Transcriptome Analysis and Identification of Lipid Genes in Physaria lindheimeri, a Genetic Resource for Hydroxy Fatty Acids in Seed Oil

 , and
, and 
Abstract
:1. Introduction
2. Results and Discussion
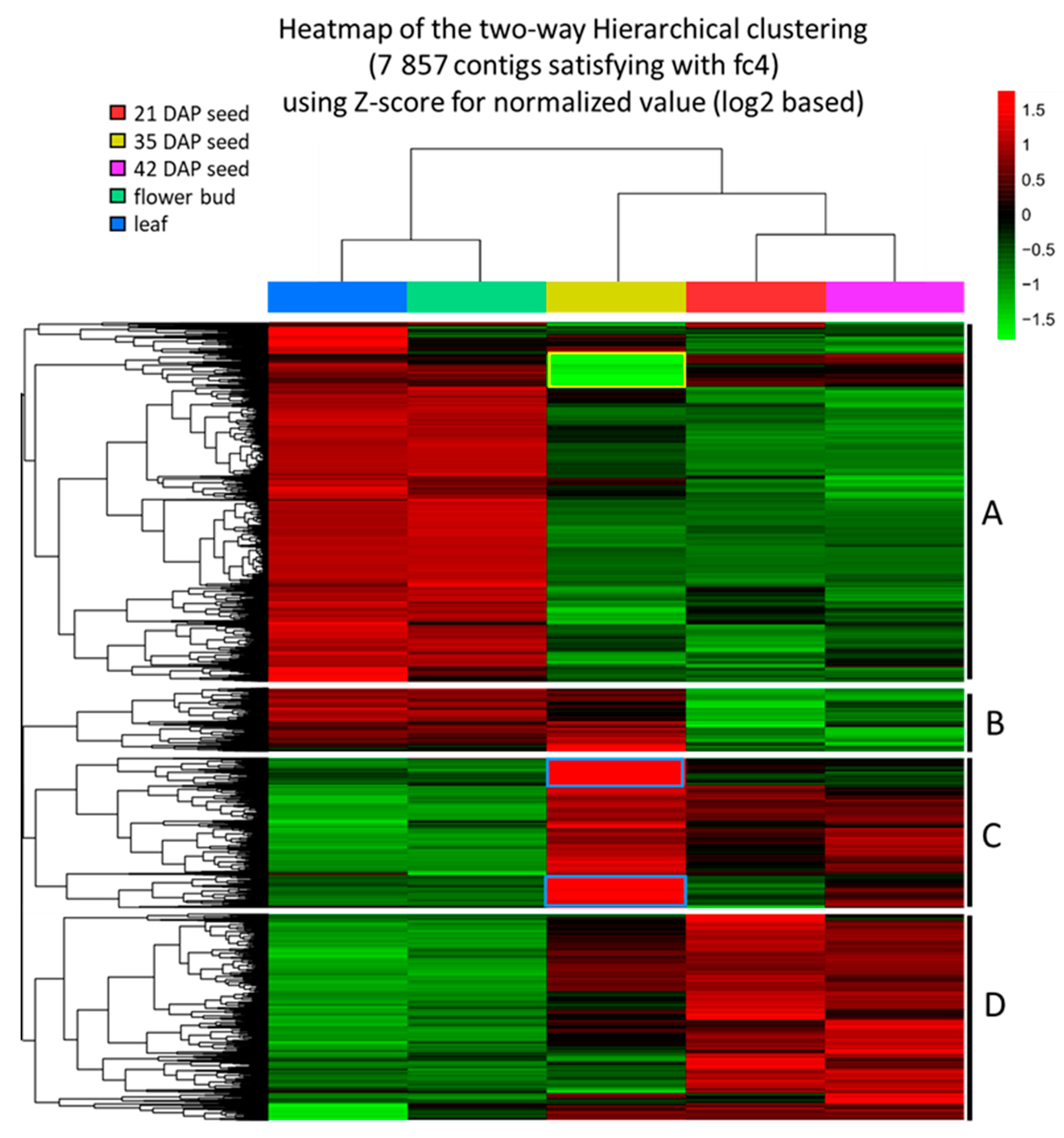
2.1. Analysis of Transcriptomes, Gene Ontology and Differentially Expressed Genes (DEGs)
2.2. Mining and Characterization of Genes Involved in FA and TAG Biosynthesis during P. lindheimeri Seed Development
2.3. De novo Fatty Acid Biosynthesis in Plastids
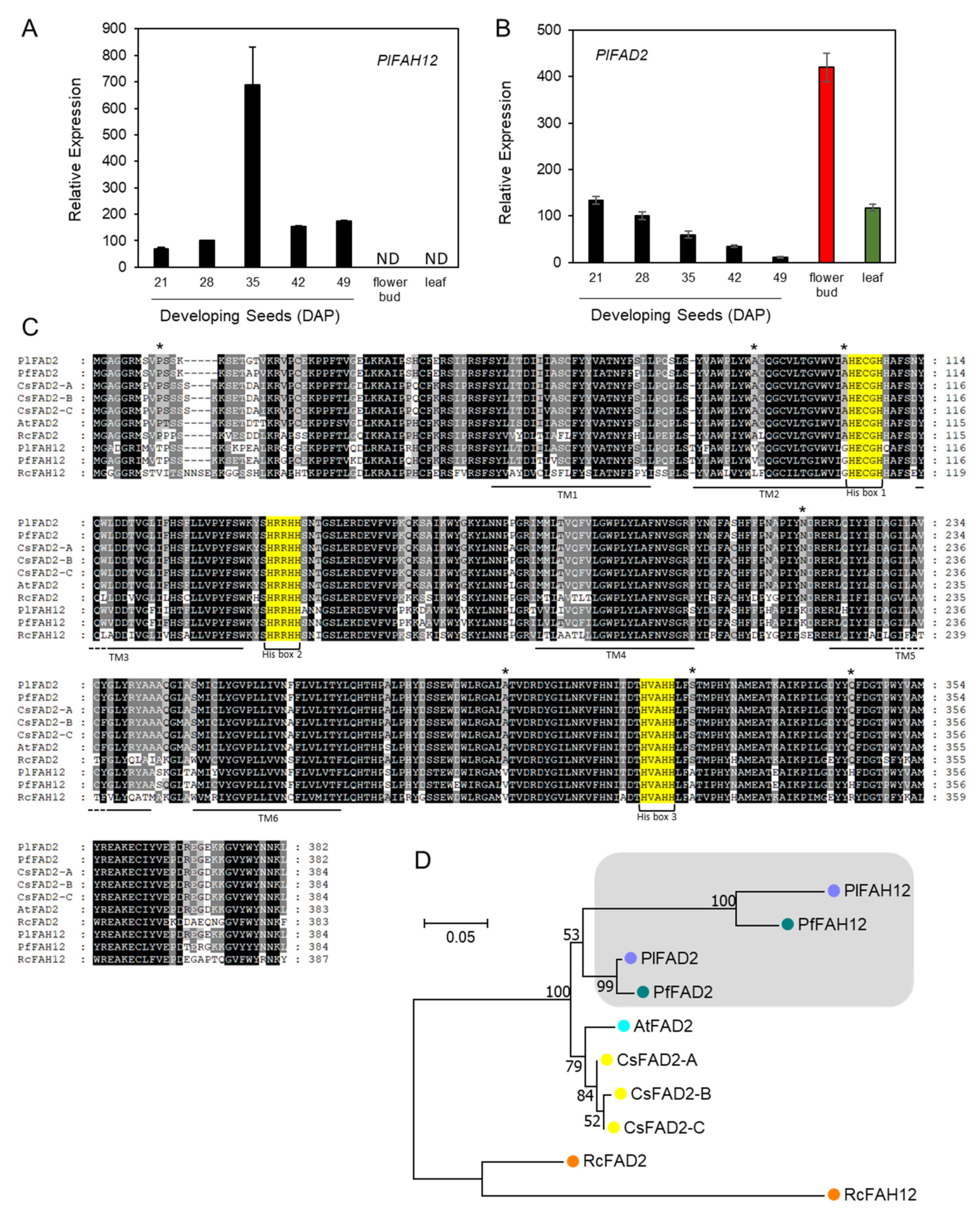
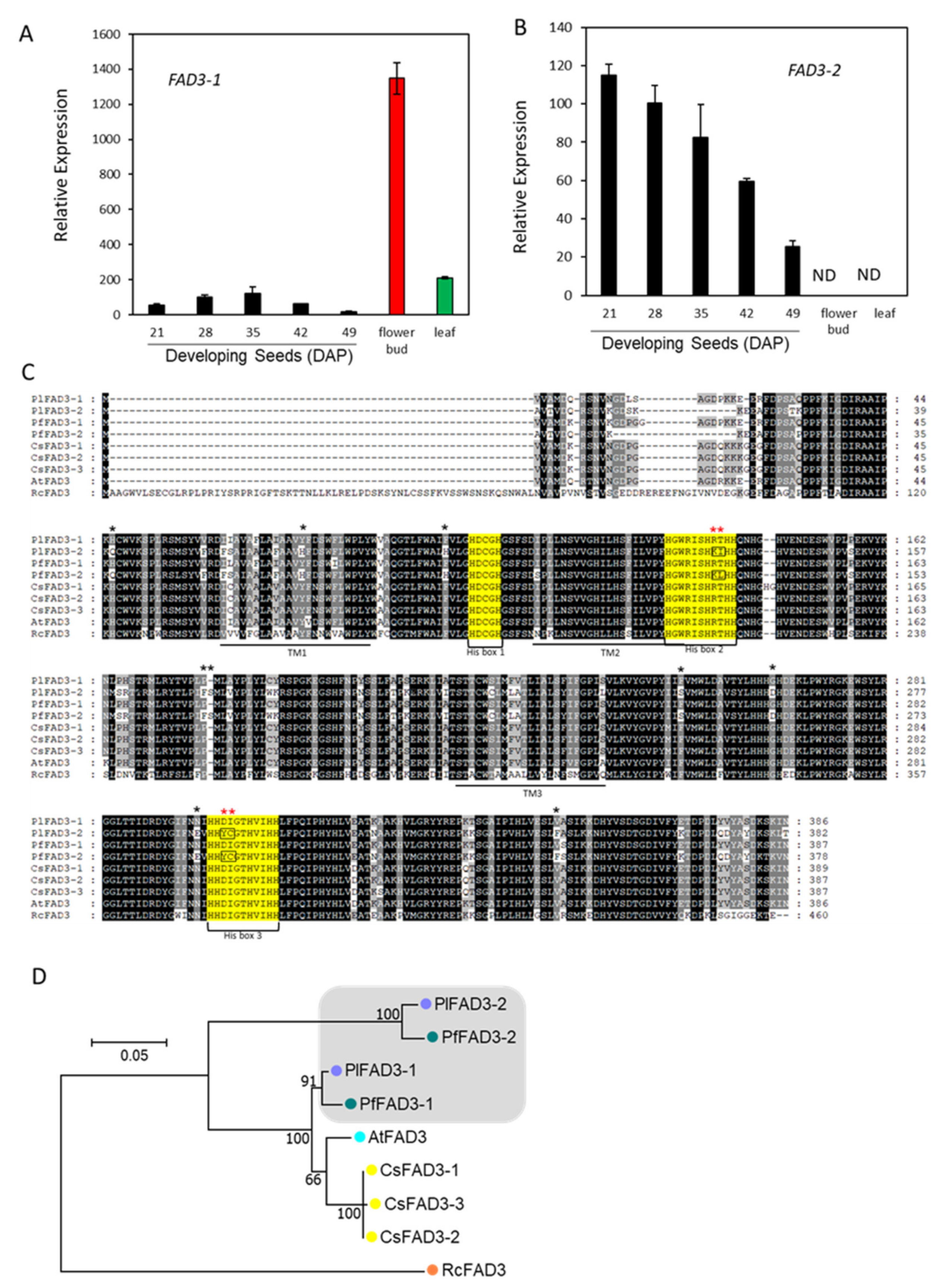
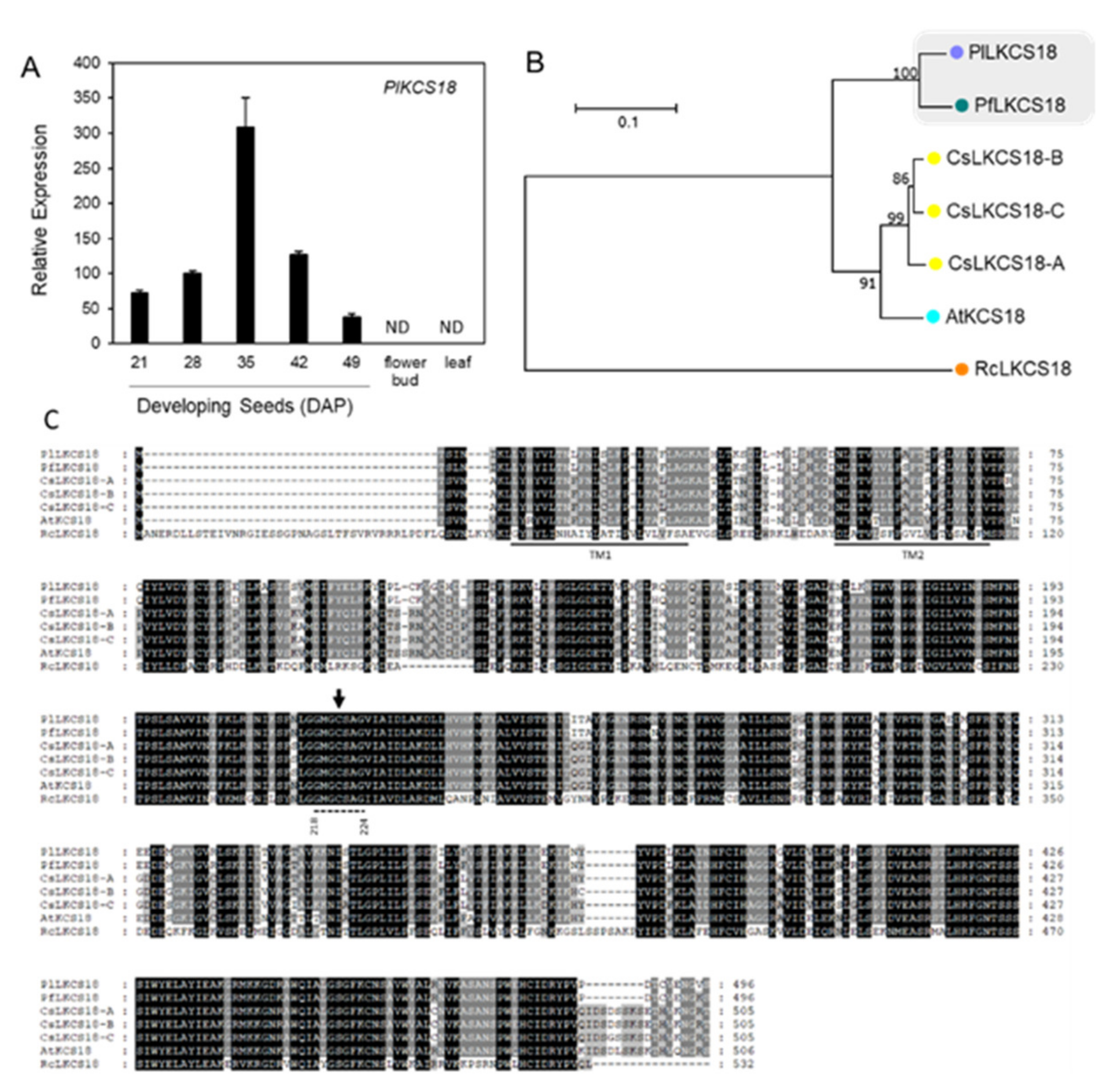
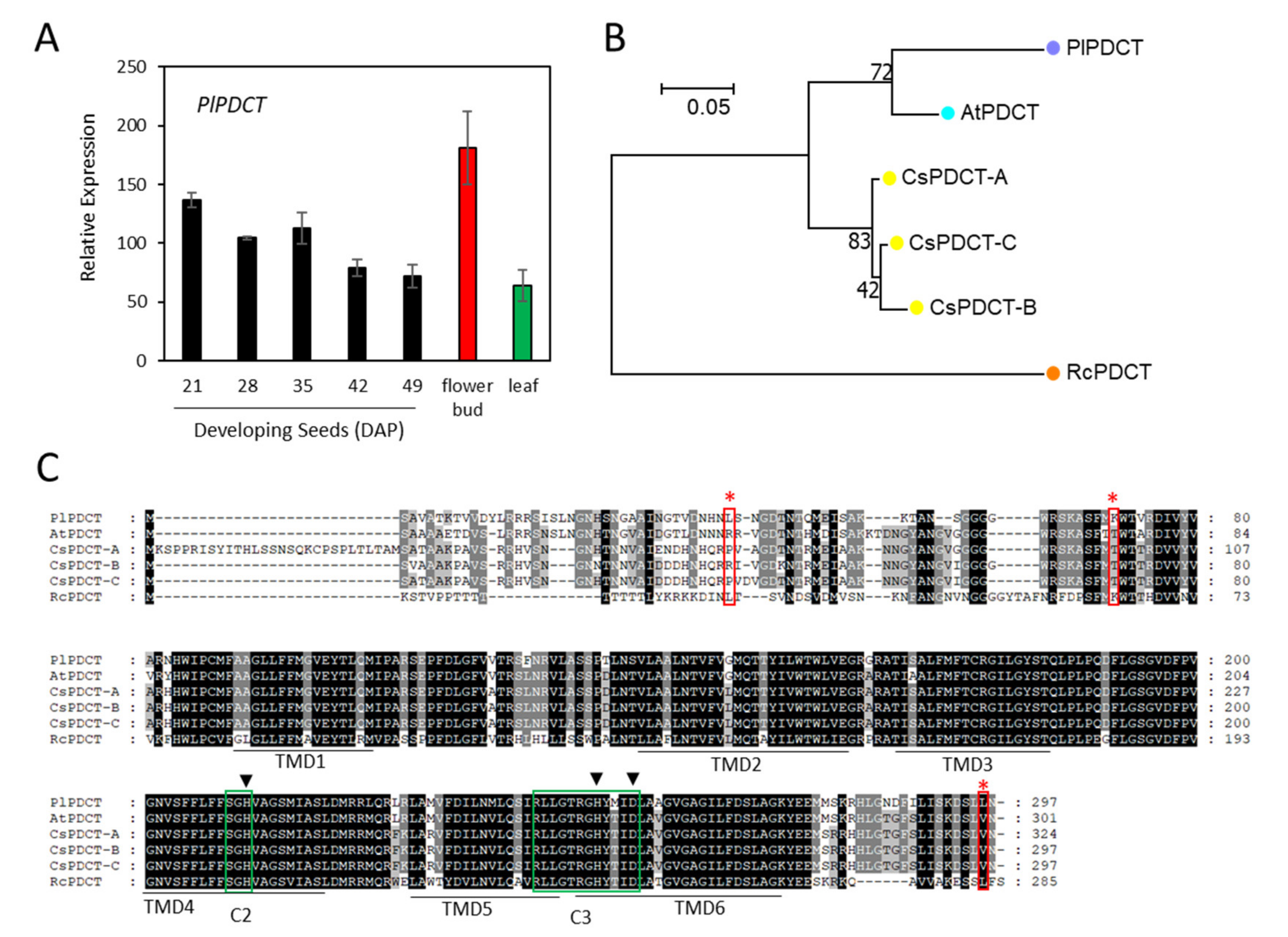
2.4. Hydroxylase and ER-Associated FA Desaturases
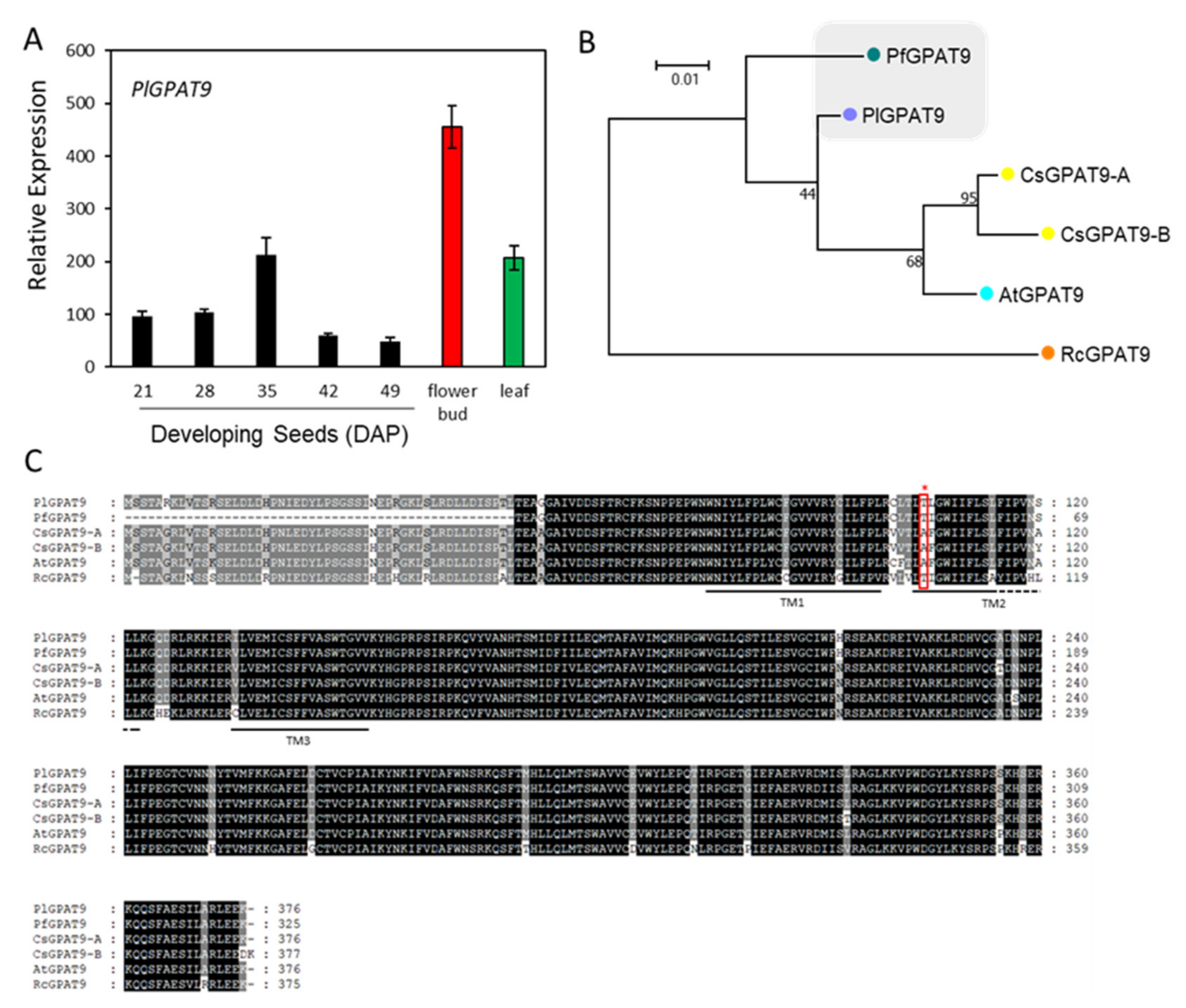
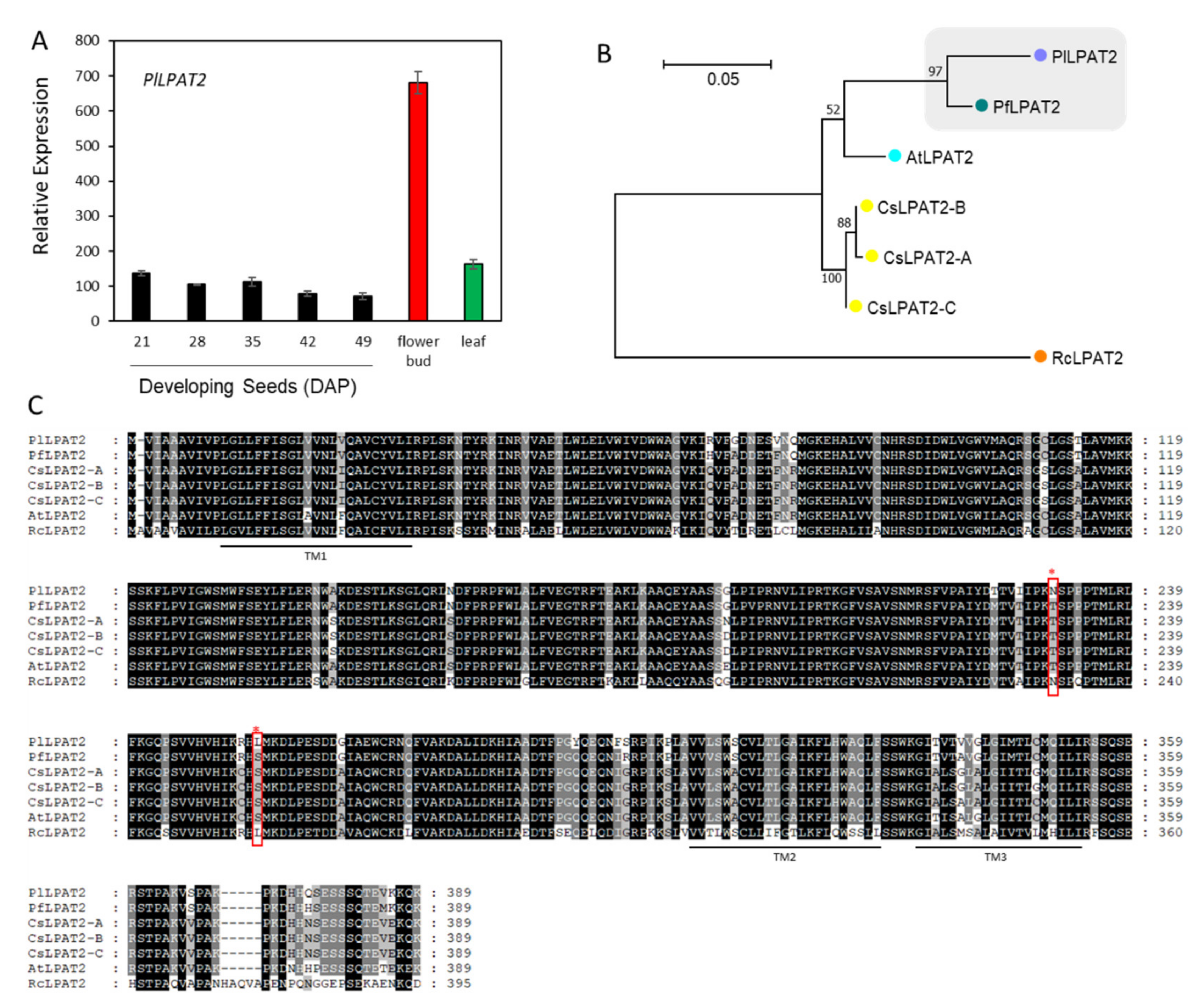
2.5. Lyso-PC Acyltransferase, FA-Condensing Enzyme, and Phospholipases A
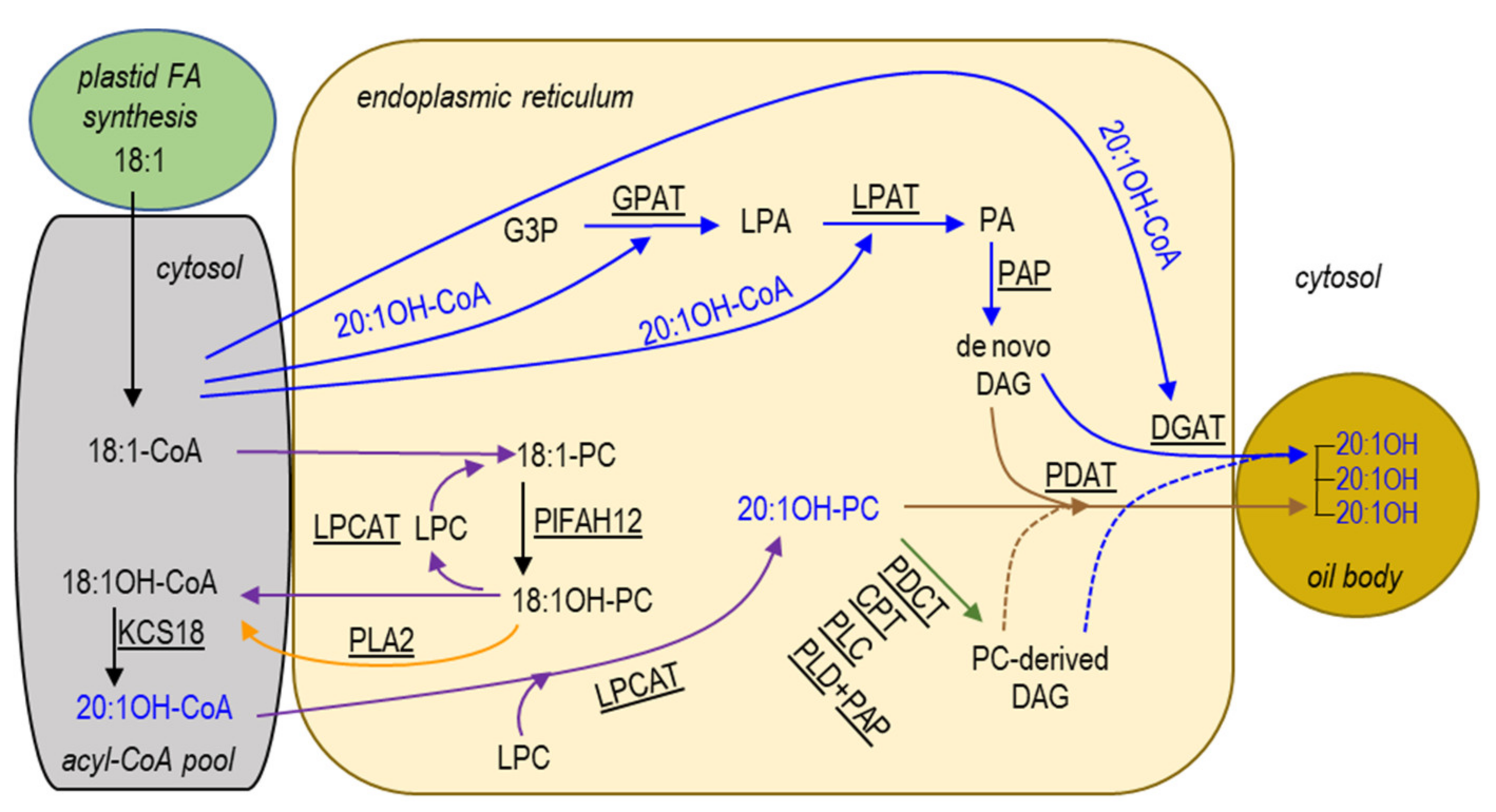
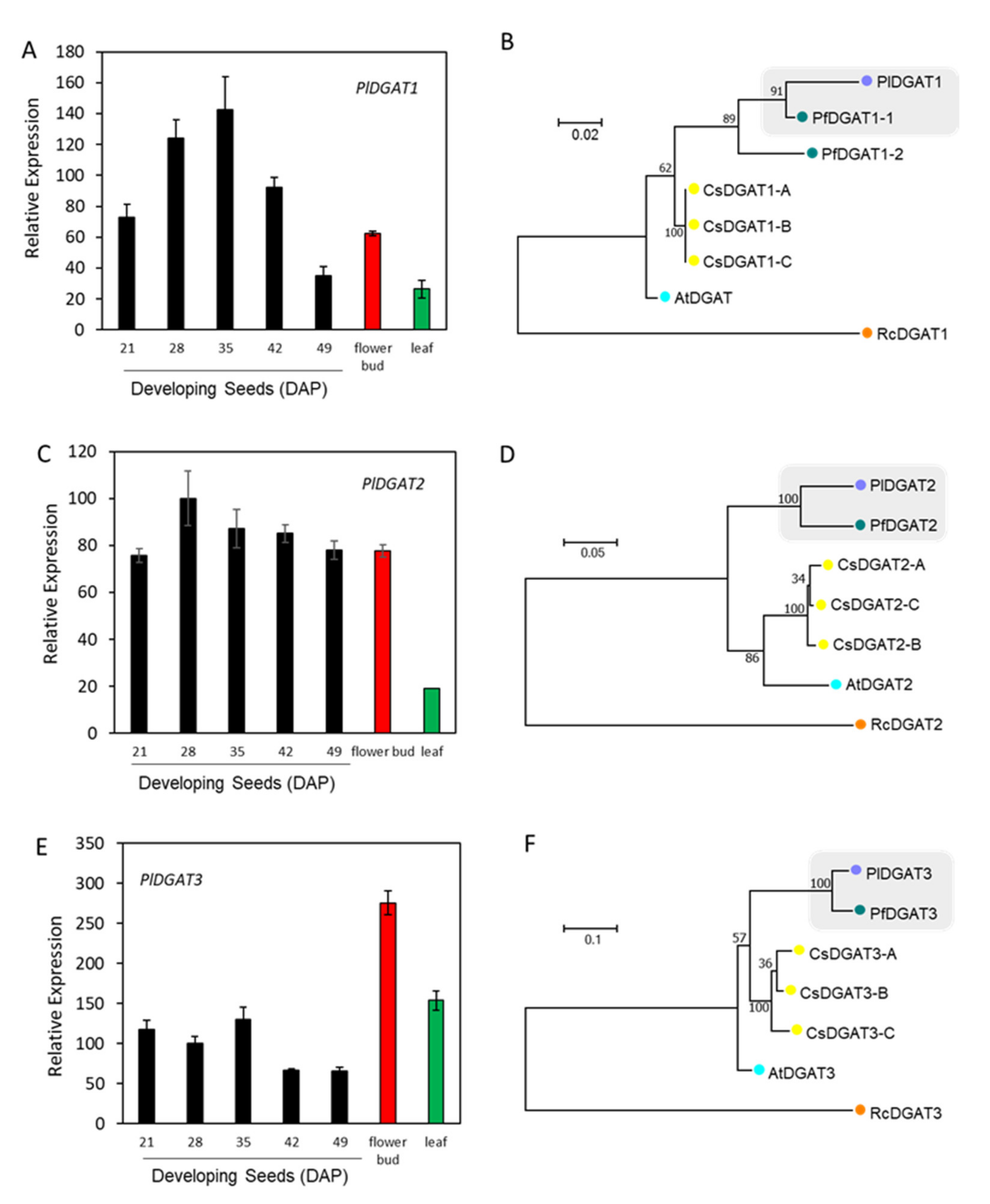
2.6. Kennedy Pathway for TAG Synthesis in the ER
2.7. Acyl-CoA-Independent and PC-Mediated Pathways for TAG Synthesis
3. Materials and Methods
3.1. Plant Materials and RNA Extraction
3.2. cDNA Library Construction
3.3. Sequencing and Sequence Quality Check
3.4. Transcriptome Analysis
3.5. Protein Sequence Analysis
3.6. Quantitative PCR
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Caupin, H.J. Products from castor oil: Past, present and future. In Lipid Technologies and Applications; Gunstone, F.D., Padley, F.B., Eds.; Marcel Dekker: New York, NY, USA, 1997. [Google Scholar]
- Roetheli, J.C. Lesquerella as a Source of Hydroxy Fatty Acids for Industrial Products; Growing Industrial Materials Series; USDA CSRS: Washington, DC, USA, 1991. Available online: https://www.ars.usda.gov/ARSUserFiles/53442010/NewCrops/Lesquerella/PDF/LesquerellaBooklet.pdf (accessed on 5 January 2021).
- Chen, G.Q.; He, X.; McKeon, T.A. A Simple and Sensitive Assay for Distinguishing the Expression of Ricin andRicinus communisAgglutinin Genes in Developing Castor Seed (R. communis L.). J. Agric. Food Chem. 2005, 53, 2358–2361. [Google Scholar] [CrossRef] [PubMed]
- Lord, M.J.; He, X.; Liao, L.P.; McKeon, T.A. Ricin: Mechanisms of cytotoxicity. Toxicol. Rev. 2003, 22, 53–64. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.Q.; He, X.; Liao, L.P.; McKeon, T.A. 2S albumin gene expression in castor plant (Ricinus communis L.). J. Am. Oil Chem. Soc. 2004, 81, 867–872. [Google Scholar] [CrossRef]
- Machado, O.L.T.; Silva, J.G., Jr. An allergenic 2S storage protein from Ricinus communis seeds which is a part of the 2S albumin precursor predicted by C-DNA data. Braz. J. Med. Biol. Res. 1992, 25, 567–582. [Google Scholar]
- Youle, R.J.; Huang, A.H.C. Albumin Storage Proteins in the Protein Bodies of Castor Bean. Plant Physiol. 1978, 61, 13–16. [Google Scholar] [CrossRef] [Green Version]
- Chen, G.Q.; Lin, J.-T.; Lu, C. Hydroxy fatty acid synthesis and lipid gene expression during seed development in Lesquerella fendleri. Ind. Crop Prod. 2011, 34, 1286–1292. [Google Scholar] [CrossRef]
- Hayes, D.G.; Carlson, K.D.; Kleiman, R. The isolation of hydroxy acids from lesquerella oil lipolysate by a saponification/extraction technique. J. Am. Oil Chem. Soc. 1996, 73, 1113–1119. [Google Scholar] [CrossRef]
- Dierig, D.; Wang, G.; McCloskey, W.; Thorp, K.; Isbell, T.; Ray, D.; Foster, M. Lesquerella: New crop development and commercialization in the U.S. Ind. Crop Prod. 2011, 34, 1381–1385. [Google Scholar] [CrossRef]
- Smith, C.R., Jr.; Wilson, T.L.; Miwa, T.K.; Zobel, H.; Lohmar, R.L.; Wolff, I.A. Lesquerolic acid. A new hydroxy acid from Lesquerella seed oil. J. Org. Chem. 1961, 26, 2903–2905. [Google Scholar] [CrossRef]
- Von Cruz, M.V.; Dierig, D.A. Germplasm improvement to develop commercially viable lines of the new oilseed crop lesquerella. In Industrial Crops: Breeding for Bioenergy and Bioproducts; Springer: New York, NY, USA, 2015; pp. 315–334. [Google Scholar]
- Dauk, M.; Lam, P.; Kunst, L.; Smith, M.A. A FAD2 homologue from Lesquerella lindheimeri has predominantly fatty acid hydroxylase activity. Plant Sci. 2007, 173, 43–49. [Google Scholar] [CrossRef] [Green Version]
- Jenderek, M.; Dierig, D.; Isbell, T. Fatty-acid profile of Lesquerella germplasm in the National Plant Germplasm System collection. Ind. Crop Prod. 2009, 29, 154–164. [Google Scholar] [CrossRef]
- Chen, G.Q.; Riiff, T.J.; Johnson, K.; Morales, E.; Kim, H.U.; Lee, K.-R.; Lin, J.-T. Seed development and hydroxy fatty acid biosynthesis in Physaria lindheimeri. Ind. Crop Prod. 2017, 108, 410–415. [Google Scholar] [CrossRef]
- Dierig, D.A.; Tomasi, P.M.; Salywon, A.M.; Ray, D.T. Improvement in hydroxy fatty acid seed oil content and other traits from interspecific hybrids of three Lesquerella species: Lesquerella fendleri, L. pallida, and L. lindheimeri. Euphytica 2004, 139, 199–206. [Google Scholar] [CrossRef]
- Chen, G.Q. Effective Reduction of Chimeric Tissue in Transgenics for the Stable Genetic Transformation of Lesquerella fendleri. HortScience 2011, 46, 86–90. [Google Scholar] [CrossRef] [Green Version]
- Chen, G.Q.; Van Erp, H.; Martin-Moreno, J.; Johnson, K.; Morales, E.; Browse, J.; Eastmond, P.J.; Lin, J.-T. Expression of Castor LPAT2 Enhances Ricinoleic Acid Content at the sn-2 Position of Triacylglycerols in Lesquerella Seed. Int. J. Mol. Sci. 2016, 17, 507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kokotou, M.G.; Kokotos, A.C.; Gkikas, D.; Mountanea, O.G.; Mantzourani, C.; Almutairi, A.; Lei, X.; Ramanadham, S.; Politis, P.K.; Kokotos, G. Saturated Hydroxy Fatty Acids Exhibit a Cell Growth Inhibitory Activity and Suppress the Cytokine-Induced β-Cell Apoptosis. J. Med. Chem. 2020, 63, 12666–12681. [Google Scholar] [CrossRef] [PubMed]
- Kokotou, M.G.; Mantzourani, C.; Bourboula, A.; Mountanea, O.G.; Kokotos, G. A Liquid Chromatography-High Resolution Mass Spectrometry (LC-HRMS) Method for the Determination of Free Hydroxy Fatty Acids in Cow and Goat Milk. Molecules 2020, 25, 3947. [Google Scholar] [CrossRef]
- Li-Beisson, Y.; Shorrosh, B.; Beisson, F.; Andersson, M.X.; Arondel, V.; Bates, P.D.; Baud, S.; Bird, D.; Debono, A.; Durrett, T.P.; et al. Acyl-Lipid Metabolism. Arab. Book 2013, 11, e0161. [Google Scholar] [CrossRef] [Green Version]
- Bates, P.D.; Browse, J. The pathway of triacylglycerol synthesis through phosphatidylcholine in Arabidopsis produces a bottleneck for the accumulation of unusual fatty acids in transgenic seeds. Plant J. 2011, 68, 387–399. [Google Scholar] [CrossRef]
- Lands, W.E. Lipid Metabolism. Annu. Rev. Biochem. 1965, 34, 313–346. [Google Scholar] [CrossRef]
- Lager, I.; Lindberg Yilmaz, J.; Zhou, X.-R.; Jasieniecka, K.; Kazachkov, M.; Wang, P.; Zou, J.; Weselake, R.; Smith, M.A.; Bayon, S.; et al. Plant acyl-CoA:lysophosphatidylcholine acyltransferases (LPCATs) have different specificities in their forward and reverse reactions. J. Biol. Chem. 2013, 288, 36902–36914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bates, P.D. Understanding the control of acyl flux through the lipid metabolic network of plant oil biosynthesis. Biochim. et Biophys. Acta (BBA) Mol. Cell Biol. Lipids 2016, 1861, 1214–1225. [Google Scholar] [CrossRef]
- Bafor, M.; A Smith, M.; Jonsson, L.; Stobart, K.; Stymne, S. Ricinoleic acid biosynthesis and triacylglycerol assembly in microsomal preparations from developing castor-bean (Ricinus communis) endosperm. Biochem. J. 1991, 280, 507–514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreau, R.A.; Stumpf, P.K. Recent Studies of the Enzymic Synthesis of Ricinoleic Acid by Developing Castor Beans. Plant Physiol. 1981, 67, 672–676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van De Loo, F.J.; Broun, P.; Turner, S.; Somerville, C. An oleate 12-hydroxylase from Ricinus communis L. is a fatty acyl desaturase homolog. Proc. Natl. Acad. Sci. USA 1995, 92, 6743–6747. [Google Scholar] [CrossRef] [Green Version]
- Bayon, S.; Chen, G.; Weselake, R.J.; Browse, J. A Small Phospholipase A2-α from Castor Catalyzes the Removal of Hydroxy Fatty Acids from Phosphatidylcholine in Transgenic Arabidopsis Seeds. Plant Physiol. 2015, 167, 1259–1270. [Google Scholar] [CrossRef] [Green Version]
- Moon, H.; Smith, M.A.; Kunst, L. A condensing enzyme from the seeds of Lesquerella fendleri that specifically elongates hydroxy fatty acids. Plant Physiol. 2001, 127, 1635–1643. [Google Scholar] [CrossRef]
- Weiss, S.B.; Kennedy, E.P. The enzymatic synthesis of triglycerides. J. Am. Chem. Soc. 1956, 78, 3550. [Google Scholar] [CrossRef]
- Weiss, S.B.; Kennedy, E.P.; Kiyasu, J.Y. The enzymatic synthesis of triglycerides. J. Biol. Chem. 1960, 235, 40–44. [Google Scholar] [CrossRef]
- Kennedy, E.P. Biosynthesis of complex lipids. Fed. Proc. 1961, 20, 934–940. [Google Scholar]
- Dahlqvist, A.; Ståhl, U.; Lenman, M.; Banas, A.; Lee, M.; Sandager, L.; Ronne, H.; Stymne, S. Phospholipid:diacylglycerol acyltransferase: An enzyme that catalyzes the acyl-CoA-independent formation of triacylglycerol in yeast and plants. Proc. Natl. Acad. Sci. USA 2000, 97, 6487–6492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.U.; Lee, K.-R.; Go, Y.S.; Jung, J.H.; Suh, M.-C.; Kim, J.B. Endoplasmic Reticulum-Located PDAT1-2 from Castor Bean Enhances Hydroxy Fatty Acid Accumulation in Transgenic Plants. Plant Cell Physiol. 2011, 52, 983–993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Erp, H.; Bates, P.D.; Burgal, J.; Shockey, J. Castor phospholipid:diacylglycerol acyltransferase facilitates efficient metabolism of hydroxy fatty acids in transgenic Arabidopsis. Plant Physiol. 2011, 155, 683–693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okuley, J.; Lightner, J.; Feldmann, K.; Yadav, N.; Lark, E. Arabidopsis FAD2 gene encodes the enzyme that is essential for polyunsaturated lipid synthesis. Plant Cell 1994, 6, 147. [Google Scholar] [PubMed]
- Arondel, V.; Lemieux, B.; Hwang, I.; Gibson, S.; Goodman, H.M.; Somerville, C.R. Map-based cloning of a gene controlling omega-3 fatty acid desaturation in Arabidopsis. Science 1992, 258, 1353. [Google Scholar] [CrossRef]
- Yadav, N.S.; Wierzbicki, A.; Aegerter, M.; Caster, C.S.; Perez-Grau, L.; Kinney, A.J.; Hitz, W.D.; Booth, J.R., Jr.; Schweiger, B.; Stecca, K.L.; et al. Cloning of higher plant omega-3 fatty acid desaturases. Plant Physiol. 1993, 103, 467–476. [Google Scholar] [CrossRef] [Green Version]
- Lu, C.; Xin, Z.; Ren, Z.; Miquel, M.; Browse, J. An enzyme regulating triacylglycerol composition is encoded by the ROD1 gene of Arabidopsis. Proc. Natl. Acad. Sci. USA 2009, 106, 18837–18842. [Google Scholar] [CrossRef] [Green Version]
- Hu, Z.; Ren, Z.; Lu, C. The Phosphatidylcholine Diacylglycerol Cholinephosphotransferase Is Required for Efficient Hydroxy Fatty Acid Accumulation in Transgenic Arabidopsis. Plant Physiol. 2012, 158, 1944–1954. [Google Scholar] [CrossRef] [Green Version]
- Slack, C.R.; Campbell, L.C.; Browse, J.A.; Roughan, P.G. Some evidence for the reversibility of the cholinephosphotransferasecatalysed reaction in developing linseed cotyledons in vivo. Biochim. Biophys. Acta (BBA) Lipids Lipid Metab. 1983, 754, 10–20. [Google Scholar] [CrossRef]
- Lee, K.-R.; Chen, G.Q.; Kim, H.U. Current progress towards the metabolic engineering of plant seed oil for hydroxy fatty acids production. Plant Cell Rep. 2015, 34, 603–615. [Google Scholar] [CrossRef]
- Broun, P.; Somerville, C.; Morell, M.K.; Blennow, A.; Kosar-Hashemi, B.; Samuel, M.S. Accumulation of Ricinoleic, Lesquerolic, and Densipolic Acids in Seeds of Transgenic Arabidopsis Plants That Express a Fatty Acyl Hydroxylase cDNA from Castor Bean. Plant Physiol. 1997, 113, 933–942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, C.; Fulda, M.; Wallis, J.G.; Browse, J. A high-throughput screen for genes from castor that boost hydroxy fatty acid accumulation in seed oils of transgenic Arabidopsis. Plant J. 2006, 45, 847–856. [Google Scholar] [CrossRef] [PubMed]
- Burgal, J.; Shockey, J.; Lu, C.; Dyer, J.; Larson, T.; Graham, I.; Browse, J. Metabolic engineering of hydroxy fatty acid production in plants: RcDGAT2 drives dramatic increases in ricinoleate levels in seed oil. Plant Biotechnol. J. 2008, 6, 819–831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aryal, N.; Lu, C. A Phospholipase C-Like Protein From Ricinus communis Increases Hydroxy Fatty Acids Accumulation in Transgenic Seeds of Camelina sativa. Front. Plant Sci. 2018, 9, 1576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shockey, J.; Lager, I.; Stymne, S.; Kotapati, H.K.; Sheffield, J.; Mason, C.; Bates, P.D. Specialized lysophosphatidic acid acyltransferases contribute to unusual fatty acid accumulation in exotic Euphorbiaceae seed oils. Planta 2019, 249, 1285–1299. [Google Scholar] [CrossRef]
- Kim, H.U.; Park, M.-E.; Lee, K.-R.; Suh, M.C.; Chen, G.Q. Variant castor lysophosphatidic acid acyltransferases acylate ricinoleic acid in seed oil. Ind. Crop Prod. 2020, 150, 112245. [Google Scholar] [CrossRef]
- Lunn, D.; Wallis, J.G.; Browse, J. Tri-Hydroxy-Triacylglycerol Is Efficiently Produced by Position-Specific Castor Acyltransferases. Plant Physiol. 2019, 179, 1050–1063. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.; Chen, G.; Mietkiewska, E.; Song, Z.; Caldo, K.M.P.; Singer, S.D.; Dyer, J.; Smith, M.; McKeon, T.; Weselake, R.J. Castor patatin-like phospholipase A IIIβ facilitates removal of hydroxy fatty acids from phosphatidylcholine in transgenic Arabidopsis seeds. Plant Mol. Biol. 2019, 101, 521–536. [Google Scholar] [CrossRef]
- Xu, Y.; Caldo, K.M.P.; Singer, S.D.; Mietkiewska, E.; Greer, M.S.; Tian, B.; Dyer, J.M.; Smith, M.; Zhou, X.R.; Qiu, X.; et al. Physaria fendleri and Ricinus communis LCAT-like phospholipases selectively cleave hydroxy acyl chains from phosphatidylcholine. Plant J. 2020. [Google Scholar] [CrossRef]
- Lin, J.-T.; Turner, C.; Liao, L.P.; McKeon, T.A. Identification and Quantification of the Molecular Species of Acylglycerols in Castor Oil by HPLC Using ELSD. J. Liq. Chromatogr. Relat. Technol. 2003, 26, 773–780. [Google Scholar] [CrossRef]
- Hayes, D.G.; Kleiman, R. A detailed triglyceride analysis ofLesquerella fendleri oil: Column chromatographic fractionation followed by supercritical fluid chromatography. J. Am. Oil Chem. Soc. 1996, 73, 267–269. [Google Scholar] [CrossRef]
- Chen, G.Q.; Lin, J.-T.; Van Erp, H.; Johnson, K.; Lu, C. Regiobiochemical analysis reveals the role of castor LPAT2 in the accumulation of hydroxy fatty acids in transgenic lesquerella seeds. Biocatal. Agric. Biotechnol. 2020, 25, 101617. [Google Scholar] [CrossRef]
- Chan, A.P.; Crabtree, J.; Zhao, Q.; Lorenzi, H.; Orvis, J.; Puiu, D.; Melake-Berhan, A.; Jones, K.M.; Redman, J.C.; Chen, G.; et al. Draft genome sequence of the oilseed species Ricinus communis. Nat. Biotechnol. 2010, 28, 951–956. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Troncoso-Ponce, M.A.; Kilaru, A.; Cao, X.; Durrett, T.P.; Fan, J.; Jensen, J.K.; Thrower, N.A.; Pauly, M.; Wilkerson, C.; Ohlrogge, J.B. Comparative deep transcriptional profiling of four developing oilseeds. Plant J. 2011, 68, 1014–1027. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.U.; Chen, G.Q. Identification of hydroxy fatty acid and triacylglycerol metabolism-related genes in lesquerella through seed transcriptome analysis. BMC Genom. 2015, 16, 230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horn, P.J.; Liu, J.; Cocuron, J.-C.; McGlew, K.; Thrower, N.A.; Larson, M.; Lu, C.; Alonso, A.P.; Ohlrogge, J. Identification of multiple lipid genes with modifications in expression and sequence associated with the evolution of hydroxy fatty acid accumulation in Physaria fendleri. Plant J. 2016, 86, 322–348. [Google Scholar] [CrossRef]
- McGinnis, S.; Madden, T.L. BLAST: At the core of a powerful and diverse set of sequence analysis tools. Nucleic Acids Res. 2004, 32, W20–W25. [Google Scholar] [CrossRef]
- Trapnell, C.; Williams, B.A.; Pertea, G.; Mortazavi, A.; Kwan, G.; Van Baren, M.J.; Salzberg, S.L.; Wold, B.J.; Pachter, L. Transcript assembly and quantification by RNA-Seq reveals unannotated transcripts and isoform switching during cell differentiation. Nat. Biotechnol. 2010, 28, 511–515. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Ilarslan, H.; Brachova, L.; Qian, H.-R.; Li, L.; Che, P.; Wurtele, E.S.; Nikolau, B.J. Reverse-Genetic Analysis of the Two Biotin-Containing Subunit Genes of the Heteromeric Acetyl-Coenzyme A Carboxylase in Arabidopsis Indicates a Unidirectional Functional Redundancy. Plant Physiol. 2010, 155, 293–314. [Google Scholar] [CrossRef] [Green Version]
- Schnurr, J.A.; Shockey, J.M.; De Boer, G.-J.; Browse, J.A. Fatty Acid Export from the Chloroplast. Molecular Characterization of a Major Plastidial Acyl-Coenzyme A Synthetase from Arabidopsis. Plant Physiol. 2002, 129, 1700–1709. [Google Scholar] [CrossRef] [Green Version]
- Zhao, L.; Katavic, V.; Li, F.; Haughn, G.W.; Kunst, L. Insertional mutant analysis reveals that long-chain acyl-CoA synthetase 1 (LACS1), but not LACS8, functionally overlaps with LACS9 in Arabidopsis seed oil biosynthesis. Plant J. 2010, 64, 1048–1058. [Google Scholar] [CrossRef] [PubMed]
- Broun, P.; Shanklin, J.; Whittle, E.; Somerville, C. Catalytic Plasticity of Fatty Acid Modification Enzymes Underlying Chemical Diversity of Plant Lipids. Science 1998, 282, 1315. [Google Scholar] [CrossRef] [PubMed]
- Broun, P.; Boddupalli, S.; Somerville, C. A bifunctional oleate 12-hydroxylase: Desaturase fromLesquerella fendleri. Plant J. 1998, 13, 201–210. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.-R.; Kim, E.-H.; Jeon, I.; Lee, Y.; Chen, G.Q.; Kim, H.U. Lesquerella FAD3-1 gene is responsible for the biosynthesis of trienoic acid and dienoic hydroxy fatty acids in seed oil. Ind. Crop Prod. 2019, 134, 257–264. [Google Scholar] [CrossRef] [Green Version]
- Los, D.A.; Murata, N. Structure and expression of fatty acid desaturases. Biochim. Biophys. Acta (BBA) Lipids Lipid Metab. 1998, 1394, 3–15. [Google Scholar] [CrossRef]
- Broadwater, J.A.; Whittle, E.; Shanklin, J. Desaturation and hydroxylation. Residues 148 and 324 of Arabidopsis FAD2, in addition to substrate chain length, exert a major influence in partitioning of catalytic specificity. J. Biol. Chem. 2002, 277, 15613–15620. [Google Scholar] [CrossRef] [Green Version]
- Yurchenko, O.P.; Park, S.; Ilut, D.C.; Inmon, J.J.; Millhollon, J.C.; Liechty, Z.; Page, J.T.; Jenks, M.A.; Chapman, K.D.; Udall, J.A.; et al. Genome-wide analysis of the omega-3 fatty acid desaturase gene family in Gossypium. BMC Plant Biol. 2014, 14, 312. [Google Scholar] [CrossRef] [Green Version]
- Ghanevati, M.; Jaworski, J.G. Active-site residues of a plant membrane-bound fatty acid elongase beta-ketoacyl-CoA synthase, FAE1 KCS. Biochim. Biophys. Acta 2001, 1530, 77–85. [Google Scholar] [CrossRef]
- Smith, M.A.; Moon, H.; Chowrira, G.; Kunst, L. Heterologous expression of a fatty acid hydroxylase gene in developing seeds of Arabidopsis thaliana. Planta 2003, 217, 507–516. [Google Scholar] [CrossRef]
- Snapp, A.R.; Kang, J.; Qi, X.; Lu, C. A fatty acid condensing enzyme from Physaria fendleri increases hydroxy fatty acid accumulation in transgenic oilseeds of Camelina sativa. Planta 2014, 240, 599–610. [Google Scholar] [CrossRef]
- Shockey, J.; Regmi, A.; Cotton, K.; Adhikari, N.D.; Browse, J.; Bates, P.D. Identification of Arabidopsis GPAT9 (At5g60620) as an Essential Gene Involved in Triacylglycerol Biosynthesis. Plant Physiol. 2016, 170, 163–179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singer, S.D.; Chen, G.; Mietkiewska, E.; Tomasi, P.; Jayawardhane, K.; Dyer, J.M.; Weselake, R.J. Arabidopsis GPAT9 contributes to synthesis of intracellular glycerolipids but not surface lipids. J. Exp. Bot. 2016, 67, 4627–4638. [Google Scholar] [CrossRef] [PubMed]
- Waschburger, E.; Kulcheski, F.R.; Veto, N.M.; Margis, R.; Margis-Pinheiro, M.; Turchetto-Zolet, A.C. Genome-wide analysis of the Glycerol-3-Phosphate Acyltransferase (GPAT) gene family reveals the evolution and diversification of plant GPATs. Genet. Mol. Biol. 2018, 41 (Suppl. 1), 355–370. [Google Scholar] [CrossRef] [PubMed]
- Frentzen, M. Acyltransferases from basic science to modified seed oils. Lipid/Fett 1998, 100, 161–166. [Google Scholar] [CrossRef]
- Ohlrogge, J.B.; Thrower, N.; Mhaske, V.; Stymne, S.; Baxter, M.; Yang, W.; Liu, J.; Shaw, K.; Shorrosh, B.; Zhang, M.; et al. PlantFAdb: A resource for exploring hundreds of plant fatty acid structures synthesized by thousands of plants and their phylogenetic relationships. Plant J. 2018, 96, 1299–1308. [Google Scholar] [CrossRef] [Green Version]
- Dyer, J.M.; Chapital, D.C.; Kuan, J.-C.W.; Mullen, R.T.; Turner, C.; McKeon, T.A.; Pepperman, A.B. Molecular Analysis of a Bifunctional Fatty Acid Conjugase/Desaturase from Tung. Implications for the Evolution of Plant Fatty Acid Diversity1. Plant Physiol. 2002, 130, 2027–2038. [Google Scholar] [CrossRef] [Green Version]
- Jonas, M.; Ketlogetswe, C.; Gandure, J. Variation of Jatropha curcas seed oil content and fatty acid composition with fruit maturity stage. Heliyon 2020, 6, e03285. [Google Scholar] [CrossRef]
- Kim, H.U.; Huang, A.H. Plastid Lysophosphatidyl Acyltransferase Is Essential for Embryo Development in Arabidopsis. Plant Physiol. 2004, 134, 1206–1216. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.U.; Li, Y.; Huang, A.H. Ubiquitous and Endoplasmic Reticulum–Located Lysophosphatidyl Acyltransferase, LPAT2, Is Essential for Female but Not Male Gametophyte Development in Arabidopsis. Plant Cell 2005, 17, 1073–1089. [Google Scholar] [CrossRef] [Green Version]
- Bourgis, F.; Kader, J.-C.; Barret, P.; Renard, M.; Robinson, D.; Robinson, C.; Delseny, M.; Roscoe, T.J. A Plastidial Lysophosphatidic Acid Acyltransferase from Oilseed Rape. Plant Physiol. 1999, 120, 913–922. [Google Scholar] [CrossRef] [Green Version]
- Phillips, B.E.; Smith, C.R.; Tallent, W.H. Glycerides ofLimnanthes douglasii seed oil. Lipids 1971, 6, 93–99. [Google Scholar] [CrossRef]
- Balachandran, C.; Arumughan, C.; Mathew, A.G. Distribution of major chemical constituents and fatty acids in different regions of coconut endosperm. J. Am. Oil Chem. Soc. 1985, 62, 1583–1586. [Google Scholar] [CrossRef]
- Kim, H.J.; Silva, J.E.; Iskandarov, U.; Andersson, M.; Cahoon, E.B.; Mockaitis, K.; Cahoon, E.B. Structurally divergent lysophosphatidic acid acyltransferases with high selectivity for saturated medium chain fatty acids from Cuphea seeds. Plant J. 2015, 84, 1021–1033. [Google Scholar] [CrossRef] [PubMed]
- Brough, C.L.; Coventry, J.M.; Christie, W.W.; Kroon, J.T.M.; Brown, A.P.; Barsby, T.L.; Slabas, A.R. Towards the genetic engineering of triacylglycerols of defined fatty acid composition: Major changes in erucic acid content at the sn-2 position affected by the introduction of a 1-acyl-sn-glycerol-3-phosphate acyltransferase from Limnanthes douglasii into oil seed rape. Mol. Breed. 1996, 2, 133–142. [Google Scholar]
- Knutzon, D.S.; Hayes, T.R.; Wyrick, A.; Xiong, H.; Davies, H.M.; Voelker, T.A. Lysophosphatidic Acid Acyltransferase from Coconut Endosperm Mediates the Insertion of Laurate at the sn-2 Position of Triacylglycerols in Lauric Rapeseed Oil and Can Increase Total Laurate Levels. Plant Physiol. 1999, 120, 739–746. [Google Scholar] [CrossRef] [Green Version]
- Lassner, M.W.; Levering, C.K.; Davies, H.M.; Knutzon, D.S. Lysophosphatidic Acid Acyltransferase from Meadowfoam Mediates Insertion of Erucic Acid at the sn-2 Position of Triacylglycerol in Transgenic Rapeseed Oil. Plant Physiol. 1995, 109, 1389–1394. [Google Scholar] [CrossRef] [Green Version]
- Arroyo-Caro, J.M.; Chileh, T.; Kazachkov, M.; Zou, J.; Alonso, D.L.; García-Maroto, F. The multigene family of lysophosphatidate acyltransferase (LPAT)-related enzymes in Ricinus communis. Cloning and molecular characterization of two LPAT genes that are expressed in castor seeds. Plant Sci. 2013, 199–200, 29–40. [Google Scholar] [CrossRef]
- Yu, X.-H.; Prakash, R.R.; Sweet, M.; Shanklin, J. Coexpressing Escherichia coli cyclopropane synthase with Sterculia foetida Lysophosphatidic acid acyltransferase enhances cyclopropane fatty acid accumulation. Plant Physiol. 2013, 164, 455–465. [Google Scholar] [CrossRef] [Green Version]
- Yu, X.-H.; Cahoon, R.E.; Horn, P.J.; Shi, H.; Prakash, R.R.; Cai, Y.; Hearney, M.; Chapman, K.D.; Cahoon, E.B.; Schwender, J.; et al. Identification of bottlenecks in the accumulation of cyclic fatty acids in camelina seed oil. Plant Biotechnol. J. 2018, 16, 926–938. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Caldo, K.M.P.; Pal-Nath, D.; Ozga, J.; Lemieux, M.J.; Weselake, R.J.; Chen, G. Properties and Biotechnological Applications of Acyl-CoA:diacylglycerol Acyltransferase and Phospholipid:diacylglycerol Acyltransferase from Terrestrial Plants and Microalgae. Lipids 2018, 53, 663–688. [Google Scholar] [CrossRef]
- Aymé, L.; Arragain, S.; Canonge, M.; Baud, S.; Touati, N.; Bimai, O.; Jagic, F.; Louis-Mondésir, C.; Briozzo, P.; Fontecave, M.; et al. Arabidopsis thaliana DGAT3 is a [2Fe-2S] protein involved in TAG biosynthesis. Sci. Rep. 2018, 8, 17254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernández, M.L.; Whitehead, L.; He, Z.; Gazda, V.; Gilday, A.D.; Kozhevnikova, E.; Vaistij, F.E.; Larson, T.R.; Graham, I. A Cytosolic Acyltransferase Contributes to Triacylglycerol Synthesis in Sucrose-Rescued Arabidopsis Seed Oil Catabolism Mutants. Plant Physiol. 2012, 160, 215–225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chi, X.; Hu, R.; Zhang, X.; Chen, M.; Chen, N.; Pan, L.; Wang, T.; Wang, M.; Yang, Z.; Wang, Q.; et al. Cloning and Functional Analysis of Three Diacylglycerol Acyltransferase Genes from Peanut (Arachis hypogaea L.). PLoS ONE 2014, 9, e105834. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Siloto, R.M.; Lehner, R.; Stone, S.J.; Weselake, R.J. Acyl-CoA:diacylglycerol acyltransferase: Molecular biology, biochemistry and biotechnology. Prog. Lipid Res. 2012, 51, 350–377. [Google Scholar] [CrossRef] [PubMed]
- Cao, H. Structure-Function Analysis of Diacylglycerol Acyltransferase Sequences from 70 Organisms. BMC Res. Notes 2011, 4, 249. [Google Scholar] [CrossRef] [Green Version]
- Aznar-Moreno, J.A.; Denolf, P.; Van Audenhove, K.; De Bodt, S.; Engelen, S.; Fahy, D.; Wallis, J.G.; Browse, J. Type 1 diacylglycerol acyltransferases of Brassica napus preferentially incorporate oleic acid into triacylglycerol. J. Exp. Bot. 2015, 66, 6497–6506. [Google Scholar] [CrossRef] [Green Version]
- Lager, I.; Jeppson, S.; Gippert, A.-L.; Feussner, I.; Stymne, S.; Marmon, S. Acyltransferases Regulate Oil Quality in Camelina sativa Through Both Acyl Donor and Acyl Acceptor Specificities. Front. Plant Sci. 2020, 11, 1144. [Google Scholar] [CrossRef]
- Demski, K.; Jeppson, S.; Lager, I.; Misztak, A.; Jasieniecka-Gazarkiewicz, K.; Waleron, M.; Stymne, S.; Banaś, A. Isoforms of Acyl-CoA:Diacylglycerol Acyltransferase2 Differ Substantially in Their Specificities toward Erucic Acid1. Plant Physiol. 2019, 181, 1468–1479. [Google Scholar] [CrossRef]
- Kroon, J.T.M.; Wei, W.; Simon, W.J.; Slabas, A.R. Identification and functional expression of a type 2 acyl-CoA:diacylglycerol acyltransferase (DGAT2) in developing castor bean seeds which has high homology to the major triglyceride biosynthetic enzyme of fungi and animals. Phytochemistry 2006, 67, 2541–2549. [Google Scholar] [CrossRef]
- Shockey, J.M.; Gidda, S.K.; Chapital, D.C.; Kuan, J.-C.; Dhanoa, P.K.; Bland, J.M.; Rothstein, S.J.; Mullen, R.T.; Dyer, J.M. Tung Tree DGAT1 and DGAT2 Have Nonredundant Functions in Triacylglycerol Biosynthesis and Are Localized to Different Subdomains of the Endoplasmic Reticulum. Plant Cell 2006, 18, 2294–2313. [Google Scholar] [CrossRef] [Green Version]
- Li, R.; Yu, K.; Jamboonsri, W. DGAT1, DGAT2 and PDAT Expression in Seeds and Other Tissues of Epoxy and Hydroxy Fatty Acid Accumulating Plants. Lipids 2010, 45, 145–157. [Google Scholar] [CrossRef] [PubMed]
- Jeppson, S.; Demski, K.; Carlsson, A.S.; Zhu, L.H.; Banaś, A.; Stymne, S.; Lager, I. Crambe hispanica Subsp. abyssinica Diacylglycerol Acyltransferase Specificities Towards Diacylglycerols and Acyl-CoA Reveal Combinatorial Effects That Greatly Affect Enzymatic Activity and Specificity. Front. Plant Sci. 2019, 10, 1442. [Google Scholar] [CrossRef] [PubMed]
- Durrett, T.P.; McClosky, D.D.; Tumaney, A.W.; Elzinga, D.A.; Ohlrogge, J.; Pollard, M. A distinct DGAT with sn-3 acetyltransferase activity that synthesizes unusual, reduced-viscosity oils in Euonymus and transgenic seeds. Proc. Natl. Acad. Sci. USA 2010, 107, 9464–9469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Rice, A.; McGlew, K.; Shaw, V.; Park, H.; Clemente, T.E.; Pollard, M.; Ohlrogge, J.; Durrett, T.P. Metabolic engineering of oilseed crops to produce high levels of novel acetyl glyceride oils with reduced viscosity, freezing point and calorific value. Plant Biotechnol. J. 2015, 13, 858–865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Q.; Siloto, R.M.; Snyder, C.L.; Weselake, R.J. Functional and topological analysis of yeast acyl-CoA:diacylglycerol acyltransferase 2, an endoplasmic reticulum enzyme essential for triacylglycerol biosynthesis. J. Biol. Chem. 2011, 286, 13115–13126. [Google Scholar] [CrossRef] [Green Version]
- Stone, S.; Levin, M.; Farese, R.V. Membrane Topology and Identification of Key Functional Amino Acid Residues of Murine Acyl-CoA:Diacylglycerol Acyltransferase-2*. J. Biol. Chem. 2006, 281, 40273–40282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, R.; Yang, T.; Wang, R.; Liu, A. Characterisation of DGAT1 and DGAT2 from Jatropha curcas and their functions in storage lipid biosynthesis. Funct. Plant Biol. 2014, 41, 321–329. [Google Scholar] [CrossRef] [Green Version]
- Stahl, U.; Carlsson, A.S.; Lenman, M.; Dahlqvist, A.; Huang, B.; Banaś, W.; Banaś, A.; Stymne, S. Cloning and Functional Characterization of a Phospholipid:Diacylglycerol Acyltransferase from Arabidopsis. Plant Physiol. 2004, 135, 1324–1335. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; Fan, J.; Taylor, D.C.; Ohlrogge, J.B. DGAT1 and PDAT1 acyltransferases have overlapping functions in Arabidopsis triacylglycerol biosynthesis and are essential for normal pollen and seed development. Plant Cell 2009, 21, 3885–3901. [Google Scholar] [CrossRef] [Green Version]
- Fan, J.; Yan, C.; Xu, C. Phospholipid:diacylglycerol acyltransferase-mediated triacylglycerol biosynthesis is crucial for protection against fatty acid-induced cell death in growing tissues of Arabidopsis. Plant J. 2013, 76, 930–942. [Google Scholar] [CrossRef]
- Fan, J.; Yan, C.; Zhang, X.; Xu, C. Dual role for phospholipid: Diacylglycerol acyltransferase: Enhancing fatty acid synthesis and diverting fatty acids from membrane lipids to triacylglycerol in Arabidopsis leaves. Plant Cell 2013, 25, 3506–3518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, J.; Yan, C.; Roston, R.; Shanklin, J.; Xu, C. Arabidopsis lipins, PDAT1 acyltransferase, and SDP1 triacylglycerol lipase synergistically direct fatty acids toward β-oxidation, thereby maintaining membrane lipid homeostasis. Plant Cell 2014, 26, 4119–4134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mueller, S.P.; Unger, M.; Guender, L.; Fekete, A.; Mueller, M.J. Phospholipid:Diacylglycerol Acyltransferase-Mediated Triacylglyerol Synthesis Augments Basal Thermotolerance. Plant Physiol. 2017, 175, 486–497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, L.; Mao, X.; Zhao, K.; Ji, X.; Ji, C.; Xue, J.; Li, R. Characterisation of phospholipid: Diacylglycerol acyltransferases (PDATs) from Camelina sativa and their roles in stress responses. Biol. Open 2017, 6, 1024–1034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aznar-Moreno, J.A.; Durrett, T.P. Simultaneous Targeting of Multiple Gene Homeologs to Alter Seed Oil Production in Camelina sativa. Plant Cell Physiol. 2017, 58, 1260–1267. [Google Scholar] [CrossRef] [PubMed]
- Marmon, S.; Sturtevant, D.; Herrfurth, C.; Chapman, K.D.; Stymne, S.; Feussner, I. Two Acyltransferases Contribute Differently to Linolenic Acid Levels in Seed Oil. Plant Physiol. 2017, 173, 2081–2095. [Google Scholar] [CrossRef] [Green Version]
- Banaś, W.; Garcia, A.S.; Banaś, A.; Stymne, S. Activities of acyl-CoA:diacylglycerol acyltransferase (DGAT) and phospholipid:diacylglycerol acyltransferase (PDAT) in microsomal preparations of developing sunflower and safflower seeds. Planta 2013, 237, 1627–1636. [Google Scholar] [CrossRef] [Green Version]
- Pan, X.; Siloto, R.M.P.; Wickramarathna, A.D.; Mietkiewska, E.; Weselake, R.J. Identification of a Pair of Phospholipid:Diacylglycerol Acyltransferases from Developing Flax (Linum usitatissimum L.) Seed Catalyzing the Selective Production of Trilinolenin. J. Biol. Chem. 2013, 288, 24173–24188. [Google Scholar] [CrossRef] [Green Version]
- Chellamuthu, M.; Kumaresan, K.; Subramanian, S.; Muthumanickam, H. Functional Analysis of Sesame Diacylglycerol Acyltransferase and Phospholipid: Diacylglycerol Acyltransferase Genes Using In Silico and In Vitro Approaches. Plant Mol. Biol. Rep. 2019, 37, 146–156. [Google Scholar] [CrossRef]
- Peelman, F.; Vinaimont, N.; Vanloo, B.; Labeur, C.; Rosseneu, M.; Verhee, A.; Verschelde, J.L.; Vandekerckhove, J.; Tavernier, J.; Seguret-Mace, S.; et al. A proposed architecture for lecithin cholesterol acyl transferase (LCAT): Identification of the catalytic triad and molecular modeling. Protein Sci. 1998, 7, 587–599. [Google Scholar] [CrossRef]
- Glukhova, A.; Hinkovska-Galcheva, V.; Kelly, R.; Abe, A.; Shayman, J.A.; Tesmer, J.J. Structure and function of lysosomal phospholipase A2 and lecithin:cholesterol acyltransferase. Nat. Commun. 2015, 6, 6250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piper, D.E.; Romanow, W.G.; Gunawardane, R.N.; Fordstrom, P.; Masterman, S.; Pan, O.; Thibault, S.T.; Zhang, R.; Meininger, D.; Schwarz, M.; et al. The high-resolution crystal structure of human LCAT1. J. Lipid Res. 2015, 56, 1711–1719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Falarz, L.J.; Xu, Y.; Caldo, K.M.P.; Garroway, C.J.; Singer, S.D.; Chen, G. Characterization of the diversification of phospholipid:diacylglycerol acyltransferases in the green lineage. Plant J. 2020, 103, 2025–2038. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.-H.; Cai, Y.; Chai, J.; Schwender, J.; Shanklin, J. Expression of a Lychee PHOSPHATIDYLCHOLINE: DIACYLGLYCEROL CHOLINEPHOSPHOTRANSFERASE with an Escherichia coli CYCLOPROPANE SYNTHASE enhances cyclopropane fatty acid accumulation in Camelina Seeds. Plant Physiol. 2019, 180, 1351–1361. [Google Scholar] [CrossRef] [Green Version]
- Sigal, Y.J.; McDermott, M.I.; Morris, A.J. Integral membrane lipid phosphatases/phosphotransferases: Common structure and diverse functions. Biochem. J. 2005, 387, 281–293. [Google Scholar] [CrossRef]
- Bates, P.D.; Fatihi, A.; Snapp, A.R.; Carlsson, A.S.; Browse, J.; Lu, C. Acyl Editing and Headgroup Exchange Are the Major Mechanisms That Direct Polyunsaturated Fatty Acid Flux into Triacylglycerols. Plant Physiol. 2012, 160, 1530–1539. [Google Scholar] [CrossRef] [Green Version]
- Yang, W.; Wang, G.; Li, J.; Bates, P.D.; Wang, X.; Allen, D.K. Phospholipase Dζ Enhances Diacylglycerol Flux into Triacylglycerol. Plant Physiol. 2017, 174, 110–123. [Google Scholar] [CrossRef] [Green Version]
- Chen, G.Q.; Vang, L.; Lin, J.-T. Seed Development in Lesquerella fendleri (L.). HortScience 2009, 44, 1415–1418. [Google Scholar] [CrossRef] [Green Version]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef] [Green Version]
- Bolstad, B.; Irizarry, R.; Astrand, M.; Speed, T. A comparison of normalization methods for high density oligonucleotide array data based on variance and bias. Bioinformatics 2003, 19, 185–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kolde, R.; Laur, S.; Adler, P.; Vilo, J. Robust rank aggregation for gene list integration and meta-analysis. Bioinformatics 2012, 28, 573–580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef] [PubMed]
- Walley, J.W.; Shen, Z.; Sartor, R.; Wu, K.J.; Osborn, J.; Smith, L.G.; Briggs, S.P. Reconstruction of protein networks from an atlas of maize seed proteotypes. Proc. Natl. Acad. Sci. USA 2013, 110, E4808–E4817. [Google Scholar] [CrossRef] [Green Version]
- Schwender, J.; König, C.; Klapperstück, M.; Heinzel, N.; Munz, E.; Hebbelmann, I.; Hay, J.O.; Denolf, P.; De Bodt, S.; Redestig, H.; et al. Transcript abundance on its own cannot be used to infer fluxes in central metabolism. Front. Plant Sci. 2014, 5, 668. [Google Scholar] [CrossRef] [Green Version]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Developing Seed (DAP) a | Flower Bud | Leaf | Total | |||
|---|---|---|---|---|---|---|
| 21 | 35 | 42 | ||||
| raw reads | 129,121,576 | 99,873,058 | 127,855,980 | 99,729,366 | 96,034,602 | 552,614,582 |
| clean reads b | 125,026,944 | 97,400,124 | 123,850,518 | 98,065,392 | 95,366,514 | 539,709,492 |
| ratio c (%) | 96.8 | 97.5 | 96.9 | 98.3 | 99.3 | 97.7 |
| Number | |
|---|---|
| Numbers of unique genes transcripts (longest isoforms) | 85,948 |
| Numbers of transcripts (contigs) | 129,633 |
| Average length of transcripts (bp) | 814 (bp) |
| N50 a transcript size (bp) | 1373 (bp) |
| Total assembled bases | 105,504,114 (nt) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, G.Q.; Kim, W.N.; Johnson, K.; Park, M.-E.; Lee, K.-R.; Kim, H.U. Transcriptome Analysis and Identification of Lipid Genes in Physaria lindheimeri, a Genetic Resource for Hydroxy Fatty Acids in Seed Oil. Int. J. Mol. Sci. 2021, 22, 514. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22020514
Chen GQ, Kim WN, Johnson K, Park M-E, Lee K-R, Kim HU. Transcriptome Analysis and Identification of Lipid Genes in Physaria lindheimeri, a Genetic Resource for Hydroxy Fatty Acids in Seed Oil. International Journal of Molecular Sciences. 2021; 22(2):514. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22020514
Chicago/Turabian StyleChen, Grace Q., Won Nyeong Kim, Kumiko Johnson, Mid-Eum Park, Kyeong-Ryeol Lee, and Hyun Uk Kim. 2021. "Transcriptome Analysis and Identification of Lipid Genes in Physaria lindheimeri, a Genetic Resource for Hydroxy Fatty Acids in Seed Oil" International Journal of Molecular Sciences 22, no. 2: 514. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22020514