Conditioned Medium from Human Amnion-Derived Mesenchymal Stromal/Stem Cells Attenuating the Effects of Cold Ischemia-Reperfusion Injury in an In Vitro Model Using Human Alveolar Epithelial Cells

 , ,
, ,  , ,
, ,
Abstract
:1. Introduction
2. Results
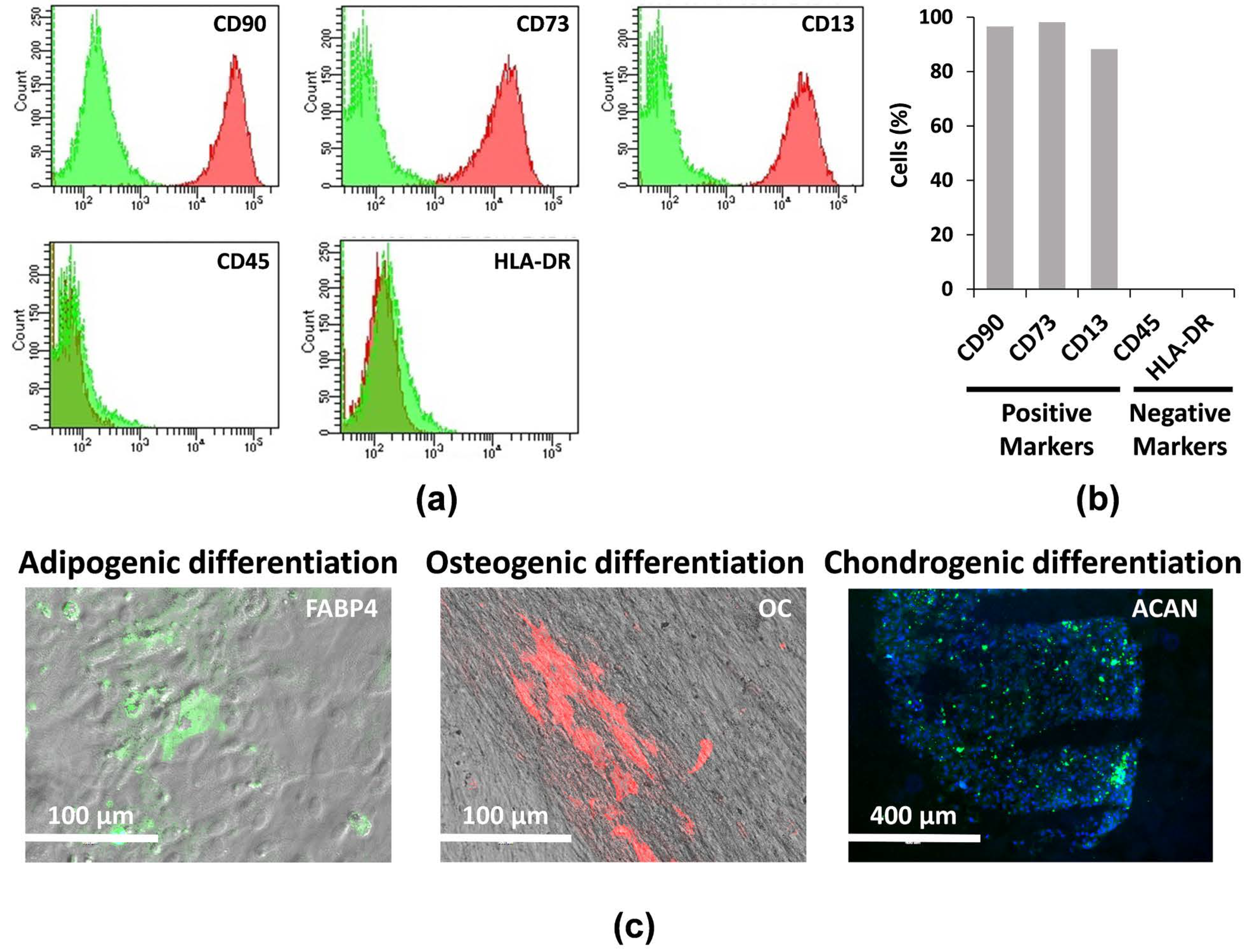
2.1. Characterization, Culture, and Differentiation of hAMSCs
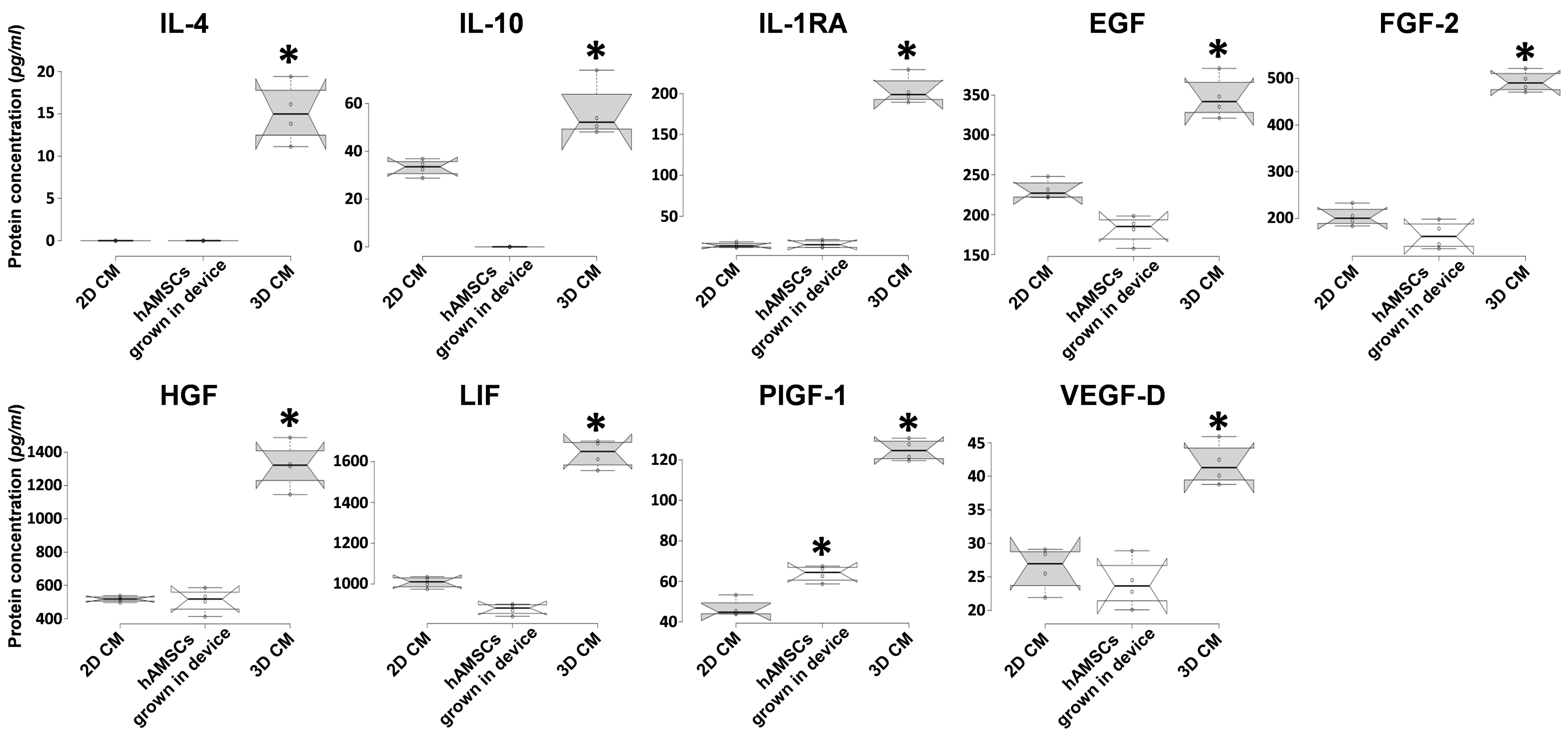
2.2. Conditioned Medium Derived from 3D hAMSC Cultures Showing Increased Production of Both Growth and Immunomodulatory Factors
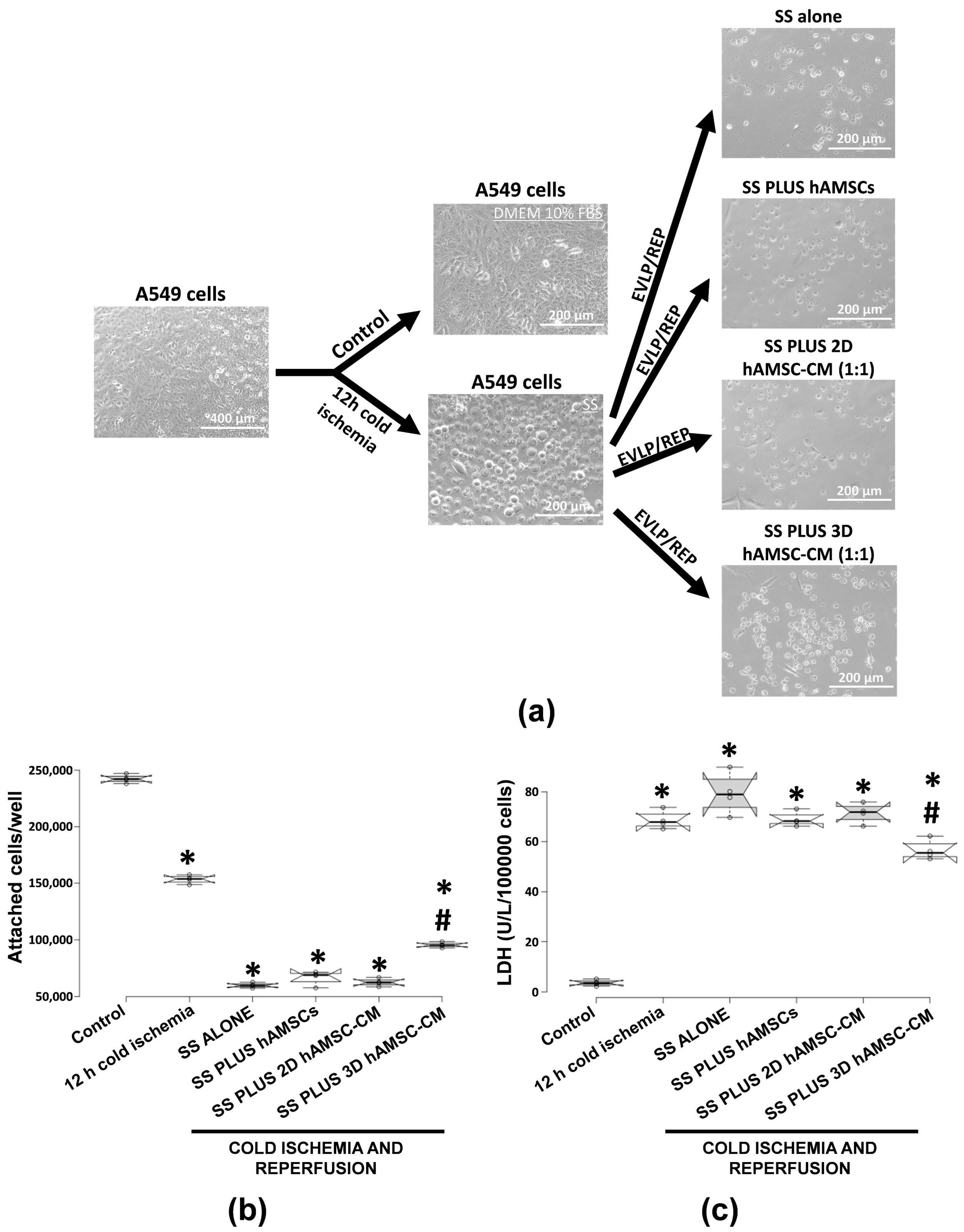
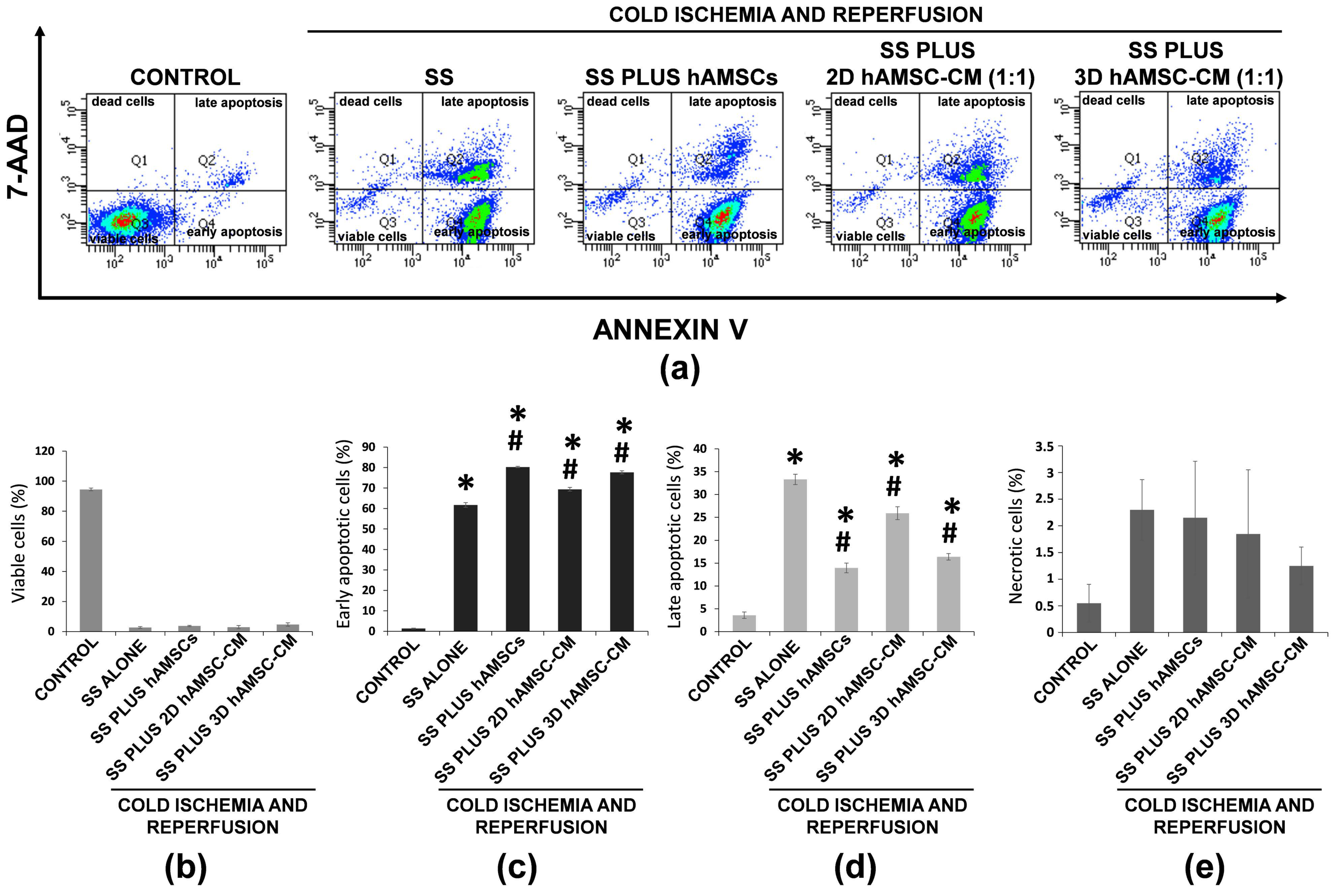
2.3. Protective Effects of Encapsulated-hAMSCs and hAMSC-CM against Adverse Effects Induced by IRI on the Morphology and Viability of Human Alveolar Basal Epithelial Cells
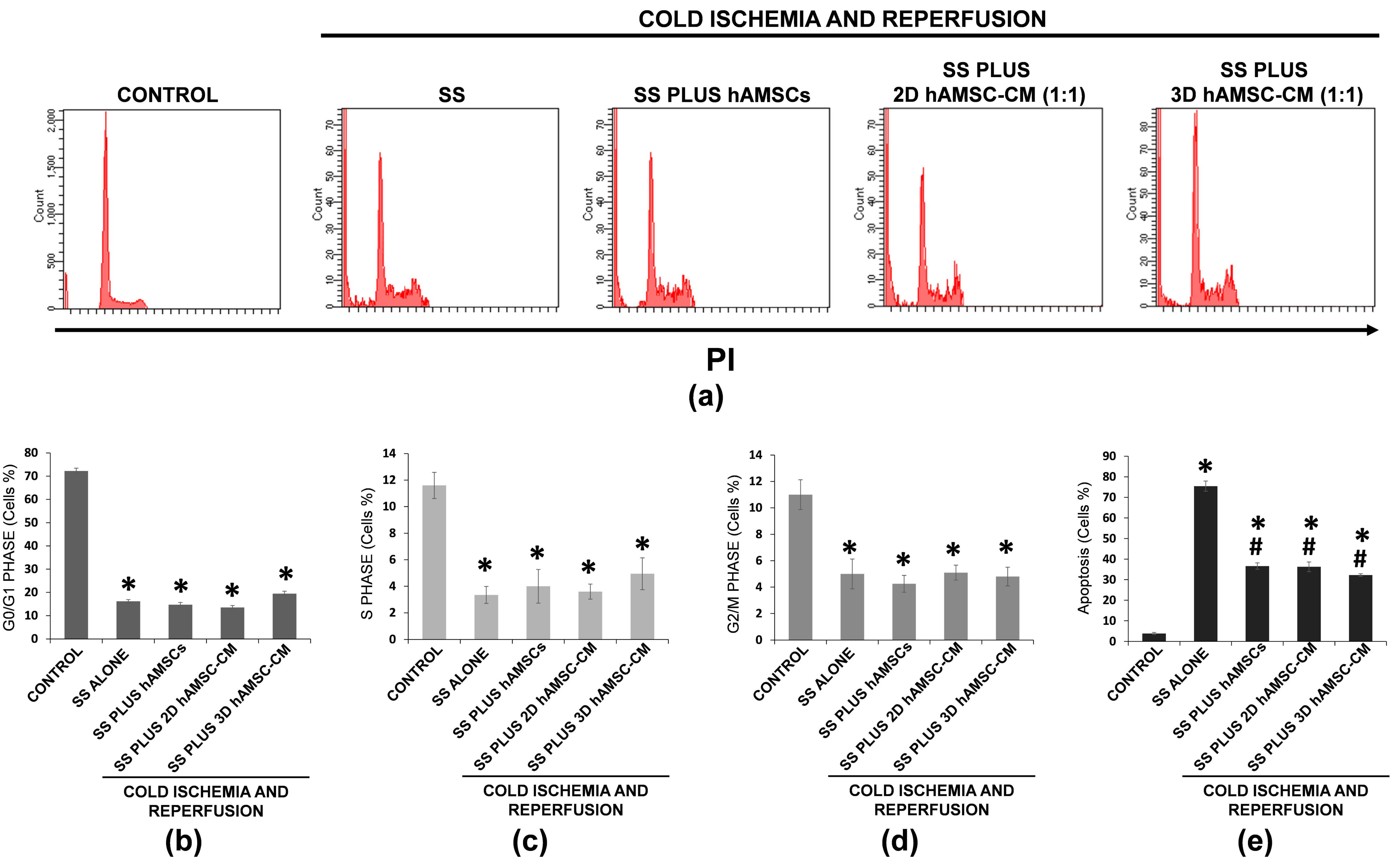
2.4. Effects of Encapsulated-hAMSCs and hAMSC-CM against Adverse Effects Induced by IRI on Apoptosis and Cell Cycle Progression of Human Alveolar Basal Epithelial Cells
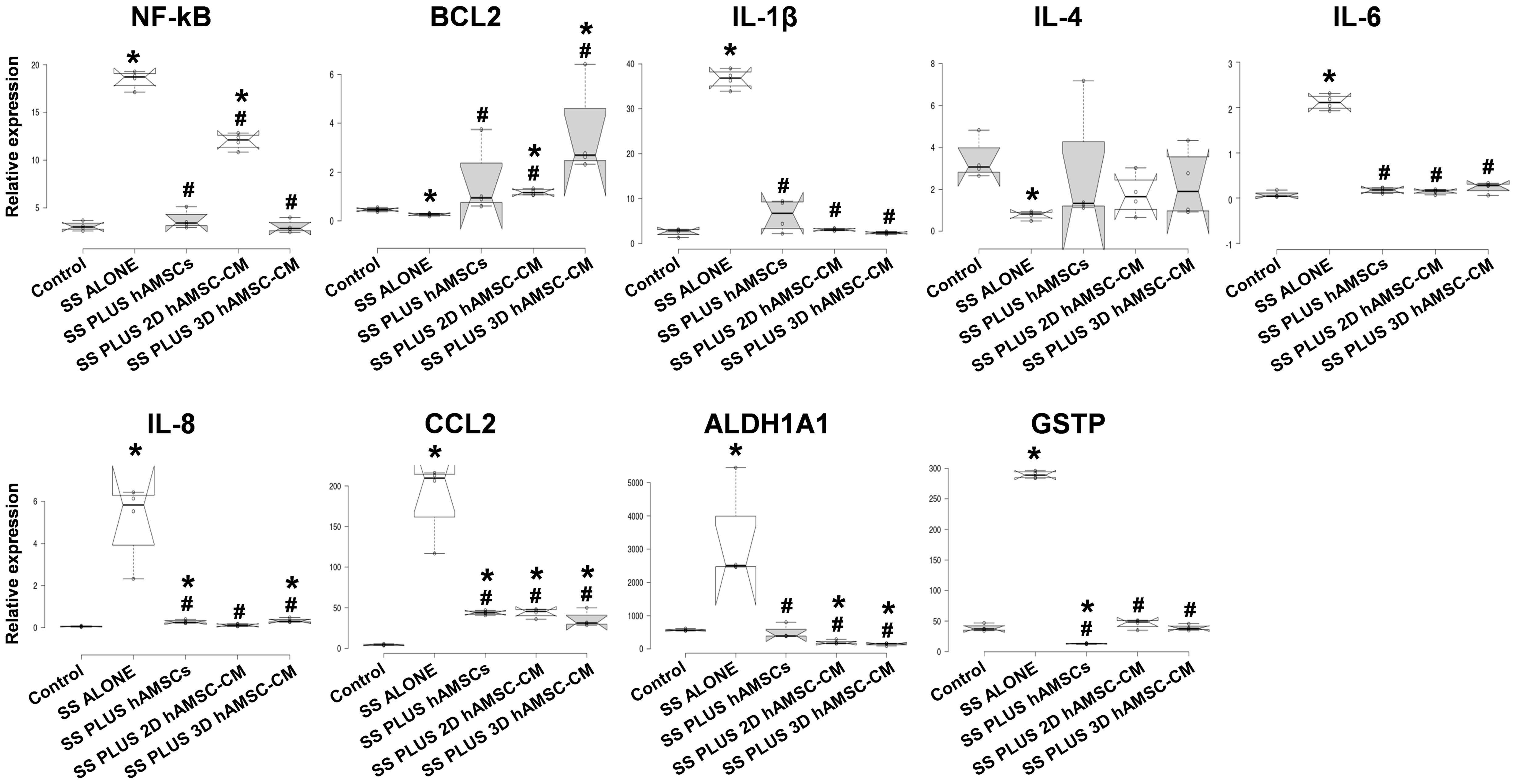
2.5. Effects of Encapsulated-hAMSCs and hAMSC-CM against Adverse Effects Induced by IRI on Expression of Inflammatory, Apoptotic, and Antioxidant Mediators in Human Alveolar Basal Epithelial Cells
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Isolation and Culture of Human Amnion-Derived Mesenchymal Stem/Stromal Cells
4.3. Device Preparation
4.4. Flow Cytometric Phenotypic Analysis
4.5. Induction of Osteogenic, Adipogenic, and Chondrogenic Differentiation
4.6. Formation of Mesenchymal Stromal/Stem Cell Spheroids
4.7. Conditioned Media Preparation
4.8. Gene Expression Analysis
4.9. Protein Expression Analysis
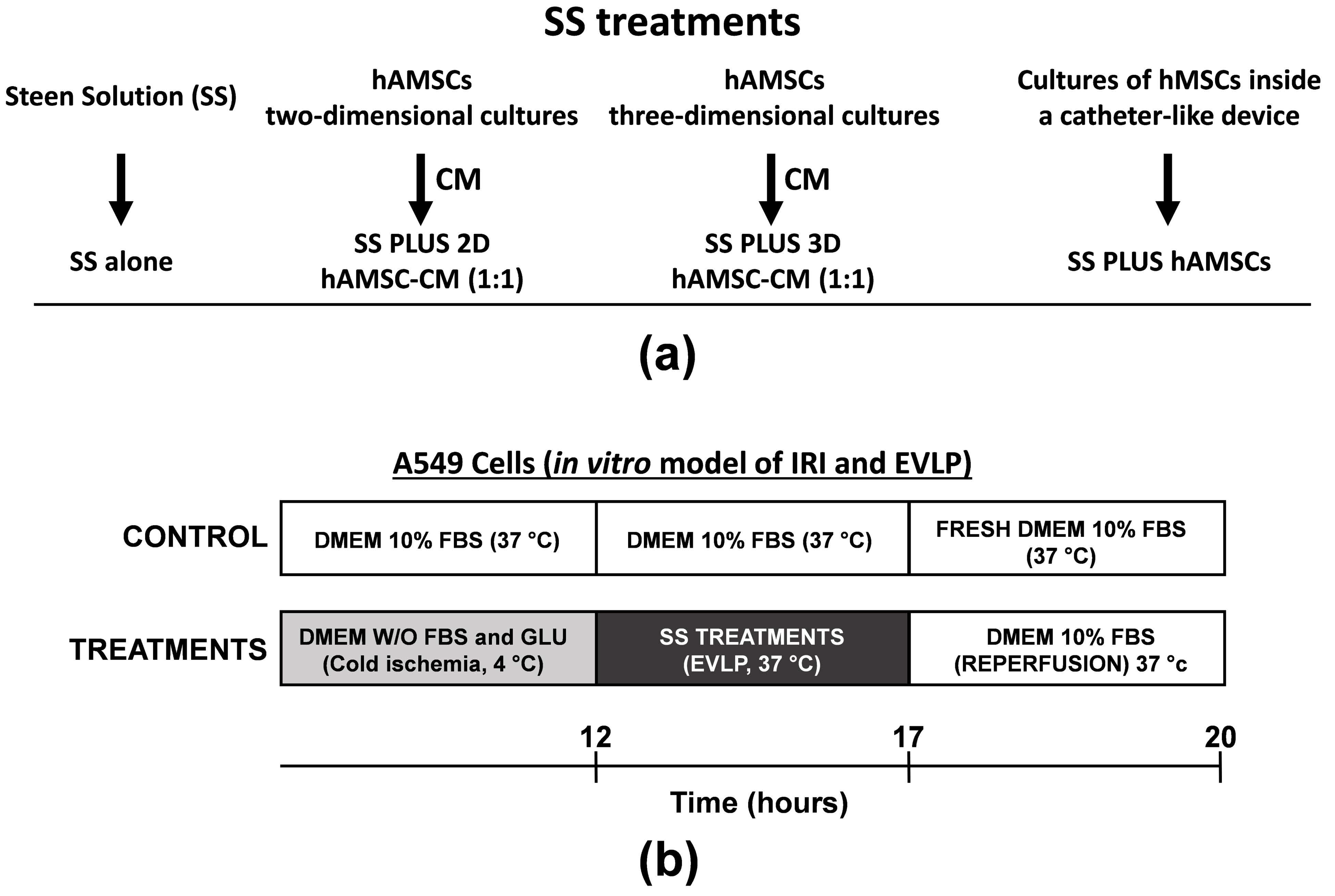
4.10. In Vitro Protocol of Cold Ischemia, Normothermic Recovery (EVLP), and Reperfusion
4.11. Analysis of Cell Death
4.12. Cell Cycle Analysis
4.13. Statistics
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| LTx | Lung transplantation |
| IRI | Ischemia-reperfusion injury |
| EVLP | Ex vivo lung perfusion |
| MSCs | Mesenchymal stromal/stem cells |
| hAMSCs | Human amnion-derived mesenchymal stromal/stem cells |
| CLAD | Chronic lung allograft dysfunction |
| PGD | Primary lung graft dysfunction |
| ALI | Acute lung injury |
| CM | Conditioned medium |
| PD-MSCs | Placenta-derived mesenchymal stromal/stem cells |
| 2D | Two-dimensional |
| 3D | Three-dimensional |
| LDH | Lactate dehydrogenase |
| PI | Propidium iodide |
| NRF2 | Nuclear factor erythroid 2-related factor 2 |
| 7AAD | 7-Aminoactinomycin D |
References
- Thabut, G.; Mal, H. Outcomes after lung transplantation. J. Thorac. Dis. 2017, 9, 2684–2691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bozso, S.; Vasanthan, V.; Luc, J.G.; Kinaschuk, K.; Freed, D.; Nagendran, J. Lung transplantation from donors after circulatory death using portable ex vivo lung perfusion. Can. Respir. J. 2015, 22, 47–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, F.; Date, H. Update on ischemia-reperfusion injury in lung transplantation. Curr. Opin. Organ Transplant. 2015, 20, 515–520. [Google Scholar] [CrossRef] [PubMed]
- Laubach, V.E.; Sharma, A.K. Mechanisms of lung ischemia-reperfusion injury. Curr. Opin. Organ Transplant. 2016, 21, 246–252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawashima, M.; Juvet, S.C. The role of innate immunity in the long-term outcome of lung transplantation. Ann. Transl. Med. 2020, 8, 412. [Google Scholar] [CrossRef] [PubMed]
- De Perrot, M.; Keshavjee, S. Lung transplantation. Lung preservation. Chest Surg. Clin. N. Am. 2003, 13, 443–462. [Google Scholar] [CrossRef]
- Charles, E.J.; Huerter, M.E.; Wagner, C.E.; Sharma, A.K.; Zhao, Y.; Stoler, M.H.; Mehaffey, J.H.; Isbell, J.M.; Lau, C.L.; Tribble, C.G.; et al. Donation After Circulatory Death Lungs Transplantable Up to Six Hours After Ex Vivo Lung Perfusion. Ann. Thorac. Surg. 2016, 102, 1845–1853. [Google Scholar] [CrossRef] [Green Version]
- Cypel, M.; Yeung, J.C.; Donahoe, L.; Chen, M.; Zamel, R.; Hoetzenecker, K.; Yasufuku, K.; de Perrot, M.; Pierre, A.F.; Waddell, T.K.; et al. Normothermic ex vivo lung perfusion: Does the indication impact organ utilization and patient outcomes after transplantation? J. Thorac. Cardiovasc. Surg. 2019, 159, 346–355.e1. [Google Scholar] [CrossRef] [Green Version]
- Huerter, M.E.; Sharma, A.K.; Zhao, Y.; Charles, E.J.; Kron, I.L.; Laubach, V.E. Attenuation of Pulmonary Ischemia-Reperfusion Injury by Adenosine A2B Receptor Antagonism. Ann. Thorac. Surg. 2016, 102, 385–393. [Google Scholar] [CrossRef] [Green Version]
- Martens, A.; Boada, M.; Vanaudenaerde, B.M.; Verleden, S.E.; Vos, R.; Verleden, G.M.; Verbeken, E.K.; Van Raemdonck, D.; Schols, D.; Claes, S.; et al. Steroids can reduce warm ischemic reperfusion injury in a porcine donation after circulatory death model with ex vivo lung perfusion evaluation. Transpl. Int. Off. J. Eur. Soc. Organ Transplant. 2016, 29, 1237–1246. [Google Scholar] [CrossRef] [Green Version]
- Motoyama, H.; Chen, F.; Hijiya, K.; Kondo, T.; Ohsumi, A.; Yamada, T.; Sato, M.; Aoyama, A.; Bando, T.; Date, H. Plasmin administration during ex vivo lung perfusion ameliorates lung ischemia-reperfusion injury. J. Heart Lung Transplant. Off. Publ. Int. Soc. Heart Transplant. 2014, 33, 1093–1099. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmelzer, E.; Miceli, V.; Chinnici, C.M.; Bertani, A.; Gerlach, J.C. Effects of Mesenchymal Stem Cell Coculture on Human Lung Small Airway Epithelial Cells. BioMed Res. Int. 2020, 2020, 9847579. [Google Scholar] [CrossRef] [PubMed]
- Abreu, S.C.; Antunes, M.A.; Xisto, D.G.; Cruz, F.F.; Branco, V.C.; Bandeira, E.; Kitoko, J.Z.; de Araujo, A.F.; Dellatorre-Texeira, L.; Olsen, P.C.; et al. Bone Marrow, Adipose, and Lung Tissue-Derived Murine Mesenchymal Stromal Cells Release Different Mediators and Differentially Affect Airway and Lung Parenchyma in Experimental Asthma. Stem Cells Transl. Med. 2017, 6, 1557–1567. [Google Scholar] [CrossRef] [PubMed]
- Morrison, T.J.; Jackson, M.V.; Cunningham, E.K.; Kissenpfennig, A.; McAuley, D.F.; O’Kane, C.M.; Krasnodembskaya, A.D. Mesenchymal Stromal Cells Modulate Macrophages in Clinically Relevant Lung Injury Models by Extracellular Vesicle Mitochondrial Transfer. Am. J. Respir. Crit. Care Med. 2017, 196, 1275–1286. [Google Scholar] [CrossRef] [PubMed]
- Marrazzo, P.; Crupi, A.N.; Alviano, F.; Teodori, L.; Bonsi, L. Exploring the roles of MSCs in infections: Focus on bacterial diseases. J. Mol. Med. 2019, 97, 437–450. [Google Scholar] [CrossRef]
- Sun, C.K.; Yen, C.H.; Lin, Y.C.; Tsai, T.H.; Chang, L.T.; Kao, Y.H.; Chua, S.; Fu, M.; Ko, S.F.; Leu, S.; et al. Autologous transplantation of adipose-derived mesenchymal stem cells markedly reduced acute ischemia-reperfusion lung injury in a rodent model. J. Transl. Med. 2011, 9, 118. [Google Scholar] [CrossRef] [Green Version]
- Tian, W.; Liu, Y.; Zhang, B.; Dai, X.; Li, G.; Li, X.; Zhang, Z.; Du, C.; Wang, H. Infusion of mesenchymal stem cells protects lung transplants from cold ischemia-reperfusion injury in mice. Lung 2015, 193, 85–95. [Google Scholar] [CrossRef]
- Wittwer, T.; Rahmanian, P.; Choi, Y.H.; Zeriouh, M.; Karavidic, S.; Neef, K.; Christmann, A.; Piatkowski, T.; Schnapper, A.; Ochs, M.; et al. Mesenchymal stem cell pretreatment of non-heart-beating-donors in experimental lung transplantation. J. Cardiothorac. Surg. 2014, 9, 151. [Google Scholar] [CrossRef] [Green Version]
- La Francesca, S.; Ting, A.E.; Sakamoto, J.; Rhudy, J.; Bonenfant, N.R.; Borg, Z.D.; Cruz, F.F.; Goodwin, M.; Lehman, N.A.; Taggart, J.M.; et al. Multipotent adult progenitor cells decrease cold ischemic injury in ex vivo perfused human lungs: An initial pilot and feasibility study. Transplant. Res. 2014, 3, 19. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.W.; Fang, X.; Gupta, N.; Serikov, V.; Matthay, M.A. Allogeneic human mesenchymal stem cells for treatment of E. coli endotoxin-induced acute lung injury in the ex vivo perfused human lung. Proc. Natl. Acad. Sci. USA 2009, 106, 16357–16362. [Google Scholar] [CrossRef] [Green Version]
- Mordant, P.; Nakajima, D.; Kalaf, R.; Iskender, I.; Maahs, L.; Behrens, P.; Coutinho, R.; Iyer, R.K.; Davies, J.E.; Cypel, M.; et al. Mesenchymal stem cell treatment is associated with decreased perfusate concentration of interleukin-8 during ex vivo perfusion of donor lungs after 18-hour preservation. J. Heart Lung Transplant. Off. Publ. Int. Soc. Heart Transplant. 2016, 35, 1245–1254. [Google Scholar] [CrossRef] [PubMed]
- Walter, S.G.; Randau, T.M.; Hilgers, C.; Haddouti, E.M.; Masson, W.; Gravius, S.; Burger, C.; Wirtz, D.C.; Schildberg, F.A. Molecular and Functional Phenotypes of Human Bone Marrow-Derived Mesenchymal Stromal Cells Depend on Harvesting Techniques. Int. J. Mol. Sci. 2020, 21, 4382. [Google Scholar] [CrossRef] [PubMed]
- Burja, B.; Barlic, A.; Erman, A.; Mrak-Poljsak, K.; Tomsic, M.; Sodin-Semrl, S.; Lakota, K. Human mesenchymal stromal cells from different tissues exhibit unique responses to different inflammatory stimuli. Curr. Res. Transl. Med. 2020, 68, 217–224. [Google Scholar] [CrossRef] [PubMed]
- Tesarova, L.; Jaresova, K.; Simara, P.; Koutna, I. Umbilical Cord-Derived Mesenchymal Stem Cells Are Able to Use bFGF Treatment and Represent a Superb Tool for Immunosuppressive Clinical Applications. Int. J. Mol. Sci. 2020, 21, 5366. [Google Scholar] [CrossRef] [PubMed]
- Parolini, O.; Alviano, F.; Bagnara, G.P.; Bilic, G.; Buhring, H.J.; Evangelista, M.; Hennerbichler, S.; Liu, B.; Magatti, M.; Mao, N.; et al. Concise review: Isolation and characterization of cells from human term placenta: Outcome of the first international Workshop on Placenta Derived Stem Cells. Stem Cells 2008, 26, 300–311. [Google Scholar] [CrossRef] [Green Version]
- Bulati, M.; Miceli, V.; Gallo, A.; Amico, G.; Carcione, C.; Pampalone, M.; Conaldi, P.G. The Immunomodulatory Properties of the Human Amnion-Derived Mesenchymal Stromal/Stem Cells Are Induced by INF-gamma Produced by Activated Lymphomonocytes and Are Mediated by Cell-To-Cell Contact and Soluble Factors. Front. Immunol. 2020, 11, 54. [Google Scholar] [CrossRef] [Green Version]
- Fan, X.L.; Zhang, Y.; Li, X.; Fu, Q.L. Mechanisms underlying the protective effects of mesenchymal stem cell-based therapy. Cell. Mol. Life Sci. 2020, 77, 2771–2794. [Google Scholar] [CrossRef] [Green Version]
- Miceli, V.; Pampalone, M.; Vella, S.; Carreca, A.P.; Amico, G.; Conaldi, P.G. Comparison of Immunosuppressive and Angiogenic Properties of Human Amnion-Derived Mesenchymal Stem Cells between 2D and 3D Culture Systems. Stem Cells Int. 2019, 2019, 7486279. [Google Scholar] [CrossRef]
- Stavely, R.; Nurgali, K. The emerging antioxidant paradigm of mesenchymal stem cell therapy. Stem Cells Transl. Med. 2020, 9, 985–1006. [Google Scholar] [CrossRef]
- Arrigoni, C.; D’Arrigo, D.; Rossella, V.; Candrian, C.; Albertini, V.; Moretti, M. Umbilical Cord MSCs and Their Secretome in the Therapy of Arthritic Diseases: A Research and Industrial Perspective. Cells 2020, 9, 1343. [Google Scholar] [CrossRef]
- Maleki, S.N.; Aboutaleb, N.; Nazarinia, D.; Allahverdi Beik, S.; Qolamian, A.; Nobakht, M. Conditioned medium obtained from human amniotic membrane-derived mesenchymal stem cell attenuates heart failure injury in rats. Iran. J. Basic Med Sci. 2019, 22, 1253–1258. [Google Scholar]
- Ertl, J.; Pichlsberger, M.; Tuca, A.C.; Wurzer, P.; Fuchs, J.; Geyer, S.H.; Maurer-Gesek, B.; Weninger, W.J.; Pfeiffer, D.; Bubalo, V.; et al. Comparative study of regenerative effects of mesenchymal stem cells derived from placental amnion, chorion and umbilical cord on dermal wounds. Placenta 2018, 65, 37–46. [Google Scholar] [CrossRef] [PubMed]
- Shu, J.; He, X.; Li, H.; Liu, X.; Qiu, X.; Zhou, T.; Wang, P.; Huang, X. The Beneficial Effect of Human Amnion Mesenchymal Cells in Inhibition of Inflammation and Induction of Neuronal Repair in EAE Mice. J. Immunol. Res. 2018, 2018, 5083797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tuca, A.C.; Ertl, J.; Hingerl, K.; Pichlsberger, M.; Fuchs, J.; Wurzer, P.; Pfeiffer, D.; Bubalo, V.; Parvizi, D.; Kamolz, L.P.; et al. Comparison of Matrigel and Matriderm as a carrier for human amnion-derived mesenchymal stem cells in wound healing. Placenta 2016, 48, 99–103. [Google Scholar] [CrossRef]
- Miceli, V.; Chinnici, C.M.; Bulati, M.; Pampalone, M.; Amico, G.; Schmelzer, E.; Gerlach, J.C.; Conaldi, P.G. Comparative study of the production of soluble factors in human placenta-derived mesenchymal stromal/stem cells grown in adherent conditions or as aggregates in a catheter-like device. Biochem. Biophys. Res. Commun. 2020, 522, 171–176. [Google Scholar] [CrossRef]
- Silini, A.; Parolini, O.; Huppertz, B.; Lang, I. Soluble factors of amnion-derived cells in treatment of inflammatory and fibrotic pathologies. Curr. Stem Cell Res. Ther. 2013, 8, 6–14. [Google Scholar] [CrossRef] [Green Version]
- Danieli, P.; Malpasso, G.; Ciuffreda, M.C.; Cervio, E.; Calvillo, L.; Copes, F.; Pisano, F.; Mura, M.; Kleijn, L.; de Boer, R.A.; et al. Conditioned medium from human amniotic mesenchymal stromal cells limits infarct size and enhances angiogenesis. Stem Cells Transl. Med. 2015, 4, 448–458. [Google Scholar] [CrossRef]
- Cargnoni, A.; Ressel, L.; Rossi, D.; Poli, A.; Arienti, D.; Lombardi, G.; Parolini, O. Conditioned medium from amniotic mesenchymal tissue cells reduces progression of bleomycin-induced lung fibrosis. Cytotherapy 2012, 14, 153–161. [Google Scholar] [CrossRef] [Green Version]
- Rossi, D.; Pianta, S.; Magatti, M.; Sedlmayr, P.; Parolini, O. Characterization of the conditioned medium from amniotic membrane cells: Prostaglandins as key effectors of its immunomodulatory activity. PLoS ONE 2012, 7, e46956. [Google Scholar] [CrossRef]
- Bartosh, T.J.; Ylostalo, J.H.; Mohammadipoor, A.; Bazhanov, N.; Coble, K.; Claypool, K.; Lee, R.H.; Choi, H.; Prockop, D.J. Aggregation of human mesenchymal stromal cells (MSCs) into 3D spheroids enhances their antiinflammatory properties. Proc. Natl. Acad. Sci. USA 2010, 107, 13724–13729. [Google Scholar] [CrossRef] [Green Version]
- Ylostalo, J.H.; Bartosh, T.J.; Coble, K.; Prockop, D.J. Human mesenchymal stem/stromal cells cultured as spheroids are self-activated to produce prostaglandin E2 that directs stimulated macrophages into an anti-inflammatory phenotype. Stem Cells 2012, 30, 2283–2296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Francesco, F.; Tirino, V.; Desiderio, V.; Ferraro, G.; D’Andrea, F.; Giuliano, M.; Libondi, G.; Pirozzi, G.; De Rosa, A.; Papaccio, G. Human CD34/CD90 ASCs are capable of growing as sphere clusters, producing high levels of VEGF and forming capillaries. PLoS ONE 2009, 4, e6537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, E.J.; Park, S.J.; Kang, S.K.; Kim, G.H.; Kang, H.J.; Lee, S.W.; Jeon, H.B.; Kim, H.S. Spherical bullet formation via E-cadherin promotes therapeutic potency of mesenchymal stem cells derived from human umbilical cord blood for myocardial infarction. Mol. Ther. J. Am. Soc. Gene Ther. 2012, 20, 1424–1433. [Google Scholar] [CrossRef] [Green Version]
- De Perrot, M.; Liu, M.; Waddell, T.K.; Keshavjee, S. Ischemia-reperfusion-induced lung injury. Am. J. Respir. Crit. Care Med. 2003, 167, 490–511. [Google Scholar] [CrossRef] [PubMed]
- Tabarelli, W.; Bonatti, H.; Tabarelli, D.; Eller, M.; Muller, L.; Ruttmann, E.; Lass-Florl, C.; Larcher, C.; Geltner, C. Long term complications following 54 consecutive lung transplants. J. Thorac. Dis. 2016, 8, 1234–1244. [Google Scholar] [CrossRef] [Green Version]
- Christie, J.D.; Carby, M.; Bag, R.; Corris, P.; Hertz, M.; Weill, D.; ISHLT Working Group on Primary Lung Graft Dysfunction. Report of the ISHLT Working Group on Primary Lung Graft Dysfunction part II: Definition. A consensus statement of the International Society for Heart and Lung Transplantation. J. Heart Lung Transplant. Off. Publ. Int. Soc. Heart Transplant. 2005, 24, 1454–1459. [Google Scholar] [CrossRef]
- Kalogeris, T.; Baines, C.P.; Krenz, M.; Korthuis, R.J. Ischemia/Reperfusion. Compr. Physiol. 2016, 7, 113–170. [Google Scholar] [CrossRef] [Green Version]
- Carden, D.L.; Granger, D.N. Pathophysiology of ischaemia-reperfusion injury. J. Pathol. 2000, 190, 255–266. [Google Scholar] [CrossRef]
- Behnke, J.; Kremer, S.; Shahzad, T.; Chao, C.M.; Bottcher-Friebertshauser, E.; Morty, R.E.; Bellusci, S.; Ehrhardt, H. MSC Based Therapies—New Perspectives for the Injured Lung. J. Clin. Med. 2020, 9, 682. [Google Scholar] [CrossRef] [Green Version]
- Qin, H.; Zhao, A. Mesenchymal stem cell therapy for acute respiratory distress syndrome: From basic to clinics. Protein Cell 2020, 11, 707–722. [Google Scholar] [CrossRef]
- Lin, K.C.; Yip, H.K.; Shao, P.L.; Wu, S.C.; Chen, K.H.; Chen, Y.T.; Yang, C.C.; Sun, C.K.; Kao, G.S.; Chen, S.Y.; et al. Combination of adipose-derived mesenchymal stem cells (ADMSC) and ADMSC-derived exosomes for protecting kidney from acute ischemia-reperfusion injury. Int. J. Cardiol. 2016, 216, 173–185. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.S.; Choi, S.J.; Ahn, S.Y.; Sung, D.K.; Sung, S.I.; Yoo, H.S.; Oh, W.I.; Park, W.S. Timing of umbilical cord blood derived mesenchymal stem cells transplantation determines therapeutic efficacy in the neonatal hyperoxic lung injury. PLoS ONE 2013, 8, e52419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, H.; Hu, J.; Yan, Q.; Zhu, J.; Zhu, Z.; Chen, Y.; Sun, J.; Zhang, R. Bone marrow-derived mesenchymal stem cells rescue injured H9c2 cells via transferring intact mitochondria through tunneling nanotubes in an in vitro simulated ischemia/reperfusion model. Mol. Med. Rep. 2016, 13, 1517–1524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, M.; Jue, S.S.; Cho, Y.A.; Kim, E.C. Comparison of the effects of human dental pulp stem cells and human bone marrow-derived mesenchymal stem cells on ischemic human astrocytes in vitro. J. Neurosci. Res. 2015, 93, 973–983. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, T.; Hoshikawa, Y.; Ishibashi, N.; Suzuki, H.; Notsuda, H.; Watanabe, Y.; Noda, M.; Kanehira, M.; Ohkouchi, S.; Kondo, T.; et al. Mesenchymal stem cells attenuate ischemia-reperfusion injury after prolonged cold ischemia in a mouse model of lung transplantation: A preliminary study. Surg. Today 2017, 47, 425–431. [Google Scholar] [CrossRef]
- Martens, A.; Ordies, S.; Vanaudenaerde, B.M.; Verleden, S.E.; Vos, R.; Van Raemdonck, D.E.; Verleden, G.M.; Roobrouck, V.D.; Claes, S.; Schols, D.; et al. Immunoregulatory effects of multipotent adult progenitor cells in a porcine ex vivo lung perfusion model. Stem Cell Res. Ther. 2017, 8, 159. [Google Scholar] [CrossRef] [Green Version]
- Haarer, J.; Johnson, C.L.; Soeder, Y.; Dahlke, M.H. Caveats of mesenchymal stem cell therapy in solid organ transplantation. Transpl. Int. Off. J. Eur. Soc. Organ Transplant. 2015, 28, 1–9. [Google Scholar] [CrossRef]
- Lalu, M.M.; McIntyre, L.; Pugliese, C.; Fergusson, D.; Winston, B.W.; Marshall, J.C.; Granton, J.; Stewart, D.J. Canadian Critical Care Trials Group. Safety of cell therapy with mesenchymal stromal cells (SafeCell): A systematic review and meta-analysis of clinical trials. PLoS ONE 2012, 7, e47559. [Google Scholar] [CrossRef]
- Akyurekli, C.; Le, Y.; Richardson, R.B.; Fergusson, D.; Tay, J.; Allan, D.S. A systematic review of preclinical studies on the therapeutic potential of mesenchymal stromal cell-derived microvesicles. Stem Cell Rev. Rep. 2015, 11, 150–160. [Google Scholar] [CrossRef]
- Xagorari, A.; Siotou, E.; Yiangou, M.; Tsolaki, E.; Bougiouklis, D.; Sakkas, L.; Fassas, A.; Anagnostopoulos, A. Protective effect of mesenchymal stem cell-conditioned medium on hepatic cell apoptosis after acute liver injury. Int. J. Clin. Exp. Pathol. 2013, 6, 831–840. [Google Scholar]
- Hwang, B.; Liles, W.C.; Waworuntu, R.; Mulligan, M.S. Pretreatment with bone marrow-derived mesenchymal stromal cell-conditioned media confers pulmonary ischemic tolerance. J. Thorac. Cardiovasc. Surg. 2016, 151, 841–849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliva, J. Therapeutic Properties of Mesenchymal Stem Cell on Organ Ischemia-Reperfusion Injury. Int. J. Mol. Sci. 2019, 20, 5511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, C.; Fu, W.L.; Zhang, D.D.; Xing, W.W.; Xia, W.R.; Wei, Z.; Zou, M.J.; Zhu, X.M.; Xu, D.G. The protective role of IL-1Ra on intestinal ischemia reperfusion injury by anti-oxidative stress via Nrf2/HO-1 pathway in rat. Biomed. J. 2019, 42, 36–45. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Tian, S.F.; Guo, Y.; Niu, X.; Hu, B.; Guo, S.C.; Wang, N.S.; Wang, Y. Transplantation of induced pluripotent stem cell-derived renal stem cells improved acute kidney injury. Cell Biosci. 2015, 5, 45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toyohara, T.; Mae, S.; Sueta, S.; Inoue, T.; Yamagishi, Y.; Kawamoto, T.; Kasahara, T.; Hoshina, A.; Toyoda, T.; Tanaka, H.; et al. Cell Therapy Using Human Induced Pluripotent Stem Cell-Derived Renal Progenitors Ameliorates Acute Kidney Injury in Mice. Stem Cells Transl. Med. 2015, 4, 980–992. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Zhang, J.; Song, Y.; Yang, P.; Yang, Y.; Huang, Z.; Wang, K. Deficiency of anti-inflammatory cytokine IL-4 leads to neural hyperexcitability and aggravates cerebral ischemia-reperfusion injury. Acta Pharm. Sin. B 2020, 10, 1634–1645. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, F.; Nagai, N.; Tang, Z.; Zhang, S.; Scotney, P.; Lennartsson, J.; Zhu, C.; Qu, Y.; Fang, C.; et al. VEGF-B inhibits apoptosis via VEGFR-1-mediated suppression of the expression of BH3-only protein genes in mice and rats. J. Clin. Investig. 2008, 118, 913–923. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Cao, C.; Xin, J.; Lv, P.; Chen, D.; Li, S.; Yang, H.; Chen, C.; Liu, B.; Li, Q. Treatment with placental growth factor attenuates myocardial ischemia/reperfusion injury. PLoS ONE 2018, 13, e0202772. [Google Scholar] [CrossRef]
- Papiewska-Pajak, I.; Balcerczyk, A.; Stec-Martyna, E.; Koziolkiewicz, W.; Boncela, J. Vascular endothelial growth factor-D modulates oxidant-antioxidant balance of human vascular endothelial cells. J. Cell. Mol. Med. 2017, 21, 1139–1149. [Google Scholar] [CrossRef] [Green Version]
- Zouein, F.A.; Kurdi, M.; Booz, G.W. LIF and the heart: Just another brick in the wall? Eur. Cytokine Netw. 2013, 24, 11–19. [Google Scholar] [CrossRef]
- Ege, T.; Us, M.H.; Sungun, M.; Duran, E. Cytokine response in lower extremity ischaemia/reperfusion. J. Int. Med. Res. 2004, 32, 124–131. [Google Scholar] [CrossRef] [Green Version]
- Sung, F.L.; Zhu, T.Y.; Au-Yeung, K.K.; Siow, Y.L.; Karmin, O. Enhanced MCP-1 expression during ischemia/reperfusion injury is mediated by oxidative stress and NF-kappaB. Kidney Int. 2002, 62, 1160–1170. [Google Scholar] [CrossRef] [PubMed]
- Evangelista, A.F.; Vannier-Santos, M.A.; de Assis Silva, G.S.; Silva, D.N.; Juiz, P.J.L.; Nonaka, C.K.V.; Dos Santos, R.R.; Soares, M.B.P.; Villarreal, C.F. Bone marrow-derived mesenchymal stem/stromal cells reverse the sensorial diabetic neuropathy via modulation of spinal neuroinflammatory cascades. J. Neuroinflamm. 2018, 15, 189. [Google Scholar] [CrossRef] [PubMed]
- Leonard, M.O.; Kieran, N.E.; Howell, K.; Burne, M.J.; Varadarajan, R.; Dhakshinamoorthy, S.; Porter, A.G.; O’Farrelly, C.; Rabb, H.; Taylor, C.T. Reoxygenation-specific activation of the antioxidant transcription factor Nrf2 mediates cytoprotective gene expression in ischemia-reperfusion injury. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2006, 20, 2624–2626. [Google Scholar] [CrossRef]
- Schmelzer, E.; McKeel, D.T.; Gerlach, J.C. Characterization of Human Mesenchymal Stem Cells from Different Tissues and Their Membrane Encasement for Prospective Transplantation Therapies. BioMed Res. Int. 2019, 2019, 6376271. [Google Scholar] [CrossRef] [PubMed]
- Cypel, M.; Yeung, J.C.; Liu, M.; Anraku, M.; Chen, F.; Karolak, W.; Sato, M.; Laratta, J.; Azad, S.; Madonik, M.; et al. Normothermic ex vivo lung perfusion in clinical lung transplantation. N. Engl. J. Med. 2011, 364, 1431–1440. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward (5′-3′) | Reverse (5′-3′) | GenBank Accession ID | Amplicon Length (bp) |
|---|---|---|---|---|
| NF-kB | TACTCTGGCGCAGAAATTAGGTC | CTGTCTCGGAGCTCGTCTATTTG | NM_003998.3 | 263 |
| BCL2 | ATCGCCCTGTGGATGACTGAG | CAGCCAGGAGAAATCAAACAGAGG | NM_000633.2 | 128 |
| IL-1β | TCCAGGGACAGGATATGGAG | TCTTTCAACACGCAGGACAG | NM_000576.2 | 132 |
| IL-4 | CTTTGCTGCCTCCAAGAACAC | GCGAGTGTCCTTCTCATGGT | NM_000589.3 | 96 |
| IL-6 | TGTGAAAGCAGCAAAGAGGC | TGGGTCAGGGGTGGTTATT | NM_000600.4 | 275 |
| IL-8 | CCACCGGAGCACTCCATAAG | GATGGTTCCTTCCGGTGGTT | NM_000584.3 | 96 |
| CCL2 | CGCGAGCTATAGAAGAATCAC | TTGGGTTGTGGAGTGAGTGT | NM_002982.3 | 176 |
| ALDH1A1 | CAAGATCCAGGGCCGTACAA | GGAGGAAACCCTGCCTCTTTT | NM_000689.4 | 233 |
| GSTP | CCCTACACCGTGGTCTATTTCC | GAGGCTTTGAGTGAGCCCT | NM_000852.3 | 133 |
| GAPDH | TCAAGAAGGTGGTGAAGCAGG | ACCAGGAAATGAGCTTGACAAA | NM_002046.6 | 167 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miceli, V.; Bertani, A.; Chinnici, C.M.; Bulati, M.; Pampalone, M.; Amico, G.; Carcione, C.; Schmelzer, E.; Gerlach, J.C.; Conaldi, P.G. Conditioned Medium from Human Amnion-Derived Mesenchymal Stromal/Stem Cells Attenuating the Effects of Cold Ischemia-Reperfusion Injury in an In Vitro Model Using Human Alveolar Epithelial Cells. Int. J. Mol. Sci. 2021, 22, 510. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22020510
Miceli V, Bertani A, Chinnici CM, Bulati M, Pampalone M, Amico G, Carcione C, Schmelzer E, Gerlach JC, Conaldi PG. Conditioned Medium from Human Amnion-Derived Mesenchymal Stromal/Stem Cells Attenuating the Effects of Cold Ischemia-Reperfusion Injury in an In Vitro Model Using Human Alveolar Epithelial Cells. International Journal of Molecular Sciences. 2021; 22(2):510. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22020510
Chicago/Turabian StyleMiceli, Vitale, Alessandro Bertani, Cinzia Maria Chinnici, Matteo Bulati, Mariangela Pampalone, Giandomenico Amico, Claudia Carcione, Eva Schmelzer, Jörg C. Gerlach, and Pier Giulio Conaldi. 2021. "Conditioned Medium from Human Amnion-Derived Mesenchymal Stromal/Stem Cells Attenuating the Effects of Cold Ischemia-Reperfusion Injury in an In Vitro Model Using Human Alveolar Epithelial Cells" International Journal of Molecular Sciences 22, no. 2: 510. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22020510