
Characterizing Binding Interactions That Are Essential for Selective Transport through the Nuclear Pore Complex

 , , , and
, , , and
Abstract
:1. Introduction
2. Results
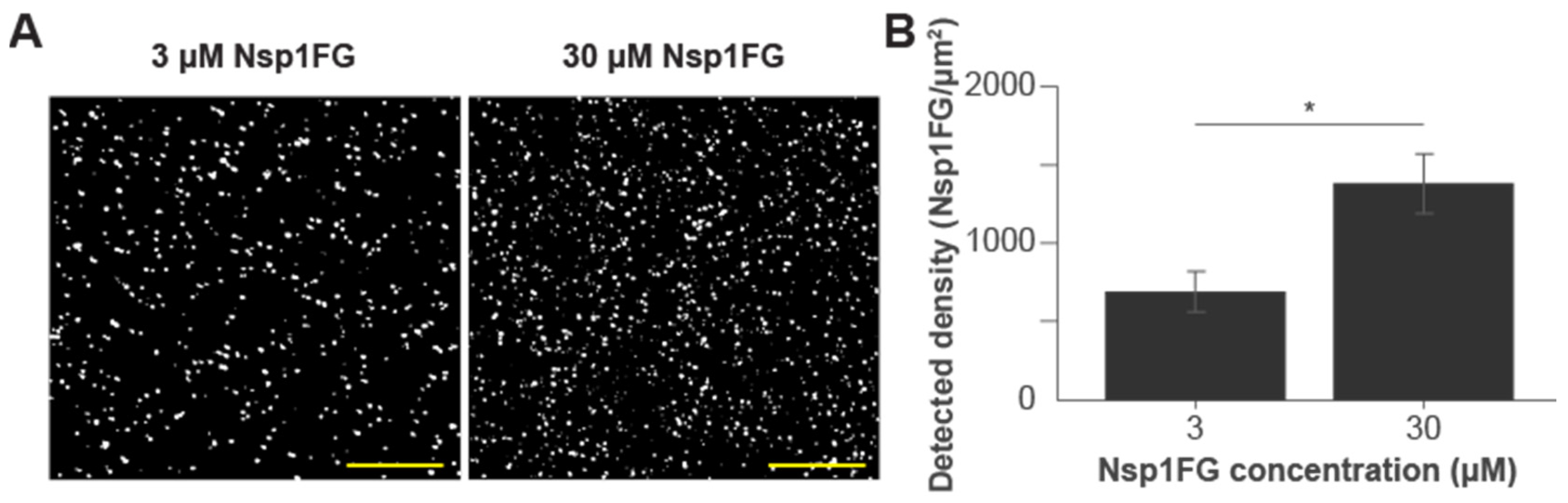
2.1. Characterizing the Stationary Phase of the NPC Barrier Mimic
2.2. Molecular Dynamic Simulations to Determine Radii of Gyration
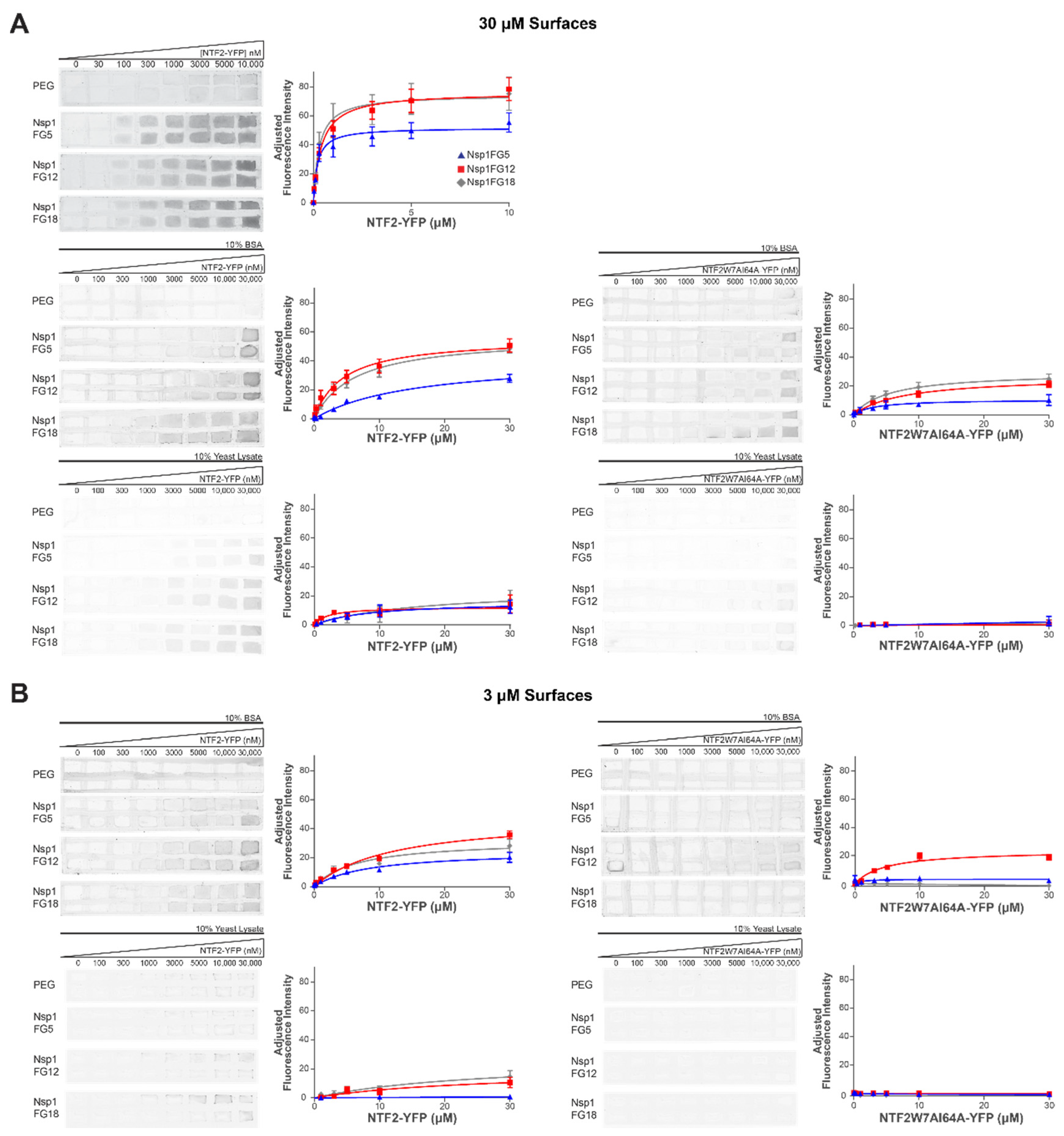
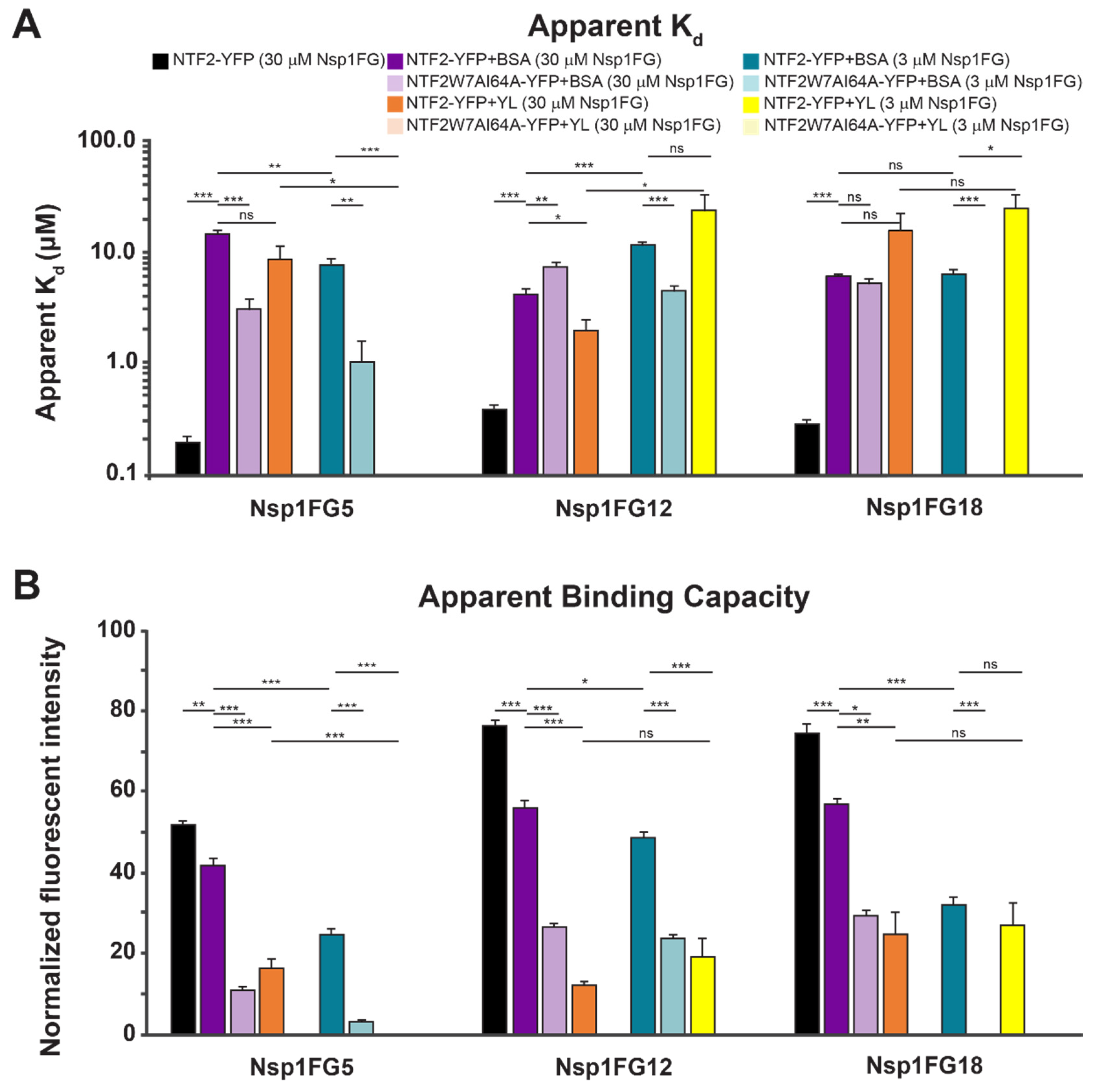
2.3. Binding of NTF2-YFP onto Nsp1FG Monolayers
2.4. Agent-Based Modeling Suggests Multivalent NTF2-Nsp1FG Interactions
3. Discussion
4. Materials and Methods
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Alberts, B. Molecular Biology of the Cell; Garland Publishing: New York, NY, USA, 2014. [Google Scholar]
- Alber, F.; Dokudovskaya, S.; Veenhoff, L.M.; Zhang, W.Z.; Kipper, J.; Devos, D.; Suprapto, A.; Karni-Schmidt, O.; Williams, R.; Chait, B.T.; et al. The molecular architecture of the nuclear pore complex. Nature 2007, 450, 695–701. [Google Scholar] [CrossRef]
- Cautain, B.; Hill, R.; de Pedro, N.; Link, W. Components and regulation of nuclear transport processes. FEBS J. 2015, 282, 445–462. [Google Scholar] [CrossRef]
- Lim, R.Y.H.; Aebi, U.; Fahrenkrog, B. Towards reconciling structure and function in the nuclear pore complex. Histochem. Cell Biol. 2008, 129, 105–116. [Google Scholar] [CrossRef] [Green Version]
- Cronshaw, J.A.; Krutchinsky, A.N.; Zhang, W.Z.; Chait, B.T.; Matunis, M.J. Proteomic analysis of the mammalian nuclear pore complex. J. Cell Biol. 2002, 158, 915–927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alber, F.; Dokudovskaya, S.; Veenhoff, L.M.; Zhang, W.Z.; Kipper, J.; Devos, D.; Suprapto, A.; Karni-Schmidt, O.; Williams, R.; Chait, B.T.; et al. Determining the architectures of macromolecular assemblies. Nature 2007, 450, 683–694. [Google Scholar] [CrossRef] [PubMed]
- Rout, M.P.; Aitchison, J.D.; Magnasco, M.O.; Chait, B.T. Virtual gating and nuclear transport: The hole picture. Trends Cell Biol. 2003, 13, 622–628. [Google Scholar] [CrossRef] [PubMed]
- Zilman, A.; Di Talia, S.; Chait, B.T.; Rout, M.P.; Magnasco, M.O. Efficiency, selectivity, and robustness of nucleocytoplasmic transport. PLoS Comput. Biol. 2007, 3, 1281–1290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, R.Y.H.; Huang, N.P.; Koser, J.; Deng, J.; Lau, K.H.A.; Schwarz-Herion, K.; Fahrenkrog, B.; Aebi, U. Flexible phenylalanine-glycine nucleoporins as entropic barriers to nucleocytoplasmic transport. Proc. Natl. Acad. Sci. USA 2006, 103, 9512–9517. [Google Scholar] [CrossRef] [Green Version]
- Ghavami, A.; Veenhoff, L.M.; van der Giessen, E.; Onck, P.R. Probing the disordered domain of the nuclear pore complex through coarse-grained molecular dynamics simulations. Biophys. J. 2014, 107, 1393–1402. [Google Scholar] [CrossRef] [Green Version]
- Dange, T.; Grunwald, D.; Grunwald, A.; Peters, R.; Kubitscheck, U. Autonomy and robustness of translocation through the nuclear pore complex: A single-molecule study. J. Cell Biol. 2008, 183, 77–86. [Google Scholar] [CrossRef] [Green Version]
- Kubitscheck, U.; Grunwald, D.; Hoekstra, A.; Rohleder, D.; Kues, T.; Siebrasse, J.P.; Peters, R. Nuclear transport of single molecules: Dwell times at the nuclear pore complex. J. Cell Biol. 2005, 168, 233–243. [Google Scholar] [CrossRef]
- Yang, W.; Gelles, J.; Musser, S. Imaging of single-molecule translocation through nuclear pore complexes. Proc. Natl. Acad. Sci. USA 2004, 101, 12887–12892. [Google Scholar] [CrossRef] [Green Version]
- Ribbeck, K.; Gorlich, D. Kinetic analysis of translocation through nuclear pore complexes. EMBO J. 2001, 20, 1320–1330. [Google Scholar] [CrossRef] [PubMed]
- Milles, S.; Mercadante, D.; Aramburu, I.V.; Jensen, M.R.; Banterle, N.; Koehler, C.; Tyagi, S.; Clarke, J.; Shammas, S.L.; Blackledge, M.; et al. Plasticity of an Ultrafast Interaction between Nucleoporins and Nuclear Transport Receptors. Cell 2015, 163, 734–745. [Google Scholar] [CrossRef] [Green Version]
- Hough, L.E.; Dutta, K.; Sparks, S.; Temel, D.B.; Kamal, A.; Tetenbaum-Novatt, J.; Rout, M.P.; Cowburn, D. The molecular mechanism of nuclear transport revealed by atomic-scale measurements. eLife 2015, 4. [Google Scholar] [CrossRef]
- Moussavi-Baygi, R.; Mofrad, M.R. Rapid Brownian Motion Primes Ultrafast Reconstruction of Intrinsically Disordered Phe-Gly Repeats Inside the Nuclear Pore Complex. Sci. Rep. 2016, 6, 29991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakiyama, Y.; Mazur, A.; Kapinos, L.E.; Lim, R.Y. Spatiotemporal dynamics of the nuclear pore complex transport barrier resolved by high-speed atomic force microscopy. Nat. Nanotechnol. 2016, 11, 719–723. [Google Scholar] [CrossRef]
- Hayama, R.; Sparks, S.; Hecht, L.M.; Dutta, K.; Karp, J.M.; Cabana, C.M.; Rout, M.P.; Cowburn, D. Thermodynamic characterization of the multivalent interactions underlying rapid and selective translocation through the nuclear pore complex. J. Biol. Chem. 2018, 293, 4555–4563. [Google Scholar] [CrossRef] [Green Version]
- Yamada, J.; Phillips, J.L.; Patel, S.; Goldfien, G.; Calestagne-Morelli, A.; Huang, H.; Reza, R.; Acheson, J.; Krishnan, V.V.; Newsam, S.; et al. A Bimodal Distribution of Two Distinct Categories of Intrinsically Disordered Structures with Separate Functions in FG Nucleoporins. Mol. Cell. Proteom. 2010, 9, 2205–2224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ando, D.; Zandi, R.; Kim, Y.W.; Colvin, M.; Rexach, M.; Gopinathan, A. Nuclear pore complex protein sequences determine overall copolymer brush structure and function. Biophys. J. 2014, 106, 1997–2007. [Google Scholar] [CrossRef] [Green Version]
- Tagliazucchi, M.; Peleg, O.; Kroger, M.; Rabin, Y.; Szleifer, I. Effect of charge, hydrophobicity, and sequence of nucleoporins on the translocation of model particles through the nuclear pore complex. Proc. Natl. Acad. Sci. USA 2013, 110, 3363–3368. [Google Scholar] [CrossRef] [Green Version]
- Xylourgidis, N.; Fornerod, M. Acting out of character: Regulatory roles of nuclear pore complex proteins. Dev. Cell 2009, 17, 617–625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakuma, S.; D’Angelo, M.A. The roles of the nuclear pore complex in cellular dysfunction, aging and disease. Semin. Cell Dev. Biol. 2017, 68, 72–84. [Google Scholar] [CrossRef] [PubMed]
- Jamali, T.; Jamali, Y.; Mehrbod, M.; Mofrad, M.R.K. Nuclear pore complex: Biochemistry and biophysics of nucleocytoplasmic transport in health and disease. In International Review of Cell and Molecular Biology; Jeon, K.W., Ed.; Elsevier Academic Press Inc.: San Diego, CA, USA, 2011; Volume 287, pp. 233–286. [Google Scholar]
- Ding, B.; Sepehrimanesh, M. Nucleocytoplasmic Transport: Regulatory Mechanisms and the Implications in Neurodegeneration. Int. J. Mol. Sci. 2021, 22, 4165. [Google Scholar] [CrossRef] [PubMed]
- Bitetto, G.; Di Fonzo, A. Nucleo–cytoplasmic transport defects and protein aggregates in neurodegeneration. Transl. Neurodegener. 2020, 9, 25. [Google Scholar] [CrossRef] [PubMed]
- Basel-Vanagaite, L.; Muncher, L.; Straussberg, R.; Pasmanik-Chor, M.; Yahav, M.; Rainshtein, L.; Walsh, C.A.; Magal, N.; Taub, E.; Drasinover, V.; et al. Mutated nup62 causes autosomal recessive infantile bilateral striatal necrosis. Ann. Neurol. 2006, 60, 214–222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, I.S.; Kim, D.H.; Han, S.M.; Chin, M.U.; Nam, H.J.; Cho, H.P.; Choi, S.Y.; Song, B.J.; Kim, E.R.; Bae, Y.S.; et al. Truncated form of importin alpha identified in breast cancer cell inhibits nuclear import of p53. J. Biol. Chem. 2000, 275, 23139–23145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terlecki-Zaniewicz, S.; Humer, T.; Eder, T.; Schmoellerl, J.; Heyes, E.; Manhart, G.; Kuchynka, N.; Parapatics, K.; Liberante, F.G.; Müller, A.C.; et al. Biomolecular condensation of NUP98 fusion proteins drives leukemogenic gene expression. Nat. Struct. Mol. Biol. 2021, 28, 190–201. [Google Scholar] [CrossRef]
- Martinez, N.; Alonso, A.; Moragues, M.D.; Ponton, J.; Schneider, J. The nuclear pore complex protein Nup88 is overexpressed in tumor cells. Cancer Res. 1999, 59, 5408–5411. [Google Scholar]
- Grima, J.C.; Daigle, J.G.; Arbez, N.; Cunningham, K.C.; Zhang, K.; Ochaba, J.; Geater, C.; Morozko, E.; Stocksdale, J.; Glatzer, J.C.; et al. Mutant Huntingtin Disrupts the Nuclear Pore Complex. Neuron 2017, 94, 93–107.e106. [Google Scholar] [CrossRef] [Green Version]
- Tetenbaum-Novatt, J.; Hough, L.E.; Mironska, R.; McKenney, A.S.; Rout, M.P. Nucleocytoplasmic transport: A role for non-specific competition in karyopherin-nucleoporin interactions. Mol. Cell Proteom. 2012, 11, 31–46. [Google Scholar] [CrossRef] [Green Version]
- Lowe, A.R.; Tang, J.H.; Yassif, J.; Graf, M.; Huang, W.Y.; Groves, J.T.; Weis, K.; Liphardt, J.T. Importin-beta modulates the permeability of the nuclear pore complex in a Ran-dependent manner. eLife 2015, 4, e04052. [Google Scholar] [CrossRef] [PubMed]
- Hoogenboom, B.W.; Hough, L.E.; Lemke, E.A.; Lim, R.Y.H.; Onck, P.R.; Zilman, A. Physics of the nuclear pore complex: Theory, modeling and experiment. Phys. Rep. 2021, 921, 1–53. [Google Scholar] [CrossRef]
- Zilman, A.; Di Talia, S.; Jovanovic-Talisman, T.; Chait, B.; Rout, M.; Magnasco, M.O. Enhancement of transport selectivity through nano-channels by non-specific competition. PLoS Comp. Biol. 2010, 6, e1000804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jovanovic-Talisman, T.; Zilman, A. Protein Transport by the Nuclear Pore Complex: Simple Biophysics of a Complex Biomachine. Biophys. J. 2017, 113, 6–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jovanovic-Talisman, T.; Tetenbaum-Novatt, J.; McKenney, A.S.; Zilman, A.; Peters, R.; Rout, M.P.; Chait, B.T. Artificial nanopores that mimic the transport selectivity of the nuclear pore complex. Nature 2009, 457, 1023–1027. [Google Scholar] [CrossRef]
- Kowalczyk, S.W.; Kapinos, L.; Blosser, T.R.; Magalhaes, T.; van Nies, P.; Lim, R.Y.H.; Dekker, C. Single-molecule transport across an individual biomimetic nuclear pore complex. Nat. Nanotechnol. 2011, 6, 433–438. [Google Scholar] [CrossRef]
- Fisher, P.D.E.; Shen, Q.; Akpinar, B.; Davis, L.K.; Chung, K.K.H.; Baddeley, D.; Saric, A.; Melia, T.J.; Hoogenboom, B.W.; Lin, C.; et al. A Programmable DNA Origami Platform for Organizing Intrinsically Disordered Nucleoporins within Nanopore Confinement. ACS Nano 2018, 12, 1508–1518. [Google Scholar] [CrossRef]
- Celetti, G.; Paci, G.; Caria, J.; VanDelinder, V.; Bachand, G.; Lemke, E.A. The liquid state of FG-nucleoporins mimics permeability barrier properties of nuclear pore complexes. J. Cell Biol. 2019, 219, e201907157. [Google Scholar] [CrossRef]
- Wolf, C.; Mofrad, M.R. On the octagonal structure of the nuclear pore complex: Insights from coarse-grained models. Biophys. J. 2008, 95, 2073–2085. [Google Scholar] [CrossRef] [Green Version]
- Azimi, M.; Jamali, Y.; Mofrad, M.R. Accounting for diffusion in agent based models of reaction-diffusion systems with application to cytoskeletal diffusion. PLoS ONE 2011, 6, e25306. [Google Scholar] [CrossRef] [PubMed]
- Moussavi-Baygi, R.; Jamali, Y.; Karimi, R.; Mofrad, M.R. Biophysical coarse-grained modeling provides insights into transport through the nuclear pore complex. Biophys. J. 2011, 100, 1410–1419. [Google Scholar] [CrossRef] [Green Version]
- Peyro, M.; Dickson, A.M.; Mofrad, M.R.K. Nucleoporins’ exclusive amino acid sequence features regulate their transient interaction with and selectivity of cargo complexes in the nuclear pore. Mol. Biol. Cell 2021, in press. [Google Scholar] [CrossRef] [PubMed]
- Kapinos, L.E.; Schoch, R.L.; Wagner, R.S.; Schleicher, K.D.; Lim, R.Y.H. Karyopherin-centric control of nuclear pores based on molecular occupancy and kinetic analysis of multivalent binding with FG nucleoporins. Biophys. J. 2014, 106, 1751–1762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zahn, R.; Osmanović, D.; Ehret, S.; Callis, C.A.; Frey, S.; Stewart, M.; You, C.; Görlich, D.; Hoogenboom, B.W.; Richter, R.P. A physical model describing the interaction of nuclear transport receptors with FG nucleoporin domain assemblies. eLife 2016, 5, e14119. [Google Scholar] [CrossRef] [PubMed]
- Wagner, R.S.; Kapinos, L.E.; Marshall, N.J.; Stewart, M.; Lim, R.Y. Promiscuous binding of Karyopherinbeta1 modulates FG nucleoporin barrier function and expedites NTF2 transport kinetics. Biophys. J. 2015, 108, 918–927. [Google Scholar] [CrossRef] [Green Version]
- Vovk, A.; Gu, C.; Opferman, M.G.; Kapinos, L.E.; Lim, R.Y.; Coalson, R.D.; Jasnow, D.; Zilman, A. Simple biophysics underpins collective conformations of the intrinsically disordered proteins of the Nuclear Pore Complex. eLife 2016, 5, e10785. [Google Scholar] [CrossRef]
- Eisele, N.B.; Frey, S.; Piehler, J.; Gorlich, D.; Richter, R.P. Ultrathin nucleoporin phenylalanine-glycine repeat films and their interaction with nuclear transport receptors. EMBO Rep. 2010, 11, 366–372. [Google Scholar] [CrossRef]
- Hayama, R.; Sorci, M.; Keating Iv, J.J.; Hecht, L.M.; Plawsky, J.L.; Belfort, G.; Chait, B.T.; Rout, M.P. Interactions of nuclear transport factors and surface-conjugated FG nucleoporins: Insights and limitations. PLoS ONE 2019, 14, e0217897. [Google Scholar] [CrossRef] [Green Version]
- Schoch, R.L.; Kapinos, L.E.; Lim, R.Y.H. Nuclear transport receptor binding avidity triggers a self-healing collapse transition in FG-nucleoporin molecular brushes. Proc. Natl. Acad. Sci. USA 2012, 109, 16911–16916. [Google Scholar] [CrossRef] [Green Version]
- Davis, L.K.; Šarić, A.; Hoogenboom, B.W.; Zilman, A. Physical modeling of multivalent interactions in the nuclear pore complex. Biophys. J. 2021, 120, 1565–1577. [Google Scholar] [CrossRef] [PubMed]
- Hurt, E.C. A novel nucleoskeletal-like protein located at the nuclear periphery is required for the life cycle of Saccharomyces cerevisiae. EMBO J. 1988, 7, 4323–4334. [Google Scholar] [CrossRef] [PubMed]
- Davis, L.K.; Ford, I.J.; Šarić, A.; Hoogenboom, B.W. Intrinsically disordered nuclear pore proteins show ideal-polymer morphologies and dynamics. Phys. Rev. E 2020, 101, 022420. [Google Scholar] [CrossRef] [PubMed]
- Gamini, R.; Han, W.; Stone, J.E.; Schulten, K. Assembly of Nsp1 nucleoporins provides insight into nuclear pore complex gating. PLoS Comput. Biol. 2014, 10, e1003488. [Google Scholar] [CrossRef]
- Ribbeck, K.; Lipowsky, G.; Kent, H.M.; Stewart, M.; Gorlich, D. NTF2 mediates nuclear import of Ran. EMBO J. 1998, 17, 6587–6598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clarkson, W.D.; Kent, H.M.; Stewart, M. Separate binding sites on nuclear transport factor 2 (NTF2) for GDP-Ran and the phenylalanine-rich repeat regions of nucleoporins p62 and Nsp1p. J. Mol. Biol. 1996, 263, 517–524. [Google Scholar] [CrossRef]
- Cushman, I.; Bowman, B.R.; Sowa, M.E.; Lichtarge, O.; Quiocho, F.A.; Moore, M.S. Computational and biochemical identification of a nuclear pore complex binding site on the nuclear transport carrier NTF2. J. Mol. Biol. 2004, 344, 303–310. [Google Scholar] [CrossRef]
- Siebrasse, J.P.; Peters, R. Rapid translocation of NTF2 through the nuclear pore of isolated nuclei and nuclear envelopes. EMBO Rep. 2002, 3, 887–892. [Google Scholar] [CrossRef] [Green Version]
- Knockenhauer, K.E.; Schwartz, T.U. The Nuclear Pore Complex as a Flexible and Dynamic Gate. Cell 2016, 164, 1162–1171. [Google Scholar] [CrossRef] [Green Version]
- Schleicher, K.D.; Dettmer, S.L.; Kapinos, L.E.; Pagliara, S.; Keyser, U.F.; Jeney, S.; Lim, R.Y. Selective transport control on molecular velcro made from intrinsically disordered proteins. Nat. Nanotechnol. 2014, 9, 525–530. [Google Scholar] [CrossRef]
- Bayliss, R.; Littlewood, T.; Stewart, M. Structural basis for the interaction between FxFG nucleoporin repeats and importin-beta in nuclear trafficking. Cell 2000, 102, 99–108. [Google Scholar] [CrossRef] [Green Version]
- Golfetto, O.; Wakefield, D.L.; Cacao, E.E.; Avery, K.N.; Kenyon, V.; Jorand, R.; Tobin, S.J.; Biswas, S.; Gutierrez, J.; Clinton, R.; et al. A Platform To Enhance Quantitative Single Molecule Localization Microscopy. J. Am. Chem. Soc. 2018, 140, 12785–12797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moran, U.; Phillips, R.; Milo, R. SnapShot: Key numbers in biology. Cell 2010, 141, 1262–1262.e1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peters, R. Translocation through the nuclear pore: Kaps pave the way. Bioessays 2009, 31, 466–477. [Google Scholar] [CrossRef]
- Pyhtila, B.; Rexach, M. A gradient of affinity for the karyopherin Kap95p along the yeast nuclear pore complex. J. Biol. Chem. 2003, 278, 42699–42709. [Google Scholar] [CrossRef] [Green Version]
- Jorand, R.; Biswas, S.; Wakefield, D.L.; Tobin, S.J.; Golfetto, O.; Hilton, K.; Ko, M.; Ramos, J.W.; Small, A.R.; Chu, P.; et al. Molecular signatures of mu opioid receptor and somatostatin receptor 2 in pancreatic cancer. Mol. Biol. Cell 2016, 27, 3659–3672. [Google Scholar] [CrossRef] [Green Version]
- Ghavami, A.; van der Giessen, E.; Onck, P.R. Coarse-Grained Potentials for Local Interactions in Unfolded Proteins. J. Chem. Theory Comput. 2013, 9, 432–440. [Google Scholar] [CrossRef]
- Pronk, S.; Páll, S.; Schulz, R.; Larsson, P.; Bjelkmar, P.; Apostolov, R.; Shirts, M.R.; Smith, J.C.; Kasson, P.M.; van der Spoel, D.; et al. GROMACS 4.5: A high-throughput and highly parallel open source molecular simulation toolkit. Bioinformatics 2013, 29, 845–854. [Google Scholar] [CrossRef]
- Humphrey, W.; Dalke, A.; Schulten, K. VMD: Visual molecular dynamics. J. Mol. Graph. 1996, 14, 33–38. [Google Scholar] [CrossRef]
- Soheilypour, M.; Mofrad, M.R.K. Agent-Based Modeling in Molecular Systems Biology. Bioessays 2018, 40, e1800020. [Google Scholar] [CrossRef]
- Soheilypour, M.; Mofrad, M.R. Regulation of RNA-binding proteins affinity to export receptors enables the nuclear basket proteins to distinguish and retain aberrant mRNAs. Sci. Rep. 2016, 6, 35380. [Google Scholar] [CrossRef] [PubMed]
- Azimi, M.; Mofrad, M.R. Higher nucleoporin-Importinbeta affinity at the nuclear basket increases nucleocytoplasmic import. PLoS ONE 2013, 8, e81741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azimi, M.; Bulat, E.; Weis, K.; Mofrad, M.R. An agent-based model for mRNA export through the nuclear pore complex. Mol. Biol. Cell 2014, 25, 3643–3653. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Nsp1FG Variant | Radius of Gyration (nm) |
|---|---|
| Nsp1FG5 | 4.3 |
| Nsp1FG12 | 6.7 |
| Input Kd1 | Input Kd2 | Input Kd3 | Apparent Kd | Apparent Bmax | |
|---|---|---|---|---|---|
| Nsp1FG5 + BSA 3 μM surfaces | 5.4 | 16.2 | -- | 8.16 ± 0.26 | 24.15 ± 0.19 |
| Nsp1FG5 + BSA 30 μM surfaces | 10.2 | 38.25 | -- | 15.16 ± 1.03 | 45.63 ± 1.90 |
| Nsp1FG12 + BSA 3 μM surfaces | 5.45 | 5.45 | 40 | 11.87 ± 0.35 | 50.59 ± 2.66 |
| Nsp1FG12 + BSA 30 μM surfaces | 2.2 | 4.89 | -- | 4.35 ± 0.24 | 58.23 ± 3.81 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lennon, K.M.; Soheilypour, M.; Peyro, M.; Wakefield, D.L.; Choo, G.E.; Mofrad, M.R.K.; Jovanovic-Talisman, T. Characterizing Binding Interactions That Are Essential for Selective Transport through the Nuclear Pore Complex. Int. J. Mol. Sci. 2021, 22, 10898. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms221910898
Lennon KM, Soheilypour M, Peyro M, Wakefield DL, Choo GE, Mofrad MRK, Jovanovic-Talisman T. Characterizing Binding Interactions That Are Essential for Selective Transport through the Nuclear Pore Complex. International Journal of Molecular Sciences. 2021; 22(19):10898. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms221910898
Chicago/Turabian StyleLennon, Kathleen M., Mohammad Soheilypour, Mohaddeseh Peyro, Devin L. Wakefield, Grace E. Choo, Mohammad R. K. Mofrad, and Tijana Jovanovic-Talisman. 2021. "Characterizing Binding Interactions That Are Essential for Selective Transport through the Nuclear Pore Complex" International Journal of Molecular Sciences 22, no. 19: 10898. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms221910898