
TmSpz-like Plays a Fundamental Role in Response to E. coli but Not S. aureus or C. albican Infection in Tenebrio molitor via Regulation of Antimicrobial Peptide Production


 , , , and
, , , and
Abstract
:1. Introduction
2. Results
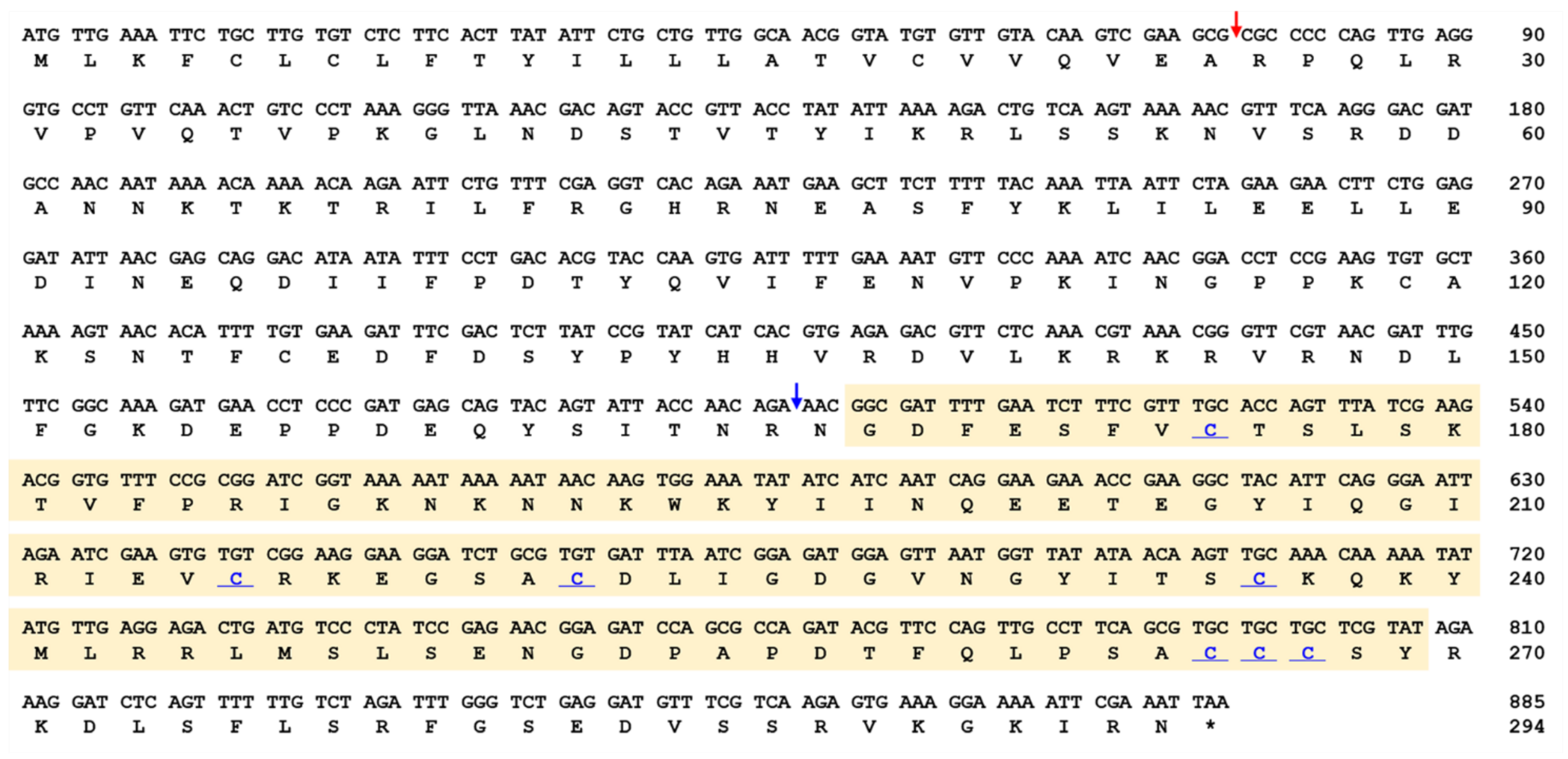
2.1. Cloning and Sequence Analysis of TmSpz-like
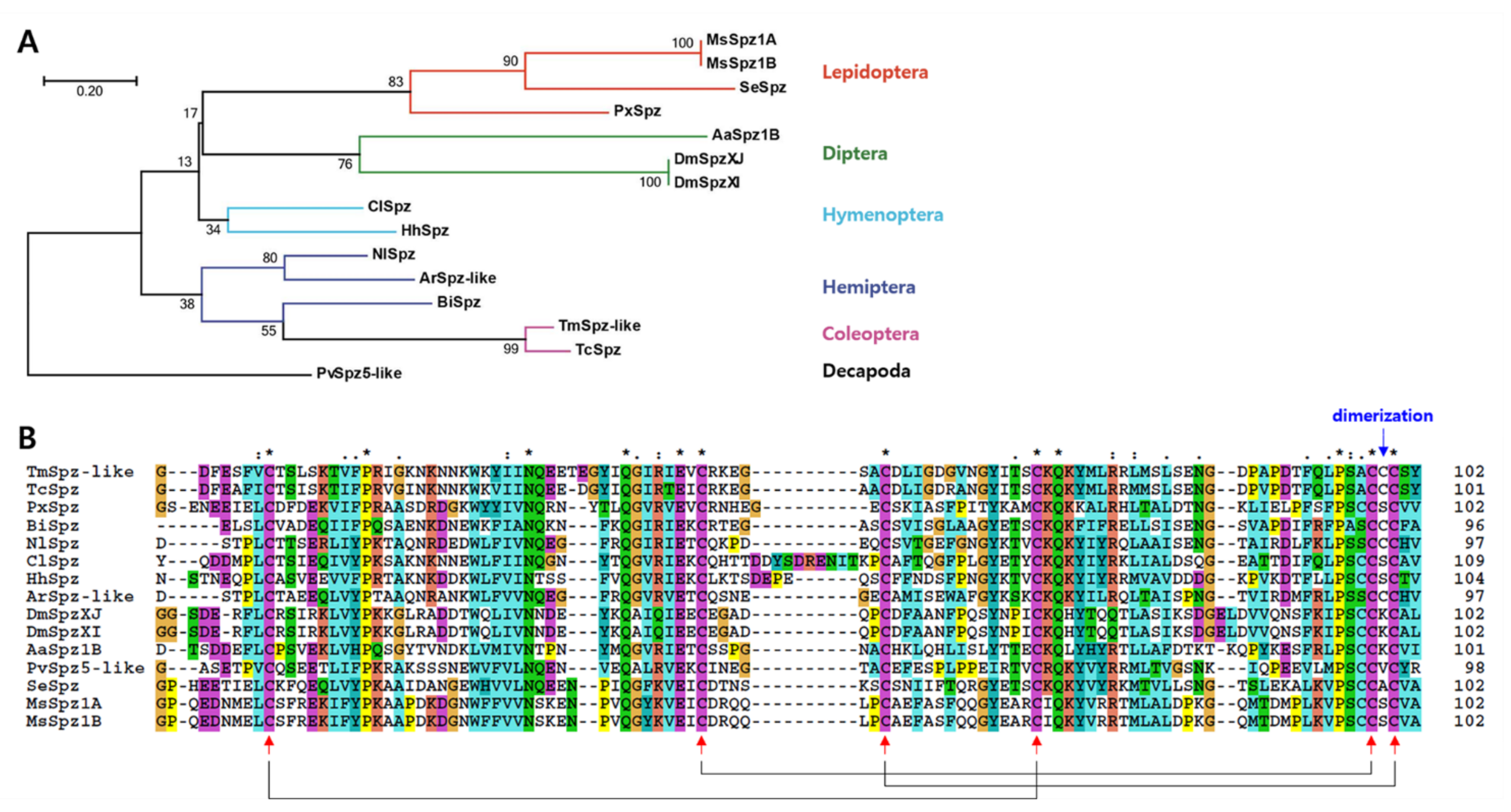
2.2. Phylogenetic Analysis of TmSpz-like
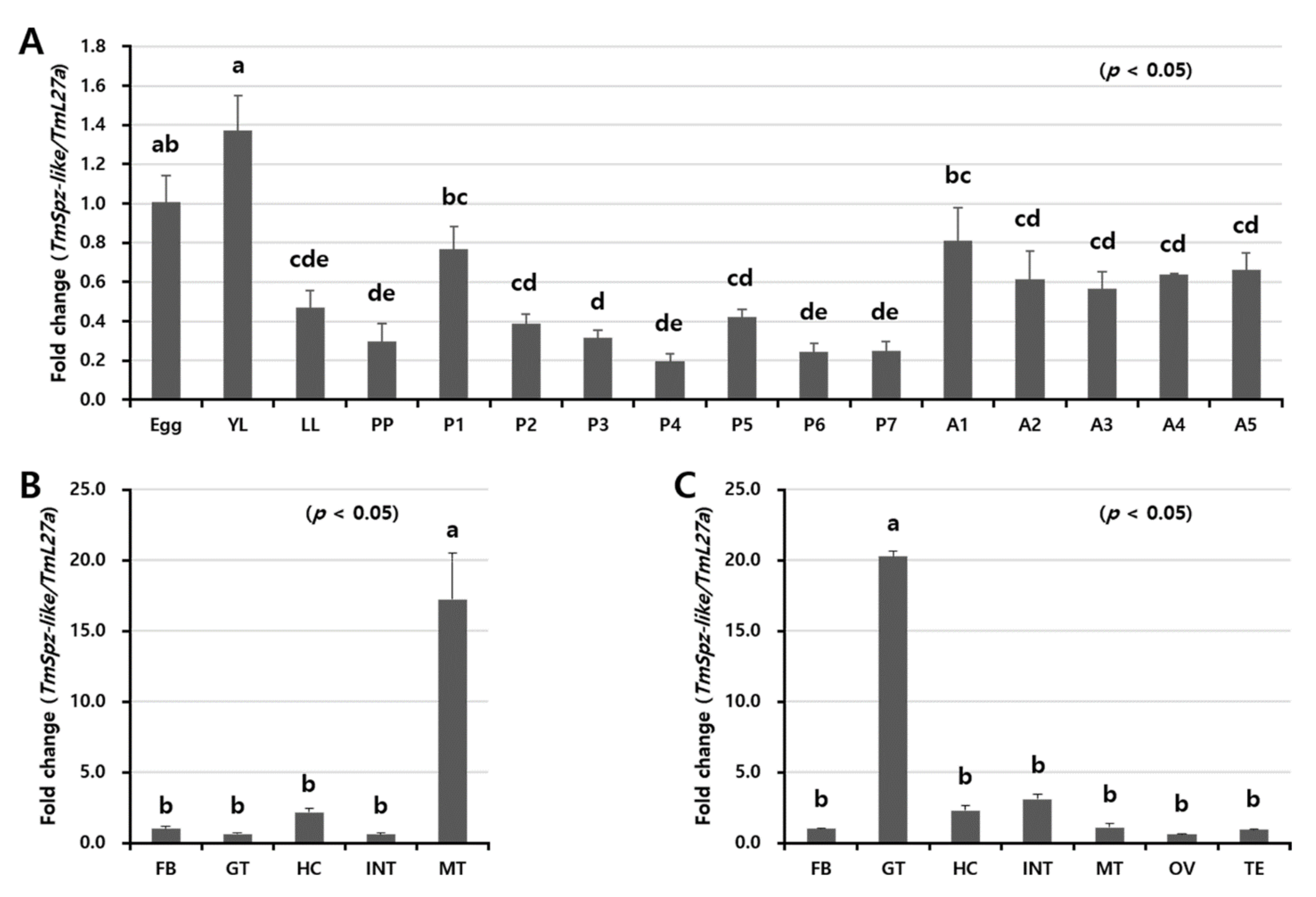
2.3. Developmental and Tissue-Specific Expression Patterns of TmSpz-like
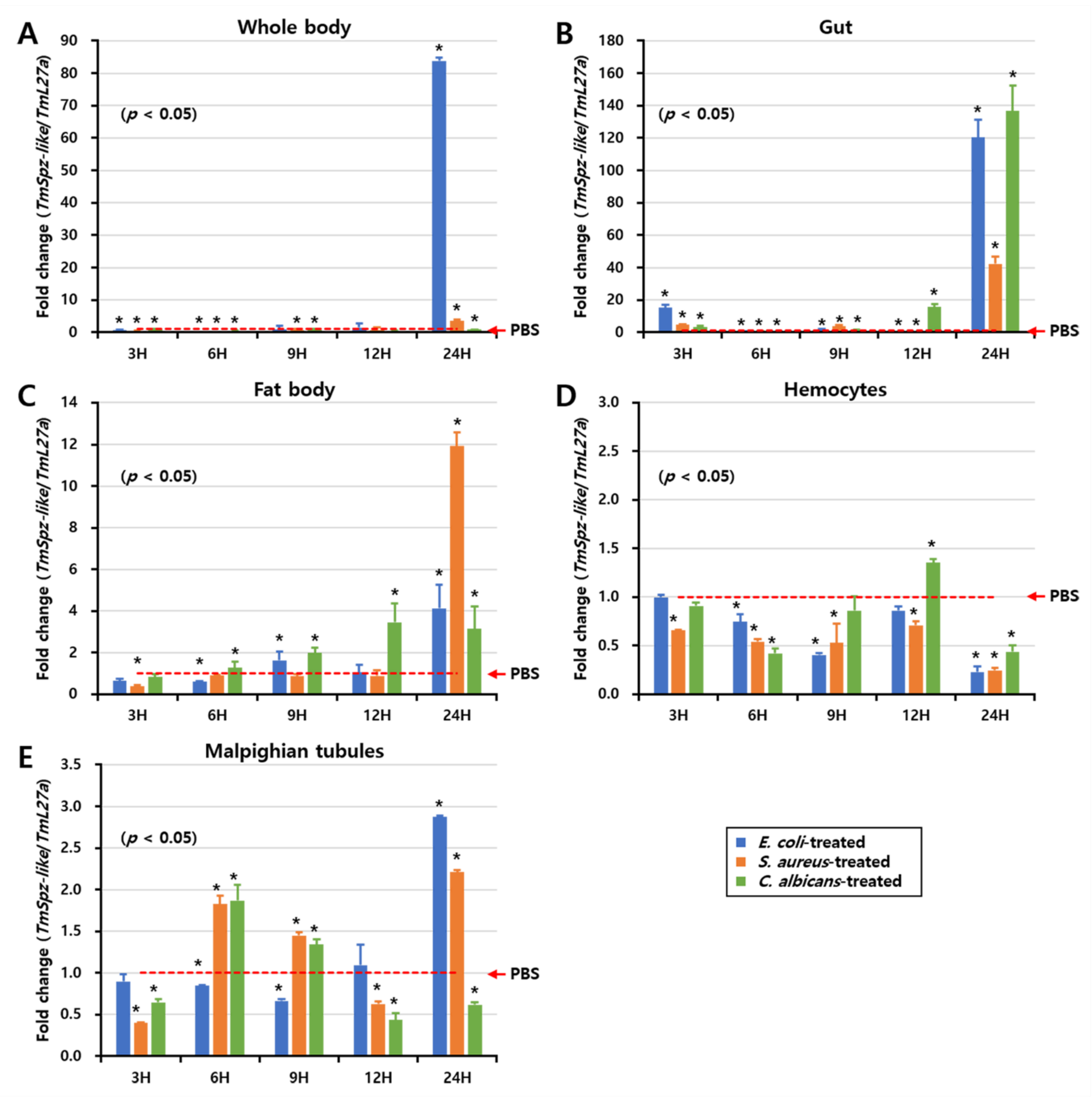
2.4. Effect of Microbial Infection on the Expression Patterns of TmSpz-like
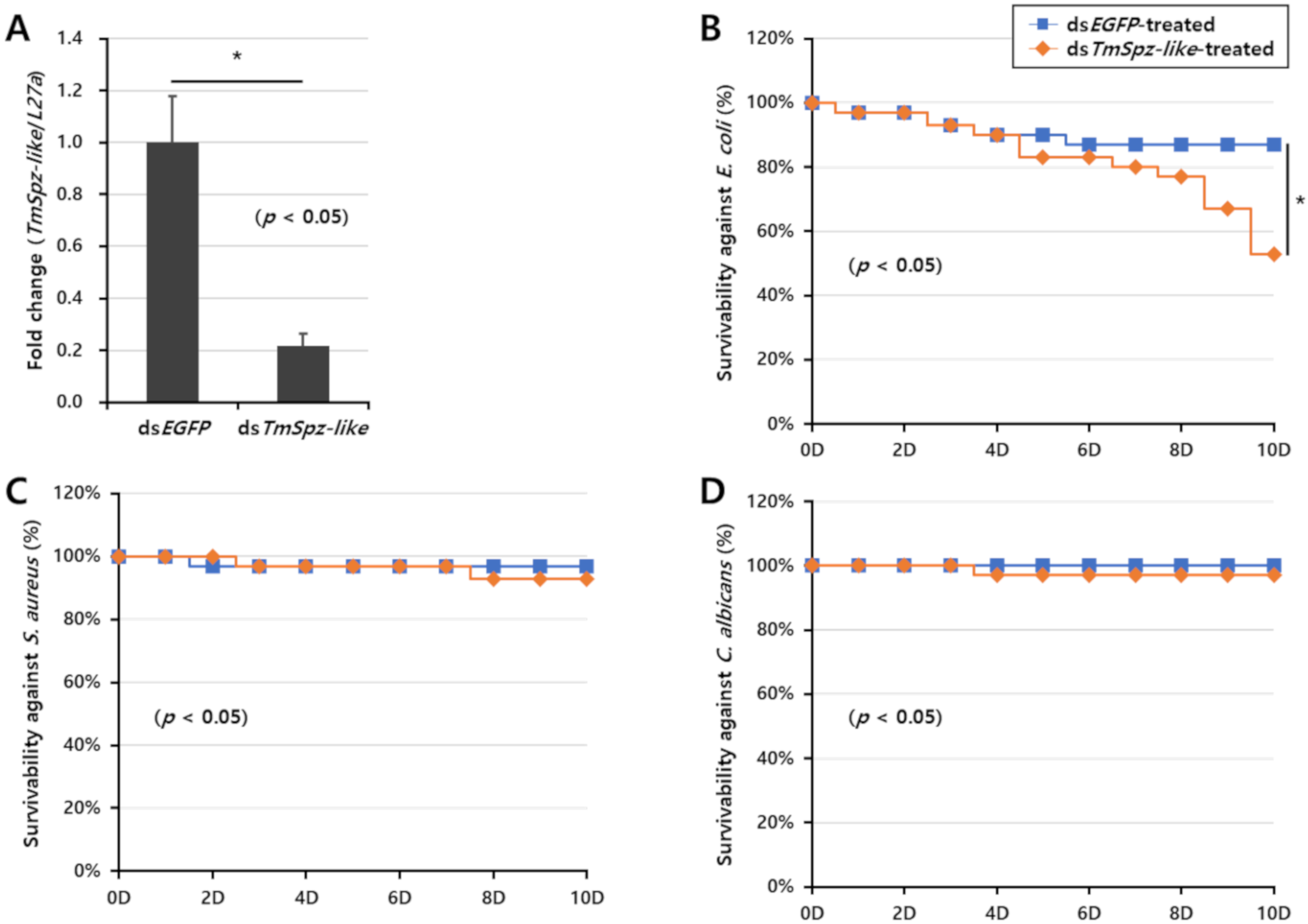
2.5. TmSpz-like Knockdown Increased the Mortality of E. coli-Infected T. Molitor Larvae
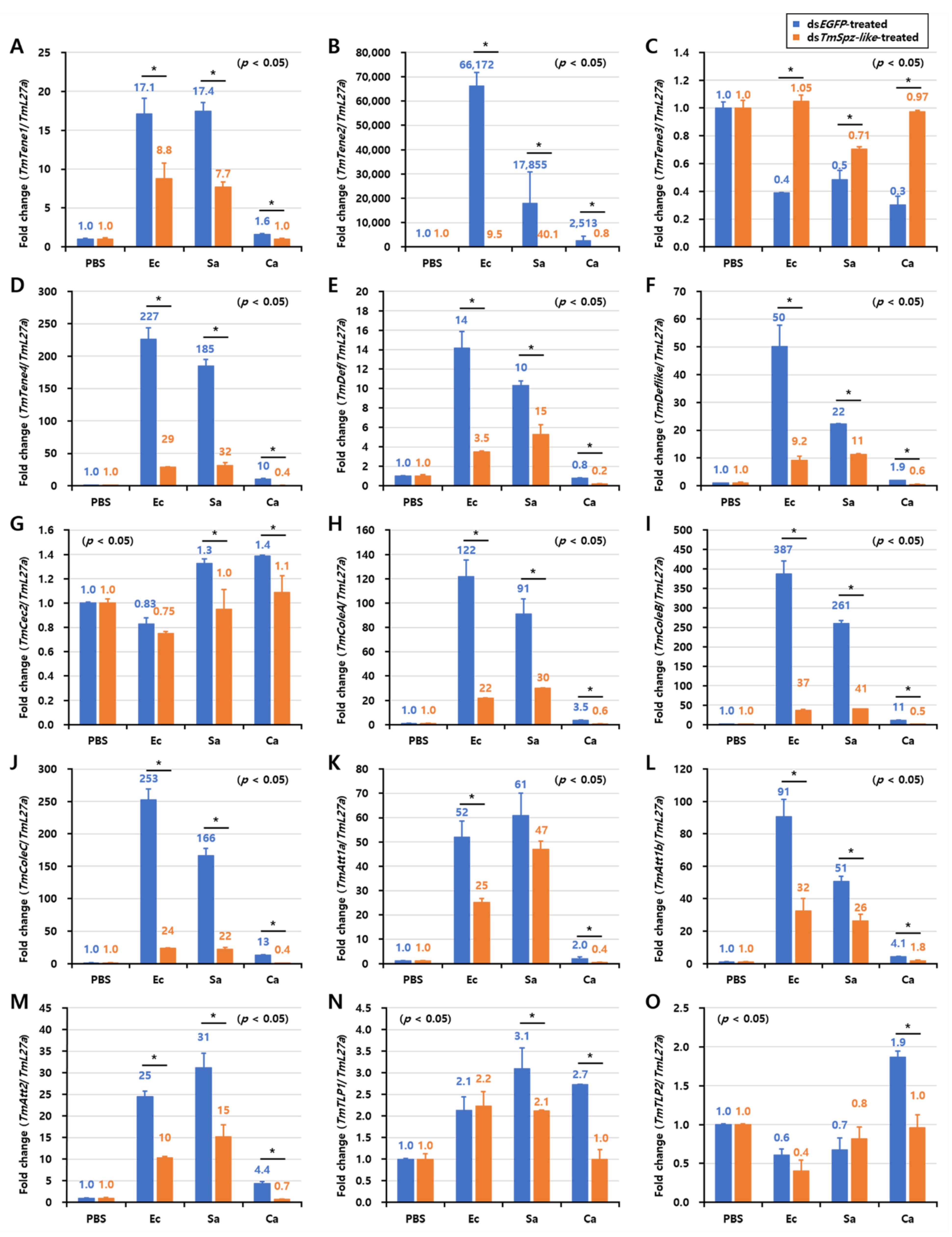
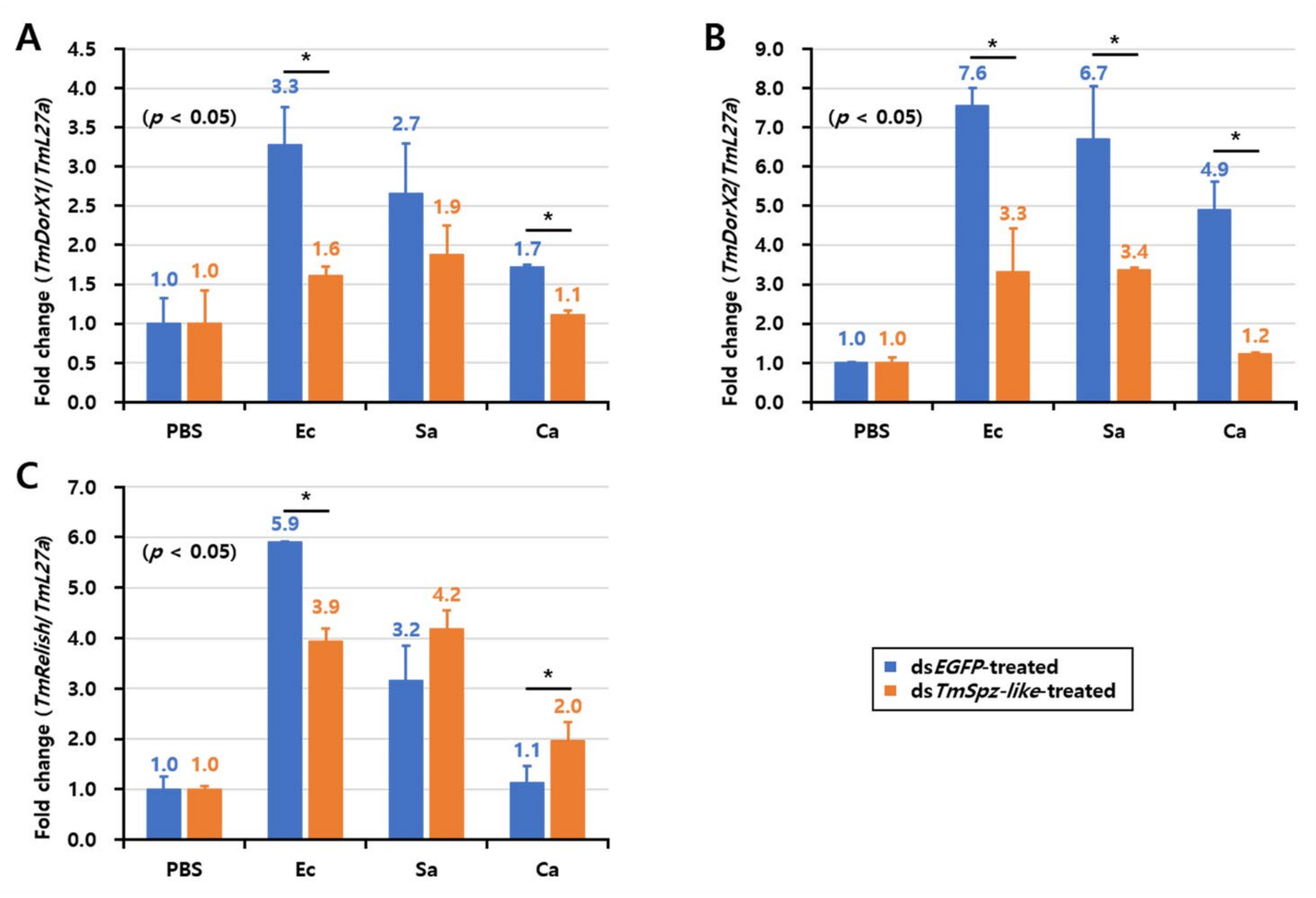
2.6. Effects of TmSpz-like Knockdown on the Expression of AMPs and NF-κB
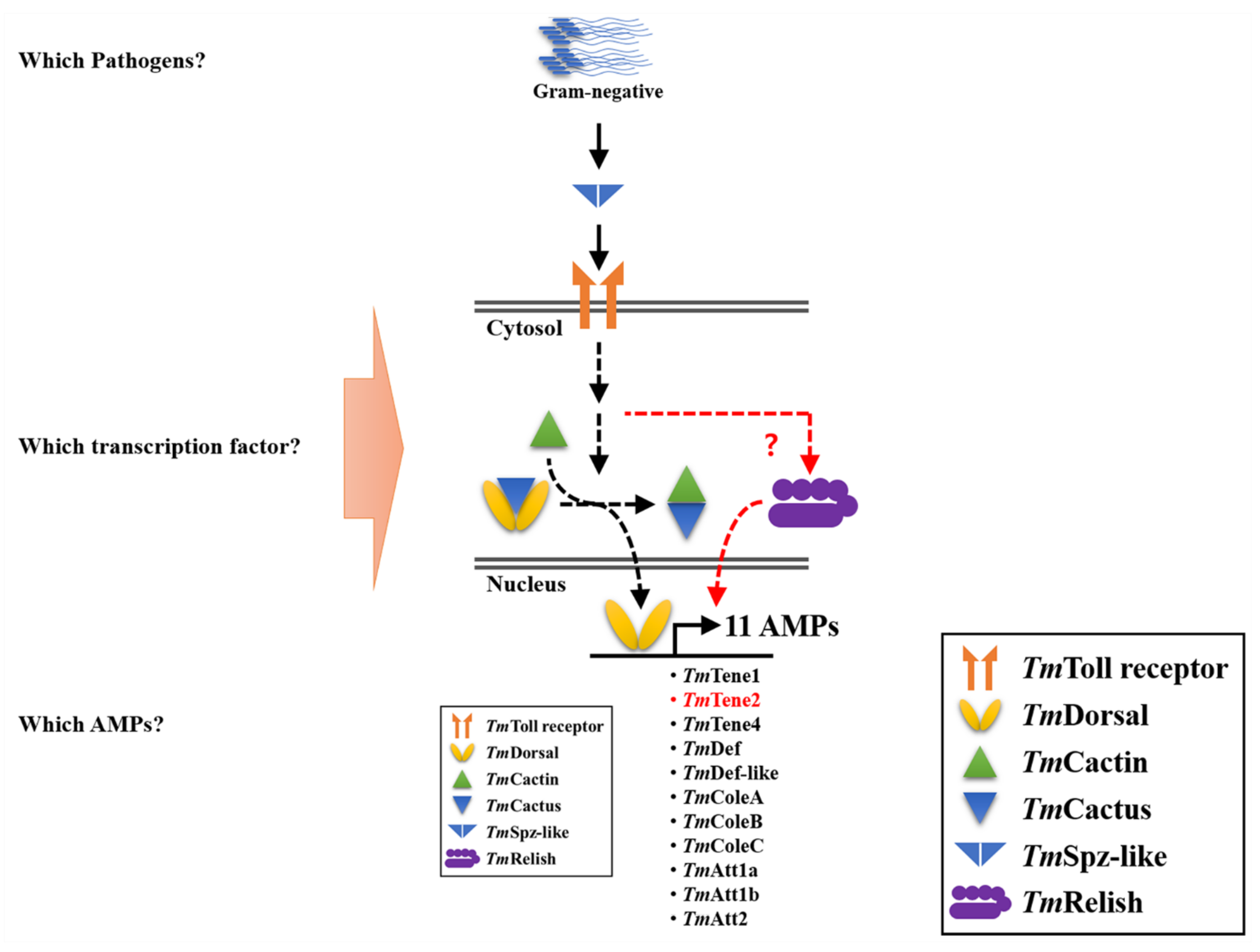
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Insect Rearing
5.2. Microorganisms
5.3. In Silico Identification and Cloning of the Full-Length TmSpz-like cDNA
5.4. Domain and Phylogenetic Analyses
5.5. Analysis of TmSpz-like Expression in Different Developmental Stages and Tissues
5.6. RNAi Analysis
5.7. Effect of TmSpz-like Knockdown on Transcriptional Regulation of AMP and NF-κB
5.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lemaitre, B.; Hoffmann, J. The host defense of drosophila melanogaster. Annu. Rev. Immunol. 2007, 25, 697–743. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.H.; Kim, S.J.; Kan, H.; Kwon, H.M.; Roh, K.B.; Jiang, R.; Yang, Y.; Park, J.W.; Lee, H.H.; Ha, N.C.; et al. A three-step proteolytic cascade mediates the activation of the peptidoglycan-induced toll pathway in an insect. J. Biol. Chem. 2008, 283, 7599–7607. [Google Scholar] [CrossRef] [PubMed]
- Roh, K.B.; Kim, C.H.; Lee, H.; Kwon, H.M.; Park, J.W.; Ryu, J.H.; Kurokawa, K.; Ha, N.C.; Lee, W.J.; Lemaitre, B.; et al. Proteolytic cascade for the activation of the insect toll pathway induced by the fungal cell wall component. J. Biol. Chem. 2009, 284, 19474–19481. [Google Scholar] [CrossRef] [PubMed]
- Keshavarz, M.; Jo, Y.H.; Edosa, T.T.; Bae, Y.M.; Han, Y.S. Tmpgrp-sa regulates antimicrobial response to bacteria and fungi in the fat body and gut of tenebrio molitor. Int. J. Mol. Sci. 2020, 21, 2113. [Google Scholar] [CrossRef] [PubMed]
- Edosa, T.T.; Jo, Y.H.; Keshavarz, M.; Kim, I.S.; Han, Y.S. Biosurfactants induce antimicrobial peptide production through the activation oftmspatzles intenebrio molitor. Int. J. Mol. Sci. 2020, 21, 6090. [Google Scholar] [CrossRef]
- Yu, B.; Sang, Q.; Pan, G.; Li, C.; Zhou, Z. A toll-spatzle pathway in the immune response of bombyx mori. Insects 2020, 11, 586. [Google Scholar] [CrossRef]
- Huang, X.; Xu, Y.; Zhang, Y.; Liu, J.; Luo, S.; Wu, J. Spatzle4 gene of silkworm, bombyx mori: Identification, immune response, and the effect of rna interference on the antimicrobial peptides’ expression in the integument. Saudi J. Biol. Sci. 2018, 25, 1817–1825. [Google Scholar] [CrossRef]
- Muhammad, A.; Habineza, P.; Wang, X.; Xiao, R.; Ji, T.; Hou, Y.; Shi, Z. Spatzle homolog-mediated toll-like pathway regulates innate immune responses to maintain the homeostasis of gut microbiota in the red palm weevil, rhynchophorus ferrugineus olivier (coleoptera: Dryophthoridae). Front. Microbiol. 2020, 11, 846. [Google Scholar] [CrossRef]
- Edosa, T.T.; Jo, Y.H.; Keshavarz, M.; Bae, Y.M.; Kim, D.H.; Lee, Y.S.; Han, Y.S. Tmspz4 plays an important role in regulating the production of antimicrobial peptides in response to escherichia coli and candida albicans infections. Int. J. Mol. Sci. 2020, 21, 1878. [Google Scholar] [CrossRef]
- Keshavarz, M.; Jo, Y.H.; Patnaik, B.B.; Park, K.B.; Ko, H.J.; Kim, C.E.; Edosa, T.T.; Lee, Y.S.; Han, Y.S. Tmrelish is required for regulating the antimicrobial responses to escherichia coli and staphylococcus aureus in tenebrio molitor. Sci. Rep. 2020, 10, 4258. [Google Scholar] [CrossRef]
- Edosa, T.T.; Jo, Y.H.; Keshavarz, M.; Bae, Y.M.; Kim, D.H.; Lee, Y.S.; Han, Y.S. Tmspz6 is essential for regulating the immune response to escherichia coli and staphylococcus aureus infection in tenebrio molitor. Insects 2020, 11, 105. [Google Scholar] [CrossRef]
- Nonaka, S.; Kawamura, K.; Hori, A.; Salim, E.; Fukushima, K.; Nakanishi, Y.; Kuraishi, T. Characterization of spz5 as a novel ligand for drosophila toll-1 receptor. Biochem. Biophys. Res. Commun. 2018, 506, 510–515. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, M.; Li, C.F.; He, Z.; Lu, Y.Z.; Liu, X.S.; Wang, Y.F.; Ip, Y.T.; Strand, M.R.; Yu, X.Q. Toll family members bind multiple spatzle proteins and activate antimicrobial peptide gene expression in drosophila. J. Biol. Chem. 2019, 294, 10172–10181. [Google Scholar] [CrossRef]
- Zhong, X.; Xu, X.X.; Yi, H.Y.; Lin, C.; Yu, X.Q. A toll-spatzle pathway in the tobacco hornworm, manduca sexta. Insect Biochem. Mol. Biol. 2012, 42, 514–524. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Jiang, Y.R.; Wang, Y.; Li, X.S.; Yang, R.S.; Yu, Z.G.; Qin, L. The toll signaling pathway in the chinese oak silkworm, antheraea pernyi: Innate immune responses to different microorganisms. PLoS ONE 2016, 11, e0160200. [Google Scholar] [CrossRef]
- Luna, C.; Wang, X.L.; Huang, Y.M.; Zhang, J.A.; Zheng, L.B. Characterization of four toll related genes during development and immune responses in anopheles gambiae. Insect Biochem. Mol. 2002, 32, 1171–1179. [Google Scholar] [CrossRef]
- Shin, S.W.; Bian, G.; Raikhel, A.S. A toll receptor and a cytokine, toll5a and spz1c, are involved in toll antifungal immune signaling in the mosquito aedes aegypti. J. Biol. Chem. 2006, 281, 39388–39395. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.H.; Liang, J.P.; Gu, Z.H.; Wan, D.H.; Weng, S.P.; Yu, X.Q.; He, J.G. Molecular cloning, characterization and expression analysis of two novel tolls (lvtoll2 and lvtoll3) and three putative spatzle-like toll ligands (lvspz1-3) from litopenaeus vannamei. Dev. Comp. Immunol. 2012, 36, 359–371. [Google Scholar] [CrossRef]
- Yuan, K.; Yuan, F.H.; Weng, S.P.; He, J.G.; Chen, Y.H. Identification and functional characterization of a novel spatzle gene in litopenaeus vannamei. Dev. Comp. Immunol. 2017, 68, 46–57. [Google Scholar] [CrossRef]
- Shi, X.Z.; Zhang, R.R.; Jia, Y.P.; Zhao, X.F.; Yu, X.Q.; Wang, J.X. Identification and molecular characterization of a spatzle-like protein from chinese shrimp (fenneropenaeus chinensis). Fish Shellf. Immunol. 2009, 27, 610–617. [Google Scholar] [CrossRef]
- Jo, Y.H.; Kim, Y.J.; Park, K.B.; Seong, J.H.; Kim, S.G.; Park, S.; Noh, M.Y.; Lee, Y.S.; Han, Y.S. Tmcactin plays an important role in gram-negative and -positive bacterial infection by regulating expression of 7 amp genes in tenebrio molitor. Sci. Rep. 2017, 7, 46459. [Google Scholar] [CrossRef]
- Jo, Y.H.; Patnaik, B.B.; Hwang, J.; Park, K.B.; Ko, H.J.; Kim, C.E.; Bae, Y.M.; Jung, W.J.; Lee, Y.S.; Han, Y.S. Regulation of the expression of nine antimicrobial peptide genes by tmimd confers resistance against gram-negative bacteria. Sci. Rep. 2019, 9, 10138. [Google Scholar] [CrossRef]
- Ko, H.J.; Jo, Y.H.; Patnaik, B.B.; Park, K.B.; Kim, C.E.; Keshavarz, M.; Jang, H.A.; Lee, Y.S.; Han, Y.S. Ikkgamma/nemo is required to confer antimicrobial innate immune responses in the yellow mealworm, tenebrio molitor. Int. J. Mol. Sci. 2020, 21, 6734. [Google Scholar] [CrossRef]
- Parker, J.S.; Mizuguchi, K.; Gay, N.J. A family of proteins related to spatzle, the toll receptor ligand, are encoded in the drosophila genome. Proteins 2001, 45, 71–80. [Google Scholar] [CrossRef]
- Valanne, S.; Wang, J.H.; Ramet, M. The drosophila toll signaling pathway. J. Immunol. 2011, 186, 649–656. [Google Scholar] [CrossRef] [PubMed]
- Parthier, C.; Stelter, M.; Ursel, C.; Fandrich, U.; Lilie, H.; Breithaupt, C.; Stubbs, M.T. Structure of the toll-spatzle complex, a molecular hub in drosophila development and innate immunity. Proc. Natl. Acad. Sci. USA 2014, 111, 6281–6286. [Google Scholar] [CrossRef]
- Moussian, B.; Roth, S. Dorsoventral axis formation in the drosophila embryo--shaping and transducing a morphogen gradient. Curr. Biol. 2005, 15, R887–R899. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Jo, Y.H.; Park, K.B.; Ko, H.J.; Kim, C.E.; Bae, Y.M.; Kim, B.; Jun, S.A.; Bang, I.S.; Lee, Y.S.; et al. Tmtoll-7 plays a crucial role in innate immune responses against gram-negative bacteria by regulating 5 amp genes in tenebrio molitor. Front. Immunol. 2019, 10, 310. [Google Scholar] [CrossRef]
- Borst, D.W.; Bollenbacher, W.E.; O’Connor, J.D.; King, D.S.; Fristrom, J.W. Ecdysone levels during metamorphosis of drosophila melanogaster. Dev. Biol. 1974, 39, 308–316. [Google Scholar] [CrossRef]
- Verma, P.; Tapadia, M.G. Immune response and anti-microbial peptides expression in malpighian tubules of drosophila melanogaster is under developmental regulation. PLoS ONE 2012, 7, e40714. [Google Scholar] [CrossRef]
- Shukla, A.; Tapadia, M.G. Differential localization and processing of apoptotic proteins in malpighian tubules of drosophila during metamorphosis. Eur. J. Cell. Biol. 2011, 90, 72–80. [Google Scholar] [CrossRef]
- Yu, Y.; Park, J.W.; Kwon, H.M.; Hwang, H.O.; Jang, I.H.; Masuda, A.; Kurokawa, K.; Nakayama, H.; Lee, W.J.; Dohmae, N.; et al. Diversity of innate immune recognition mechanism for bacterial polymeric meso-diaminopimelic acid-type peptidoglycan in insects. J. Biol. Chem. 2010, 285, 32937–32945. [Google Scholar] [CrossRef]
- Sun, J.J.; Xu, S.; He, Z.H.; Shi, X.Z.; Zhao, X.F.; Wang, J.X. Activation of toll pathway is different between kuruma shrimp and drosophila. Front. Immunol. 2017, 8, 1151. [Google Scholar] [CrossRef]
- An, C.J.; Jiang, H.B.; Kanost, M.R. Proteolytic activation and function of the cytokine spatzle in the innate immune response of a lepidopteran insect, manduca sexta. FEBS J. 2010, 277, 148–162. [Google Scholar] [CrossRef] [PubMed]
- Bulet, P.; Cociancich, S.; Dimarcq, J.L.; Lambert, J.; Reichhart, J.M.; Hoffmann, D.; Hetru, C.; Hoffmann, J.A. Insect immunity. Isolation from a coleopteran insect of a novel inducible antibacterial peptide and of new members of the insect defensin family. J. Biol. Chem. 1991, 266, 24520–24525. [Google Scholar] [CrossRef]
- Carlsson, A.; Nystrom, T.; de Cock, H.; Bennich, H. Attacin—An insect immune protein--binds lps and triggers the specific inhibition of bacterial outer-membrane protein synthesis. Microbiology 1998, 144 Pt 8, 2179–2188. [Google Scholar] [CrossRef] [PubMed]
- Bechinger, B.; Lohner, K. Detergent-like actions of linear amphipathic cationic antimicrobial peptides. Biochim. Biophys. Acta 2006, 1758, 1529–1539. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, L.; Xu, X.X.; Freed, S.F.; Gao, Y.F.; Yu, J.; Wang, S.; Ju, W.Y.; Zhang, Y.Q.; Jin, F.L. Cecropins from plutella xylostella and their interaction with metarhizium anisopliae. PLoS ONE 2015, 10, e0142451. [Google Scholar] [CrossRef]
- Ali Mohammadie Kojour, M.; Jang, H.A.; Edosa, T.T.; Keshavarz, M.; Kim, B.B.; Bae, Y.M.; Patnaik, B.B.; Han, Y.S.; Jo, Y.H. Identification, in silico characterization, and expression analysis of tenebrio molitor cecropin-2. Entomol. Res. 2021, 51, 74–82. [Google Scholar] [CrossRef]
- Jang, H.A.; Park, K.B.; Kim, B.B.; Ali Mohammadie Kojour, M.; Bae, Y.M.; Baliarsingh, S.; Lee, Y.S.; Han, Y.S.; Jo, Y.H. Bacterial but not fungal challenge up-regulates the transcription of coleoptericin genes in tenebrio molitor. Entomol. Res. 2020, 50, 440–449. [Google Scholar] [CrossRef]
- Iqbal, I.; Tripathi, R.K.; Wilkins, O.; Singh, J. Thaumatin-like protein (tlp) gene family in barley: Genome-wide exploration and expression analysis during germination. Genes 2020, 11, 1080. [Google Scholar] [CrossRef] [PubMed]
- Maistrou, S.; Paris, V.; Jensen, A.B.; Rolff, J.; Meyling, N.V.; Zanchi, C. A constitutively expressed antifungal peptide protects tenebrio molitor during a natural infection by the entomopathogenic fungus beauveria bassiana. Dev. Comp. Immunol. 2018, 86, 26–33. [Google Scholar] [CrossRef]
- Zhang, R.; Zhu, Y.; Pang, X.; Xiao, X.; Zhang, R.; Cheng, G. Regulation of antimicrobial peptides in aedes aegypti aag2 cells. Front. Cell Infect. Microbiol. 2017, 7, 22. [Google Scholar] [CrossRef] [PubMed]
- Keshavarz, M.; Jo, Y.H.; Park, K.B.; Ko, H.J.; Edosa, T.T.; Lee, Y.S.; Han, Y.S. Tmdorx2 positively regulates antimicrobial peptides in tenebrio molitor gut, fat body, and hemocytes in response to bacterial and fungal infection. Sci. Rep. 2019, 9, 16878. [Google Scholar] [CrossRef]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal w and clustal x version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. Mega7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative pcr and the 2(t)(-delta delta c) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Primer Sequences (5′-3′) |
|---|---|
| TmSpz-like-ORF-Fw | CGTTTTCAGCGGCTAATTGT |
| TmSpz-like-ORF-Rv | CATAATTCCCTTTTCCCAATTT |
| TmSpz-like-T7-Fw | TAATACGACTCACTATAGGGT ATGTTCCCAAAATCAACGGA |
| TmSpz-like-T7-Rv | TAATACGACTCACTATAGGGT AATCACACGCAGATCCTTCC |
| EGFP-T7-Fw | TAATACGACTCACTATAGGGT CGTAAACGGCCACAAGTTC |
| EGFP-T7-Rv | TAATACGACTCACTATAGGGT TGCTCAGGTAGTGTTGTCG |
| TmSpz-like-qPCR-Fw | CAGTTGAGGGTGCCTGTTCA |
| TmSpz-like-qPCR-Rv | TTGTTGGCATCGTCCCTTGA |
| TmL27a-qPCR-Fw | TCATCCTGAAGGCAAAGCTCCAGT |
| TmL27a-qPCR-Rv | AGGTTGGTTAGGCAGGCACCTTTA |
| TmTenecin-1-qPCR-Fw | CAGCTGAAGAAATCGAACAAGG |
| TmTenecin-1-qPCR-Rv | CAGACCCTCTTTCCGTTACAGT |
| TmTenecin-2-qPCR-Fw | CAGCAAAACGGAGGATGGTC |
| TmTenecin-2-qPCR-Rv | CGTTGAAATCGTGATCTTGTCC |
| TmTenecin-3-qPCR-Fw | GATTTGCTTGATTCTGGTGGTC |
| TmTenecin-3-qPCR-Rv | CTGATGGCCTCCTAAATGTCC |
| TmTenecin-4-qPCR-Fw | GGACATTGAAGATCCAGGAAAG |
| TmTenecin-4-qPCR-Rv | CGGTGTTCCTTATGTAGAGCTG |
| TmDefensin-Fw | AAATCGAACAAGGCCAACAC |
| TmDefensin-Rv | GCAAATGCAGACCCTCTTTC |
| TmDefensin-like-Fw | GCGATGCCTCATGAAGATGTAG |
| TmDefensin-like-Rv | CCAATGCAAACACATTCGTC |
| TmColeoptericin-A-qPCR-Fw | GGACAGAATGGTGGATGGTC |
| TmColeoptericin-A-qPCR-Rv | CTCCAACATTCCAGGTAGGC |
| TmColeoptericin-B-qPCR-Fw | CAGCTGTTGCCCACAAGTG |
| TmColeoptericin-B-qPCR-Rv | CTCAACGTTGGTCCTGGTGT |
| TmColeoptericin-C-qPCR-Fw | GGACGGTTCTGATCTTCTTGAT |
| TmColeoptericin-C-qPCR-Rv | CAGCTGTTTGTTTGTTCTCGTC |
| TmAttacin-1a-Fw | GAAACGAAATGGAAGGTGGA |
| TmAttacin-1a-Rv | TGCTTCGGCAGACAATACAG |
| TmAttacin-1b-Fw | GAGCTGTGAATGCAGGACAA |
| TmAttacin-1b-Rv | CCCTCTGATGAAACCTCCAA |
| TmAttacin-2-Fw | AACTGGGATATTCGCACGTC |
| TmAttacin-2-Rv | CCCTCCGAAATGTCTGTTGT |
| TmCecropin-2-Fw | TACTAGCAGCGCCAAAACCT |
| TmCecropin-2-Rv | CTGGAACATTAGGCGGAGAA |
| TmThaumatin-like protein-1-Fw | CTCAAAGGACACGCAGGACT |
| TmThaumatin-like protein-1-Rv | ACTTTGAGCTTCTCGGGACA |
| TmThaumatin-like protein-2-Fw | CCGTCTGGCTAGGAGTTCTG |
| TmThaumatin-like protein-2-Rv | ACTCCTCCAGCTCCGTTACA |
| TmDorX1_qPCR_Fw | AGCGTTGAGGTTTCGGTATG |
| TmDorX1_qPCR_Rv | TCTTTGGTGACGCAAGACAC |
| TmDorX2_qPCR_Fw | ACACCCCCGAAATCACAAAC |
| TmDorX2_qPCR_Rv | TTTCAGAGCGCCAGGTTTTG |
| TmRelish_qPCR_Fw | AGCGTCAAGTTGGAGCAGAT |
| TmRelish_qPCR_Rv | GTCCGGACCTCAAGTGT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jang, H.A.; Patnaik, B.B.; Ali Mohammadie Kojour, M.; Kim, B.B.; Bae, Y.M.; Park, K.B.; Lee, Y.S.; Jo, Y.H.; Han, Y.S. TmSpz-like Plays a Fundamental Role in Response to E. coli but Not S. aureus or C. albican Infection in Tenebrio molitor via Regulation of Antimicrobial Peptide Production. Int. J. Mol. Sci. 2021, 22, 10888. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms221910888
Jang HA, Patnaik BB, Ali Mohammadie Kojour M, Kim BB, Bae YM, Park KB, Lee YS, Jo YH, Han YS. TmSpz-like Plays a Fundamental Role in Response to E. coli but Not S. aureus or C. albican Infection in Tenebrio molitor via Regulation of Antimicrobial Peptide Production. International Journal of Molecular Sciences. 2021; 22(19):10888. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms221910888
Chicago/Turabian StyleJang, Ho Am, Bharat Bhusan Patnaik, Maryam Ali Mohammadie Kojour, Bo Bae Kim, Young Min Bae, Ki Beom Park, Yong Seok Lee, Yong Hun Jo, and Yeon Soo Han. 2021. "TmSpz-like Plays a Fundamental Role in Response to E. coli but Not S. aureus or C. albican Infection in Tenebrio molitor via Regulation of Antimicrobial Peptide Production" International Journal of Molecular Sciences 22, no. 19: 10888. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms221910888