
Dependence of Graphene Oxide (GO) Toxicity on Oxidation Level, Elemental Composition, and Size

 , , and
, , and
Abstract
:1. Introduction
2. Results
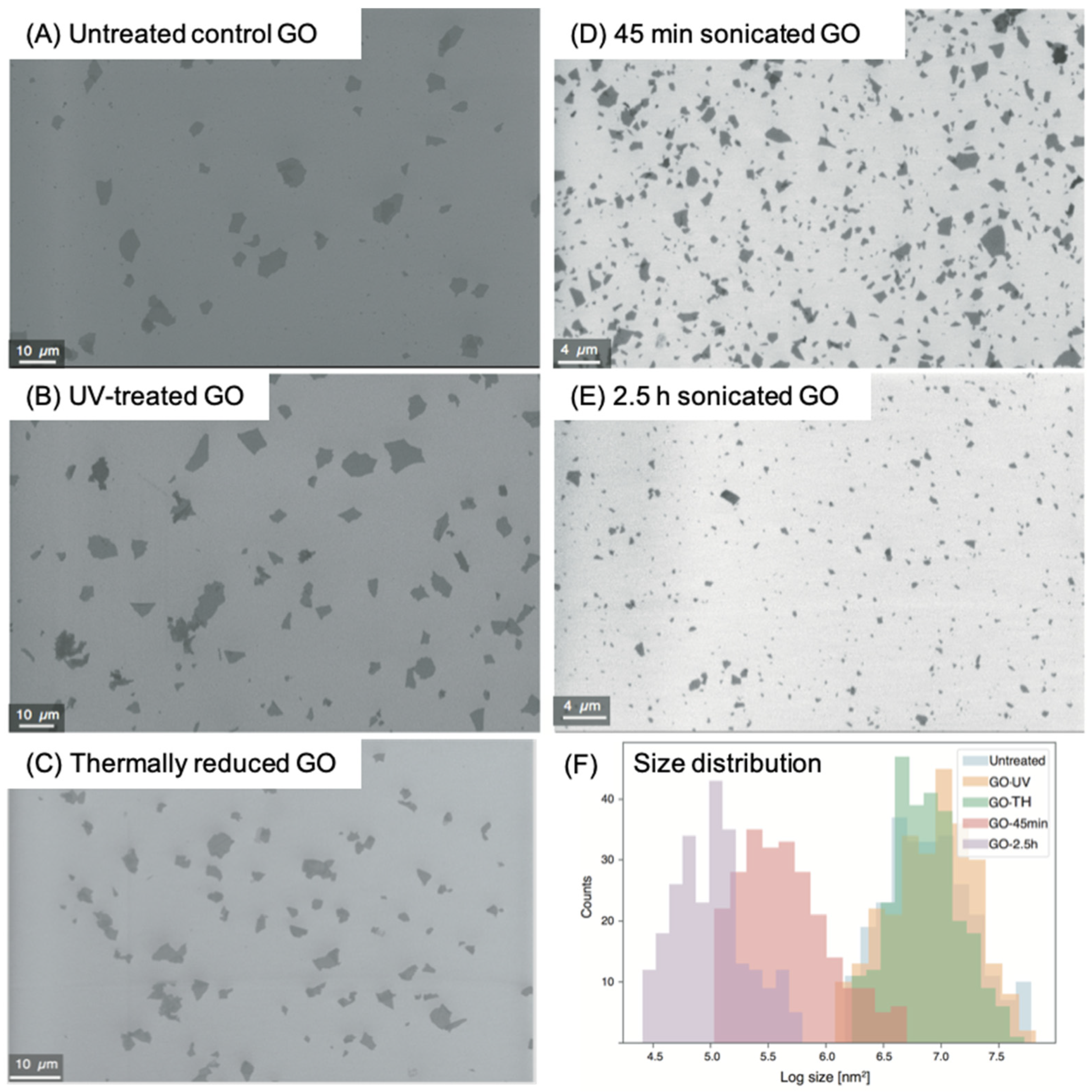
2.1. Treatment and Characterization of Graphene Oxides
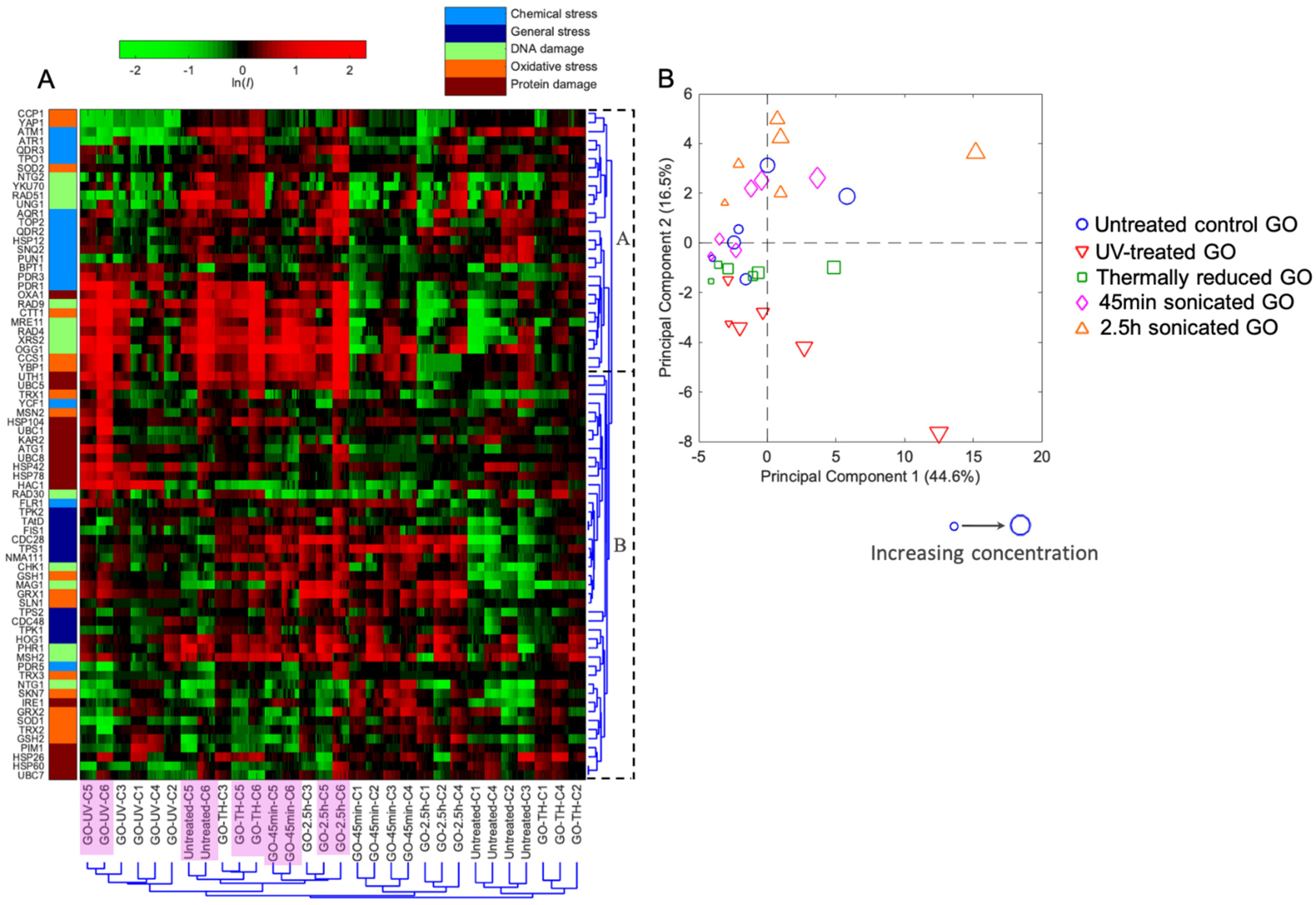
2.2. Distinctive Toxicity Profiles among Five GOs with Varying Physiochemical Properties Revealed by the Quantitative Toxicogenomic Analysis
2.3. Insights into Toxicity Mechanisms Revealed by Comparison among Various GOs
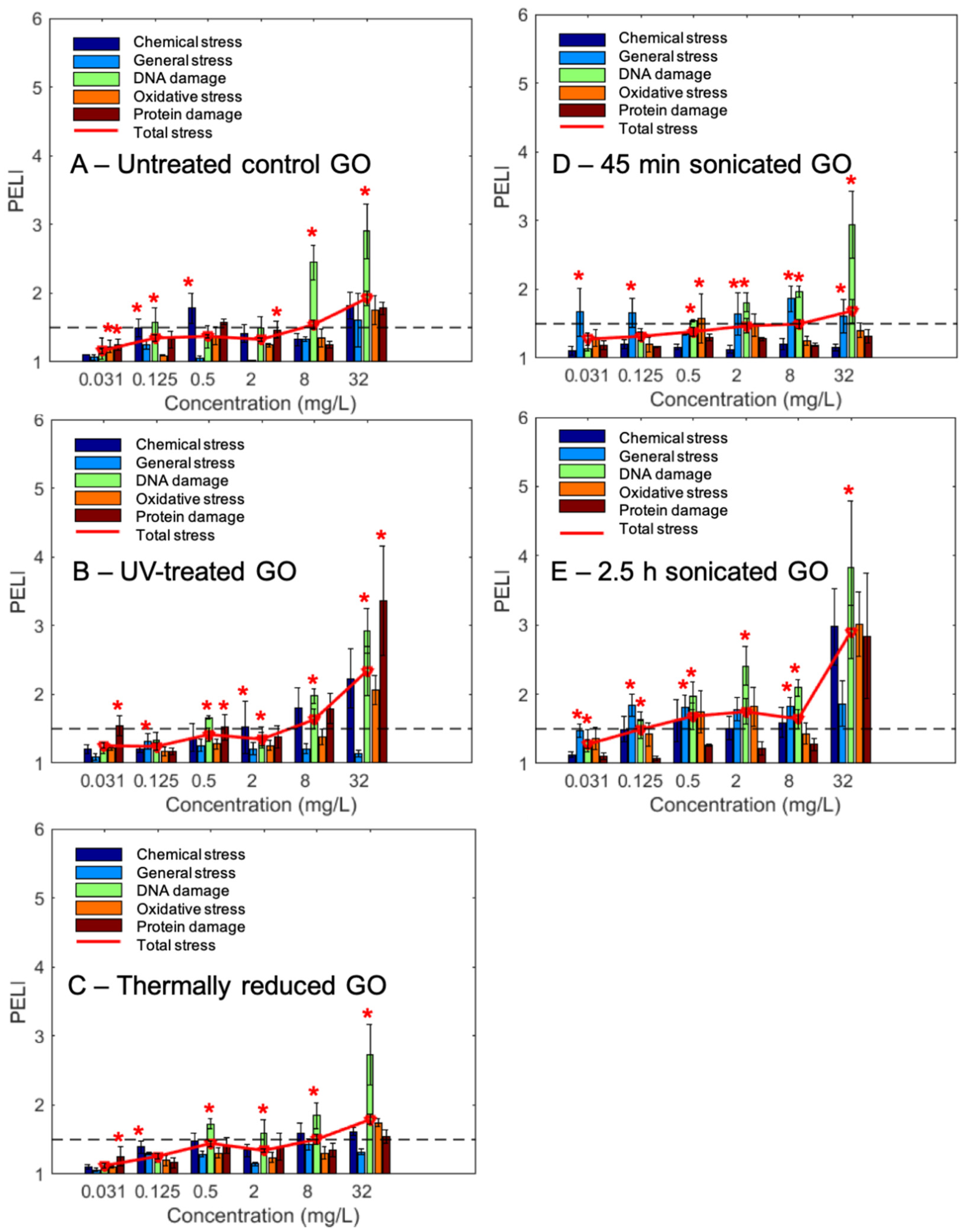
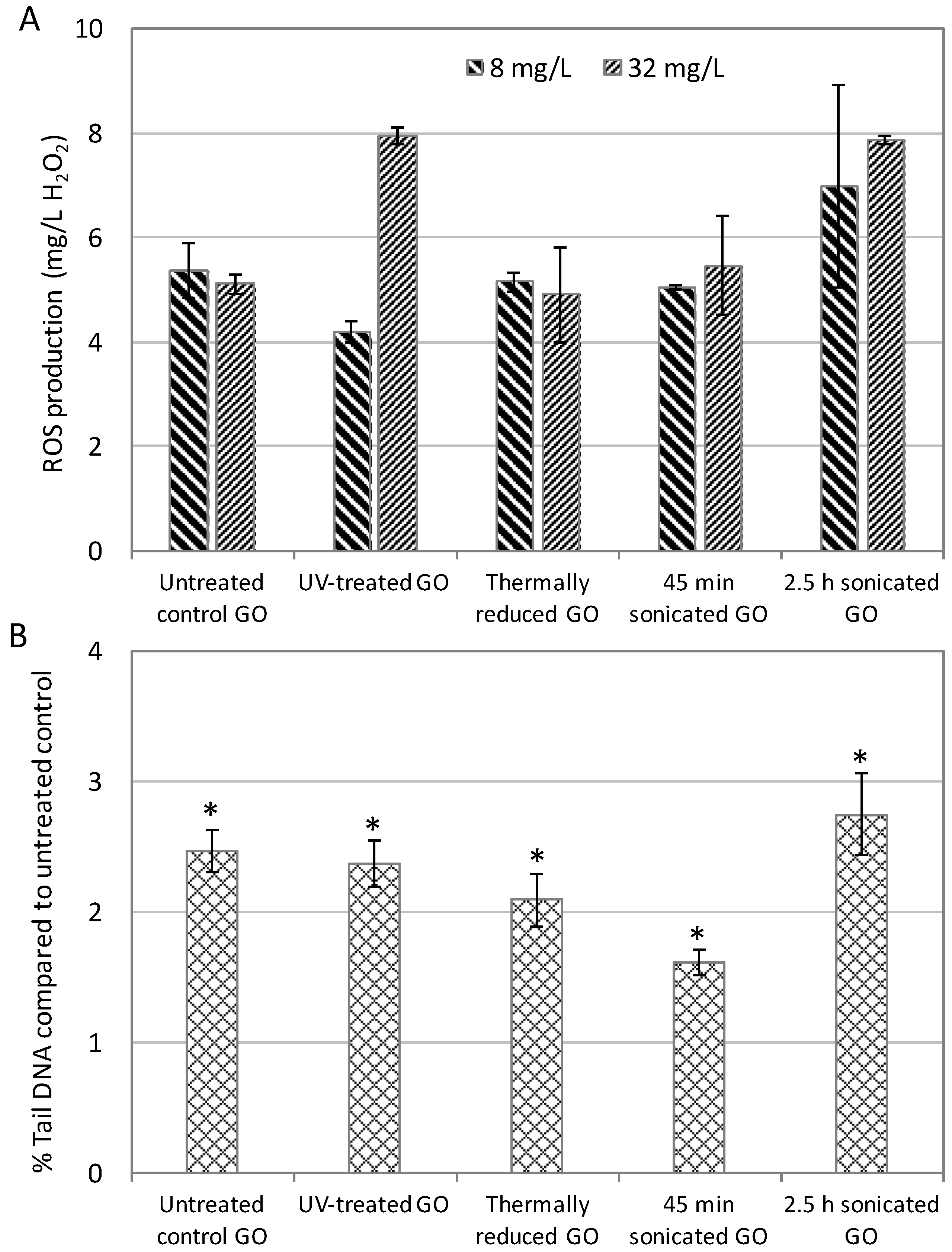
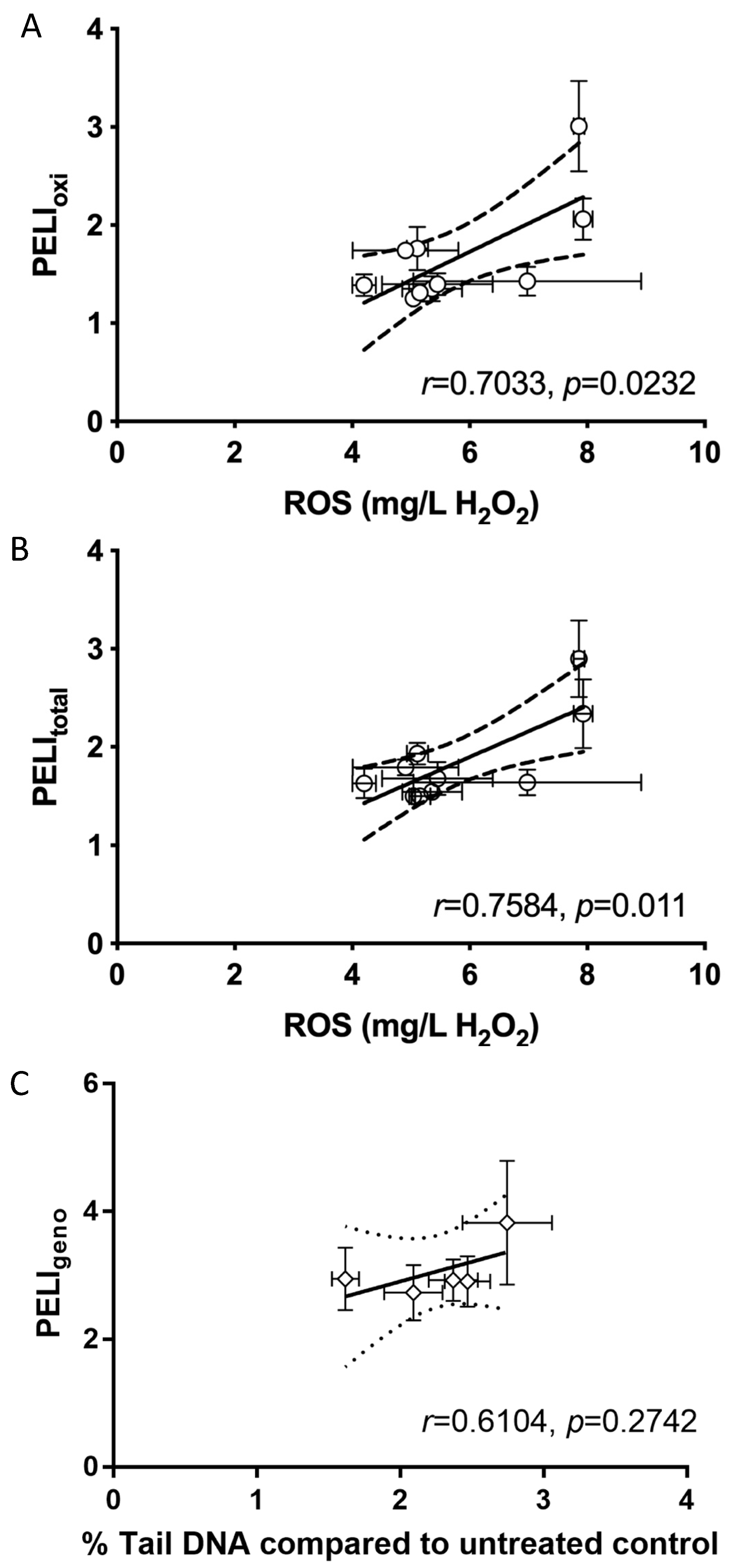
2.4. Molecular Toxicity Endpoints Correlated with Conventional Phenotypic Toxicity Endpoints of GOs
3. Discussion
3.1. Toxicity Mechanisms of the Untreated GO
3.2. Effect of UV Treatment and Thermal Reduction on Toxicity of GOs
3.3. Impact of Size on GO Toxicity
4. Materials and Methods
4.1. Nanomaterials Information and Preparation
4.2. GO Characterization
4.3. Toxicogenomic Assay and Quantitative Molecular Endpoint Derivation
4.4. Intracellular ROS Production Measurement
4.5. DNA Damage Alkaline Comet Assay in Human A549 Cells
4.6. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Kemp, K.C.; Seema, H.; Saleh, M.; Le, N.H.; Mahesh, K.; Chandra, V.; Kim, K.S. Environmental applications using graphene composites: Water remediation and gas adsorption. Nanoscale 2013, 5, 3149–3171. [Google Scholar] [CrossRef] [Green Version]
- Goenka, S.; Sant, V.; Sant, S. Graphene-based nanomaterials for drug delivery and tissue engineering. J. Control. Release 2014, 173, 75–88. [Google Scholar] [CrossRef]
- Akhavan, O.; Ghaderi, E. Toxicity of Graphene and Graphene Oxide Nanowalls Against Bacteria. ACS Nano 2010, 4, 5731–5736. [Google Scholar] [CrossRef]
- Park, S.; Ruoff, R.S. Chemical methods for the production of graphenes. Nat. Nanotechnol. 2009, 4, 217–224. [Google Scholar] [CrossRef]
- Nogueira, P.F.M.; Nakabayashi, D.; Zucolotto, V. The effects of graphene oxide on green algae Raphidocelis subcapitata. Aquat. Toxicol. 2015, 166, 29–35. [Google Scholar] [CrossRef]
- Wang, L.; Lee, K.; Sun, Y.Y.; Lucking, M.; Chen, Z.F.; Zhao, J.J.; Zhang, S.B.B. Graphene Oxide as an Ideal Substrate for Hydrogen Storage. ACS Nano 2009, 3, 2995–3000. [Google Scholar] [CrossRef] [PubMed]
- Scheuermann, G.M.; Rumi, L.; Steurer, P.; Bannwarth, W.; Mulhaupt, R. Palladium Nanoparticles on Graphite Oxide and Its Functionalized Graphene Derivatives as Highly Active Catalysts for the Suzuki-Miyaura Coupling Reaction. J. Am. Chem. Soc. 2009, 131, 8262–8270. [Google Scholar] [CrossRef] [PubMed]
- Eda, G.; Fanchini, G.; Chhowalla, M. Large-area ultrathin films of reduced graphene oxide as a transparent and flexible electronic material. Nat. Nanotechnol. 2008, 3, 270–274. [Google Scholar] [CrossRef] [PubMed]
- Perreault, F.; Tousley, M.E.; Elimelech, M. Thin-Film Composite Polyamide Membranes Functionalized with Biocidal Graphene Oxide Nanosheets. Environ. Sci. Technol. Lett. 2014, 1, 71–76. [Google Scholar] [CrossRef]
- Hu, W.B.; Peng, C.; Luo, W.J.; Lv, M.; Li, X.M.; Li, D.; Huang, Q.; Fan, C.H. Graphene-Based Antibacterial Paper. ACS Nano 2010, 4, 4317–4323. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.H.; Yang, H.H.; Zhu, C.L.; Chen, X.; Chen, G.N. A Graphene Platform for Sensing Biomolecules. Angew. Chem. Int. Ed. 2009, 48, 4785–4787. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.B.; Zhang, Q.; Chu, X.; Chen, T.T.; Ge, J.; Yu, R.Q. Graphene Oxide-Peptide Conjugate as an Intracellular Protease Sensor for Caspase-3 Activation Imaging in Live Cells. Angew. Chem. Int. Ed. 2011, 50, 7065–7069. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Robinson, J.T.; Sun, X.M.; Dai, H.J. PEGylated nanographene oxide for delivery of water-insoluble cancer drugs. J. Am. Chem. Soc. 2008, 130, 10876–10877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, C.H.; Zhu, C.L.; Li, J.; Liu, J.J.; Chen, X.; Yang, H.H. Using graphene to protect DNA from cleavage during cellular delivery. Chem. Commun. 2010, 46, 3116–3118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, X.M.; Liu, Z.; Welsher, K.; Robinson, J.T.; Goodwin, A.; Zaric, S.; Dai, H.J. Nano-Graphene Oxide for Cellular Imaging and Drug Delivery. Nano Res. 2008, 1, 203–212. [Google Scholar] [CrossRef] [Green Version]
- Chowdhury, I.; Duch, M.C.; Mansukhani, N.D.; Hersam, M.C.; Bouchard, D. Colloidal Properties and Stability of Graphene Oxide Nanomaterials in the Aquatic Environment. Environ. Sci. Technol. 2013, 47, 6288–6296. [Google Scholar] [CrossRef]
- Handy, R.D.; van den Brink, N.; Chappell, M.; Muhling, M.; Behra, R.; Dusinska, M.; Simpson, P.; Ahtiainen, J.; Jha, A.N.; Seiter, J.; et al. Practical considerations for conducting ecotoxicity test methods with manufactured nanomaterials: What have we learnt so far? Ecotoxicology 2012, 21, 933–972. [Google Scholar] [CrossRef] [Green Version]
- Sanchez, V.C.; Jachak, A.; Hurt, R.H.; Kane, A.B. Biological Interactions of Graphene-Family Nanomaterials: An Interdisciplinary Review. Chem. Res. Toxicol. 2012, 25, 15–34. [Google Scholar] [CrossRef] [Green Version]
- Fisher, C.; Rider, A.E.; Han, Z.J.; Kumar, S.; Levchenko, I.; Ostrikov, K. Applications and Nanotoxicity of Carbon Nanotubes and Graphene in Biomedicine. J. Nanomater. 2012, 14, 122. [Google Scholar] [CrossRef] [Green Version]
- Nel, A.; Xia, T.; Madler, L.; Li, N. Toxic potential of materials at the nanolevel. Science 2006, 311, 622–627. [Google Scholar] [CrossRef] [Green Version]
- Thurnherr, T.; Brandenberger, C.; Fischer, K.; Diener, L.; Manser, P.; Maeder-Althaus, X.; Kaiser, J.P.; Krug, H.F.; Rothen-Rutishauser, B.; Wick, P. A comparison of acute and long-term effects of industrial multiwalled carbon nanotubes on human lung and immune cells in vitro. Toxicol. Lett. 2011, 200, 176–186. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.J.; Yin, J.F.; Liang, Y.; Yuan, S.P.; Wang, F.B.; Song, M.Y.; Wang, H.L. Oxidative stress and immunotoxicity induced by graphene oxide in zebrafish. Aquat. Toxicol. 2016, 174, 54–60. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.L.; Yang, S.T.; Liu, J.H.; Dong, E.; Wang, Y.W.; Cao, A.N.; Liu, Y.F.; Wang, H.F. In vitro toxicity evaluation of graphene oxide on A549 cells. Toxicol. Lett. 2011, 200, 201–210. [Google Scholar] [CrossRef] [PubMed]
- Long, Z.F.; Ji, J.; Yang, K.; Lin, D.H.; Wu, F.C. Systematic and Quantitative Investigation of the Mechanism of Carbon Nanotubes’ Toxicity toward Algae. Environ. Sci. Technol. 2012, 46, 8458–8466. [Google Scholar] [CrossRef]
- Hondroulis, E.; Zhang, Z.Q.; Chen, C.Y.; Li, C.Z. Impedance Based Nanotoxicity Assessment of Graphene Nanomaterials at the Cellular and Tissue Level. Anal. Lett. 2012, 45, 272–282. [Google Scholar] [CrossRef]
- Perreault, F.; de Faria, A.F.; Nejati, S.; Elimelech, M. Antimicrobial Properties of Graphene Oxide Nanosheets: Why Size Matters. ACS Nano 2015, 9, 7226–7236. [Google Scholar] [CrossRef]
- Barbolina, I.; Woods, C.R.; Lozano, N.; Kostarelos, K.; Novoselov, K.S.; Roberts, I.S. Purity of graphene oxide determines its antibacterial activity. Material 2016, 3, 025025. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.H.; Yang, S.T.; Wang, H.F.; Chang, Y.L.; Cao, A.N.; Liu, Y.F. Effect of size and dose on the biodistribution of graphene oxide in mice. Nanomedicine 2012, 7, 1801–1812. [Google Scholar] [CrossRef]
- Liu, S.B.; Hu, M.; Zeng, T.H.; Wu, R.; Jiang, R.R.; Wei, J.; Wang, L.; Kong, J.; Chen, Y. Lateral Dimension-Dependent Antibacterial Activity of Graphene Oxide Sheets. Langmuir 2012, 28, 12364–12372. [Google Scholar] [CrossRef]
- Sydlik, S.A.; Jhunjhunwala, S.; Webber, M.J.; Anderson, D.G.; Langer, R. In Vivo Compatibility of Graphene Oxide with Differing Oxidation States. ACS Nano 2015, 9, 3866–3874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, Z.L.; Xie, C.J.; Zhang, P.; Zhang, J.Z.; Wang, G.H.; He, X.; Ma, Y.H.; Zhao, B.; Zhang, Z.Y. Toxicity and transformation of graphene oxide and reduced graphene oxide in bacteria biofilm. Sci. Total Environ. 2017, 580, 1300–1308. [Google Scholar] [CrossRef]
- National Research Council. Guide for the Care and Use of Laboratory Animals, 8th ed.; National Academies Press: Washington, DC, USA, 2011. [Google Scholar]
- Krewski, D.; Acosta, D., Jr.; Andersen, M.; Anderson, H.; Bailar, J.C., 3rd; Boekelheide, K.; Brent, R.; Charnley, G.; Cheung, V.G.; Green, S., Jr.; et al. Toxicity testing in the 21st century: A vision and a strategy. J. Toxicol. Environ. Health Part B Crit. Rev. 2010, 13, 51–138. [Google Scholar] [CrossRef]
- Fourches, D.; Pu, D.; Tropsha, A. Exploring quantitative nanostructure-activity relationships (QNAR) modeling as a tool for predicting biological effects of manufactured nanoparticles. Comb. Chem. High Throughput Screen. 2011, 14, 217–225. [Google Scholar] [CrossRef]
- Schmidt, C.W. TOX 21: New dimensions of toxicity testing. Environ. Health Perspect. 2009, 117, A348–A353. [Google Scholar] [CrossRef] [Green Version]
- Fujita, K.; Fukuda, M.; Fukui, H.; Horie, M.; Endoh, S.; Uchida, K.; Shichiri, M.; Morimoto, Y.; Ogami, A.; Iwahashi, H. Intratracheal instillation of single-wall carbon nanotubes in the rat lung induces time-dependent changes in gene expression. Nanotoxicology 2015, 9, 290–301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shannahan, J.H.; Brown, J.M.; Chen, R.; Ke, P.C.; Lai, X.Y.; Mitra, S.; Witzmann, F.A. Comparison of Nanotube-Protein Corona Composition in Cell Culture Media. Small 2013, 9, 2171–2181. [Google Scholar] [CrossRef] [PubMed]
- Blazer-Yost, B.L.; Banga, A.; Amos, A.; Chernoff, E.; Lai, X.Y.; Li, C.; Mitra, S.; Witzmann, F.A. Effect of carbon nanoparticles on renal epithelial cell structure, barrier function, and protein expression. Nanotoxicology 2011, 5, 354–371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nicholson, J.K.; Lindon, J.C. Systems biology—Metabonomics. Nature 2008, 455, 1054–1056. [Google Scholar] [CrossRef]
- Sui, J.; Zhang, Y.Q.; Li, C.Y.; Fu, Y.Y.; Ma, S.M.; Tang, M.; Yin, L.H.; Pu, Y.P.; Liang, G.Y. Metabolic Characteristics in Serum of Rats Intratracheally Instilled with Multi-Walled Carbon Nanotubes (MWCNT). J. Nanosci. Nanotechnol. 2017, 17, 9236–9243. [Google Scholar] [CrossRef]
- Gou, N.; Gu, A.Z. A New Transcriptional Effect Level Index (TELI) for Toxicogenomics-based Toxicity Assessment. Environ. Sci. Technol. 2011, 45, 5410–5417. [Google Scholar] [CrossRef]
- Onnis-Hayden, A.; Weng, H.; He, M.; Hansen, S.; Ilyin, V.; Lewis, K.; Gu, A.Z. Prokaryotic real-time gene expression profiling for toxicity assessment. Environ. Sci. Technol. 2009, 43, 4574–4581. [Google Scholar] [CrossRef]
- Gou, N.; Onnis-Hayden, A.; Gu, A.Z. Mechanistic toxicity assessment of nanomaterials by whole-cell-array stress genes expression analysis. Environ. Sci. Technol. 2010, 44, 5964–5970. [Google Scholar] [CrossRef] [PubMed]
- Lan, J.; Gou, N.; Gao, C.; He, M.; Gu, A.Z. Comparative and mechanistic genotoxicity assessment of nanomaterials via a quantitative toxicogenomics approach across multiple species. Environ. Sci. Technol. 2014, 48, 12937–12945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lan, J.; Gou, N.; Rahman, S.M.; Gao, C.; He, M.; Gu, A.Z. A Quantitative Toxicogenomics Assay for High-throughput and Mechanistic Genotoxicity Assessment and Screening of Environmental Pollutants. Environ. Sci. Technol. 2016, 50, 3202–3214. [Google Scholar] [CrossRef] [PubMed]
- Lan, J.; Hu, M.; Gao, C.; Alshawabkeh, A.; Gu, A.Z. Toxicity Assessment of 4-Methyl-1-cyclohexanemethanol and Its Metabolites in Response to a Recent Chemical Spill in West Virginia, USA. Environ. Sci. Technol. 2015, 49, 6284–6293. [Google Scholar] [CrossRef] [PubMed]
- Lan, J.; Rahman, S.M.; Gou, N.; Jiang, T.; Plewa, M.J.; Alshawabkeh, A.; Gu, A.Z. Genotoxicity Assessment of Drinking Water Disinfection Byproducts by DNA Damage and Repair Pathway Profiling Analysis. Environ. Sci. Technol. 2018, 52, 6565–6575. [Google Scholar] [CrossRef] [PubMed]
- Jiang, T.; Amadei, C.A.; Gou, N.; Lin, Y.; Lan, J.; Vecitis, C.D.; Gu, A.Z. Toxicity of single-walled carbon nanotubes (SWCNTs): Effect of lengths, functional groups and electronic structures revealed by a quantitative toxicogenomics assay. Environ. Sci. Nano 2020, 7, 1348–1364. [Google Scholar] [CrossRef]
- Jiang, T.; Lin, Y.; Amadei, C.A.; Gou, N.; Rahman, S.M.; Lan, J.; Vecitis, C.D.; Gu, A.Z. Comparative and mechanistic toxicity assessment of structure-dependent toxicity of carbon-based nanomaterials. J. Hazard. Mater. 2021, 418, 126282. [Google Scholar] [CrossRef]
- Ding, Z.J.; Zhang, Z.J.; Ma, H.W.; Chen, Y.Y. In Vitro Hemocompatibility and Toxic Mechanism of Graphene Oxide on Human Peripheral Blood T Lymphocytes and Serum Albumin. ACS Appl. Mater. Interfaces 2014, 6, 19797–19807. [Google Scholar] [CrossRef]
- Olive, P.L.; Banáth, J.P. The comet assay: A method to measure DNA damage in individual cells. Nat. Protoc. 2006, 1, 23–29. [Google Scholar] [CrossRef]
- Collins, A.R. The comet assay for DNA damage and repair. Mol. Biotechnol. 2004, 26, 249. [Google Scholar] [CrossRef]
- Chen, Y.M.; Hu, X.G.; Sun, J.; Zhou, Q.X. Specific nanotoxicity of graphene oxide during zebrafish embryogenesis. Nanotoxicology 2016, 10, 42–52. [Google Scholar] [CrossRef]
- Wang, A.X.; Pu, K.F.; Dong, B.; Liu, Y.; Zhang, L.M.; Zhang, Z.J.; Duan, W.; Zhu, Y.M. Role of surface charge and oxidative stress in cytotoxicity and genotoxicity of graphene oxide towards human lung fibroblast cells. J. Appl. Toxicol. 2013, 33, 1156–1164. [Google Scholar] [CrossRef] [PubMed]
- Qiao, Y.; An, J.C.; Ma, L.Y. Single Cell Array Based Assay for in Vitro Genotoxicity Study of Nanomaterials. Anal. Chem. 2013, 85, 4107–4112. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.W.; Chen, L.Y.; Hu, X.; Feng, S.C.; Huang, L.; Quan, G.P.; Wei, X.; Yang, S.T. Toxicity of graphene oxide to white moss Leucobryum glaucum. RSC Adv. 2017, 7, 50287–50293. [Google Scholar] [CrossRef] [Green Version]
- Akhavan, O.; Ghaderi, E.; Akhavan, A. Size-dependent genotoxicity of graphene nanoplatelets in human stem cells. Biomaterials 2012, 33, 8017–8025. [Google Scholar] [CrossRef]
- Katsumiti, A.; Tomovska, R.; Cajaraville, M.P. Intracellular localization and toxicity of graphene oxide and reduced graphene oxide nanoplatelets to mussel hemocytes in vitro. Aquat. Toxicol. 2017, 188, 138–147. [Google Scholar] [CrossRef]
- Yu, Q.L.; Zhang, B.; Li, J.R.; Du, T.T.; Yi, X.; Li, M.C.; Chen, W.; Alvarez, P.J.J. Graphene oxide significantly inhibits cell growth at sublethal concentrations by causing extracellular iron deficiency. Nanotoxicology 2017, 11, 1102–1114. [Google Scholar] [CrossRef]
- Cheng, C.E.; Tsai, C.W.; Pei, Z.W.; Lin, T.W.; Chang, C.S.; Chien, F.S.S. UV-treated graphene oxide as anode interfacial layers for P3HT: PCBM solar cells. J. Phys. D Appl. Phys. 2015, 48, 255103. [Google Scholar] [CrossRef]
- Zhang, Y.B.; Ali, S.F.; Dervishi, E.; Xu, Y.; Li, Z.R.; Casciano, D.; Biris, A.S. Cytotoxicity Effects of Graphene and Single-Wall Carbon Nanotubes in Neural Phaeochromocytoma-Derived PC12 Cells. ACS Nano 2010, 4, 3181–3186. [Google Scholar] [CrossRef]
- Duch, M.C.; Budinger, G.R.S.; Liang, Y.T.; Soberanes, S.; Urich, D.; Chiarella, S.E.; Campochiaro, L.A.; Gonzalez, A.; Chandel, N.S.; Hersam, M.C.; et al. Minimizing Oxidation and Stable Nanoscale Dispersion Improves the Biocompatibility of Graphene in the Lung. Nano Lett. 2011, 11, 5201–5207. [Google Scholar] [CrossRef] [Green Version]
- Du, S.T.; Zhang, P.; Zhang, R.R.; Lu, Q.; Liu, L.; Bao, X.W.; Liu, H.J. Reduced graphene oxide induces cytotoxicity and inhibits photosynthetic performance of the green alga Scenedesmus obliquus. Chemosphere 2016, 164, 499–507. [Google Scholar] [CrossRef]
- Kang, Y.Y.; Liu, J.; Wu, J.R.; Yin, Q.; Liang, H.M.; Chen, A.J.; Shao, L.Q. Graphene oxide and reduced graphene oxide induced neural pheochromocytoma-derived PC12 cell lines apoptosis and cell cycle alterations via the ERK signaling pathways. Int. J. Nanomed. 2017, 12, 5501–5510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Contreras-Torres, F.F.; Rodriguez-Galvan, A.; Guerrero-Beltran, C.E.; Martinez-Loran, E.; Vazquez-Garza, E.; Ornelas-Soto, N.; Garcia-Rivas, G. Differential cytotoxicity and internalization of graphene family nanomaterials in myocardial cells. Mat. Sci. Eng. C-Mater. 2017, 73, 633–642. [Google Scholar] [CrossRef]
- Jiang, Y.; Raliya, R.; Fortner, J.D.; Biswas, P. Graphene Oxides in Water: Correlating Morphology and Surface Chemistry with Aggregation Behavior. Environ. Sci. Technol. 2016, 50, 6964–6973. [Google Scholar] [CrossRef] [PubMed]
- Schniepp, H.C.; Li, J.L.; McAllister, M.J.; Sai, H.; Herrera-Alonso, M.; Adamson, D.H.; Prud’homme, R.K.; Car, R.; Saville, D.A.; Aksay, I.A. Functionalized single graphene sheets derived from splitting graphite oxide. J. Phys. Chem. B 2006, 110, 8535–8539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hou, W.C.; Lee, P.L.; Chou, Y.C.; Wang, Y.S. Antibacterial property of graphene oxide: The role of phototransformation. Environ. Sci.-Nano 2017, 4, 647–657. [Google Scholar] [CrossRef]
- Mao, J.; Guo, R.H.; Yan, L.T. Simulation and analysis of cellular internalization pathways and membrane perturbation for graphene nanosheets. Biomaterials 2014, 35, 6069–6077. [Google Scholar] [CrossRef]
- Liu, W.T.; Bien, M.Y.; Chuang, K.J.; Chang, T.Y.; Jones, T.; BeruBe, K.; Lalev, G.; Tsai, D.H.; Chuang, H.C.; Cheng, T.J.; et al. Physicochemical and biological characterization of single-walled and double-walled carbon nanotubes in biological media. J. Hazard. Mater. 2014, 280, 216–225. [Google Scholar] [CrossRef]
- Wang, R.H.; Mikoryak, C.; Li, S.Y.; Bushdiecker, D.; Musselman, I.H.; Pantano, P.; Draper, R.K. Cytotoxicity Screening of Single-Walled Carbon Nanotubes: Detection and Removal of Cytotoxic Contaminants from Carboxylated Carbon Nanotubes. Mol. Pharm. 2011, 8, 1351–1361. [Google Scholar] [CrossRef] [Green Version]
- Nikitin, A.; Ogasawara, H.; Mann, D.; Denecke, R.; Zhang, Z.; Dai, H.; Cho, K.; Nilsson, A. Hydrogenation of single-walled carbon nanotubes. Phys. Rev. Lett. 2005, 95, 225507. [Google Scholar] [CrossRef] [PubMed]
- Hohmann, S.M.; Willem, H. Yeast Stress Responses; Springer: Berlin/Heidelberg, Germany, 2003. [Google Scholar]
- Gasch, A.P.; Spellman, P.T.; Kao, C.M.; Carmel-Harel, O.; Eisen, M.B.; Storz, G.; Botstein, D.; Brown, P.O. Genomic expression programs in the response of yeast cells to environmental changes. Mol. Biol. Cell 2000, 11, 4241–4257. [Google Scholar] [CrossRef]
- Lucau-Danila, A.; Lelandais, G.; Kozovska, Z.; Tanty, V.; Delaveau, T.; Devaux, F.; Jacq, C. Early expression of yeast genes affected by chemical stress. Mol. Cell Biol. 2005, 25, 1860–1868. [Google Scholar] [CrossRef] [Green Version]
- Huh, W.K.; Falvo, J.V.; Gerke, L.C.; Carroll, A.S.; Howson, R.W.; Weissman, J.S.; O’Shea, E.K. Global analysis of protein localization in budding yeast. Nature 2003, 425, 686–691. [Google Scholar] [CrossRef]
- Salamone, M.; Heddle, J.; Stuart, E.; Katz, M. Towards an improved micronucleus test: Studies on 3 model agents, mitomycin C, cyclophosphamide and dimethylbenzanthracene. Mutat. Res. 1980, 74, 347–356. [Google Scholar] [CrossRef]
- Godon, C.; Lagniel, G.; Lee, J.; Buhler, J.M.; Kieffer, S.; Perrot, M.; Boucherie, H.; Toledano, M.B.; Labarre, J. The H2O2 stimulon in Saccharomyces cerevisiae. J. Biol. Chem. 1998, 273, 22480–22489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ling, L.U.; Tan, K.B.; Lin, H.; Chiu, G.N. The role of reactive oxygen species and autophagy in safingol-induced cell death. Cell Death Dis. 2011, 2, e129. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Joseph, J.A. Quantifying cellular oxidative stress by dichlorofluorescein assay using microplate reader. Free Radic. Biol. Med. 1999, 27, 612–616. [Google Scholar] [CrossRef]
- Dhawan, A.; Bajpayee, M.M.; Pandey, A.K.; Parmar, D. Protocol for the single cell gel electrophoresis/comet assay for rapid genotoxicity assessment. ITRC SCGE/Comet Assay Protoc. 2009, 1077, 1–10. [Google Scholar]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S.; et al. Gene set enrichment analysis: A knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Huang, Q.; Liu, Z.P.; Wang, Y.; Wu, L.Y.; Chen, L.; Zhang, X.S. NOA: A novel Network Ontology Analysis method. Nucleic Acids Res. 2011, 39, e87. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GOs | Untreated Control | UV-Treated | Thermally Reduced | 45 min Sonicated | 2.5 h Sonicated | |
|---|---|---|---|---|---|---|
| Oxygen content (%) | 36.1 ± 0.1 | 35.0 ± 0.7 | 33.9 ± 0.1 | 36.1 ± 0.1 | 36.1 ± 0.1 | |
| O/C ratio (atomic) | 0.60 | 0.57 | 0.55 | 0.60 | 0.60 | |
| Impurities | N < 1%, S = 2–3% | N < 1%, S = 2–3% | N < 1%, S = 2–3% | N < 1%, S = 2–3% | N < 1%, S = 2–3% | |
| Monolayer (%) | >90 | >90 | >90 | >90 | >90 | |
| C-O-C and C-OH (%) | 62 ± 2 | 51 ± 2 | 57 ± 2 | 62 ± 2 | 62 ± 2 | |
| C-C and C=C (%) | 34 ± 2 | 42 ± 2 | 38 ± 2 | 34 ± 2 | 34 ± 2 | |
| C-COOH (%) | 4 ± 2 | 7 ± 2 | 6 ± 2 | 4 ± 2 | 4 ± 2 | |
| Zeta potential (mV) | −7.19 ± 0.46 | −5.46 ± 0.36 | −5.53 ± 0.36 | −6.11 ± 0.09 | −7.39 ± 0.56 | |
| Conductivity (mS/cm) | 10.21 ± 0.28 | 10.40 ± 0.29 | 10.23 ± 0.17 | 10.67 ± 0.09 | 9.72 ± 0.25 | |
| Size, Z-average (nm) | 0 h | 1331 | 1627 | 1461 | 447 | 348 |
| 2 h | 1610 | 1669 | 1479 | 471 | 352 | |
| GOs | PELI1.5 (mg/L) | TEQ1.5 | ||||||
|---|---|---|---|---|---|---|---|---|
| Chemical | General | DNA | Oxidative | Protein | Total | Geno- | Oxi- | |
| Untreated control | 0.13 | 9.17 | 2.09 | 16.01 | 0.16 | 6.06 | 0.34 | 0.21 |
| UV-treated | 1.71 | N/A | 1.42 | 12.13 | 6.32 | 4.48 | 0.51 | 0.28 |
| Thermally reduced | 1.47 | N/A | 0.98 | 18.09 | N/A | 4.85 | 0.73 | 0.19 |
| 45 min sonicated | N/A | <0.031 | 0.59 | N/A | N/A | 4.83 | 1.22 | N/A |
| 2.5 h sonicated | 4.36 | 0.07 | <0.031 | <0.031 | 13.61 | <0.031 | >23.19 | >109.77 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, T.; Amadei, C.A.; Lin, Y.; Gou, N.; Rahman, S.M.; Lan, J.; Vecitis, C.D.; Gu, A.Z. Dependence of Graphene Oxide (GO) Toxicity on Oxidation Level, Elemental Composition, and Size. Int. J. Mol. Sci. 2021, 22, 10578. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms221910578
Jiang T, Amadei CA, Lin Y, Gou N, Rahman SM, Lan J, Vecitis CD, Gu AZ. Dependence of Graphene Oxide (GO) Toxicity on Oxidation Level, Elemental Composition, and Size. International Journal of Molecular Sciences. 2021; 22(19):10578. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms221910578
Chicago/Turabian StyleJiang, Tao, Carlo Alberto Amadei, Yishan Lin, Na Gou, Sheikh Mokhlesur Rahman, Jiaqi Lan, Chad D. Vecitis, and April Z. Gu. 2021. "Dependence of Graphene Oxide (GO) Toxicity on Oxidation Level, Elemental Composition, and Size" International Journal of Molecular Sciences 22, no. 19: 10578. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms221910578