
Combination of PD98059 and TGF-β1 Efficiently Differentiates Human Urine-Derived Stem Cells into Smooth Muscle Cells


Abstract
:1. Introduction
2. Results
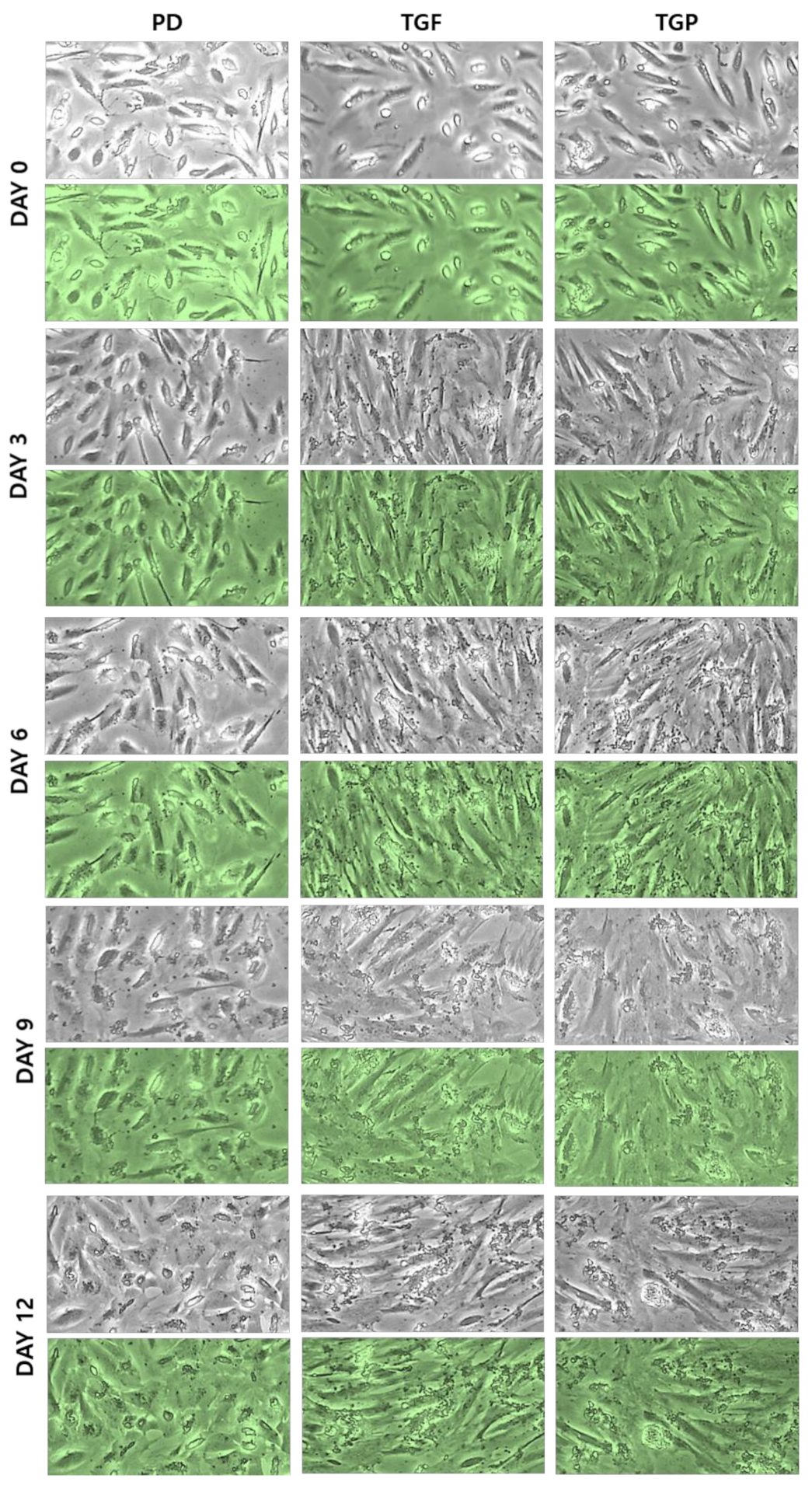
2.1. Morphological Changes of the Differentiated SMCs from hUDSCs
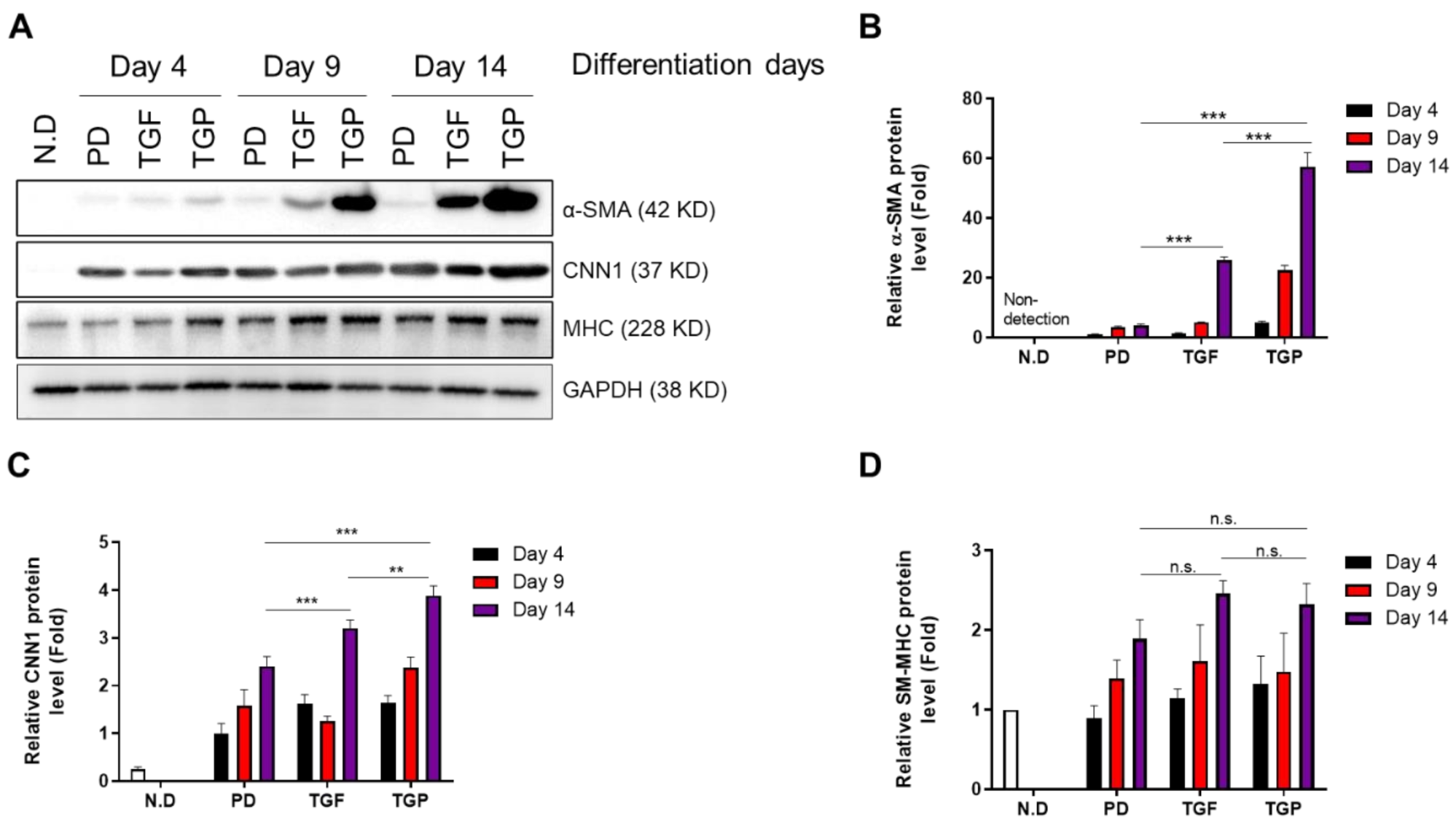
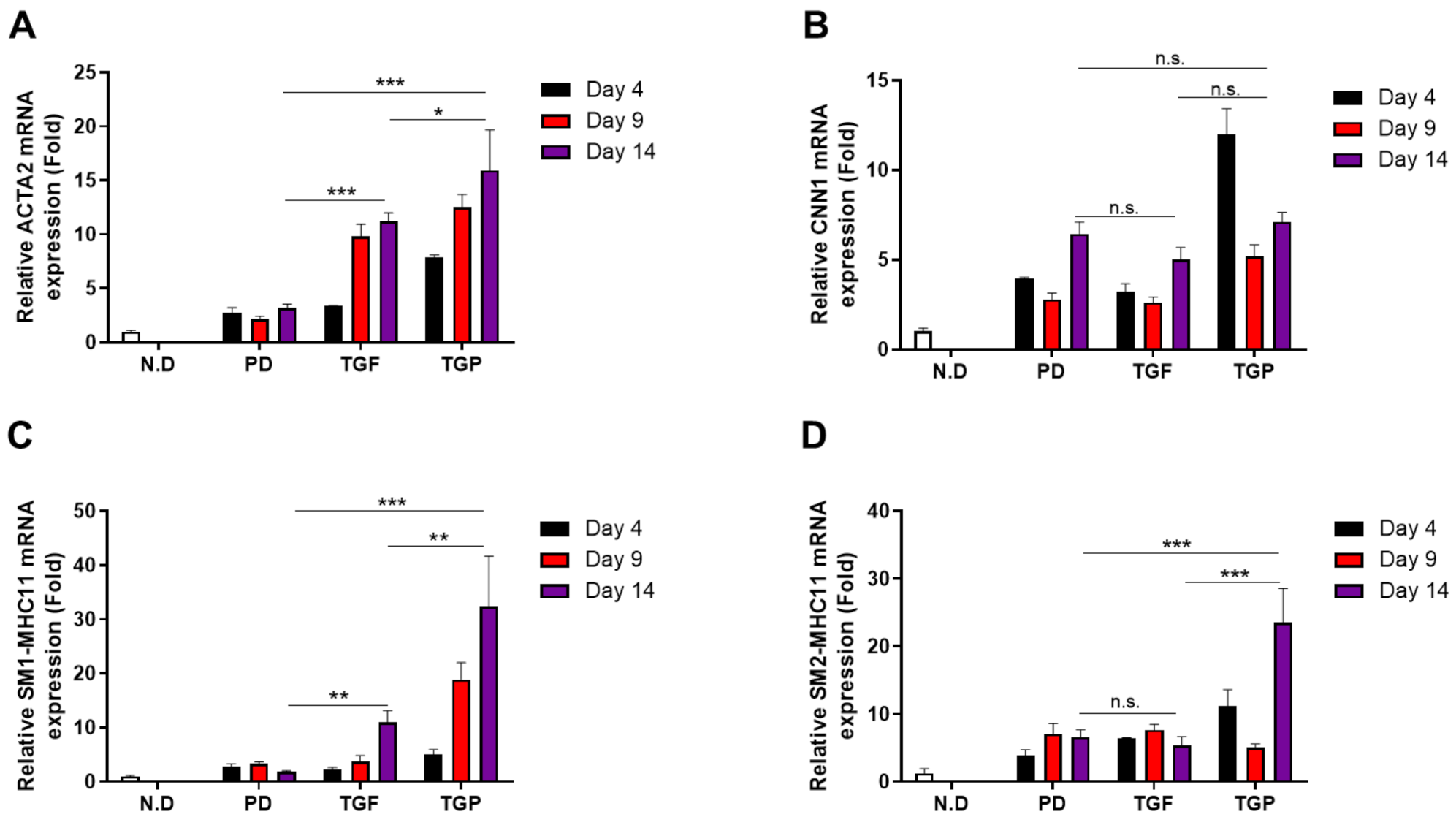
2.2. The Differentiated SMCs from hUDSCs Expressed SMC Marker Proteins, with the Highest Expression in Differentiated Cells with Combination of TGF-β1 and PD98059
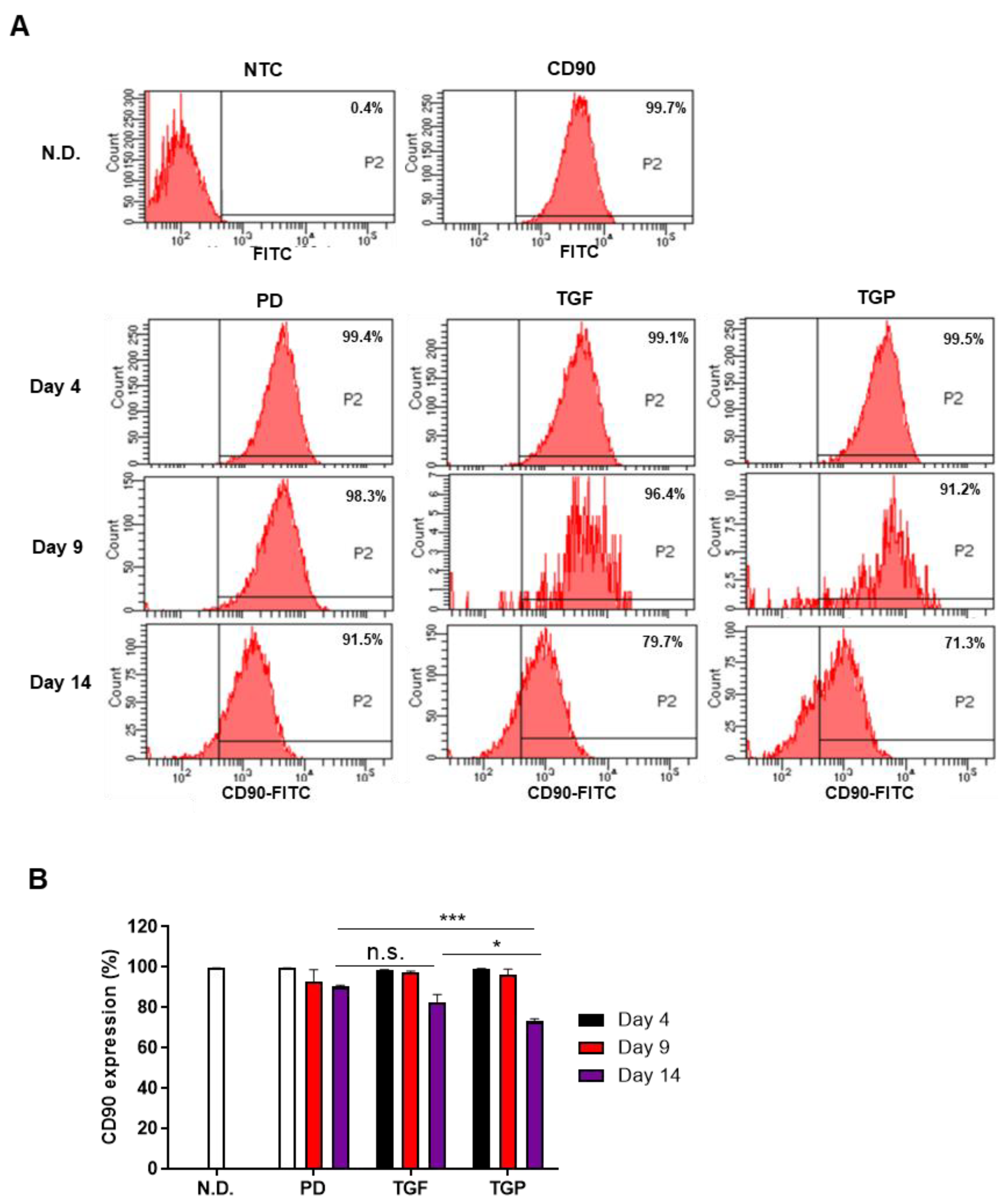
2.3. The Differentiated SMCs from hUDSCs Decreased the Expression of CD90, Mesenchymal Stem Cell Antigen, with the Highest Reduction in Differentiated Cells with a Combination of TGF-β1 and PD98059
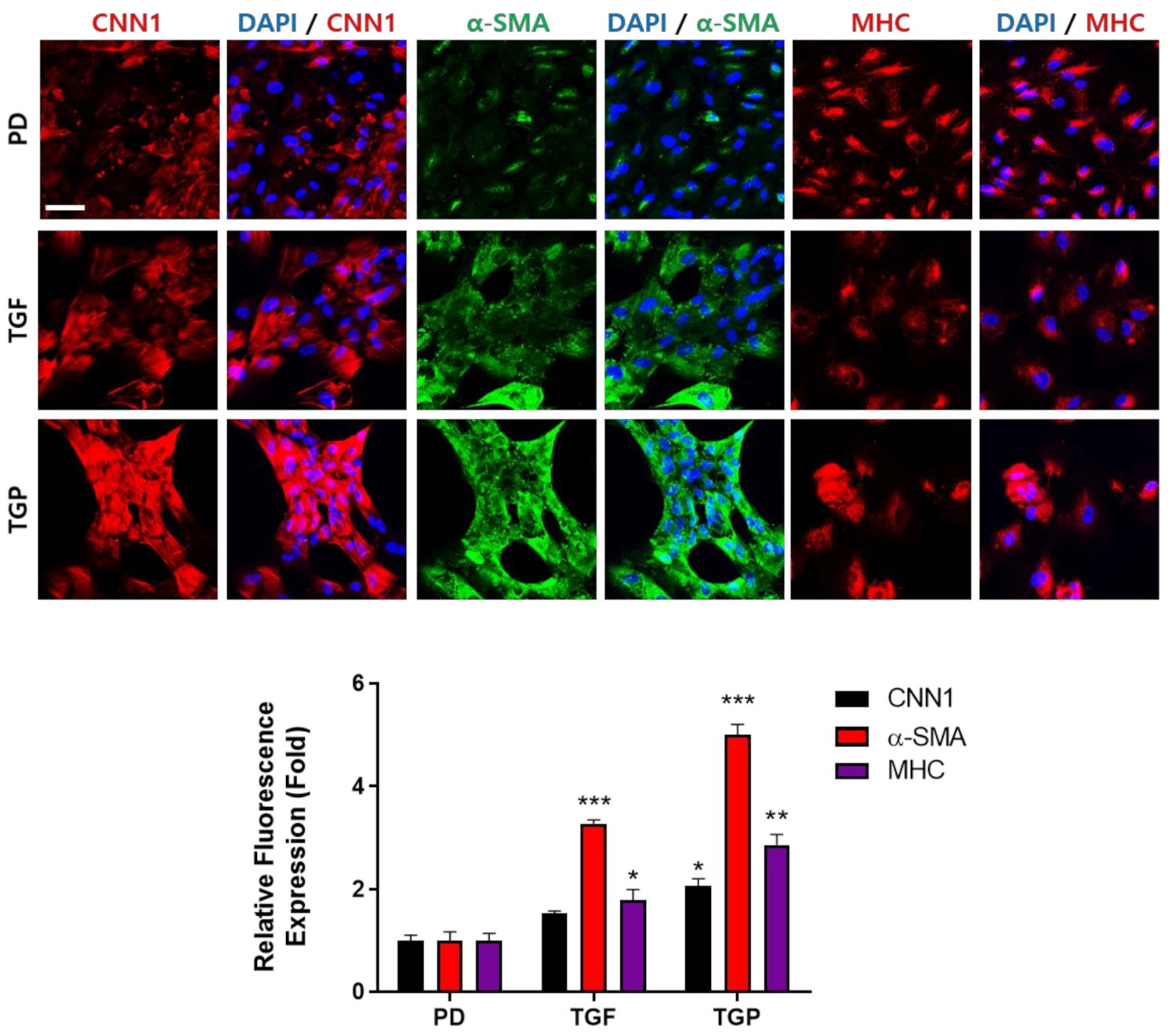
2.4. Immunocytochemical Analysis of SMC Markers in the Differentiated SMCs from hUDSCs
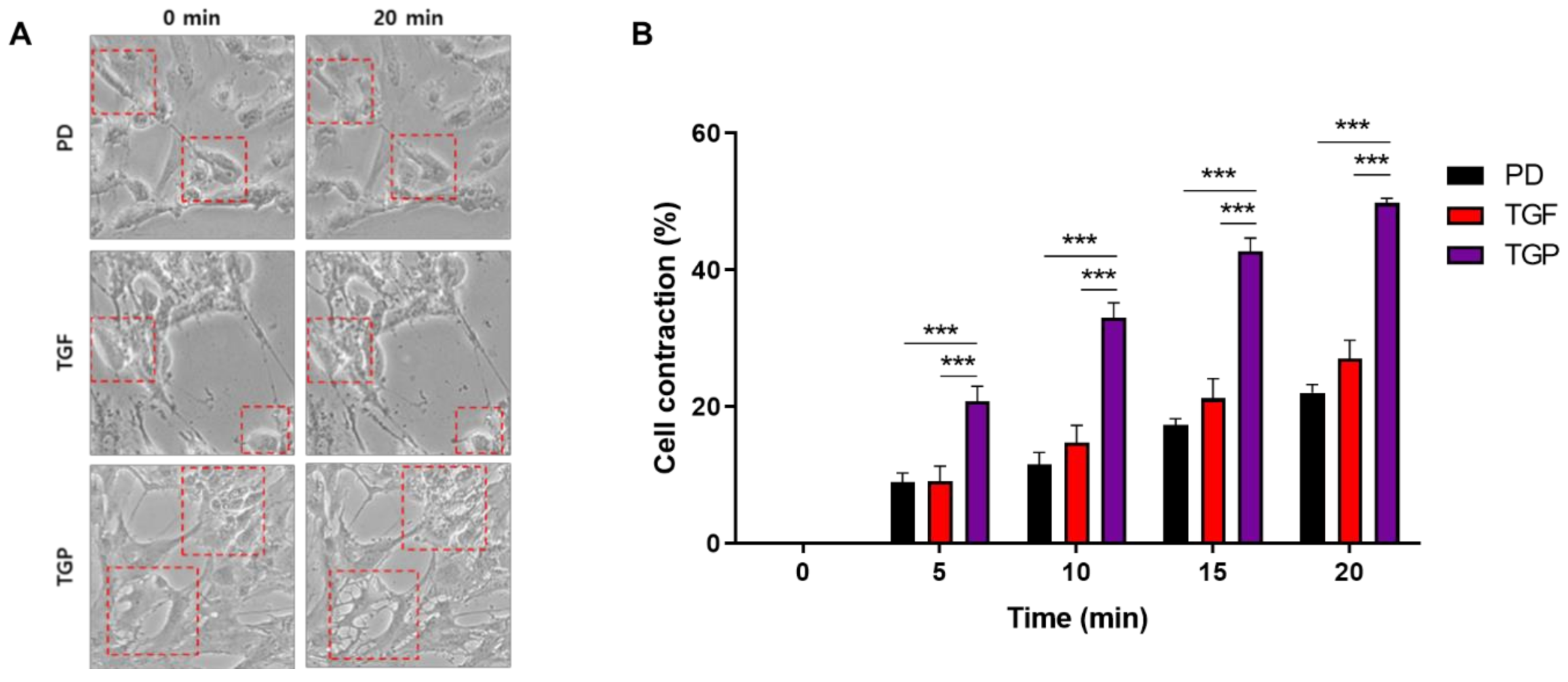
2.5. The Differentiated SMCs with a Combination of PD98059 and TGF-β1 Showed the Strongest Contracting Capacity
3. Discussion
4. Materials and Methods
4.1. Ethical Approval
4.2. Maintenance of hUDSCs
4.3. SMCs Differentiation
4.4. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
4.5. Flow Cytometry
4.6. Western Blotting
4.7. Characterization of SMCs by Immunocytochemistry
4.8. Contraction Test
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Charalambous, S.T.A. Impact of urinary incontinence on quality of life. Pelviperineology 2009, 28, 51–53. [Google Scholar]
- Luber, K.M. The definition, prevalence, and risk factors for stress urinary incontinence. Rev. Urol. 2004, 6 (Suppl. 3), S3–S9. [Google Scholar]
- Rovner, E.S.; Wein, A.J. Treatment options for stress urinary incontinence. Rev. Urol. 2004, 6 (Suppl. 3), S29–S47. [Google Scholar] [PubMed]
- Nilsson, C.G.; Kuuva, N. The tension-free vaginal tape procedure is successful in the majority of women with indications for surgical treatment of urinary stress incontinence. BJOG Int. J. Obstet. Gynaecol. 2001, 108, 414–419. [Google Scholar] [CrossRef] [PubMed]
- Wallner, C.; Dabhoiwala, N.F.; DeRuiter, M.C.; Lamers, W.H. The anatomical components of urinary continence. Eur. Urol. 2009, 55, 932–944. [Google Scholar] [CrossRef]
- Zhang, Y.; McNeill, E.; Tian, H.; Soker, S.; Andersson, K.E.; Yoo, J.J.; Atala, A. Urine derived cells are a potential source for urological tissue reconstruction. J. Urol. 2008, 180, 2226–2233. [Google Scholar] [CrossRef]
- Nickel, J.C. Lower urinary tract symptoms associated with prostatitis. Can. Urol. Assoc. J. 2012, 6, S133–S135. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.; Wei, G.; Li, P.; Zhou, X.; Zhang, Y. Urine-derived stem cells: A novel and versatile progenitor source for cell-based therapy and regenerative medicine. Genes Dis. 2014, 1, 8–17. [Google Scholar] [CrossRef] [Green Version]
- Krause, D.S.; Theise, N.D.; Collector, M.I.; Henegariu, O.; Hwang, S.; Gardner, R.; Neutzel, S.; Sharkis, S.J. Multi-organ, multi-lineage engraftment by a single bone marrow-derived stem cell. Cell 2001, 105, 369–377. [Google Scholar] [CrossRef] [Green Version]
- Covas, D.T.; Panepucci, R.A.; Fontes, A.M.; Silva, W.A., Jr.; Orellana, M.D.; Freitas, M.C.; Neder, L.; Santos, A.R.; Peres, L.C.; Jamur, M.C.; et al. Multipotent mesenchymal stromal cells obtained from diverse human tissues share functional properties and gene-expression profile with CD146+ perivascular cells and fibroblasts. Exp. Hematol. 2008, 36, 642–654. [Google Scholar] [CrossRef]
- Karaoz, E.; Okcu, A.; Unal, Z.S.; Subasi, C.; Saglam, O.; Duruksu, G. Adipose tissue-derived mesenchymal stromal cells efficiently differentiate into insulin-producing cells in pancreatic islet microenvironment both in vitro and in vivo. Cytotherapy 2013, 15, 557–570. [Google Scholar] [CrossRef]
- Zuk, P.A.; Zhu, M.; Ashjian, P.; De Ugarte, D.A.; Huang, J.I.; Mizuno, H.; Alfonso, Z.C.; Fraser, J.K.; Benhaim, P.; Hedrick, M.H. Human adipose tissue is a source of multipotent stem cells. Mol. Biol. Cell 2002, 13, 4279–4295. [Google Scholar] [CrossRef]
- Bharadwaj, S.; Liu, G.; Shi, Y.; Wu, R.; Yang, B.; He, T.; Fan, Y.; Lu, X.; Zhou, X.; Liu, H.; et al. Multipotential differentiation of human urine-derived stem cells: Potential for therapeutic applications in urology. Stem Cells 2013, 31, 1840–1856. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Li, L.; Xing, F.; Peng, J.; Peng, K.; Wang, Y.; Xiang, Z. Human Urine-Derived Stem Cells: Potential for Cell-Based Therapy of Cartilage Defects. Stem Cells Int 2018, 2018, 4686259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, D.; Cheng, F.; Pan, S.; Liu, Z. Stem cells: A potential treatment option for kidney diseases. Stem Cell Res. Ther. 2020, 11, 249. [Google Scholar] [CrossRef] [PubMed]
- Qin, D.; Long, T.; Deng, J.; Zhang, Y. Urine-derived stem cells for potential use in bladder repair. Stem Cell Res. Ther. 2014, 5, 69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ikedo, H.; Tamaki, K.; Ueda, S.; Kato, S.; Fujii, M.; Ten Dijke, P.; Okuda, S. Smad protein and TGF-beta signaling in vascular smooth muscle cells. Int. J. Mol. Med. 2003, 11, 645–650. [Google Scholar]
- Yang, X.; Long, L.; Southwood, M.; Rudarakanchana, N.; Upton, P.D.; Jeffery, T.K.; Atkinson, C.; Chen, H.; Trembath, R.C.; Morrell, N.W. Dysfunctional Smad signaling contributes to abnormal smooth muscle cell proliferation in familial pulmonary arterial hypertension. Circ. Res. 2005, 96, 1053–1063. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, X.; Chen, S.-Y. Transforming growth factor-β and smooth muscle differentiation. World J. Biol. Chem. 2012, 3, 41–52. [Google Scholar] [CrossRef]
- Owens, G.K.; Wise, G. Regulation of differentiation/maturation in vascular smooth muscle cells by hormones and growth factors. Agents Actions Suppl. 1997, 48, 3–24. [Google Scholar]
- Chi, J.-T.; Rodriguez, E.H.; Wang, Z.; Nuyten, D.S.A.; Mukherjee, S.; van de Rijn, M.; van de Vijver, M.J.; Hastie, T.; Brown, P.O. Gene expression programs of human smooth muscle cells: Tissue-specific differentiation and prognostic significance in breast cancers. PLoS Genet. 2007, 3, 1770–1784. [Google Scholar] [CrossRef]
- Tamama, K.; Sen, C.K.; Wells, A. Differentiation of bone marrow mesenchymal stem cells into the smooth muscle lineage by blocking ERK/MAPK signaling pathway. Stem Cells Dev. 2008, 17, 897–908. [Google Scholar] [CrossRef]
- Narita, Y.; Yamawaki, A.; Kagami, H.; Ueda, M.; Ueda, Y. Effects of transforming growth factor-beta 1 and ascorbic acid on differentiation of human bone-marrow-derived mesenchymal stem cells into smooth muscle cell lineage. Cell Tissue Res. 2008, 333, 449–459. [Google Scholar] [CrossRef]
- Park, J.S.; Chu, J.S.; Tsou, A.D.; Diop, R.; Tang, Z.; Wang, A.; Li, S. The effect of matrix stiffness on the differentiation of mesenchymal stem cells in response to TGF-β. Biomaterials 2011, 32, 3921–3930. [Google Scholar] [CrossRef] [Green Version]
- Biswas, B.; Bhattacharyya, A.; Dasgupta, A.; Karmakar, A.; Mallick, N.; Sembiah, S. Urinary Incontinence, Its Risk Factors, and Quality of Life: A Study among Women Aged 50 Years and above in a Rural Health Facility of West Bengal. J. Midlife Health 2017, 8, 130–136. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.H.; Salcedo, L.B.; Damaser, M.S. Quantification of neurological and other contributors to continence in female rats. Brain Res. 2011, 1382, 198–205. [Google Scholar] [CrossRef] [Green Version]
- Lightner, D.J.; Knoedler, J.J.; Linder, B.J. Periurethral Bulking Agent Injection in the Treatment of Female Stress Urinary Incontinence. In Complications of Female Incontinence and Pelvic Reconstructive Surgery; Goldman, H.B., Ed.; Springer International Publishing: Cham, Switzerland, 2017; pp. 297–305. [Google Scholar] [CrossRef]
- Ni, J.; Li, H.; Zhou, Y.; Gu, B.; Xu, Y.; Fu, Q.; Peng, X.; Cao, N.; Fu, Q.; Jin, M.; et al. Therapeutic Potential of Human Adipose-Derived Stem Cell Exosomes in Stress Urinary Incontinence—An in Vitro and in Vivo Study. Cell. Physiol. Biochem. 2018, 48, 1710–1722. [Google Scholar] [CrossRef]
- Zambon, J.P.; Williams, K.J. Applicability of regenerative medicine and tissue engineering for the treatment of stress urinary incontinence in female patients. Neurourol. Urodyn. 2019, 38 (Suppl. 4), S76–S83. [Google Scholar] [CrossRef] [PubMed]
- Peyromaure, M.; Sebe, P.; Praud, C.; DeRocle, G.; Potin, N.; Pinset, C.; Sebille, A. Fate of implanted syngenic muscle precursor cells in striated urethral sphincter of female rats: Perspectives for treatment of urinary incontinence. Urology 2004, 64, 1037–1041. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Wen, Y.; Li, Y.H.; Wei, Y.; Green, M.; Wani, P.; Zhang, P.; Pera, R.R.; Chen, B. Smooth Muscle Precursor Cells Derived from Human Pluripotent Stem Cells for Treatment of Stress Urinary Incontinence. Stem Cells Dev. 2016, 25, 453–461. [Google Scholar] [CrossRef] [Green Version]
- Abbas, T.O.; Ali, T.A.; Uddin, S. Urine as a Main Effector in Urological Tissue Engineering—A Double-Edged Sword. Cells 2020, 9, 538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bharadwaj, S.; Liu, G.; Shi, Y.; Markert, C.; Andersson, K.-E.; Atala, A.; Zhang, Y. Characterization of Urine-Derived Stem Cells Obtained from Upper Urinary Tract for Use in Cell-Based Urological Tissue Engineering. Tissue Eng. Part A 2011, 17, 2123–2132. [Google Scholar] [CrossRef] [PubMed]
- Sinha, S.; Hoofnagle, M.H.; Kingston, P.A.; McCanna, M.E.; Owens, G.K. Transforming growth factor-β1 signaling contributes to development of smooth muscle cells from embryonic stem cells. Am. J. Physiol.-Cell Physiol. 2004, 287, C1560–C1568. [Google Scholar] [CrossRef]
- Owens, G.K.; Kumar, M.S.; Wamhoff, B.R. Molecular Regulation of Vascular Smooth Muscle Cell Differentiation in Development and Disease. Physiol. Rev. 2004, 84, 767–801. [Google Scholar] [CrossRef]
- Desmoulière, A.; Geinoz, A.; Gabbiani, F.; Gabbiani, G. Transforming growth factor-beta 1 induces alpha-smooth muscle actin expression in granulation tissue myofibroblasts and in quiescent and growing cultured fibroblasts. J. Cell Biol. 1993, 122, 103–111. [Google Scholar] [CrossRef] [Green Version]
- Woodman, L.; Siddiqui, S.; Cruse, G.; Sutcliffe, A.; Saunders, R.; Kaur, D.; Bradding, P.; Brightling, C. Mast Cells Promote Airway Smooth Muscle Cell Differentiation via Autocrine Up-Regulation of TGF-β1. J. Immunol. 2008, 181, 5001–5007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sibov, T.T.; Severino, P.; Marti, L.C.; Pavon, L.F.; Oliveira, D.M.; Tobo, P.R.; Campos, A.H.; Paes, A.T.; Amaro, E., Jr.; Gamarra, L.F.; et al. Mesenchymal stem cells from umbilical cord blood: Parameters for isolation, characterization and adipogenic differentiation. Cytotechnology 2012, 64, 511–521. [Google Scholar] [CrossRef] [Green Version]
- Moraes, D.A.; Sibov, T.T.; Pavon, L.F.; Alvim, P.Q.; Bonadio, R.S.; Da Silva, J.R.; Pic-Taylor, A.; Toledo, O.A.; Marti, L.C.; Azevedo, R.B.; et al. A reduction in CD90 (THY-1) expression results in increased differentiation of mesenchymal stromal cells. Stem Cell Res. Ther. 2016, 7, 97. [Google Scholar] [CrossRef] [Green Version]
- Long, X.; Bell, R.D.; Gerthoffer, W.T.; Zlokovic, B.V.; Miano, J.M. Myocardin is sufficient for a smooth muscle-like contractile phenotype. Arterioscler. Thromb. Vasc. Biol. 2008, 28, 1505–1510. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.Z.; Olson, E.N. Control of smooth muscle development by the myocardin family of transcriptional coactivators. Curr. Opin. Genet. Dev. 2004, 14, 558–566. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.-P.; Wang, Z.; Yanagisawa, H.; Olson, E.N. Phenotypic Modulation of Smooth Muscle Cells through Interaction of Foxo4 and Myocardin. Dev. Cell 2005, 9, 261–270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Veber, M.; Dolivo, D.; Rolle, M.; Dominko, T. Pro-myogenic and low-oxygen culture increases expression of contractile smooth muscle markers in human fibroblasts. J. Tissue Eng. Regen. Med. 2018, 12, 572–582. [Google Scholar] [CrossRef] [PubMed]
- Hwang, Y.; Cha, S.-H.; Hong, Y.; Jung, A.R.; Jun, H.-S. Direct differentiation of insulin-producing cells from human urine-derived stem cells. Int. J. Med. Sci. 2019, 16, 1668–1676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Qiu, P.; Jiao, J.; Hirai, H.; Xiong, W.; Zhang, J.; Zhu, T.; Ma, P.X.; Chen, Y.E.; Yang, B. Yes-Associated Protein Inhibits Transcription of Myocardin and Attenuates Differentiation of Vascular Smooth Muscle Cell from Cardiovascular Progenitor Cell Lineage. Stem Cells 2017, 35, 351–361. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Accession Number | Primer Name | Primer Sequence (5′-3′) | Aneling Temperature (°C) | Amplification Size (bp) |
|---|---|---|---|---|---|
| Actin alpha 2, smooth muscle (ACTA2) | NM_001613.4 | ACTA2-F ACTA2-R | ACTGCCTTGGTGTGTGACAA TCAAACCCCAATCCACAGAG | 55 | 120 |
| Calponin 1 (CNN1), | NM_001299.6 | CNN1-F CNN1-R | AGGTTAAGAACAAGCTGGCCC GAGGCCGTCCATGAAGTTGT | 55 | 113 |
| Myosin heavy chain 11 isoform SM1 (MYH11 SM1) | NM_002474.2 | MYH11-SM1-F MYH11-SM1-R | CAAGAGCAAGCTCAGGCGA CCTCCTCAGAACCATCTGCATT | 55 | 98 |
| Myosin heavy chain 11 isoform SM2 (MYH11 SM2) | NM_022844.2 | MYH11-SM2-F MYH11-SM2-R | GATGCACCAGGCGAGGAAA TGAAGTCTGCGTCTCGAGTG | 55 | 120 |
| Cyclophilin B | NM_000942.5 | Cyclo-F Cyclo-R | TGCCATCGCCAAGGAGTAG TGCACAGACGGTCACTCAAA | 55 | 57 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hwang, Y.; Cha, S.-H.; Kim, D.; Jun, H.-S. Combination of PD98059 and TGF-β1 Efficiently Differentiates Human Urine-Derived Stem Cells into Smooth Muscle Cells. Int. J. Mol. Sci. 2021, 22, 10532. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms221910532
Hwang Y, Cha S-H, Kim D, Jun H-S. Combination of PD98059 and TGF-β1 Efficiently Differentiates Human Urine-Derived Stem Cells into Smooth Muscle Cells. International Journal of Molecular Sciences. 2021; 22(19):10532. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms221910532
Chicago/Turabian StyleHwang, Yongha, Seon-Heui Cha, Donghee Kim, and Hee-Sook Jun. 2021. "Combination of PD98059 and TGF-β1 Efficiently Differentiates Human Urine-Derived Stem Cells into Smooth Muscle Cells" International Journal of Molecular Sciences 22, no. 19: 10532. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms221910532