
Anti-Osteoporotic Effect of Morroniside on Osteoblast and Osteoclast Differentiation In Vitro and Ovariectomized Mice In Vivo

 , and
, and 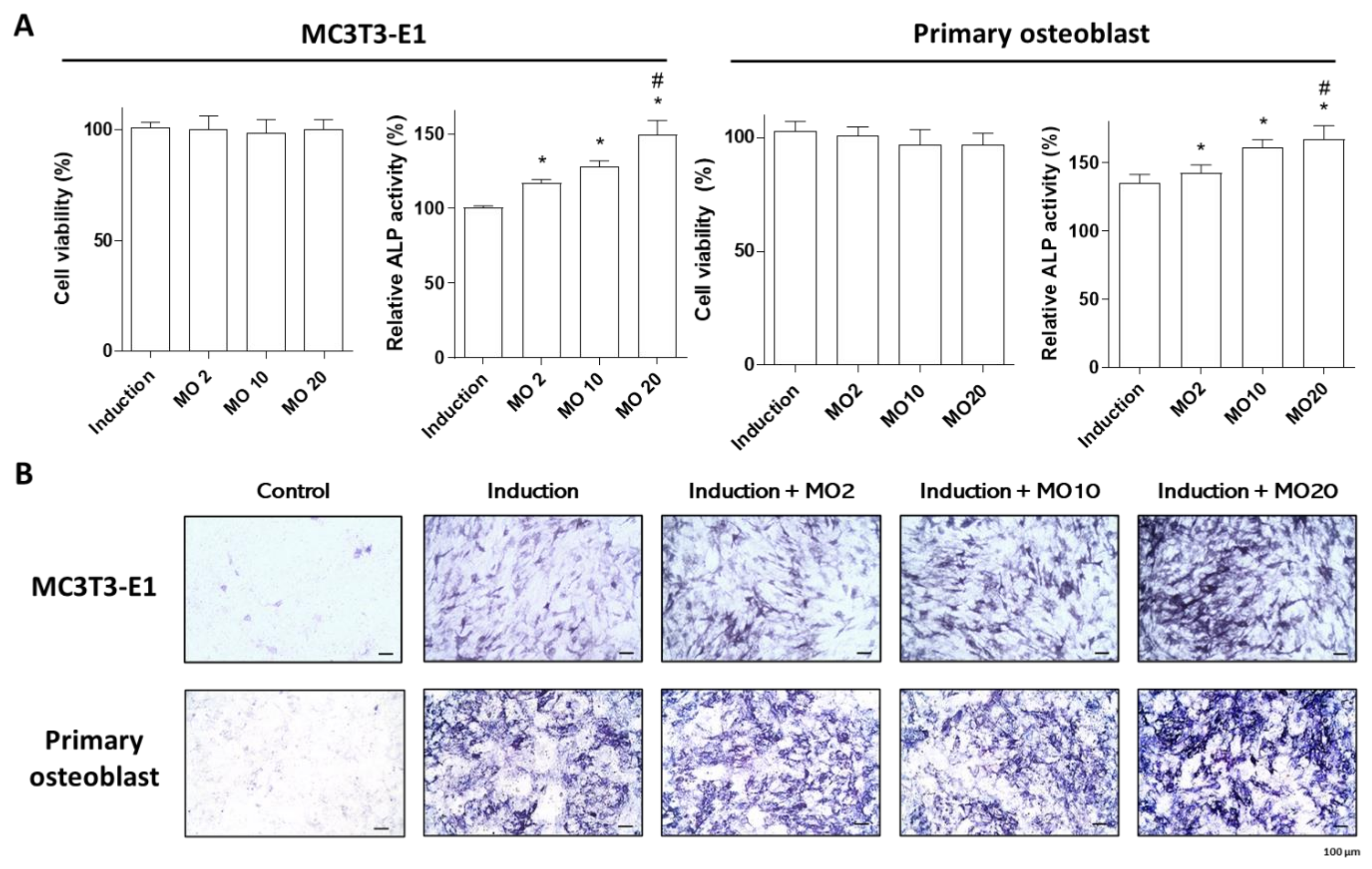{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
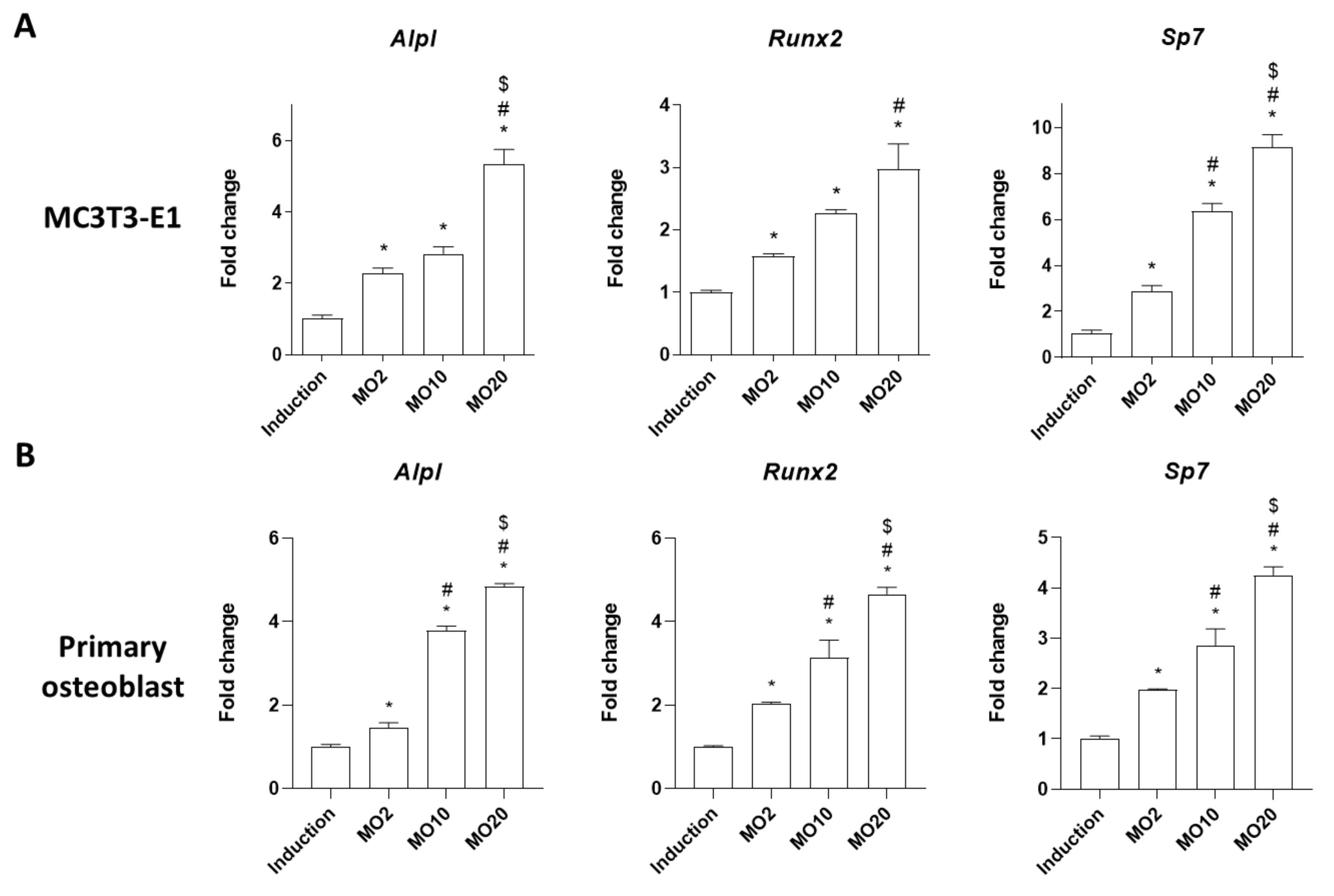
2.1. Morroniside Elevated Osteogenic Differentiation in MC3T3-E1 Cells and Mouse Primary Cultured Osteoblasts
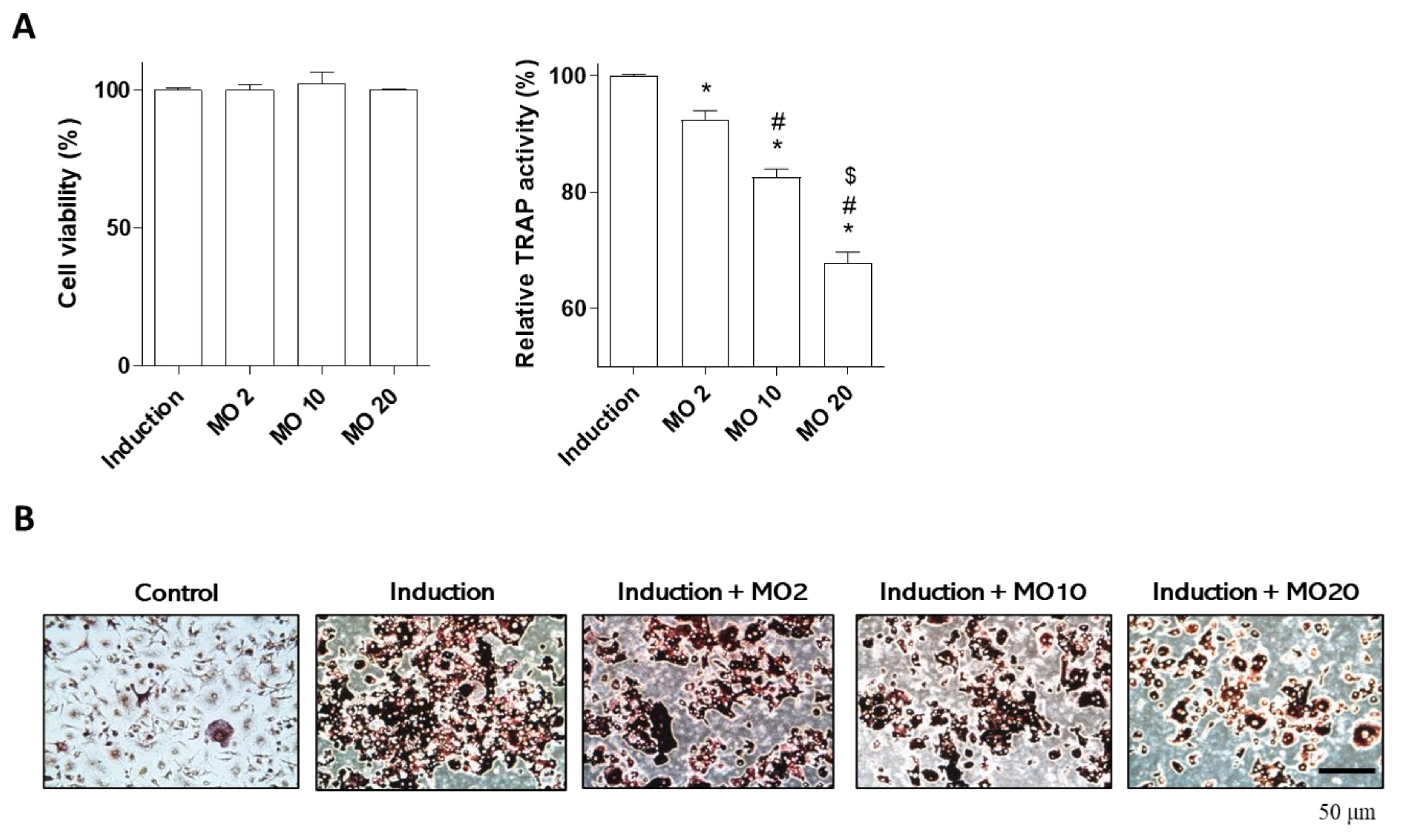
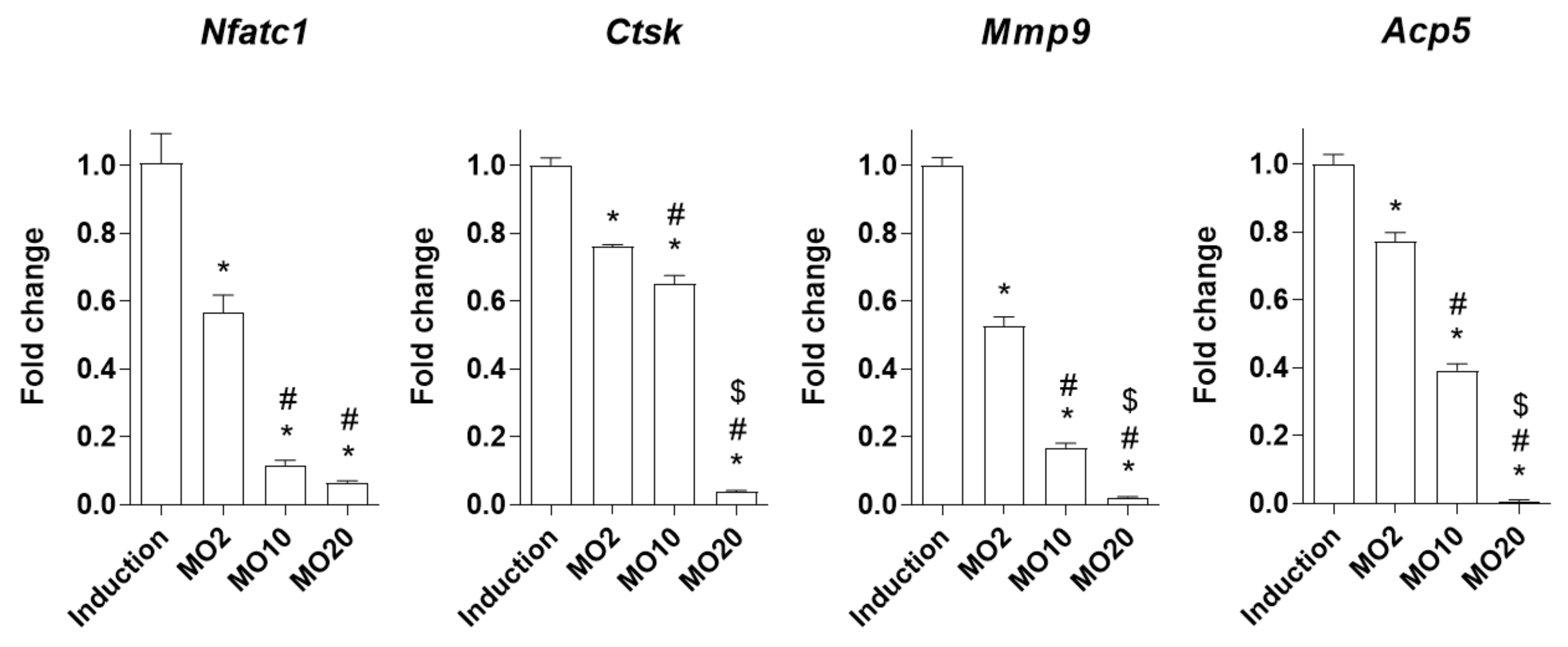
2.2. Morroniside Prevented Osteoclast Differentiation in Mouse Primary Monocytes
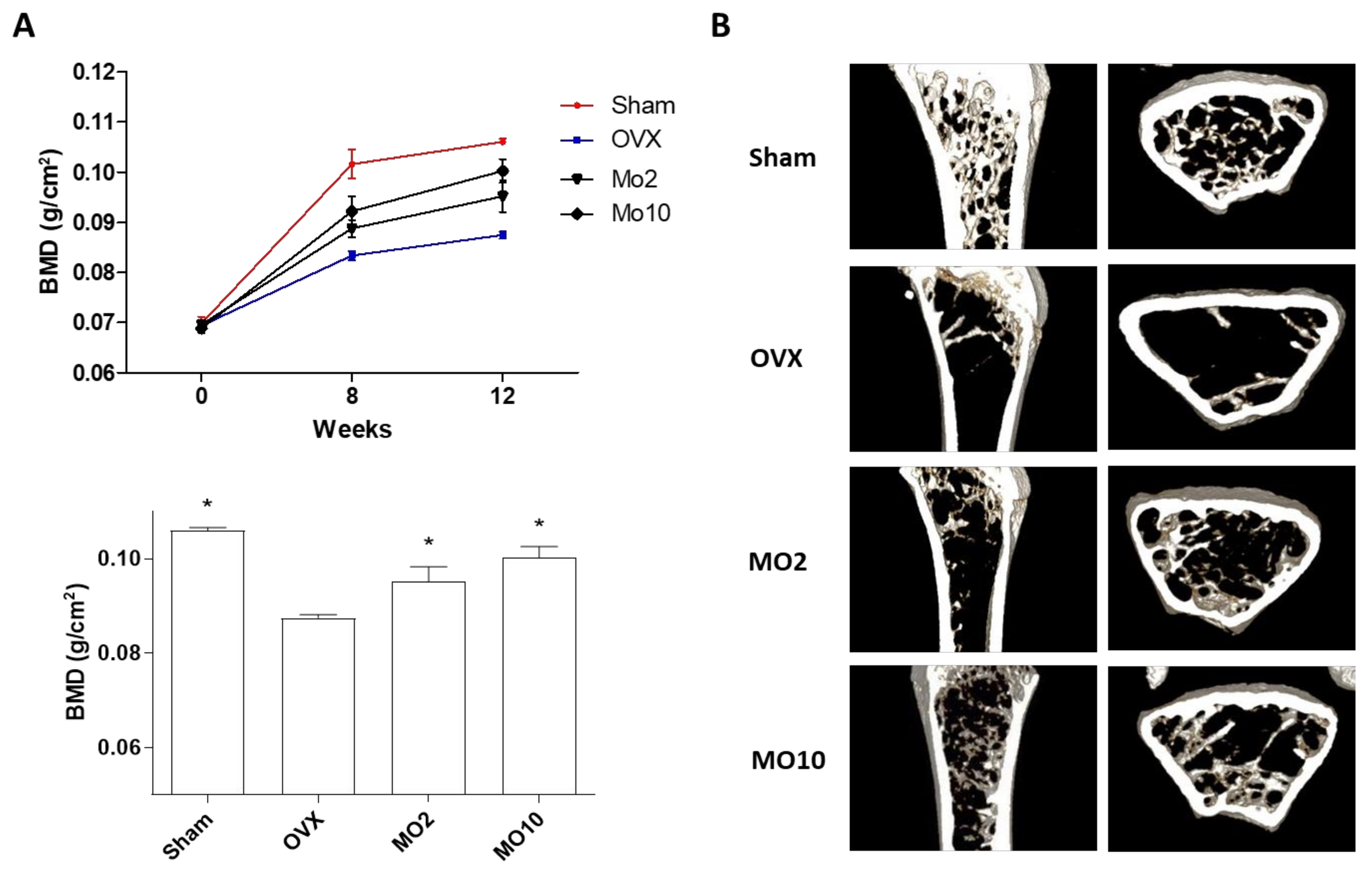
2.3. Oral Administration of Morroniside Attenuated Ovariectomy (OVX)-Induced Mouse Osteoporosis
3. Materials and Methods
3.1. Cell Culture
3.2. Osteoblast and Osteoclast Differentiation
3.3. Cell Viability
3.4. ALP/TRAP Activity and Staining
3.5. Quantitative Reverse Transcriptase Polymerase Chain Reaction (qRT-PCR) Analysis
3.6. Animal Study
3.7. Bone Mineral Density (BMD) and micro-CT Analysis
3.8. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kim, J.M.; Lin, C.; Stavre, Z.; Greenblatt, M.B.; Shim, J.H. Osteoblast-Osteoclast communication and bone homeostasis. Cells 2020, 9, 73. [Google Scholar] [CrossRef] [Green Version]
- Compston, J.E.; McClung, M.R.; Leslie, W.D. Osteoporosis. Lancet 2019, 393, 364–376. [Google Scholar] [CrossRef]
- Ensrud, K.E.; Crandall, C.J. Osteoporosis. Ann. Intern. Med. 2017, 167, ITC17–ITC32. [Google Scholar] [CrossRef]
- Khosla, S.; Hofbauer, L.C. Osteoporosis treatment: Recent developments and ongoing challenges. Lancet Diabetes Endocrinol. 2017, 5, 898–907. [Google Scholar] [CrossRef] [Green Version]
- Sozen, T.; Ozisik, L.; Basaran, N.C. An overview and management of osteoporosis. Eur. J. Rheumatol. 2017, 4, 46–56. [Google Scholar] [CrossRef]
- Xu, L.; Li, Y.; Dai, Y.; Peng, J. Natural products for the treatment of type 2 diabetes mellitus: Pharmacology and mechanisms. Pharmacol. Res. 2018, 130, 451–465. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.T.; Zheng, X.W.; Chen, S.; Shan, C.S.; Xu, Q.Q.; Zhu, J.Z.; Bao, X.Y.; Lin, Y.; Zheng, G.Q.; Wang, Y. Chinese herbal medicine for Alzheimer’s disease: Clinical evidence and possible mechanism of neurogenesis. Biochem. Pharmacol. 2017, 141, 143–155. [Google Scholar] [CrossRef] [PubMed]
- Tan, N.; Gwee, K.A.; Tack, J.; Zhang, M.; Li, Y.; Chen, M.; Xiao, Y. Herbal medicine in the treatment of functional gastrointestinal disorders: A systematic review with meta-analysis. J. Gastroenterol. Hepatol. 2020, 35, 544–556. [Google Scholar] [CrossRef] [PubMed]
- Parham, S.; Kharazi, A.Z.; Bakhsheshi-Rad, H.R.; Nur, H.; Ismail, A.F.; Sharif, S.; RamaKrishna, S.; Berto, F. Antioxidant, antimicrobial and antiviral properties of herbal materials. Antioxidants 2020, 9, 1309. [Google Scholar] [CrossRef]
- Liperoti, R.; Vetrano, D.L.; Bernabei, R.; Onder, G. Herbal medications in cardiovascular medicine. J. Am. Coll. Cardiol. 2017, 69, 1188–1199. [Google Scholar] [CrossRef]
- Pang, G.M.; Li, F.X.; Yan, Y.; Zhang, Y.; Kong, L.L.; Zhu, P.; Wang, K.F.; Zhang, F.; Liu, B.; Lu, C. Herbal medicine in the treatment of patients with type 2 diabetes mellitus. Chin. Med. J. 2019, 132, 78–85. [Google Scholar] [CrossRef]
- Han, Y.; Jung, H.W.; Park, Y.K. Selective therapeutic effect of Cornus officinalis fruits on the damage of different organs in STZ-induced diabetic rats. Am. J. Chin. Med. 2014, 42, 1169–1182. [Google Scholar] [CrossRef] [PubMed]
- Lee, N.H.; Seo, C.S.; Lee, H.Y.; Jung, D.Y.; Lee, J.K.; Lee, J.A.; Song, K.Y.; Shin, H.K.; Lee, M.Y.; Seo, Y.B.; et al. Hepatoprotective and antioxidative activities of Cornus officinalis against acetaminophen-induced hepatotoxicity in mice. Evid. Based Complement. Alternat. Med. 2012, 2012, 804924. [Google Scholar] [CrossRef] [Green Version]
- Park, E.; Lim, E.; Yeo, S.; Yong, Y.; Yang, J.; Jeong, S.Y. Anti-Menopausal effects of Cornus officinalis and Ribes fasciculatum extract in vitro and in vivo. Nutrients 2020, 12, 369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.Y.; Kim, Y.K.; Choi, M.K.; Oh, J.; Kwak, H.B.; Kim, J.J. Effect of Cornus officinalis on receptor activator of nuclear factor-kappaB ligand (RANKL)-induced osteoclast differentiation. J. Bone Metab. 2012, 19, 121–127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, J.; Zhang, Y.; Dong, L.; Gao, Q.; Yin, L.; Quan, H.; Chen, R.; Fu, X.; Lin, D. Ethnopharmacology, phytochemistry, and pharmacology of Cornus officinalis Sieb. et Zucc. J. Ethnopharmacol. 2018, 213, 280–301. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Liu, Y.; An, Z.; Ni, J. Active components and pharmacological effects of Cornus officinalis: Literature review. Front. Pharmacol. 2021, 12, 633447. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Li, X.; Lin, J.; Lin, M. Morroniside promotes the osteogenesis by activating PI3K/Akt/mTOR signaling. Biosci. Biotechnol. Biochem. 2021, 85, 332–339. [Google Scholar] [CrossRef]
- Sun, Y.; Zhu, Y.; Liu, X.; Chai, Y.; Xu, J. Morroniside attenuates high glucose-induced BMSC dysfunction by regulating the Glo1/AGE/RAGE axis. Cell Prolif. 2020, 53, e12866. [Google Scholar] [CrossRef]
- Li, M.; Wang, W.; Wang, P.; Yang, K.; Sun, H.; Wang, X. The pharmacological effects of morroniside and loganin isolated from Liuweidihuang Wan, on MC3T3-E1 cells. Molecules 2010, 15, 7403–7414. [Google Scholar] [CrossRef]
- Liu, B.; Lu, Y.; Wang, Y.; Ge, L.; Zhai, N.; Han, J. A protocol for isolation and identification and comparative characterization of primary osteoblasts from mouse and rat calvaria. Cell Tissue Bank. 2019, 20, 173–182. [Google Scholar] [CrossRef] [Green Version]
- Vimalraj, S. Alkaline phosphatase: Structure, expression and its function in bone mineralization. Gene 2020, 754, 144855. [Google Scholar] [CrossRef]
- Huang, W.; Yang, S.; Shao, J.; Li, Y.P. Signaling and transcriptional regulation in osteoblast commitment and differentiation. Front. Biosci. A J. Virtual Libr. 2007, 12, 3068–3092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Komori, T. Regulation of proliferation, differentiation and functions of osteoblasts by Runx2. Int. J. Mol. Sci. 2019, 20, 1694. [Google Scholar] [CrossRef] [Green Version]
- Komori, T. Regulation of osteoblast differentiation by transcription factors. J. Cell. Biochem. 2006, 99, 1233–1239. [Google Scholar] [CrossRef] [PubMed]
- Zuo, C.; Huang, Y.; Bajis, R.; Sahih, M.; Li, Y.P.; Dai, K.; Zhang, X. Osteoblastogenesis regulation signals in bone remodeling. Osteoporos Int. 2012, 23, 1653–1663. [Google Scholar] [CrossRef]
- Charles, J.F.; Aliprantis, A.O. Osteoclasts: More than ‘bone eaters’. Trends Mol. Med. 2014, 20, 449–459. [Google Scholar] [CrossRef] [Green Version]
- Kitaura, H.; Marahleh, A.; Ohori, F.; Noguchi, T.; Shen, W.R.; Qi, J.; Nara, Y.; Pramusita, A.; Kinjo, R.; Mizoguchi, I. Osteocyte-Related cytokines regulate osteoclast formation and bone resorption. Int. J. Mol. Sci. 2020, 21, 5169. [Google Scholar] [CrossRef]
- Yang, Y.; Yang, B. Anti-osteoporosis Effect of Ganoderma (Lingzhi) by inhibition of osteoclastogenesis. Adv. Exp. Med. Biol. 2019, 1182, 263–269. [Google Scholar] [CrossRef] [PubMed]
- Nara, Y.; Kitaura, H.; Ogawa, S.; Shen, W.R.; Qi, J.; Ohori, F.; Noguchi, T.; Marahleh, A.; Pramusita, A.; Kinjo, R.; et al. Anti-c-fms antibody prevents osteoclast formation and bone resorption in co-culture of osteoblasts and osteoclast precursors in vitro and in ovariectomized mice. Int. J. Mol. Sci. 2020, 21, 6120. [Google Scholar] [CrossRef]
- Zhuang, Y.; Sun, X.; Liu, B.; Hou, H.; Sun, Y. Effects of rambutan peel (Nepheliumlappaceum) phenolic extract on RANKL-induced differentiation of RAW264.7 cells into osteoclasts and retinoic acid-induced osteoporosis in rats. Nutrients 2020, 12, 883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gyori, D.S.; Mocsai, A. Osteoclast signal transduction during bone metastasis formation. Front. Cell. Dev. Biol. 2020, 8, 507. [Google Scholar] [CrossRef] [PubMed]
- Drissi, H.; Sanjay, A. The multifaceted osteoclast; Far and beyond bone resorption. J. Cell. Biochem. 2016, 117, 1753–1756. [Google Scholar] [CrossRef]
- Zhu, L.; Tang, Y.; Li, X.Y.; Keller, E.T.; Yang, J.; Cho, J.S.; Feinberg, T.Y.; Weiss, S.J. Osteoclast-mediated bone resorption is controlled by a compensatory network of secreted and membrane-tethered metalloproteinases. Sci. Transl. Med. 2020, 12, eaaw6143. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Wang, X.; Liu, Y.; He, A.; Jia, R. NFATc1: Functions in osteoclasts. Int. J. Biochem. Cell. Biol. 2010, 42, 576–579. [Google Scholar] [CrossRef]
- Komori, T. Animal models for osteoporosis. Eur. J. Pharmacol. 2015, 759, 287–294. [Google Scholar] [CrossRef]
- Kang, I.S.; Agidigbi, T.S.; Kwon, Y.M.; Kim, D.G.; Kim, R.I.; In, G.; Lee, M.H.; Kim, C. Effect of co-administration of Panax ginseng and Brassica oleracea on postmenopausal osteoporosis in ovariectomized mice. Nutrients 2020, 12, 2415. [Google Scholar] [CrossRef]
- Park, E.; Lee, C.G.; Han, S.J.; Yun, S.H.; Hwang, S.; Jeon, H.; Kim, J.; Choi, C.W.; Yang, S.; Jeong, S.Y. Antiosteoarthritic Effect of Morroniside in Chondrocyte Inflammation and Destabilization of Medial Meniscus-Induced Mouse Model. Int. J. Mol. Sci. 2021, 22, 2987. [Google Scholar] [CrossRef]
- Bakker, A.D.; Klein-Nulend, J. Osteoblast isolation from murine calvaria and long bones. Methods Mol. Biol. 2012, 816, 19–29. [Google Scholar] [CrossRef]
- Park, E.; Lee, C.G.; Kim, J.; Lim, E.; Hwang, S.; Yun, S.H.; Yong, Y.; Jeong, H.; Kim, J.A.; Jin, H.S.; et al. Anti-Osteoporotic effects of the herbal mixture of Cornus officinalis and Achyranthes japonica in vitro and in vivo. Plants 2020, 9, 1114. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, C.G.; Kim, J.; Yun, S.H.; Hwang, S.; Jeon, H.; Park, E.; Jeong, S.-Y. Anti-Osteoporotic Effect of Morroniside on Osteoblast and Osteoclast Differentiation In Vitro and Ovariectomized Mice In Vivo. Int. J. Mol. Sci. 2021, 22, 10642. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms221910642
Lee CG, Kim J, Yun SH, Hwang S, Jeon H, Park E, Jeong S-Y. Anti-Osteoporotic Effect of Morroniside on Osteoblast and Osteoclast Differentiation In Vitro and Ovariectomized Mice In Vivo. International Journal of Molecular Sciences. 2021; 22(19):10642. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms221910642
Chicago/Turabian StyleLee, Chang Gun, Jeonghyun Kim, Seung Hee Yun, Seokjin Hwang, Hyoju Jeon, Eunkuk Park, and Seon-Yong Jeong. 2021. "Anti-Osteoporotic Effect of Morroniside on Osteoblast and Osteoclast Differentiation In Vitro and Ovariectomized Mice In Vivo" International Journal of Molecular Sciences 22, no. 19: 10642. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms221910642