252. [Google Scholar] [CrossRef] [Green Version]
- Deng, L.; Wang, C.; Spencer, E.; Yang, L.; Braun, A.; You, J.; Slaughter, C.; Pickart, C.; Chen, Z.J. Activation of the IkappaB kinase complex by TRAF6 requires a dimeric ubiquitin-conjugating enzyme complex and a unique polyubiquitin chain. Cell 2000, 103, 351–361. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Deng, L.; Hong, M.; Akkaraju, G.R.; Inoue, J.; Chen, Z.J. TAK1 is a ubiquitin-dependent kinase of MKK and IKK. Nature 2001, 412, 346–351. [Google Scholar] [CrossRef]
- Sakai, R.; Fukuda, R.; Unida, S.; Aki, M.; Ono, Y.; Endo, A.; Kusumi, S.; Koga, D.; Fukushima, T.; Komada, M.; et al. The integral function of the endocytic recycling compartment is regulated by RFFL-mediated ubiquitylation of Rab11 effectors. J. Cell Sci. 2019, 132. [Google Scholar] [CrossRef] [Green Version]
- Bice, B.D.; Stephens, M.R.; Georges, S.J.; Venancio, A.R.; Bermant, P.C.; Warncke, A.V.; Affolter, K.E.; Hidalgo, J.R.; Angus-Hill, M.L. Environmental Enrichment Induces Pericyte and IgA-Dependent Wound Repair and Lifespan Extension in a Colon Tumor Model. Cell Rep. 2017, 19, 760–773. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Zhou, D.; Zhao, B.; Cao, Y.; Yu, J.; Yan, H.; Zhao, W.; Zhang, E.; Yang, J.; Zhong, M.; et al. Immunoglobulin A Targeting on the N-Terminal Moiety of Viral Phosphoprotein Prevents Measles Virus from Evading Interferon-beta Signaling. ACS Infect. Dis. 2020, 6, 844–856. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fan, X.; Zhou, D.; Zhao, B.; Sha, H.; Li, M.; Li, X.; Yang, J.; Yan, H. Rab11-FIP1 and Rab11-FIP5 Regulate pIgR/pIgA Transcytosis through TRIM21-Mediated Polyubiquitination. Int. J. Mol. Sci. 2021, 22, 10466. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms221910466
Fan X, Zhou D, Zhao B, Sha H, Li M, Li X, Yang J, Yan H. Rab11-FIP1 and Rab11-FIP5 Regulate pIgR/pIgA Transcytosis through TRIM21-Mediated Polyubiquitination. International Journal of Molecular Sciences. 2021; 22(19):10466. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms221910466
Chicago/Turabian StyleFan, Xuxu, Dihan Zhou, Bali Zhao, Huijun Sha, Mengxue Li, Xian Li, Jingyi Yang, and Huimin Yan. 2021. "Rab11-FIP1 and Rab11-FIP5 Regulate pIgR/pIgA Transcytosis through TRIM21-Mediated Polyubiquitination" International Journal of Molecular Sciences 22, no. 19: 10466. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms221910466