Fibulin-2: A Novel Biomarker for Differentiating Grade II from Grade I Meningiomas

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Results
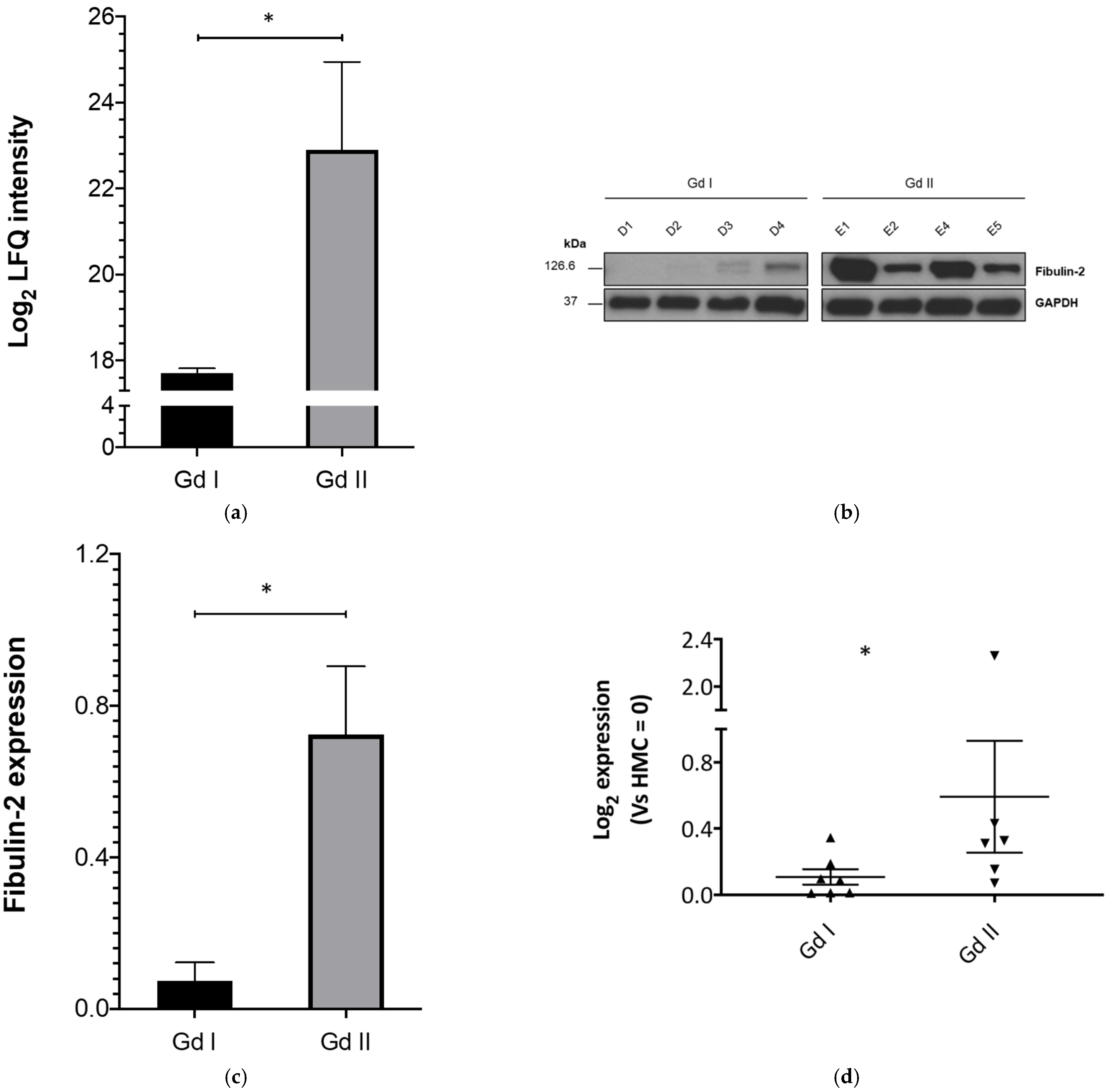
2.1. Fibulin-2 Is Significantly Differentially Expressed between Grade II and Grade I Primary Meningioma Cells
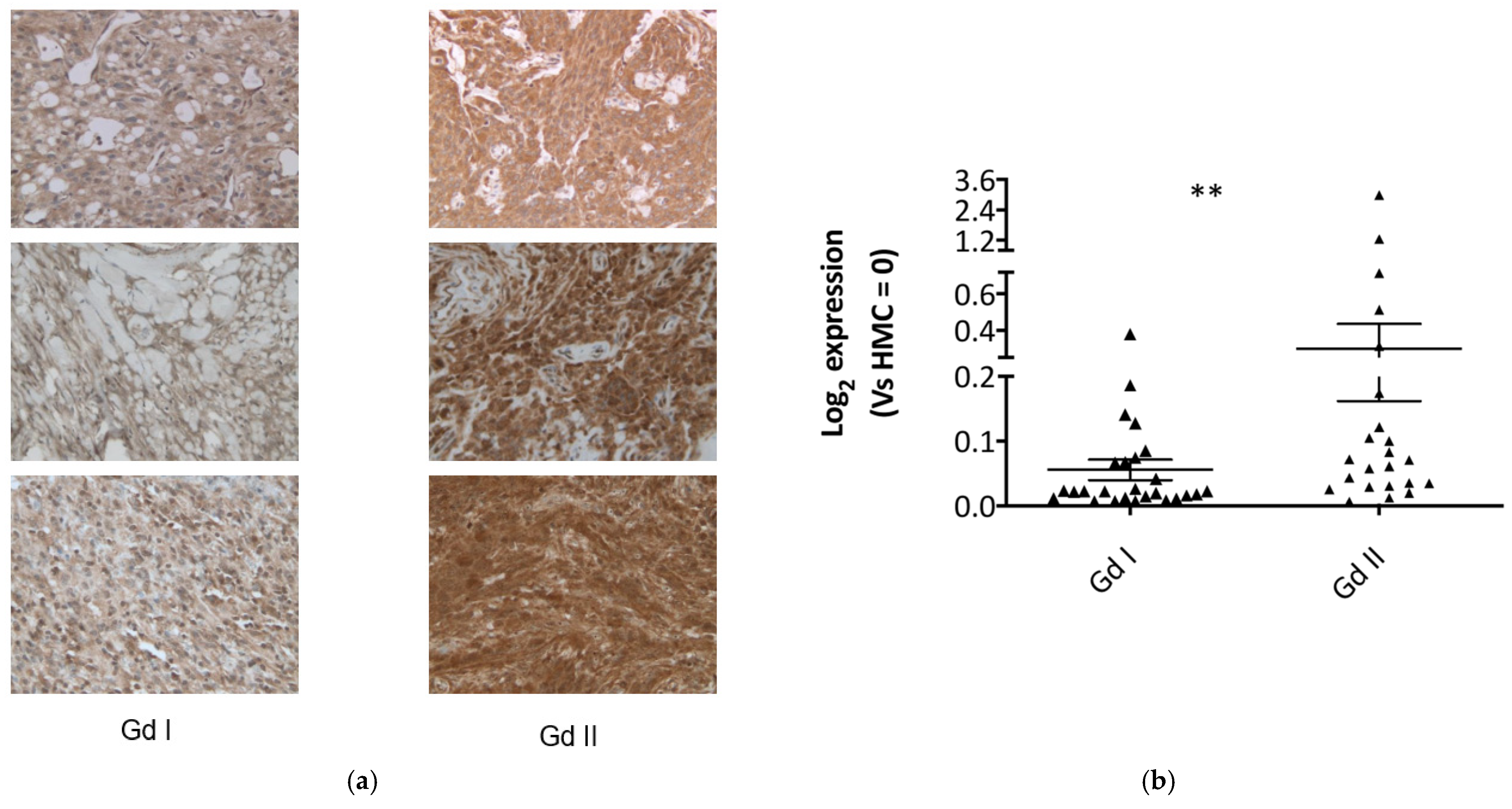
2.2. Fibulin-2 Is Significantly Overexpressed in Grade II Compared to Grade I Meningioma Tissues
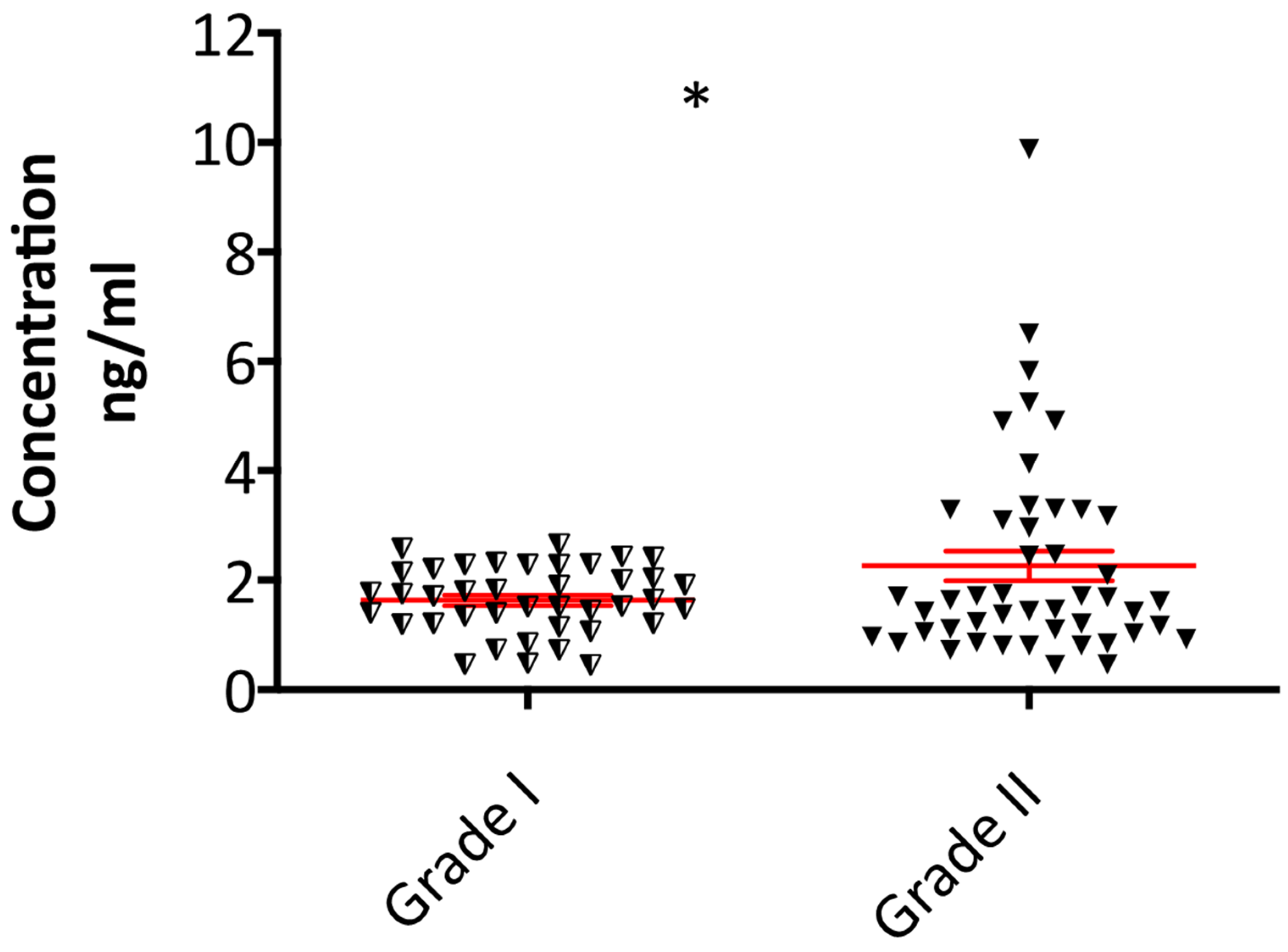
2.3. Plasma Fibulin-2 Levels Are Higher in Grade II Compared to Grade I Meningioma Patients
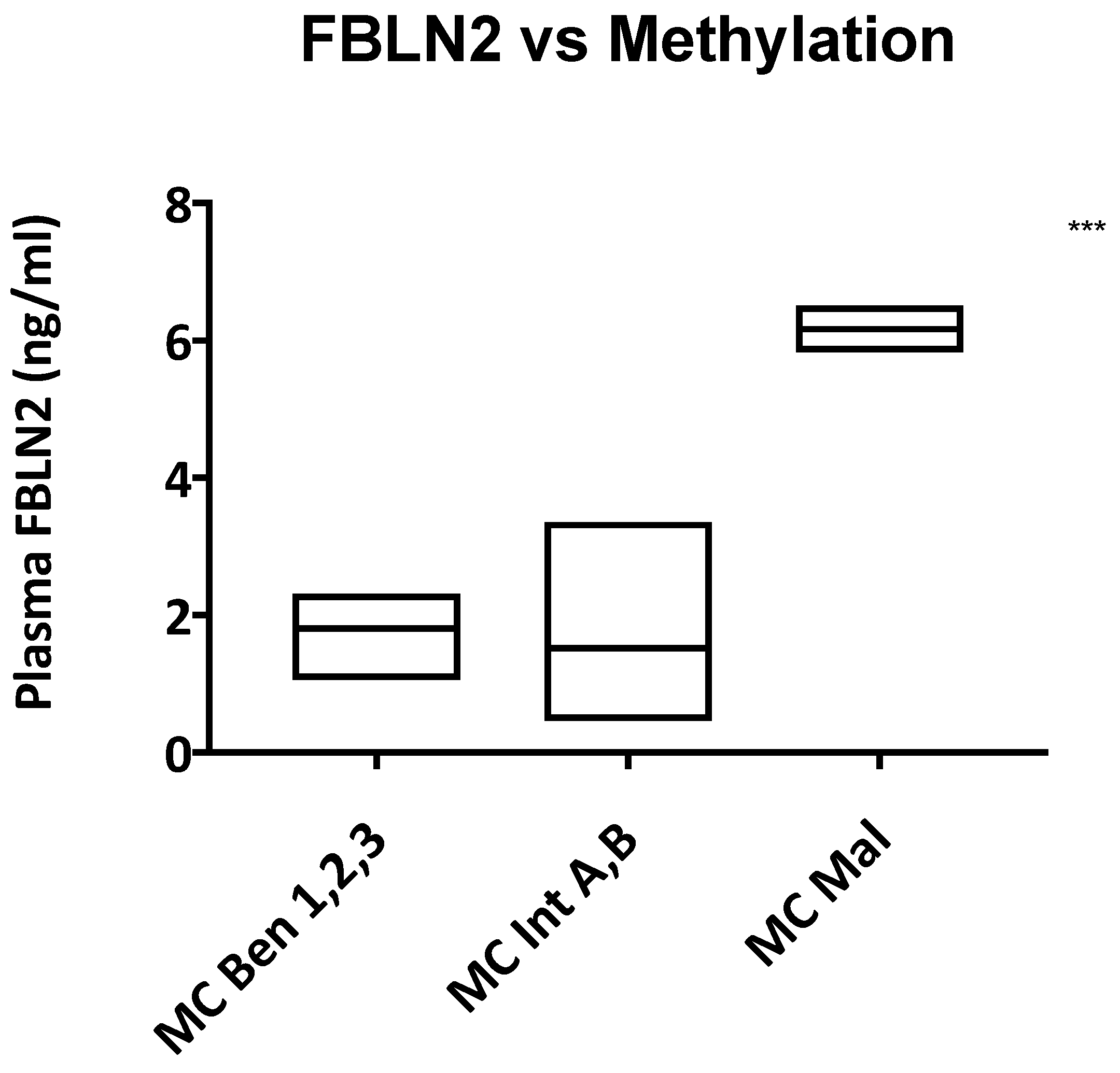
3. Discussion
4. Materials and Methods
4.1. Clinical Material and Ethical Approval
4.2. Meningioma Specimens, Tumour Digestion and Cell Culture
4.3. Western Blotting, Mass Spectrometry Proteomics and Immunohistochemistry
4.4. RNA Isolation and Gene Expression Analysis
4.5. Enzyme-Linked Immunosorbent Assay
4.6. Methylation Profiling
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| CNS | Central Nervous System |
| CDKN2A/B | Cyclin-Dependent Kinase Inhibitor 2A/2B |
| ECM | Extra-cellular Matrix |
| EDTA | Ethylenediaminetetraacetic acid |
| ELISA | Enzyme Linked Immuno-Sorbent Assay |
| FBLN2 | Fibulin-2 gene |
| Fc | Fold change |
| FFPE | Formalin-Fixed, Paraffin-Embedded |
| GAPDH | Glyceraldehyde 3-phosphate Dehydrogenase |
| Gd I | Grade I meningiomas |
| Gd II | Grade II meningiomas |
| MS | Mass spectrometry |
| MUC4 | Mucin 4 |
| NF2 | Neurofibromatosis Type 2 |
| SDS-PAGE | Sodium Dodecyl Sulfate–Polyacrylamide Gel Electrophoresis |
| RT-PCR | Reverse transcriptase polymerase chain reaction |
| RT-qPCR | Reverse transcriptase quantitative Polymerase Chain Reaction |
| WHO | World Health Organization |
References
- Norden, A.D.; Drappatz, J.; Wen, P.Y. Targeted drug therapy for meningiomas. Neurosurg. Focus 2007, 23, E12. [Google Scholar] [CrossRef]
- Kalamarides, M.; Stemmerrachamimov, A.O.; Niwakawakita, M.; Chareyre, F.; Taranchon, E.; Han, Z.-Y.; Martinelli, C.; Lusis, E.A.; Hegedus, B.; Gutmann, D.H.; et al. Identification of a progenitor cell of origin capable of generating diverse meningioma histological subtypes. Oncogene 2011, 30, 2333–2344. [Google Scholar] [CrossRef] [Green Version]
- Louis, D.N.; Perry, A.; Reifenberger, G.; Von Deimling, A.; Figarella-Branger, D.; Cavenee, W.K.; Ohgaki, H.; Wiestler, O.D.; Kleihues, P.; Ellison, D.W. The 2016 World Health Organization Classification of Tumors of the Central Nervous System: A summary. Acta Neuropathol. 2016, 131, 803–820. [Google Scholar] [CrossRef] [Green Version]
- Backer-Grøndahl, T.; Moen, B.H.; Torp, S.H. The histopathological spectrum of human meningiomas. Int. J. Clin. Exp. Pathol. 2012, 5, 231–242. [Google Scholar]
- Buerki, R.A.; Horbinski, C.; Kruser, T.; Horowitz, P.M.; James, C.D.; Lukas, R.V. An overview of meningiomas. Future Oncol. 2018, 14, 2161–2177. [Google Scholar] [CrossRef]
- Jenkinson, M.D.; Javadpour, M.; Haylock, B.J.; Young, B.; Gillard, H.; Vinten, J.; Bulbeck, H.; Das, K.; Farrell, M.; Looby, S.; et al. The ROAM/EORTC-1308 trial: Radiation versus Observation following surgical resection of Atypical Meningioma: Study protocol for a randomised controlled trial. Trials 2015, 16, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Mawrin, C.; Chung, C.; Preusser, M. Biology and Clinical Management Challenges in Meningioma. Am. Soc. Clin. Oncol. Educ. Book 2015, 35, e106–e115. [Google Scholar] [CrossRef] [PubMed]
- Bi, W.L.; Zhang, M.; Wu, W.W.; Mei, Y.; Dunn, I.F. Meningioma Genomics: Diagnostic, Prognostic, and Therapeutic Applications. Front. Surg. 2016, 3, 40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harter, P.N.; Braun, Y.; Plate, K.H. Classification of meningiomas—Advances and controversies. Chin. Clin. Oncol. 2017, 6, S2. [Google Scholar] [CrossRef]
- Zhang, Q.; Jia, G.-J.; Zhang, G.-B.; Wang, L.; Wu, Z.; Jia, W.; Hao, S.-Y.; Ni, M.; Li, D.; Wang, K.; et al. A Logistic Regression Model for Detecting the Presence of Malignant Progression in Atypical Meningiomas. World Neurosurg. 2019, 126, e392–e401. [Google Scholar] [CrossRef]
- Katz, L.M.; Hielscher, T.; Liechty, B.; Silverman, J.; Zagzag, D.; Sen, R.; Wu, P.; Golfinos, J.G.; Reuss, D.; Neidert, M.C.; et al. Loss of histone H3K27me3 identifies a subset of meningiomas with increased risk of recurrence. Acta Neuropathol. 2018, 135, 955–963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dunn, J.; Ferluga, S.; Sharma, V.; Futschik, M.; Hilton, D.A.; Adams, C.L.; Lasonder, E.; Hanemann, C.O. Proteomic analysis discovers the differential expression of novel proteins and phosphoproteins in meningioma including NEK9, HK2 and SET and deregulation of RNA metabolism. EBioMedicine 2018, 40, 77–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Champeaux, C.; Houston, D.; Dunn, L.; Resche-Rigon, M. Intracranial WHO grade I meningioma: A competing risk analysis of progression and disease-specific survival. Acta Neurochir. 2019, 161, 2541–2549. [Google Scholar] [CrossRef] [PubMed]
- Barresi, V.; Lionti, S.; Caliri, S.; Caffo, M. Histopathological features to define atypical meningioma: What does really matter for prognosis? Brain Tumor Pathol. 2018, 35, 168–180. [Google Scholar] [CrossRef] [PubMed]
- Barrett, O.C.; Hackney, J.R.; McDonald, A.M.; Willey, C.D.; Bredel, M.; Fiveash, J.B. Pathologic Predictors of Local Recurrence in Atypical Meningiomas Following Gross Total Resection. Int. J. Radiat. Oncol. 2019, 103, 453–459. [Google Scholar] [CrossRef]
- Budohoski, K.P.; Clerkin, J.; Millward, C.P.; O’Halloran, P.J.; Waqar, M.; Looby, S.; Young, A.M.H.; Guilfoyle, M.R.; Fitzroll, D.; Devadass, A.; et al. Predictors of early progression of surgically treated atypical meningiomas. Acta Neurochir. 2018, 160, 1813–1822. [Google Scholar] [CrossRef] [Green Version]
- Nowak, A.; Dziedzic, T.A.; Krych, P.; Czernicki, T.; Kunert, P.; Marchel, A. Benign versus atypical meningiomas: Risk factors predicting recurrence. Neurol. Neurochir. Pol. 2015, 49, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Sun, S.Q.; Kim, A.H.; Cai, C.; Murphy, R.K.; DeWees, T.; Sylvester, P.; Dacey, R.G.; Grubb, R.L.; Rich, K.M.; Zipfel, G.J.; et al. Management of atypical cranial meningiomas, part 1: Predictors of recurrence and the role of adjuvant radiation after gross total resection. Neurosurgery 2014, 75, 347–354. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.; Niemierko, A.; Hwang, W.L.; Stemmer-Rachamimov, A.O.; Curry, W.T.; Barker, F.G.; Martuza, R.L.; Oh, K.S.; Loeffler, J.S.; Shih, H.A. Histopathological prognostic factors of recurrence following definitive therapy for atypical and malignant meningiomas. J. Neurosurg. 2018, 128, 1123–1132. [Google Scholar] [CrossRef]
- Vranic, A.; Popović, M.; Cör, A.; Prestor, B.; Pižem, J. Mitotic Count, Brain Invasion, and Location Are Independent Predictors of Recurrence-Free Survival in Primary Atypical and Malignant Meningiomas: A Study of 86 Patients. Neurosurgery 2010, 67, 1124–1132. [Google Scholar] [CrossRef] [Green Version]
- Hardesty, D.A.; Wolf, A.B.; Brachman, D.G.; McBride, H.L.; Youssef, E.; Nakaji, P.; Porter, R.W.; Smith, K.A.; Spetzler, R.F.; Sanai, N. The impact of adjuvant stereotactic radiosurgery on atypical meningioma recurrence following aggressive microsurgical resection. J. Neurosurg. 2013, 119, 475–481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Hui, D.; Fu, X. Roles of Fibulin-2 in Carcinogenesis. Med. Sci. Monit. 2020, 26, e918099-1. [Google Scholar] [CrossRef] [PubMed]
- Baird, B.N.; Schliekelman, M.J.; Ahn, Y.-H.; Chen, Y.; Roybal, J.D.; Gill, B.J.; Mishra, D.K.; Erez, B.; O’Reilly, M.; Yang, Y.; et al. Fibulin-2 Is a Driver of Malignant Progression in Lung Adenocarcinoma. PLoS ONE 2013, 8, e67054. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yi, C.-H.; Smith, D.J.; West, W.W.; Hollingsworth, M.A. Loss of Fibulin-2 Expression Is Associated with Breast Cancer Progression. Am. J. Pathol. 2007, 170, 1535–1545. [Google Scholar] [CrossRef] [Green Version]
- Matsuyama, A.; Jotatsu, M.; Uchihashi, K.; Tsuda, Y.; Shiba, E.; Haratake, J.; Hisaoka, M. MUC4 expression in meningiomas: Under-recognized immunophenotype particularly in meningothelial and angiomatous subtypes. Histopathology 2019, 74, 276–283. [Google Scholar] [CrossRef]
- Abbritti, R.V.; Polito, F.; Cucinotta, M.; Giudice, C.L.; Caffo, M.; Tomasello, C.; Germanò, A.; Aguennouz, M. Meningiomas and Proteomics: Focus on New Potential Biomarkers and Molecular Pathways. Cancer Genom. Proteom. 2016, 13, 369–380. [Google Scholar]
- Caffo, M.; Caruso, G.; Galatioto, S.; Meli, F.; Cacciola, F.; Germanò, A.; Alafaci, C.; Tomasello, F. Immunohistochemical study of the extracellular matrix proteins laminin, fibronectin and type IV collagen in secretory meningiomas. J. Clin. Neurosci. 2008, 15, 806–811. [Google Scholar] [CrossRef]
- Law, E.W.L.; Cheung, A.K.L.; Kashuba, V.I.; Pavlova, T.V.; Zabarovsky, E.R.; Lung, H.L.; Cheng, Y.; Chua, D.; Kwong, D.L.-W.; Tsao, S.W.; et al. Anti-angiogenic and tumor-suppressive roles of candidate tumor-suppressor gene, Fibulin-2, in nasopharyngeal carcinoma. Oncogene 2012, 31, 728–738. [Google Scholar] [CrossRef] [Green Version]
- Wiemels, J.L.; Wrensch, M.; Claus, E.B. Epidemiology and etiology of meningioma. J. Neuro-Oncol. 2010, 99, 307–314. [Google Scholar] [CrossRef] [Green Version]
- Harmancı, A.S.; Youngblood, M.W.; Clark, V.E.; Coşkun, S.; Henegariu, O.; Duran, D.; Erson-Omay, E.Z.; Kaulen, L.D.; Lee, T.I.; Abraham, B.J.; et al. Integrated genomic analyses of de novo pathways underlying atypical meningiomas. Nat. Commun. 2017, 8, 14433. [Google Scholar] [CrossRef]
- Ülgen, E.; Bektaşoğlu, P.K.; Sav, M.A.; Can, Ö.; Danyeli, A.E.; Hızal, D.B.; Pamir, M.N.; Özduman, K. Meningiomas Display a Specific Immunoexpression Pattern in a Rostrocaudal Gradient: An Analysis of 366 Patients. World Neurosurg. 2019, 123, e520–e535. [Google Scholar] [CrossRef] [PubMed]
- Magill, S.; Vasudevan, H.N.; Seo, K.; Villanueva-Meyer, J.E.; Choudhury, A.; Liu, S.; Pekmezci, M.; Findakly, S.; Hilz, S.; Lastella, S.; et al. Multiplatform genomic profiling and magnetic resonance imaging identify mechanisms underlying intratumor heterogeneity in meningioma. Nat. Commun. 2020, 11, 1–15. [Google Scholar] [CrossRef]
- Ferluga, S.; Baiz, D.; Hilton, D.A.; Adams, C.L.; Ercolano, E.; Dunn, J.; Bassiri, K.; Kurian, K.M.; Hanemann, C.O. Constitutive activation of the EGFR–STAT1 axis increases proliferation of meningioma tumor cells. Neuro-Oncol. Adv. 2020, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaur, G.; Sayegh, E.T.; Larson, A.; Bloch, O.; Madden, M.; Sun, M.Z.; Barani, I.J.; James, C.D.; Parsa, A.T. Adjuvant radiotherapy for atypical and malignant meningiomas: A systematic review. Neuro-Oncology 2014, 16, 628–636. [Google Scholar] [CrossRef] [Green Version]
- Champeaux, C.; Dunn, L. World Health Organization grade II meningiomas. Acta Neurochir. 2016, 158, 921–929. [Google Scholar] [CrossRef]
- Rauch, U.; Zhou, X.-H.; Roos, G. Extracellular matrix alterations in brains lacking four of its components. Biochem. Biophys. Res. Commun. 2005, 328, 608–617. [Google Scholar] [CrossRef] [Green Version]
- Miosge, N.; Götz, W.; Sasaki, T.; Chu, M.L.; Timpl, R.; Herken, R. The extracellular matrix proteins fibulin-1 and fibulin-2 in the early human embryo. Histochem. J. 1996, 28, 109–116. [Google Scholar] [CrossRef]
- Schaeffer, J.; Tannahill, D.; Cioni, J.-M.; Rowlands, D.; Keynes, R. Identification of the extracellular matrix protein Fibulin-2 as a regulator of spinal nerve organization. Dev. Biol. 2018, 442, 101–114. [Google Scholar] [CrossRef]
- Tsuda, T.; Wu, J.; Gao, E.; Joyce, J.; Markova, D.; Dong, H.; Liu, Y.; Zhang, H.; Zou, Y.; Gao, F.; et al. Loss of fibulin-2 protects against progressive ventricular dysfunction after myocardial infarction. J. Mol. Cell. Cardiol. 2012, 52, 273–282. [Google Scholar] [CrossRef] [Green Version]
- Schliekelman, M.J.; Gibbons, D.L.; Faca, V.M.; Creighton, C.J.; Rizvi, Z.H.; Zhang, Q.; Wong, C.-H.; Wang, H.; Ungewiss, C.; Ahn, Y.-H.; et al. Targets of the Tumor Suppressor miR-200 in Regulation of the Epithelial–Mesenchymal Transition in Cancer. Cancer Res. 2011, 71, 7670–7682. [Google Scholar] [CrossRef] [Green Version]
- Ramaswamy, S.; Ross, K.N.; Lander, E.S.; Golub, T.R. A molecular signature of metastasis in primary solid tumors. Nat. Genet. 2003, 33, 49–54. [Google Scholar] [CrossRef] [PubMed]
- Dandia, H.; Tayalia, P. Immunomodulation via macrophages to fight solid tumor malignancies. FEBS J. 2020. [Google Scholar] [CrossRef] [PubMed]
- Kochi, N.; Tani, E.; Morimura, T.; Itagaki, T. Immunohistochemical study of fibronectin in human glioma and meningioma. Acta Neuropathol. 1983, 59, 119–126. [Google Scholar] [CrossRef] [PubMed]
- Corniola, M.V.; Lemée, J.-M.; Meling, T.R. Histological transformation in recurrent WHO grade I meningiomas. Sci. Rep. 2020, 10, 11220. [Google Scholar] [CrossRef] [PubMed]
- Adams, C.L.; Ercolano, E.; Ferluga, S.; Sofela, A.; Dave, F.; Negroni, C.; Kurian, K.M.; Hilton, D.A.; Hanemann, C.O. A Rapid Robust Method for Subgrouping Non-NF2 Meningiomas According to Genotype and Detection of Lower Levels of M2 Macrophages in AKT1 E17K Mutated Tumours. Int. J. Mol. Sci. 2020, 21, 1273. [Google Scholar] [CrossRef] [Green Version]
- Bassiri, K.; Ferluga, S.; Sharma, V.; Syed, N.; Adams, C.L.; Lasonder, E.; Hanemann, C.O. Global Proteome and Phospho-proteome Analysis of Merlin-deficient Meningioma and Schwannoma Identifies PDLIM2 as a Novel Therapeutic Target. EBioMedicine 2017, 16, 76–86. [Google Scholar] [CrossRef] [Green Version]
- Rappsilber, J.; Ishihama, Y.; Mann, M. Stop and Go Extraction Tips for Matrix-Assisted Laser Desorption/Ionization, Nanoelectrospray, and LC/MS Sample Pretreatment in Proteomics. Anal. Chem. 2003, 75, 663–670. [Google Scholar] [CrossRef]
- Hilton, D.A.; Ristic, N.; Hanemann, C.O. Activation of ERK, AKT and JNK signalling pathways in human schwannomas in situ. Histopathology 2009, 55, 744–749. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Sahm, F.; Schrimpf, D.; Stichel, D.; Jones, D.T.W.; Hielscher, T.; Schefzyk, S.; Okonechnikov, K.; Koelsche, C.; Reuss, D.E.; Capper, D.; et al. DNA methylation-based classification and grading system for meningioma: A multicentre, retrospective analysis. Lancet Oncol. 2017, 18, 682–694. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Clinical Features | Group | Patients | |
|---|---|---|---|
| n | (%) | ||
| Sex | Female | 123 | 69.1 |
| Male | 54 | 30.3 | |
| Unknown | 1 | 0.6 | |
| Age | Median Range | 59 27–92 | |
| WHO grade | WHO I | 87 | 48.9 |
| WHO II | 91 | 51.1 | |
| Grade I | Meningothelial | 27 | 31.0 |
| Fibrous | 25 | 28.7 | |
| Transitional | 17 | 19.5 | |
| Psammomatous | 6 | 6.9 | |
| Angiomatous | 1 | 1.1 | |
| Secretory | 1 | 1.1 | |
| Metaplastic | 1 | 1.1 | |
| Not reported | 6 | 6.9 | |
| Meningothelial + bony invasion | 1 | 1.1 | |
| Mixed | 2 | 2.3 | |
| Chordoid | 7 | 7.7 | |
| Atypical | 71 | 78.0 | |
| Clear cell | 1 | 1.1 | |
| Atypical + brain invasion | 11 | 12.1 | |
| Atypical + bone/dura invasion | 2 | 2.2 | |
| Chordoid + dura invasion | 1 | 1.1 | |
| Grade II | Atypical with extracranial extension | 1 | 1.1 |
| Atypical + focal rhabdoid features | 1 | 1.1 | |
| Location | Falcine | 10 | 5.4 |
| Cerebello-pontine angle | 4 | 2.2 | |
| Parasagittal | 15 | 8.1 | |
| Olfactory groove | 11 | 5.9 | |
| Frontal | 42 | 22.7 | |
| Occipital | 17 | 9.2 | |
| Intraventricular | 2 | 1.1 | |
| Spheno-orbital | 3 | 1.6 | |
| Sphenoid wing | 9 | 4.9 | |
| Tentorial | 4 | 2.2 | |
| Parietal | 9 | 4.9 | |
| Temporal | 9 | 4.9 | |
| Posterior fossa (not specified) | 9 | 4.9 | |
| Planum sphenoidale | 6 | 3.2 | |
| Fronto-parietal | 13 | 7.0 | |
| Fronto-temporal | 2 | 1.1 | |
| Petrous temporal (incl. petro-clival) | 3 | 1.6 | |
| Cavernous sinus | 1 | 0.5 | |
| Planum sphenoidale | 6 | 3.2 | |
| Thoracic | 2 | 1.1 | |
| Skull base (not specified) | 1 | 0.5 | |
| Not specified/stated | 7 | 3.8 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sofela, A.A.; Hilton, D.A.; Ammoun, S.; Baiz, D.; Adams, C.L.; Ercolano, E.; Jenkinson, M.D.; Kurian, K.M.; Teo, M.; Whitfield, P.C.; et al. Fibulin-2: A Novel Biomarker for Differentiating Grade II from Grade I Meningiomas. Int. J. Mol. Sci. 2021, 22, 560. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22020560
Sofela AA, Hilton DA, Ammoun S, Baiz D, Adams CL, Ercolano E, Jenkinson MD, Kurian KM, Teo M, Whitfield PC, et al. Fibulin-2: A Novel Biomarker for Differentiating Grade II from Grade I Meningiomas. International Journal of Molecular Sciences. 2021; 22(2):560. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22020560
Chicago/Turabian StyleSofela, Agbolahan A., David A. Hilton, Sylwia Ammoun, Daniele Baiz, Claire L. Adams, Emanuela Ercolano, Michael D. Jenkinson, Kathreena M. Kurian, Mario Teo, Peter C. Whitfield, and et al. 2021. "Fibulin-2: A Novel Biomarker for Differentiating Grade II from Grade I Meningiomas" International Journal of Molecular Sciences 22, no. 2: 560. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22020560