Camphor and Eucalyptol—Anticandidal Spectrum, Antivirulence Effect, Efflux Pumps Interference and Cytotoxicity

 , , , ,
, , , ,  , and
, and
Abstract
:1. Introduction
2. Results
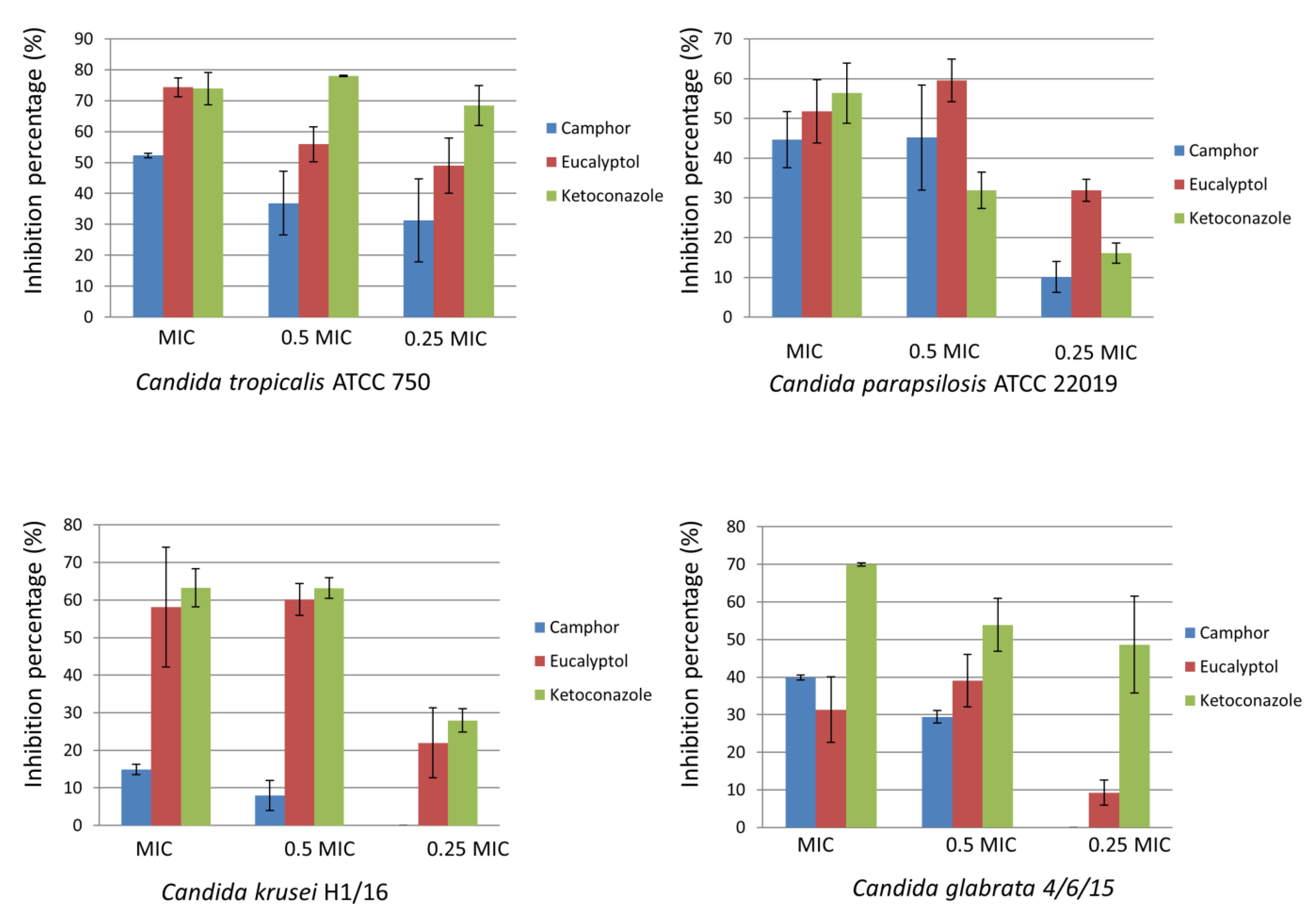
2.1. Impact of Camphor and Eucalyptol on Candida Albicans Growing in Planktonic and Biofilm Forms
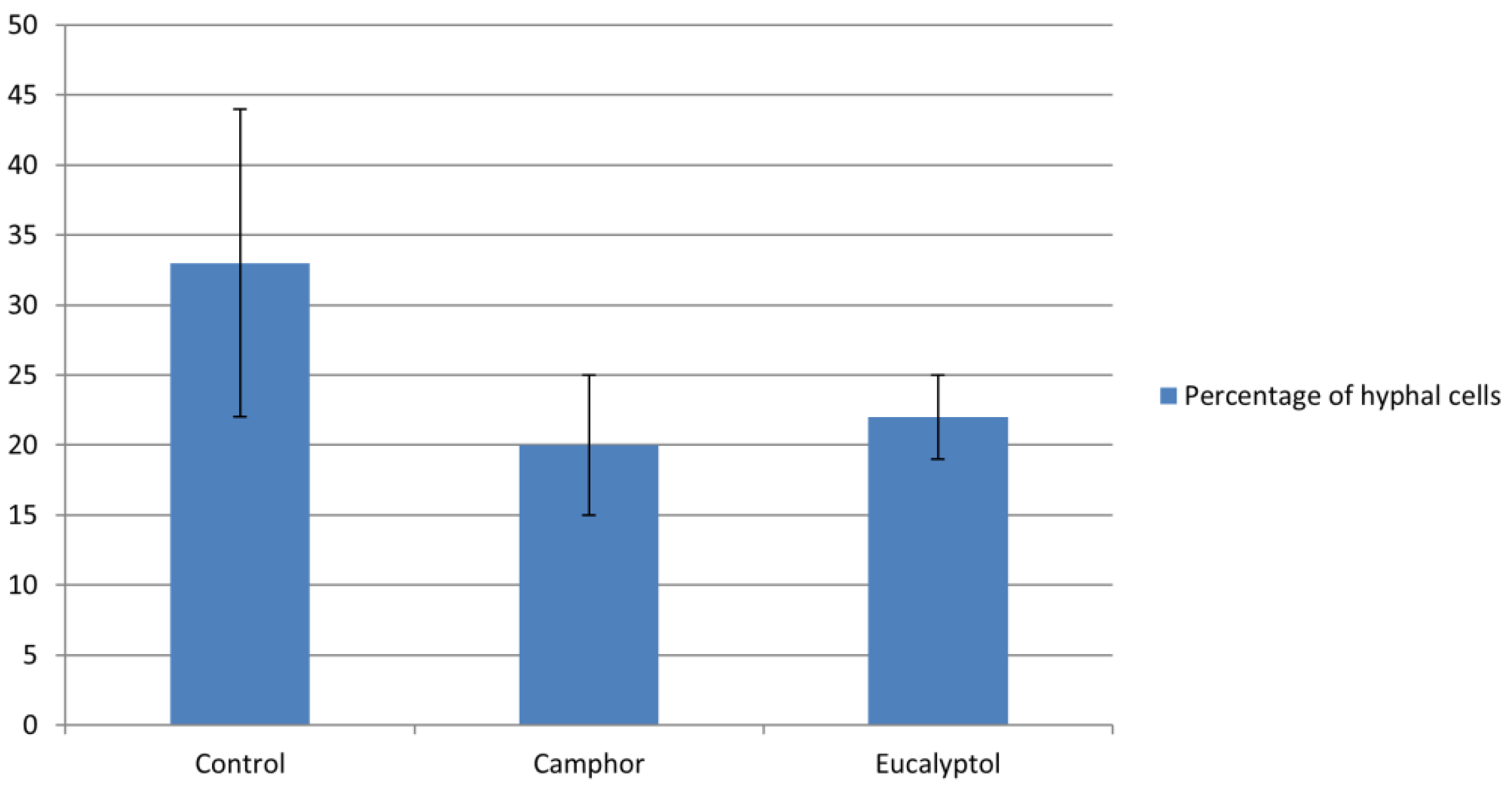
2.2. Camphor and Eucalyptol as Inhibitors of Candida Albicans Hyphae Formation
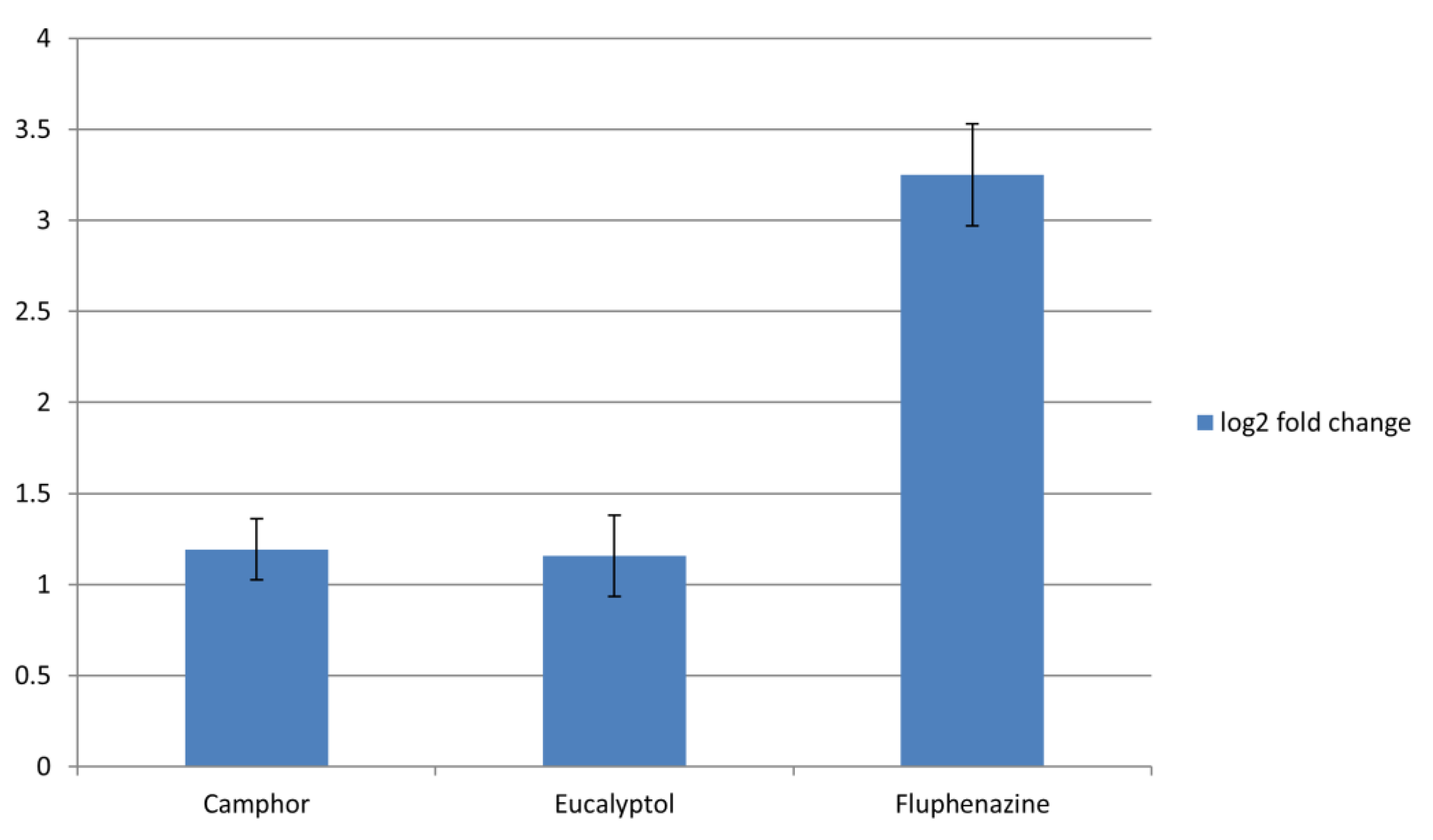
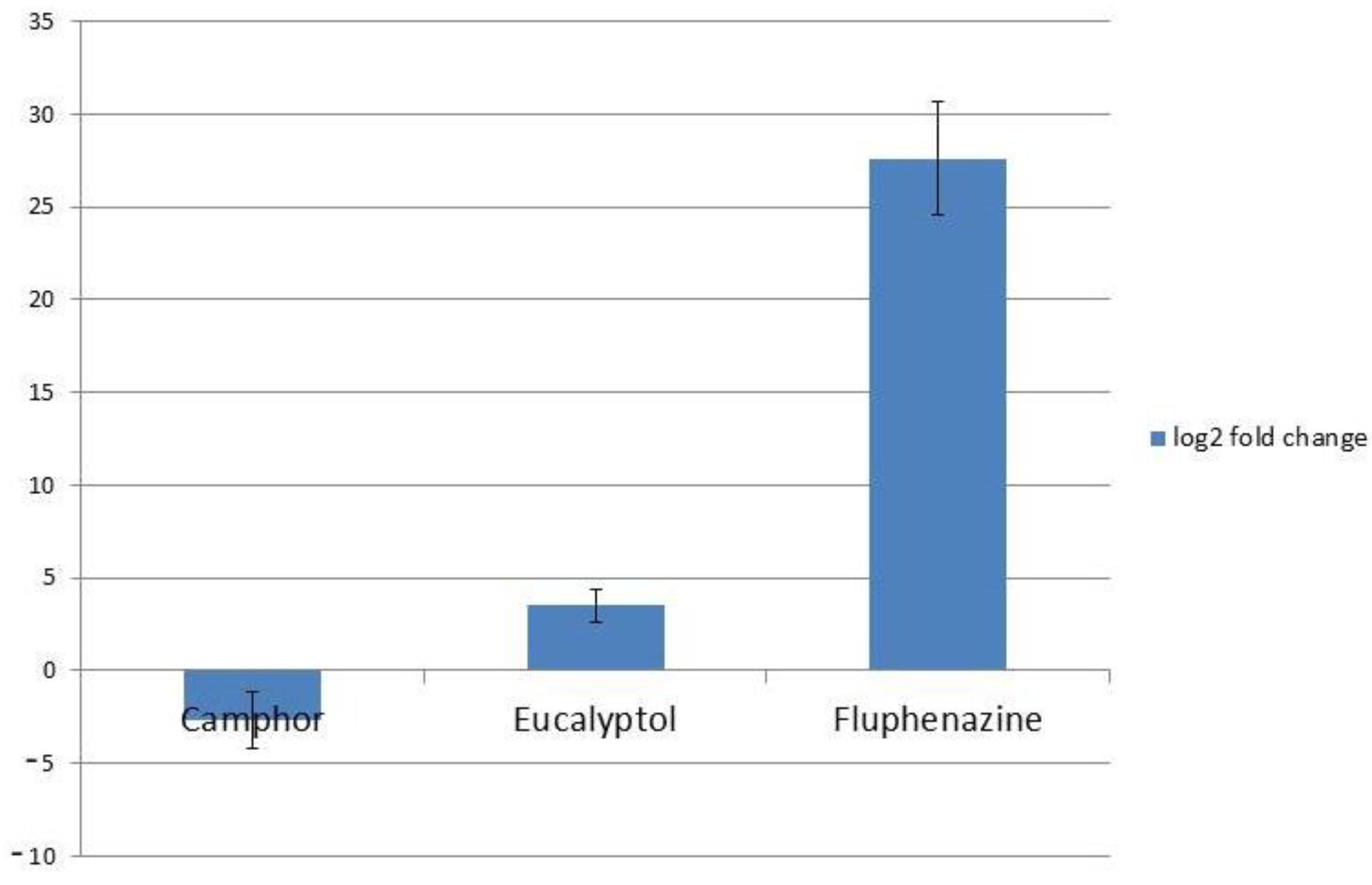
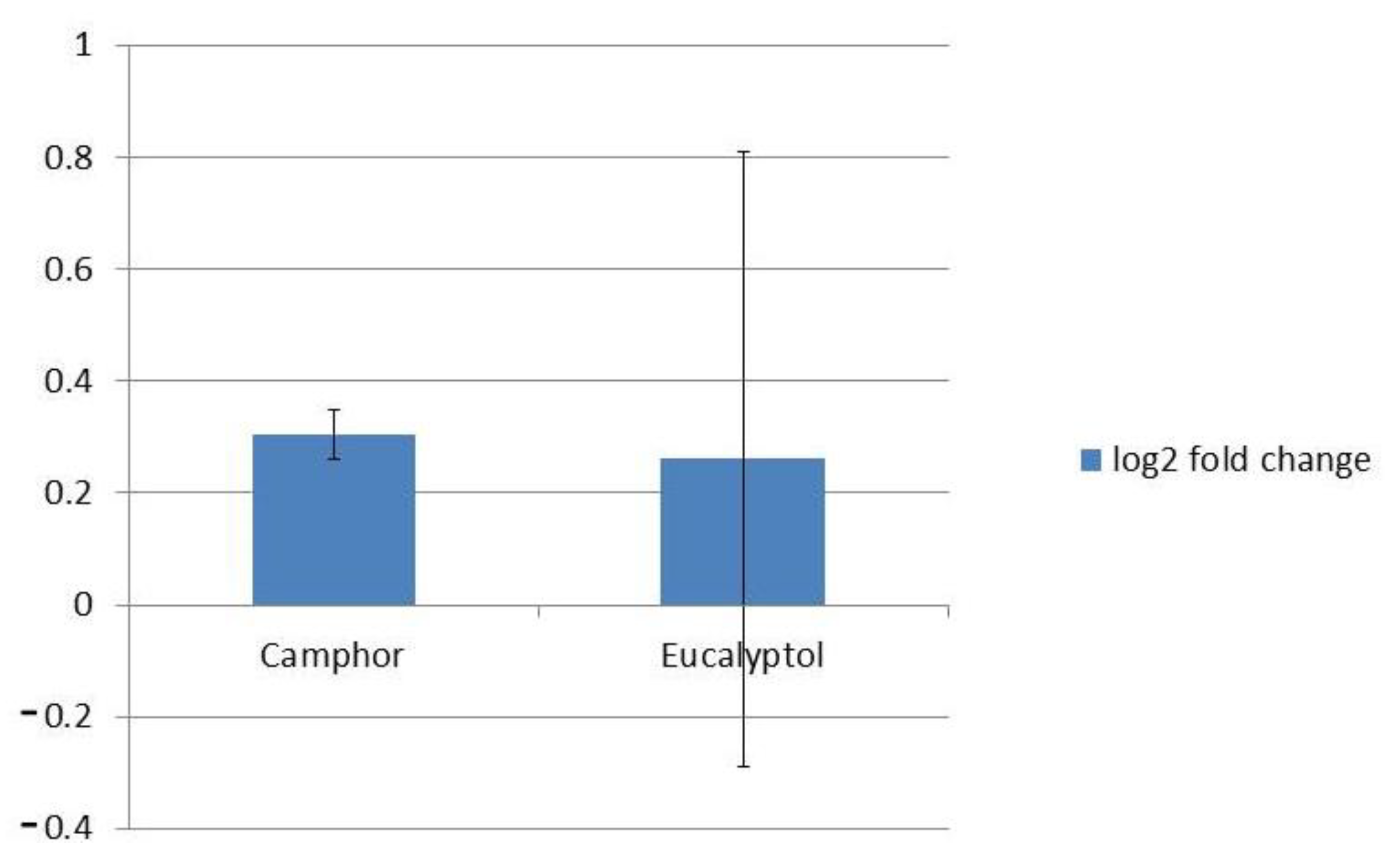
2.3. Impact on Genes Coding Fungal Efflux Pumps and Gene Involved in Ergosterol Biosynthesis
2.4. Camphor and Eucalyptol—Diverse Cytotoxicity to Porcine Liver Cells
3. Discussion
4. Materials and Methods
4.1. Microbial Culture Conditions
4.2. Microdilution Method
4.3. Antibiofilm Activity
4.4. Inhibition of Morphological Transition
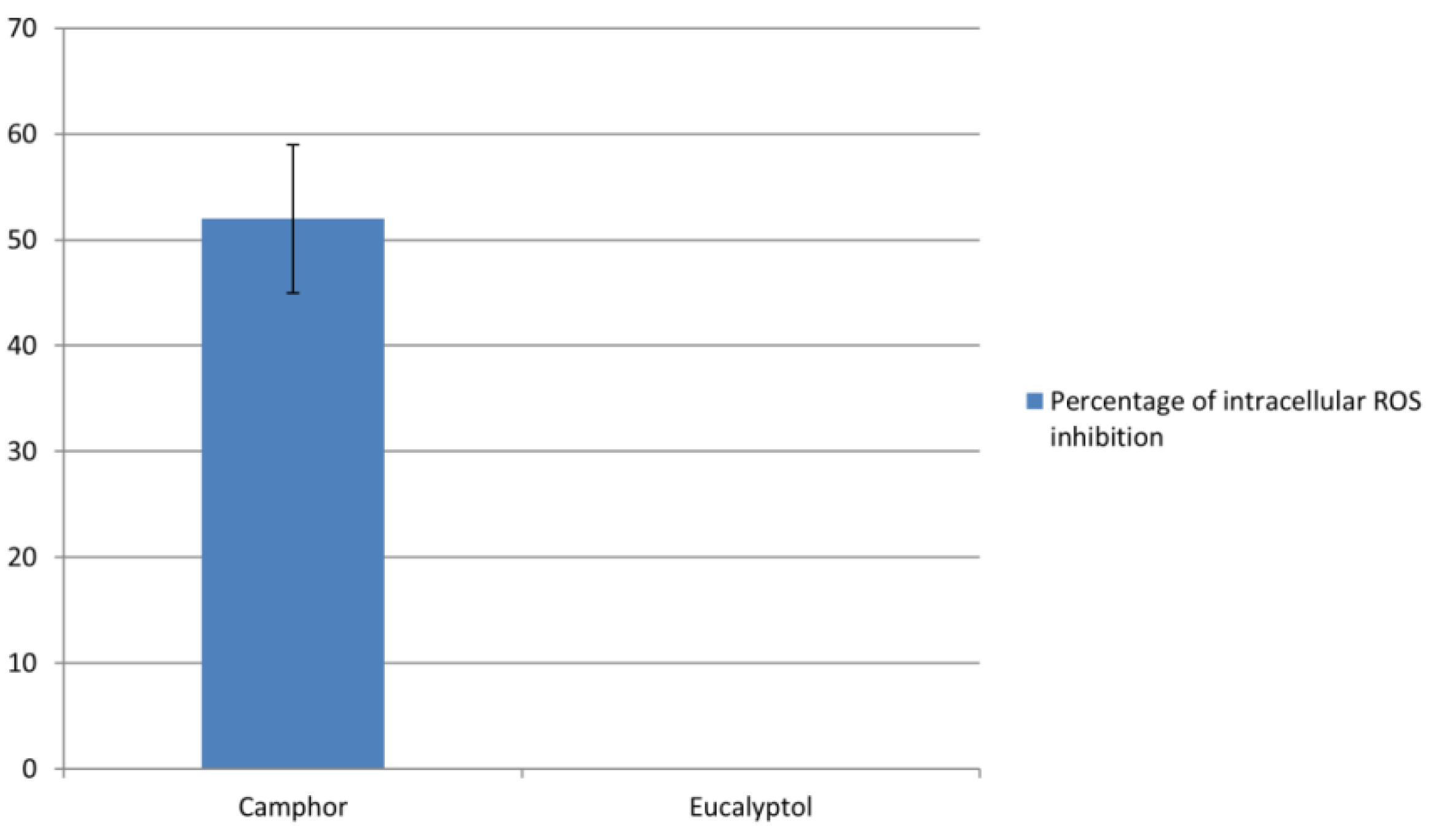
4.5. Determination of Intracellular ROS Levels in C. albicans 475/15
4.6. RNA Isolation and cDNA Synthesis
4.7. qPCR
4.8. Cytotoxicity of Compounds to Porcine Liver Primary Cells
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| ROS | Reactive oxygen species |
| MIC | Minimum inhibitory concentration |
| MFC | Minimum fungicidal concentration |
| PBS | Phosphate buffered saline |
| FC | Fold change |
References
- Janbon, G.; Quintin, J.; Lanternier, F.; d’Enfert, C. Studying fungal pathogens of humans and fungal infections: Fungal diversity and diversity of approaches. Genes Immun. 2019, 20, 403–414. [Google Scholar] [CrossRef] [PubMed]
- Nikou, S.A.; Kichik, N.; Brown, R.; Ponde, N.O.; Ho, J.; Naglik, J.R.; Richardson, J.P. Candida albicans interactions with mucosal surfaces during health and disease. Pathogens 2019, 22, 53. [Google Scholar] [CrossRef] [Green Version]
- Xiao, Z.; Wang, Q.; Zhu, F.; An, Y. Epidemiology, species distribution, antifungal susceptibility and mortality risk factors of candidemia among critically ill patients: A retrospective study from 2011 to 2017 in a teaching hospital in China. Antimicrob. Resist. Infect. Control 2019, 8, 89. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira Santos, G.C.; Vasconcelos, C.C.; Lopes, A.J.O.; de Sousa Cartágenes, M.; Filho, A.; do Nascimento, F.; Ramos, R.M.; Pires, E.; de Andrade, M.S.; Rocha, F.; et al. Candida infections and therapeutic strategies: Mechanisms of action for traditional and alternative agents. Front. Microbiol. 2018, 3, 1351. [Google Scholar] [CrossRef] [PubMed]
- Di Mambro, T.; Guerriero, I.; Aurisicchio, L.; Magnani, M.; Marra, E. The Yin and Yang of current antifungal therapeutic strategies: How can we harness our natural defenses? Front. Pharmacol. 2019, 5, 80. [Google Scholar] [CrossRef] [Green Version]
- Smiljković, M.; Kostić, M.; Stojković, D.; Glamočlija, J.; Soković, M. Could flavonoids compete with synthetic azoles in diminishing Candida albicans infections? A comparative review based on in vitro studies. Curr. Med. Chem. 2019, 26, 2536–2554. [Google Scholar] [CrossRef]
- Dadar, M.; Tiwari, R.; Karthik, K.; Chakraborty, S.; Shahali, Y.; Dhama, K. Candida albicans—Biology, molecular characterization, pathogenicity, and advances in diagnosis and control—An update. Microb. Pathog. 2018, 117, 128–138. [Google Scholar] [CrossRef]
- Zida, A.; Bamba, S.; Yacouba, A.; Ouedraogo-Traore, R.; Guiguemdé, R.T. Anti-Candida albicans natural products, sources of new antifungal drugs: A review. J. Mycol. Med. 2017, 27, 1–19. [Google Scholar] [CrossRef]
- Cavalheiro, M.; Teixeira, M.C. Candida biofilms: Threats, challenges, and promising strategies. Front. Med. 2018, 13, 28. [Google Scholar] [CrossRef] [Green Version]
- Askoura, M.; Mottawea, W.; Abujamel, T.; Taher, I. Efflux pump inhibitors (EPIs) as new antimicrobial agents against Pseudomonas aeruginosa. Libyan J. Med. 2011, 13, 6. [Google Scholar] [CrossRef] [Green Version]
- Tegos, G.P.; Haynes, M.; Strouse, J.J.; Khan, M.M.; Bologa, C.G.; Oprea, T.I.; Sklar, L.A. Microbial efflux pump inhibition: Tactics and strategies. Curr. Pharm. Des. 2011, 17, 1291–1302. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Venter, H.; Ma, S. Efflux pump inhibitors: A novel approach to combat efflux-mediated drug resistance in bacteria. Curr. Drug Targets 2016, 17, 702–719. [Google Scholar] [CrossRef]
- Sanglard, D.; Coste, A. Activity of isavuconazole and other azoles against Candida clinical isolates and yeast model systems with known azole resistance mechanisms. Antimicrob. Agents Chemother. 2016, 60, 229–238. [Google Scholar] [CrossRef] [Green Version]
- Kofla, G.; Turner, V.; Schulz, B.; Storch, U.; Froelich, D.; Rognon, B.; Coste, A.T.; Sanglard, D.; Ruhnke, M. Doxorubicin induces drug efflux pumps in Candida albicans. Med. Mycol. 2011, 49, 132–142. [Google Scholar] [CrossRef] [Green Version]
- Ivanov, M.; Kannan, A.; Stojković, D.; Glamočlija, J.; Grdadolnik, S.G.; Sanglard, D.; Soković, M. Revealing the astragalin mode of anticandidal action. EXCLI J. 2020, 19, 1436–1445. [Google Scholar] [PubMed]
- Lohberger, A.; Coste, A.T.; Sanglard, D. Distinct roles of Candida albicans drug resistance transcription factors TAC1, MRR1, and UPC2 in virulence. Eukaryot. Cell 2014, 13, 127–142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nikolić, M.; Smiljković, M.; Marković, T.; Ćirić, A.; Glamočlija, J.; Markovic, D.; Soković, M. Sensitivity of clinical isolates of Candida to essential oils from Burseraceae family. EXCLI J. 2016, 19, 280–289. [Google Scholar]
- Smiljković, M.; Dias, M.I.; Stojković, D.; Barros, L.; Bukvički, D.; Ferreira, I.; Soković, M. Characterization of phenolic compounds in tincture of edible Nepeta nuda: Development of antimicrobial mouthwash. Food Funct. 2018, 9, 5417–5425. [Google Scholar] [CrossRef] [Green Version]
- Smiljković, M.; Stanisavljević, D.; Stojković, D.; Petrović, I.; Marjanović Vicentić, J.; Popović, J.; Golic Grdadolnik, S.; Marković, D.; Sanković-Babić, S.; Glamočlija, J.; et al. Apigenin-7-O-glucoside versus apigenin: Insight into the modes of anticandidal and cytotoxic actions. EXCLI J. 2017, 16, 795–807. [Google Scholar]
- Paduch, R.; Trytek, M.; Król, S.K.; Kud, J.; Frant, M.; Kandefer-Szerszeń, M.; Fiedurek, J. Biological activity of terpene compounds produced by biotechnological methods. Pharm. Biol. 2016, 54, 1096–1107. [Google Scholar] [CrossRef] [Green Version]
- Petrović, J.; Stojković, D.; Soković, M. Terpene core in selected aromatic and edible plants: Natural health improving agents. Adv. Food Nutr. Res. 2019, 90, 423–451. [Google Scholar] [PubMed]
- Schröter, C.; Hipler, U.C.; Wilmer, A.; Künkel, W.; Wollina, U. Generation of reactive oxygen species by Candida albicans in relation to morphogenesis. Arch. Dermatol. Res. 2000, 292, 260–264. [Google Scholar] [CrossRef] [PubMed]
- Devkatte, A.N.; Zore, G.B.; Karuppayil, S.M. Potential of plant oils as inhibitors of Candida albicans growth. FEMS Yeast Res. 2005, 5, 867–873. [Google Scholar] [CrossRef] [Green Version]
- Manoharan, R.K.; Lee, J.H.; Lee, J. Antibiofilm and antihyphal activities of cedar leaf essential oil, camphor, and fenchone derivatives against Candida albicans. Front. Microbiol. 2017, 3, 1476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zuzarte, M.; Gonçalves, M.J.; Cavaleiro, C.; Canhoto, J.; Vale-Silva, L.; Silva, M.J.; Pinto, E.; Salgueiro, L. Chemical composition and antifungal activity of the essential oils of Lavandula viridis L’Her. J. Med. Microbiol. 2011, 60, 612–618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hendry, E.R.; Worthington, T.; Conway, B.R.; Lambert, P.A. Antimicrobial efficacy of eucalyptus oil and 1,8-cineole alone and in combination with chlorhexidine digluconate against microorganisms grown in planktonic and biofilm cultures. J. Antimicrob. Chemother. 2009, 64, 1219–1225. [Google Scholar] [CrossRef]
- Dalleau, S.; Cateau, E.; Bergès, T.; Berjeaud, J.M.; Imbert, C. In vitro activity of terpenes against Candida biofilms. Int. J. Antimicrob. Agents 2008, 31, 572–576. [Google Scholar] [CrossRef]
- Raut, J.S.; Shinde, R.B.; Chauhan, N.M.; Karuppayil, S.M. Terpenoids of plant origin inhibit morphogenesis, adhesion, and biofilm formation by Candida albicans. Biofouling 2013, 29, 87–96. [Google Scholar] [CrossRef]
- Bin Jantan, I.; Moharam, B.A.K.; Santhanam, J.; Jamal, J.A. Correlation between chemical composition and antifungal activity of the essential oils of eight Cinnamomum species. Pharm. Biol. 2008, 46, 406–412. [Google Scholar] [CrossRef] [Green Version]
- Abu-Darwish, M.S.; Cabral, C.; Ferreira, I.V.; Gonçalves, M.J.; Cavaleiro, C.; Cruz, M.T.; Al-bdour, T.H.; Salgueiro, L. Essential oil of common sage (Salvia officinalis L.) from Jordan: Assessment of safety in mammalian cells and its antifungal and anti-inflammatory potential. Biomed. Res. Int. 2013, 2013, 538940. [Google Scholar] [CrossRef] [Green Version]
- Soković, M.; Tzakou, O.; Pitarokili, D.; Couladis, M. Antifungal activities of selected aromatic plants growing wild in Greece. Nahrung 2002, 46, 317–320. [Google Scholar] [CrossRef]
- Yu, D.; Wang, J.; Shao, X.; Xu, F.; Wang, H. Antifungal modes of action of tea tree oil and its two characteristic components against Botrytis cinerea. J. Appl. Microbiol. 2015, 119, 1253–1262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sancineto, L.; Piccioni, M.; De Marco, S.; Pagiotti, R.; Nascimento, V.; Braga, A.L.; Santi, C.; Pietrella, D. Diphenyl diselenide derivatives inhibit microbial biofilm formation involved in wound infection. BMC Microbiol. 2016, 16, 220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merghni, A.; Noumi, E.; Hadded, O.; Dridi, N.; Panwar, H.; Ceylan, O.; Mastouri, M.; Snoussi, M. Assessment of the antibiofilm and antiquorum sensing activities of Eucalyptus globulus essential oil and its main component 1,8-cineole against methicillin-resistant Staphylococcus aureus strains. Microb. Pathog. 2018, 118, 74–80. [Google Scholar] [CrossRef]
- Cherneva, E.; Pavlovic, V.; Smelcerovic, A.; Yancheva, D. The effect of camphor and borneol on rat thymocyte viability and oxidative stress. Molecules 2012, 27, 10258–10266. [Google Scholar] [CrossRef]
- Porres-Martínez, M.; González-Burgos, E.; Carretero, M.E.; Gómez-Serranillos, M.P. Major selected monoterpenes α-pinene and 1,8-cineole found in Salvia lavandulifolia (Spanish sage) essential oil as regulators of cellular redox balance. Pharm. Biol. 2015, 53, 921–929. [Google Scholar] [CrossRef]
- Lima, S.L.; Colombo, A.L.; de Almeida, J.N., Jr. Fungal cell wall: Emerging antifungals and drug resistance. Front. Microbiol. 2019, 10, 2573. [Google Scholar] [CrossRef] [Green Version]
- Saviuc, C.; Gheorghe, I.; Coban, S.; Drumea, V.; Chifiriuc, M.; Banu, O.; Bezirtzoglou, E.; Laz, V. Rosmarinus officinalis essential oil and eucalyptol act as efflux pumps inhibitors and increase ciprofloxacin efficiency against Pseudomonas aeruginosa and Acinetobacter baumannii MDR strains. Rom. Biotechnol. Lett. 2016, 21, 11782–11790. [Google Scholar]
- Scorzoni, L.; de Paula E Silva, A.C.; Marcos, C.M.; Assato, P.A.; de Melo, W.C.; de Oliveira, H.C.; Costa-Orlandi, C.B.; Mendes-Giannini, M.J.; Fusco-Almeida, A.M. Antifungal therapy: New advances in the understanding and treatment of mycosis. Front. Microbiol. 2017, 8, 36. [Google Scholar] [CrossRef] [Green Version]
- Coste, A.T.; Karababa, M.; Ischer, F.; Bille, J.; Sanglard, D. TAC1, transcriptional activator of CDR genes, is a new transcription factor involved in the regulation of Candida albicans ABC transporters CDR1 and CDR2. Eukaryot. Cell 2004, 3, 1639–1652. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.S.; Yang, Y.L.; Wu, C.J.; Ouyang, K.J.; Tseng, K.Y.; Chen, C.G.; Wang, H.; Lo, H.J. The DNA-binding domain of CaNdt80p is required to activate CDR1 involved in drug resistance in Candida albicans. J. Med. Microbiol. 2006, 55, 1403–1411. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, A.; Khan, A.; Manzoor, N. Reversal of efflux mediated antifungal resistance underlies synergistic activity of two monoterpenes with fluconazole. Eur. J. Pharm. Sci. 2013, 23, 80–86. [Google Scholar] [CrossRef] [PubMed]
- Lorenzi, V.; Muselli, A.; Bernardini, A.F.; Berti, L.; Pagès, J.M.; Amaral, L.; Bolla, J.M. Geraniol restores antibiotic activities against multidrug-resistant isolates from gram-negative species. Antimicrob. Agents Chemother. 2009, 53, 2209–2211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dwivedi, G.R.; Maurya, A.; Yadav, D.K.; Khan, F.; Darokar, M.P.; Srivastava, S.K. Drug resistance reversal potential of ursolic acid derivatives against nalidixic acid and multidrug-resistant Escherichia coli. Chem. Biol. Drug Des. 2015, 86, 272–283. [Google Scholar] [CrossRef] [PubMed]
- Paul, S.; Singh, S.; Sharma, D.; Chakrabarti, A.; Rudramurthy, S.M.; Ghosh, A.K. Dynamics of in-vitro development of azole resistance in Candida tropicalis. J. Glob. Antimicrob. Resist. 2020, 22, 553–561. [Google Scholar] [CrossRef] [PubMed]
- Tian, J.; Ban, X.; Zeng, H.; He, J.; Chen, Y.; Wang, Y. The mechanism of antifungal action of essential oil from dill (Anethum graveolens L.) on Aspergillus flavus. PLoS ONE 2012, 7, e30147. [Google Scholar] [CrossRef] [Green Version]
- Cardoso, N.N.; Alviano, C.S.; Blank, A.F.; Romanos, M.T.; Fonseca, B.; Rozental, S.; Rodrigues, I.; Alviano, D. Synergism effect of the essential oil from Ocimum basilicum var. Maria Bonita and its major components with fluconazole and its influence on ergosterol biosynthesis. Evid. Based Complement. Alternat. Med. 2016, 2016, 5647182. [Google Scholar] [CrossRef] [Green Version]
- Kalagatur, N.K.; Nirmal Ghosh, O.S.; Sundararaj, N.; Mudili, V. Antifungal activity of chitosan nanoparticles encapsulated with Cymbopogon martinii essential oil on plant pathogenic fungi Fusarium graminearum. Front. Pharmacol. 2018, 6, 610. [Google Scholar] [CrossRef] [Green Version]
- Połeć, K.; Wójcik, A.; Flasiński, M.; Wydro, P.; Broniatowski, M.; Hąc-Wydro, K. The influence of terpinen-4-ol and eucalyptol—The essential oil components—On fungi and plant sterol monolayers. Biochim. Biophys. Acta Biomembr. 2019, 1861, 1093–1102. [Google Scholar] [CrossRef]
- Stojković, D.; Dias, M.I.; Drakulić, D.; Barros, L.; Stevanović, M.; Ferreira, I.C.F.R.; Soković, M. Methanolic extract of the herb Ononis spinosa L. is an antifungal agent with no cytotoxicity to primary human cells. Pharmaceuticals 2020, 13, 78. [Google Scholar] [CrossRef]
- Nikolić, B.; Vasilijević, B.; Mitić-Ćulafić, D.; Vuković-Gačić, B.; Knežević-Vukčević, J. Comparative study of genotoxic, antigenotoxic and cytotoxic activities of monoterpenes camphor, eucalyptol and thujone in bacteria and mammalian cells. Chem. Biol. Interact. 2015, 5, 263–271. [Google Scholar] [CrossRef] [PubMed]
- Zaccaro Scelza, M.F.; Lima Oliveira, L.R.; Carvalho, F.B.; Côrte-Real Faria, S. In vitro evaluation of macrophage viability after incubation in orange oil, eucalyptol, and chloroform. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. Endodontol. 2006, 102, e24–e27. [Google Scholar] [CrossRef] [PubMed]
- EUCAST (European Committee on Antibiotic Susceptibility). Method for Determination of Minimal Inhibitory Concentration (MIC) by Broth Dilution of Fermentative Yeasts; Discussion Document E. Dis. 7.1; European Society of Clinical Microbiology and Infectious Diseases: Taufkirchen, Germany, 2002. [Google Scholar]
- Smiljkovic, M.; Matsoukas, M.T.; Kritsi, E.; Zelenko, U.; Grdadolnik, S.G.; Calhelha, R.C.; Ferreira, I.C.F.R.; Sankovic-Babic, S.; Glamoclija, J.; Fotopoulou, T.; et al. Nitrate esters of heteroaromatic compounds as Candida albicans CYP51 enzyme inhibitors. ChemMedChem 2018, 6, 251–258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paez, P.L.; Becerra, M.C.; Albesa, I. Effect of the association of reduced glutathione and ciprofloxacin on the antimicrobial activity in Staphylococcus aureus. FEMS Microbiol. Lett. 2010, 303, 101–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanglard, D.; Ischer, F.; Calabrese, D.; Majcherczyk, P.A.; Bille, J. The ATP binding cassette transporter GeneCgCDR1 from Candida glabrata is involved in the resistance of clinical isolates to azole antifungal agents. Antimicrob. Agents Chemother. 1999, 43, 2753–2765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ivanov, M.; Kannan, A.; Stojkovic, D.; Glamoclija, J.; Calhelha, R.; Ferreira, I.; Sanglard, D.; Sokovic, M. Flavones, Flavonols, and Glycosylated Derivatives—Impact on Candida albicans Growth and Virulence, Expression of CDR1 and ERG11, Cytotoxicity. Pharmaceuticals 2021, 14, 27. [Google Scholar] [CrossRef]
- Abreu, R.M.V.; Ferreira, I.C.F.R.; Calhelha, R.C.; Lima, R.T.; Vasconcelos, M.H.; Adega, F.; Chaves, R.; Queiroz, M.J.R.P. Anti-hepatocellular carcinoma activity using human HepG2 cells and hepatotoxicity of 6-substituted methyl 3-aminothieno[3,2-b]pyridine-2-carboxylate derivatives: In vitro evaluation, cell cycle analysis and QSAR studies. Eur. J. Med. Chem. 2011, 46, 5800–5806. [Google Scholar] [CrossRef] [Green Version]
- Guimaraes, R.; Barros, L.; Duenas, M.; Calhelha, R.; Carvalho, A.M.; Santos-Buelga, C.; Queiroz, M.J.; Ferreira, I.C.F.R. Nutrients, phytochemicals and bioactivity of wild Roman chamomile: A comparison between the herb and its preparations. Food Chem. 2013, 136, 718–725. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Camphor | Eucalyptol | Ketoconazole | |||
|---|---|---|---|---|---|---|
| MIC | MFC | MIC | MFC | MIC | MFC | |
| C. albicans 475/15 | 0.125 ± 0.01 b | 0.25 ± 0.02 b | 23 ± 0.1 c | 46 ± 0.2 c | 0.0031 ± 0.0001 a | 0.0062 ± 0.0001 a |
| C. albicans 527/14 | 0.125 ± 0.01 b | 0.25 ± 0.02 b | 23 ± 0.2 c | 46 ± 0.2 c | 0.0031 ± 0.0001 a | 0.0062 ± 0.0001 a |
| C. albicans 10/15 | 0.125 ± 0.02 b | 0.25 ± 0.02 b | 23 ± 0.1 c | 46 ± 0.2 c | 0.0031 ± 0.0001 a | 0.05 ± 0.0001 a |
| C. albicans 27/15 | 0.25 ± 0.004 b | 0.5 ± 0.02 b | 6 ± 0.08 c | 12 ± 0.2 c | 0.0031 ± 0.001 a | 0.1 ± 0.01 a |
| C. albicans 532/15 | 0.125 ± 0.01 b | 0.25 ± 0.01 b | 6 ± 0.08 c | 12 ± 0.2 c | 0.0031 ± 0.0001 a | 0.0062 ± 0.0001 a |
| C. albicans 503/15 | 0.25 ± 0.008 b | 0.5 ± 0.02 b | 3 ± 0.06 c | 6 ± 0.08 c | 0.0031 ± 0.0001 a | 0.0062 ± 0.0001 a |
| C. albicans 13/15 | 0.25 ± 0.01 b | 0.5 ± 0.008 b | 6 ± 0.08 c | 12 ± 0.2 c | 0.0016 ± 0.001 a | 0.05 ± 0.002 a |
| C. albicans 16/15 | 0.125 ± 0.008 b | 0.25 ± 0.004 b | 6 ± 0.1 c | 12 ± 0.2 c | 0.0031 ± 0.001 a | 0.1 ± 0.001 a |
| C. albicans ATCC 10231 | 0.175 ± 0.02 b | 0.35 ± 0.02 b | 4 ± 0.06 c | 8 ± 0.008 c | 0.0016 ± 0.001 a | 0.0062 ± 0.001 a |
| C. tropicalis ATCC 750 | 0.175 ± 0.02 b | 0.35 ± 0.02 b | 4 ± 0.004 c | 8 ± 0.006 c | 0.0016 ± 0.002 a | 0.0062 ± 0.002 a |
| C. parapsilosis ATCC 22019 | 0.125 ± 0.003 b | 0.25 ± 0.008 b | 2 ± 0.003 c | 4 ± 0.003 c | 0.0031 ± 0.0001 a | 0.0062 ± 0.0001 a |
| C. krusei H1/16 | 0.35 ± 0.06 b | 0.7 ± 0.06 b | 4 ± 0.004 c | 8 ± 0.008 c | 0.0016 ± 0.001 a | 0.0032 ± 0.002 a |
| C. glabrata 4/6/15 | 0.175 ± 0.02 b | 0.35 ± 0.04 b | 2 ± 0.004 c | 4 ± 0.007 c | 0.0016 ± 0.001 a | 0.0062 ± 0.002 a |
| Compound | GI50 |
|---|---|
| Camphor | >400 |
| Eucalyptol | 56 ± 4 |
| Ellipticine | 3.22 ± 0.2 |
| Primer | Sequence |
| CDR1-ORF-F | ATGACTCGAGATATTTTGATA |
| CDR1-ORF-R | TTAACAGCAATGGTCTTTA |
| CDR2-ORF-F | TAGATATTTGAGCCACATG |
| CDR2-ORF-R | TTGGCATTGAAATTTTCG |
| ERG11-ORF-F | ATTGTTGAAACTGTCATTG |
| ERG11-ORF-R | CCCCTAATAATATACTGATCTG |
| ACT-ORF-F | GCATCACACTTTTTACAAT |
| ACT-ORF-R | AAACATAATTTGAGTCATCTTT |
| Probe | Sequence |
| CDR1-P2 | CATTATGAGACCTGGTGAACTTACT |
| CDR2-P2 | TTAGTCCATTCAACGGCAACATTAG |
| ERG11-P2 | TTTGTCCCTTAGTGTTACACA |
| ACT1-P2 | TTGCTCCAGAAGAACATCCAGT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ivanov, M.; Kannan, A.; Stojković, D.S.; Glamočlija, J.; Calhelha, R.C.; Ferreira, I.C.F.R.; Sanglard, D.; Soković, M. Camphor and Eucalyptol—Anticandidal Spectrum, Antivirulence Effect, Efflux Pumps Interference and Cytotoxicity. Int. J. Mol. Sci. 2021, 22, 483. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22020483
Ivanov M, Kannan A, Stojković DS, Glamočlija J, Calhelha RC, Ferreira ICFR, Sanglard D, Soković M. Camphor and Eucalyptol—Anticandidal Spectrum, Antivirulence Effect, Efflux Pumps Interference and Cytotoxicity. International Journal of Molecular Sciences. 2021; 22(2):483. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22020483
Chicago/Turabian StyleIvanov, Marija, Abhilash Kannan, Dejan S. Stojković, Jasmina Glamočlija, Ricardo C. Calhelha, Isabel C. F. R. Ferreira, Dominique Sanglard, and Marina Soković. 2021. "Camphor and Eucalyptol—Anticandidal Spectrum, Antivirulence Effect, Efflux Pumps Interference and Cytotoxicity" International Journal of Molecular Sciences 22, no. 2: 483. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22020483