Expression Levels of an Alpha-Synuclein Transcript in Blood May Distinguish between Early Dementia with Lewy Bodies and Parkinson’s Disease

 ,
,  , , , ,
, , , ,
Abstract
:1. Introduction
2. Results
2.1. Characteristics of Post-Mortem Cases
2.2. Demographic and Clinical Data of Participants
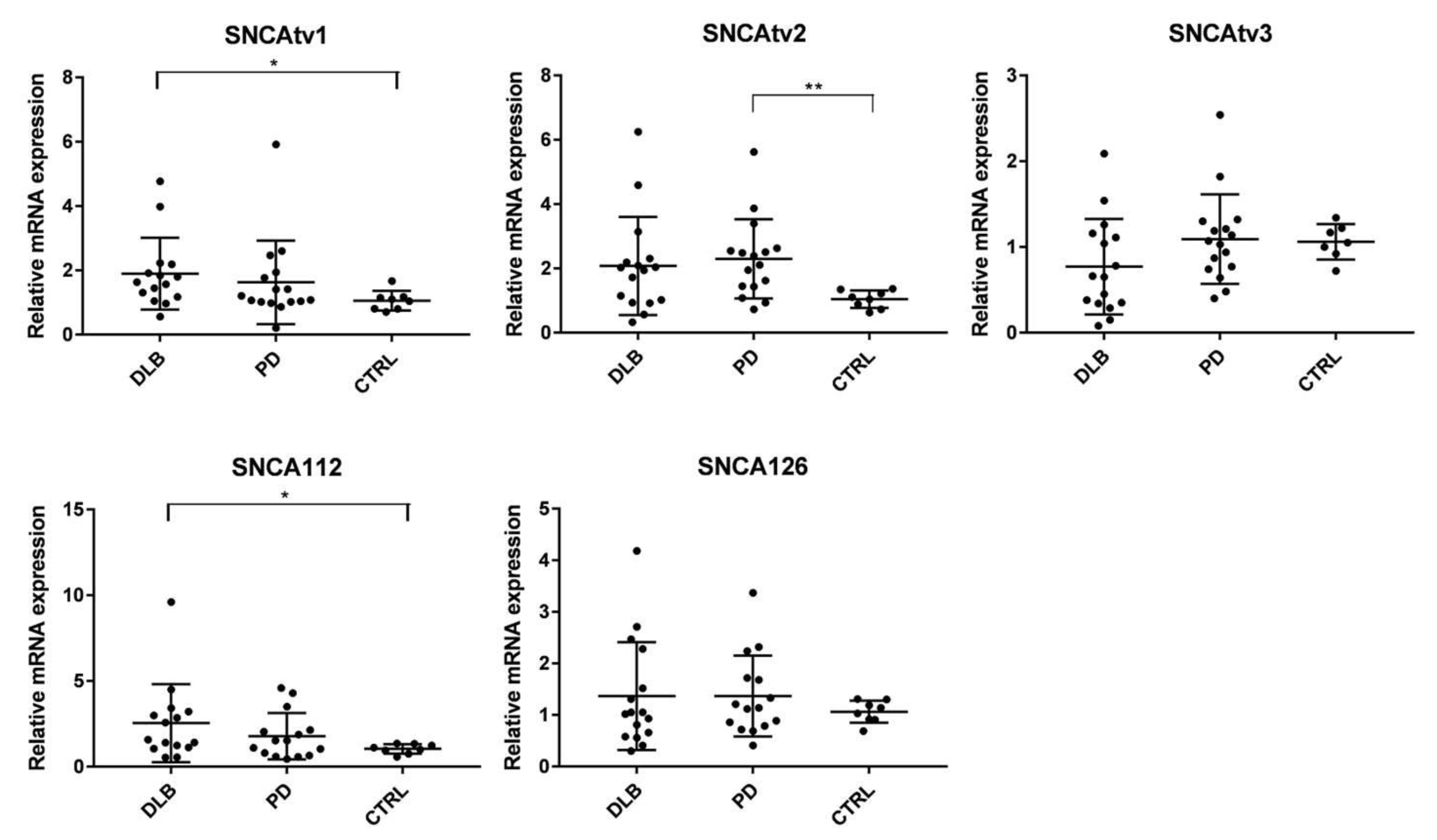
2.3. Relative SNCA Isoform Expression Levels in Different Brain Regions and Blood
2.4. SNCA Transcript Expression in the Temporal Cortex
2.5. Correlation between the Expression of SNCA Transcript Variants in Brain
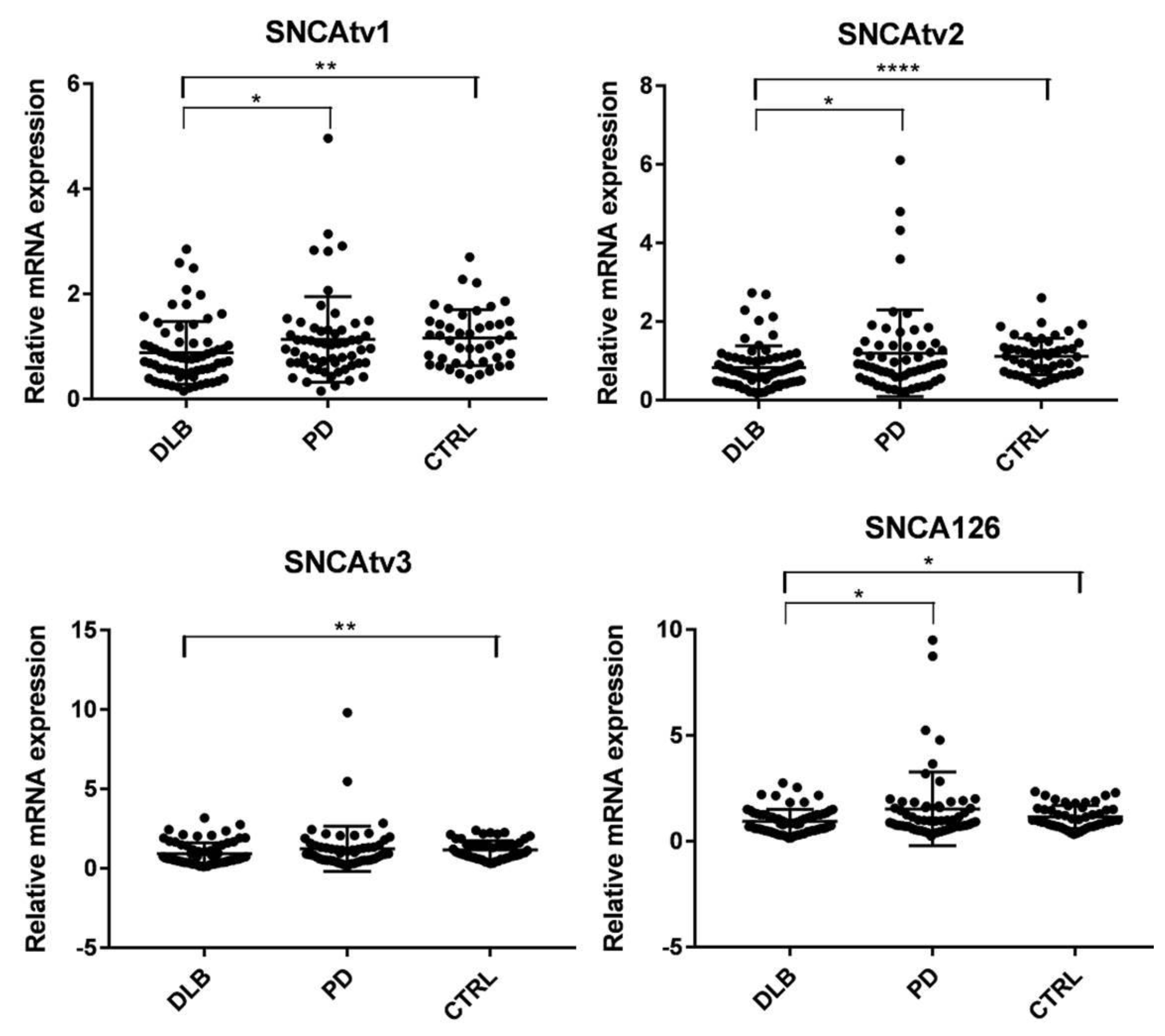
2.6. SNCA Transcript Expression in Blood
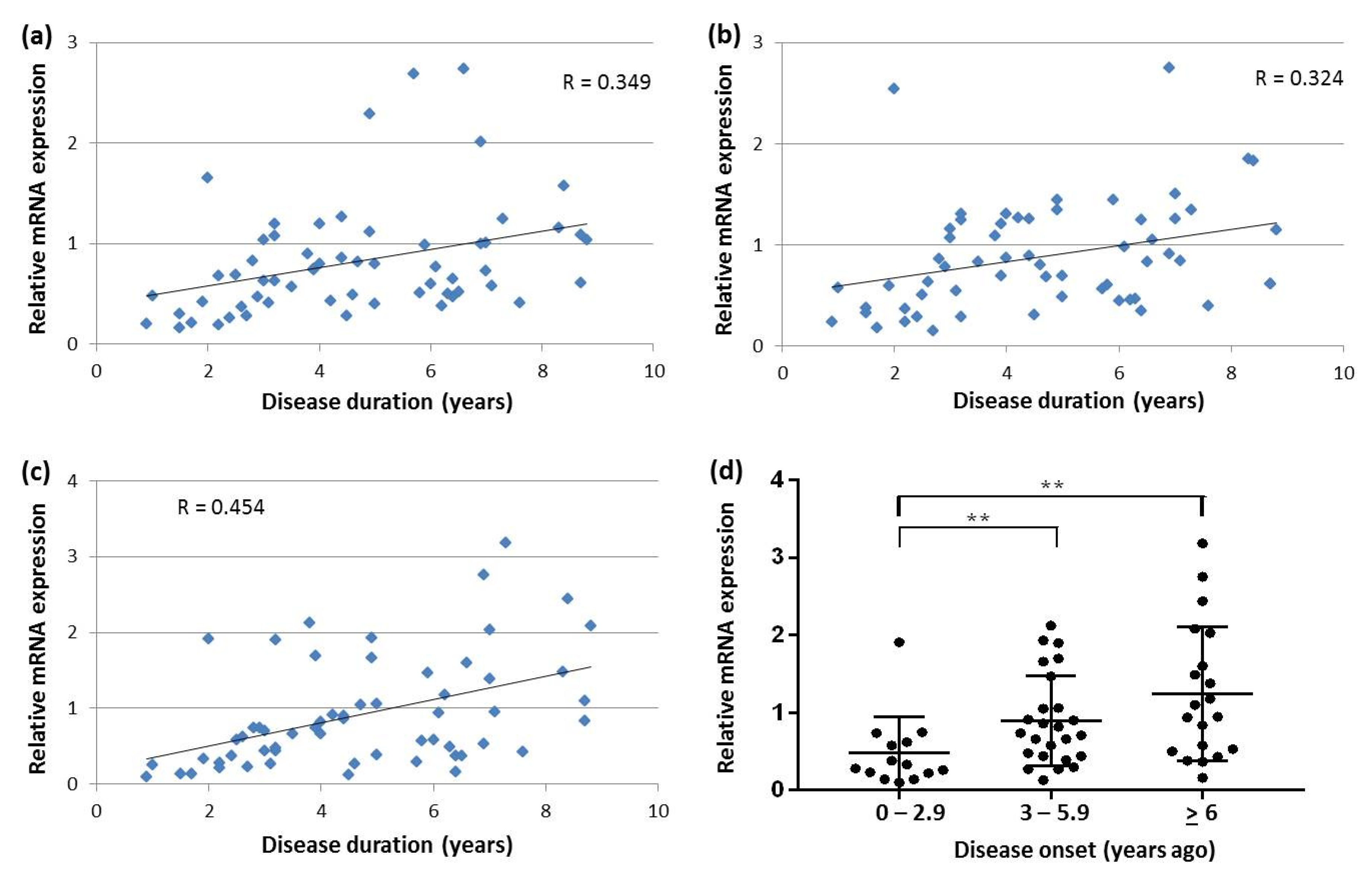
2.7. Correlation between SNCA Transcript Expression, Disease Duration and Age-at-Onset in Blood
3. Discussion
4. Materials and Methods
4.1. Source of Tissues
4.2. Participants
4.3. RNA Purification, Reverse Transcription, and Assessment of mRNA Stability
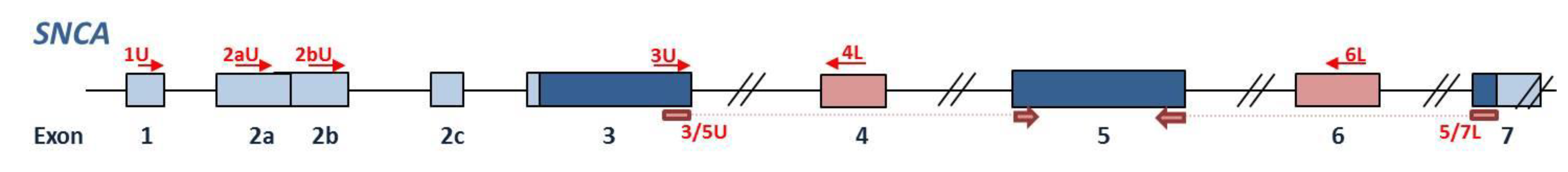
4.4. Primer Design
4.5. Real Time PCR
4.6. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| DLB | dementia with Lewy bodies |
| PD | Parkinson’s disease |
| LBD | Lewy body disorders |
| LBP | Lewy body pathology |
| AD | Alzheimer disease |
| AS | alpha-synuclein |
| LB | Lewy body |
| PDD | PD dementia |
| CSF | cerebrospinal fluid |
| SNCA | alpha-synuclein gene |
| tv | transcript variant |
| UTR | untranslated region |
| CTRL | Control subjects |
References
- Spillantini, M.G.; Crowther, R.A.; Jakes, R.; Hasegawa, M.; Goedert, M. α-Synuclein in filamentous inclusions of Lewy bodies from Parkinson’s disease and dementia with Lewy bodies. Proc. Natl. Acad. Sci. USA 1998, 95, 6469–6473. [Google Scholar] [CrossRef] [Green Version]
- Braak, H.; Braak, E. Diagnostic Criteria for Neuropathologic Assessment of Alzheimer’s Disease. Neurobiol. Aging 1997, 18, S85–S88. [Google Scholar] [CrossRef]
- Jellinger, K.A. A critical reappraisal of current staging of Lewy-related pathology in human brain. Acta Neuropathol. 2008, 116, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Jellinger, K.A. A critical evaluation of current staging of α-synuclein pathology in Lewy body disorders. Biochim. Biophys. Acta BBA Mol. Basis Dis. 2009, 1792, 730–740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferman, T.J.; Boeve, B.F. Dementia with Lewy Bodies. Neurol. Clin. 2007, 25, 741–760. [Google Scholar] [CrossRef]
- McKeith, I.G.; Boeve, B.F.; Dickson, D.W.; Halliday, G.; Taylor, J.-P.; Weintraub, D.; Aarsland, D.; Galvin, J.; Attems, J.; Ballard, C.G.; et al. Diagnosis and management of dementia with Lewy bodies: Fourth consensus report of the DLB Consortium. Neurology 2017, 89, 88–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanyu, H.; Sato, T.; Hirao, K.; Kanetaka, H.; Sakurai, H.; Iwamoto, T. Differences in clinical course between dementia with Lewy bodies and Alzheimer’s disease. Eur. J. Neurol. 2009, 16, 212–217. [Google Scholar] [CrossRef]
- Caballol, N.; Martí, M.J.; Tolosa, E. Cognitive dysfunction and dementia in Parkinson disease. Mov. Disord. 2007, 22, S358–S366. [Google Scholar] [CrossRef]
- Jellinger, K.A. Dementia with Lewy bodies and Parkinson’s disease-dementia: Current concepts and controversies. J. Neural Transm. 2018, 125, 615–650. [Google Scholar] [CrossRef]
- Colom-Cadena, M.; Grau-Rivera, O.; Planellas, L.; Cerquera, C.; Morenas, E.; Helgueta, S.; Muñoz, L.; Kulisevsky, J.; Martí, M.J.; Tolosa, E.; et al. Regional Overlap of Pathologies in Lewy Body Disorders. J. Neuropathol. Exp. Neurol. 2017, 76, 216–224. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, R.M.; Ke, Y.D.; Vucic, S.; Ittner, L.M.; Seeley, W.; Hodges, J.R.; Piguet, O.; Halliday, G.M.; Kiernan, M.C. Physiological changes in neurodegeneration—mechanistic insights and clinical utility. Nat. Rev. Neurol. 2018, 14, 259–271. [Google Scholar] [CrossRef]
- Zetterberg, H. Applying fluid biomarkers to Alzheimer’s disease. Am. J. Physiol. Physiol. 2017, 313, C3–C10. [Google Scholar] [CrossRef] [Green Version]
- Parnetti, L.; Paciotti, S.; Farotti, L.; Bellomo, G.; Sepe, F.N.; Eusebi, P. Parkinson’s and Lewy body dementia CSF biomarkers. Clin. Chim. Acta 2019, 495, 318–325. [Google Scholar] [CrossRef] [PubMed]
- Ingelsson, M. Alpha-Synuclein Oligomers—Neurotoxic Molecules in Parkinson’s Disease and Other Lewy Body Disorders. Front. Neurosci. 2016, 10, 408. [Google Scholar] [CrossRef] [Green Version]
- Uversky, V.N. Looking at the recent advances in understanding α-synuclein and its aggregation through the proteoform prism. F1000 Res. 2017, 6, 525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiba-Falek, O.; Lopez, G.J.; Nussbaum, R.L. Levels of alpha-synuclein mRNA in sporadic Parkinson disease patients. Mov. Disord. 2006, 21, 1703–1708. [Google Scholar] [CrossRef] [PubMed]
- Gründemann, J.; Schlaudraff, F.; Haeckel, O.; Liss, B. Elevated α-synuclein mRNA levels in individual UV-laser-microdissected dopaminergic substantia nigra neurons in idiopathic Parkinson’s disease. Nucleic Acids Res. 2008, 36, e38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ibáñez, P.; Bonnet, A.-M.; Débarges, B.; Lohmann, E.; Tison, F.; Agid, Y.; Dürr, A.; Brice, A.; Pollak, P. Causal relation between α-synuclein locus duplication as a cause of familial Parkinson’s disease. Lancet 2004, 364, 1169–1171. [Google Scholar] [CrossRef]
- Chartier-Harlin, M.-C.; Kachergus, J.; Roumier, C.; Mouroux, V.; Douay, X.; Lincoln, S.; Levecque, C.; Larvor, L.; Andrieux, J.; Hulihan, M.; et al. α-synuclein locus duplication as a cause of familial Parkinson’s disease. Lancet 2004, 364, 1167–1169. [Google Scholar] [CrossRef]
- Miller, D.W.; Hague, S.M.; Clarimón, J.; Baptista, M.; Gwinn, K.; Cookson, M.R.; Singleton, A.B. α-Synuclein in blood and brain from familial Parkinson disease with SNCA locus triplication. Neurology 2004, 62, 1835–1838. [Google Scholar] [CrossRef]
- Mollenhauer, B.; Caspell-Garcia, C.J.; Coffey, C.S.; Taylor, P.; Singleton, A.; Shaw, L.M.; Trojanowski, J.Q.; Frasier, M.; Simuni, T.; Iranzo, A.; et al. Longitudinal analyses of cerebrospinal fluid α-Synuclein in prodromal and early Parkinson’s disease. Mov. Disord. 2019, 34, 1354–1364. [Google Scholar] [CrossRef] [PubMed]
- Marek, K.; Chowdhury, S.; Siderowf, A.; Lasch, S.; Coffey, C.S.; Caspell-Garcia, C.; Simuni, T.; Jennings, D.; Tanner, C.M.; Trojanowski, J.Q.; et al. The Parkinson’s progression markers initiative (PPMI)—establishing a PD biomarker cohort. Ann. Clin. Transl. Neurol. 2018, 5, 1460–1477. [Google Scholar] [CrossRef] [PubMed]
- Foulds, P.G.; Diggle, P.; Mitchell, J.D.; Parker, A.; Hasegawa, M.; Masuda-Suzukake, M.; Mann, D.M.A.; Allsop, D. A longitudinal study on α-synuclein in blood plasma as a biomarker for Parkinson’s disease. Sci. Rep. 2013, 3, srep02540. [Google Scholar] [CrossRef] [PubMed]
- Gámez-Valero, A.; Beyer, K. Alternative Splicing of Alpha- and Beta-Synuclein Genes Plays Differential Roles in Synucleinopathies. Genes 2018, 9, 63. [Google Scholar] [CrossRef] [Green Version]
- Beyer, K.; Ariza, A. Alpha-Synuclein Posttranslational Modification and Alternative Splicing as a Trigger for Neurodegeneration. Mol. Neurobiol. 2013, 47, 509–524. [Google Scholar] [CrossRef]
- Rhinn, H.; Qiang, L.; Yamashita, T.; Rhee, D.; Zolin, A.; Vanti, W.B.; Abeliovich, A. Alternative α-synuclein transcript usage as a convergent mechanism in Parkinson’s disease pathology. Nat. Commun. 2012, 3, 1–11. [Google Scholar] [CrossRef]
- Braak, H.; Braak, E. Neuropathological stageing of Alzheimer-related changes. Acta Neuropathol. 1991, 82, 239–259. [Google Scholar] [CrossRef]
- Braak, H.; Del Tredici, K.; Rüb, U.; De Vos, R.A.; Steur, E.N.J.; Braak, E. Staging of brain pathology related to sporadic Parkinson’s disease. Neurobiol. Aging 2003, 24, 197–211. [Google Scholar] [CrossRef]
- McKeith, I.G.; Dickson, D.W.; Lowe, J.; Emre, M.; O’Brien, J.T.; Feldman, H.; Cummings, J.; Duda, J.E.; Lippa, C.; Perry, E.K.; et al. Diagnosis and management of dementia with Lewy bodies: Third report of the DLB consortium. Neurology 2005, 65, 1863–1872. [Google Scholar] [CrossRef] [Green Version]
- Paleologou, K.E.; Kragh, C.L.; Mann, D.M.A.; Salem, S.A.; Al-Shami, R.; Allsop, D.; Hassan, A.H.; Jensen, P.H.; El-Agnaf, O.M.A. Detection of elevated levels of soluble -synuclein oligomers in post-mortem brain extracts from patients with dementia with Lewy bodies. Brain 2009, 132, 1093–1101. [Google Scholar] [CrossRef] [Green Version]
- Vaikath, N.N.; Erskine, D.; Morris, C.M.; Majbour, N.K.; Vekrellis, K.; Li, J.-Y.; El-Agnaf, O.M.A. Heterogeneity in α-synuclein subtypes and their expression in cortical brain tissue lysates from Lewy body diseases and Alzheimer’s disease. Neuropathol. Appl. Neurobiol. 2019, 45, 597–608. [Google Scholar] [CrossRef] [PubMed]
- Beyer, K.; Lao, J.I.; Carrato, C.; Mate, J.L.; López, D.; Ferrer, I.; Ariza, A. Differential expression of α-synuclein isoforms in dementia with Lewy bodies. Neuropathol. Appl. Neurobiol. 2004, 30, 601–607. [Google Scholar] [CrossRef] [PubMed]
- Levitan, K.; Chereau, D.; Cohen, S.I.A.; Knowles, T.P.J.; Dobson, C.M.; Fink, A.L.; Anderson, J.P.; Goldstein, J.M.; Millhauser, G.L. Conserved C-Terminal Charge Exerts a Profound Influence on the Aggregation Rate of α-Synuclein. J. Mol. Biol. 2011, 411, 329–333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murray, I.V.J.; Giasson, B.I.; Quinn, S.M.; Koppaka, V.; Axelsen, P.H.; Ischiropoulos, H.; Trojanowski, J.Q.; Lee, V.M.-Y. Role of α-Synuclein Carboxy-Terminus on Fibril Formation in Vitro. Biochemistry 2003, 42, 8530–8540. [Google Scholar] [CrossRef] [PubMed]
- Nonaka, T.; Watanabe, S.T.; Iwatsubo, T.; Hasegawa, M. Seeded Aggregation and Toxicity of α-Synuclein and Tau: Cel-lular models of neurodegenerative diseases. J. Biol. Chem. 2010, 285, 34885–34898. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oueslati, A.; Ximerakis, M.; Vekrellis, K. Protein Transmission, Seeding and Degradation: Key Steps for α-Synuclein Prion-Like Propagation. Exp. Neurobiol. 2014, 23, 324–336. [Google Scholar] [CrossRef] [Green Version]
- Choi, M.L.; Gandhi, S. Crucial role of protein oligomerization in the pathogenesis of Alzheimer’s and Parkinson’s diseases. FEBS J. 2018, 285, 3631–3644. [Google Scholar] [CrossRef] [Green Version]
- Funahashi, Y.; Yoshino, Y.; Yamazaki, K.; Mori, Y.; Mori, T.; Ozaki, Y.; Sao, T.; Ochi, S.; Iga, J.-I.; Ueno, S.-I. DNA methylation changes atSNCAintron 1 in patients with dementia with Lewy bodies. Psychiatry Clin. Neurosci. 2017, 71, 28–35. [Google Scholar] [CrossRef] [Green Version]
- Locascio, J.J.; Eberly, S.; Liao, Z.; Liu, G.; Hoesing, A.N.; Duong, K.; Trisini-Lipsanopoulos, A.; Dhima, K.; Hung, A.Y.; Flaherty, A.W.; et al. Association between α-synuclein blood transcripts and early, neuroimaging-supported Parkinson’s disease. Brain 2015, 138, 2659–2671. [Google Scholar] [CrossRef]
- Pihlstrøm, L.; Berge, V.; Rengmark, A.; Toft, M. Parkinson’s disease correlates with promoter methylation in the α-synuclein gene. Mov. Disord. 2015, 30, 577–580. [Google Scholar] [CrossRef]
- Ai, S.-X.; Xu, Q.; Hu, Y.-C.; Song, C.-Y.; Guo, J.-F.; Shen, L.; Wang, C.-R.; Yu, R.-L.; Yan, X.-X.; Tang, B. Hypomethylation of SNCA in blood of patients with sporadic Parkinson’s disease. J. Neurol. Sci. 2014, 337, 123–128. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, L.; Takuma, H.; Tamaoka, A.; Kurisaki, H.; Date, H.; Tsuji, S.; Iwata, A. CpG Demethylation Enhances Alpha-Synuclein Expression and Affects the Pathogenesis of Parkinson’s Disease. PLoS ONE 2010, 5, e15522. [Google Scholar] [CrossRef] [Green Version]
- Elhadi, S.A.; Grigoletto, J.; Poli, M.; Arosio, P.; Arkadir, D.; Sharon, R. α-Synuclein in blood cells differentiates Parkinson’s disease from healthy controls. Ann. Clin. Transl. Neurol. 2019, 6, 2426–2436. [Google Scholar] [CrossRef] [PubMed]
- Cersosimo, M.G. Propagation of alpha-synuclein pathology from the olfactory bulb: Possible role in the pathogenesis of dementia with Lewy bodies. Cell Tissue Res. 2017, 373, 233–243. [Google Scholar] [CrossRef] [PubMed]
- Hughes, A.J.; Daniel, S.E.; Kilford, L.; Lees, A.J. Accuracy of clinical diagnosis of idiopathic Parkinson’s disease: A clinico-pathological study of 100 cases. J. Neurol. Neurosurg. Psychiatry 1992, 55, 181–184. [Google Scholar] [CrossRef] [Green Version]
- Barbour, R.; Kling, K.; Anderson, J.P.; Banducci, K.; Cole, T.; Diep, L.; Fox, M.; Goldstein, J.M.; Soriano, F.; Seubert, P.; et al. Red Blood Cells Are the Major Source of Alpha-Synuclein in Blood. Neurodegener. Dis. 2008, 5, 55–59. [Google Scholar] [CrossRef] [PubMed]
- Durrenberger, P.F.; Fernando, S.; Kashefi, S.N.; Ferrer, I.; Hauw, J.-J.; Seilhean, D.; Smith, C.; Walker, R.L.; Al-Sarraj, S.; Troakes, C.; et al. Effects of Antemortem and Postmortem Variables on Human Brain mRNA Quality: A BrainNet Europe Study. J. Neuropathol. Exp. Neurol. 2010, 69, 70–81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beyer, K.; Domingo-Sàbat, M.; Santos, C.; Tolosa, E.; Ferrer, I.; Ariza, A. The decrease of β-synuclein in cortical brain areas defines a molecular subgroup of dementia with Lewy bodies. Brain 2010, 133, 3724–3733. [Google Scholar] [CrossRef] [Green Version]
- Fuchs, J.; Tichopad, A.; Golub, Y.; Munz, M.; Schweitzer, K.J.; Wolf, B.; Berg, D.; Mueller, J.C.; Gasser, T. Genetic variability in the SNCA gene influences α-synuclein levels in the blood and brain. FASEB J. 2008, 22, 1327–1334. [Google Scholar] [CrossRef]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, research0034-1. [Google Scholar] [CrossRef] [Green Version]
- Marx, A.; Backes, C.; Meese, E.; Lenhof, H.-P.; Keller, A. EDISON-WMW: Exact Dynamic Programing Solution of the Wilcoxon–Mann–Whitney Test. Genom. Proteom. Bioinform. 2016, 14, 55–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| DLB (n = 16) | PD (n = 16) | CTRLs (n = 11) | p1 | |
|---|---|---|---|---|
| Mean age at death (age range) | 77.2 (60–90) | 79.9 (71–88) | 69.9 (59–81) | 0.004 |
| Gender (male/female ratio) | 1.3:1 | 1:1 | 1:1.2 | 0.329 |
| Disease onset 2 | 70.0 (56–83) | 59.9 (49–73) | - | <0.001 |
| Disease duration (range) | 6.1 (2–11) | 18.6 (3–29) | - | <0.001 |
| PM 3, hours (range) | 9:00 (3:15–21:00) | 8:10 (3:30–17:10) | 8:30 (2:15–23:30) | 0.571 |
| AD-stage 4 | 0-IV; A-C | 0-IV; A-C | ||
| PD-stage 5 | Braak 4–6 | |||
| AS pathol 6 neocortical (n, %) | 12 (75) | |||
| AS pathol limbic (n, %) | 4 (25) |
| DLB (n = 72) | PD (n = 59) | CTRLs (n = 54) | p1 | |
|---|---|---|---|---|
| Mean age (age range) | 76.0 (58–91) | 67.7 (45–84) | 71.6 (51–88) | 0.014 |
| Gender (male/female ratio) | 1.5:1 | 1.3:1 | 1:1.5 | 0.033 |
| Disease onset | 71.4 (57–86) | 62.3 (40–80) | - | <0.001 |
| Disease duration 2 (range) | 4.7 (0.9–8.7) | 5.4 (0.0–18.4) | - | 0.086 |
| MMSE 3, mean (range) | 15.1 (3–28) | - | - | - |
| UPDRS-III 4, mean (range) | - | 20.9 (5–39) | - | - |
| Parkinsonism, n (%) | 47 (79.7%) | - | - | - |
| Abnormal Dat-Spect imaging, n (%) | 55 (93.2%) | - | - | - |
| SNCAtv1 | SNCAtv2 | SNCAtv3 | SNCA112 | SNCA126 | |
|---|---|---|---|---|---|
| FC 1 | 65.5% 1 | 22.3% | 10.4% | 0.5% | 1.3% |
| 2 | 52.2% | 20.9% | 20.9% | 0.3% | 0.7% |
| Ca 3 | 63.4% | 25.3% | 9.5% | 0.6% | 1.2% |
| Pt 4 | 63.1% | 31.6% | 3.4% | 0.7% | 1.2% |
| Blood | 60.2% | 20.4% | 15.7% | 1.2% | 2.4% |
| SNCAtv1 | SNCAtv2 | SNCAtv3 | SNCA126 | |
|---|---|---|---|---|
| SNCAtv2 | 0.745 p < 0.0001 | |||
| SNCAtv3 | −0.266 p = 0.141 | −0.007 p = 0.934 | ||
| SNCA126 | 0.715 p < 0.0001 | 0.814 p < 0.0001 | −0.202 p = 0.276 | |
| SNCA112 | 0.814 p < 0.0001 | 0.834 p < 0.0001 | −0.262 p = 0.162 | 0.735 p < 0.0001 |
| Correlation between | DLB | PD |
|---|---|---|
| SNCAtv1 and SNCAtv2 | 0.81 (p < 0.0001) | 0.89 (p < 0.0001) |
| SNCAtv1 and SNCAtv3 | 0.44 (p = 0.0005) | 0.77 (p < 0.0001) |
| SNCAtv2 and SNCAtv3 | 0.46 (p < 0.0001) | 0.64 (p < 0.0001) |
| SNCAtv1 and SNCA126 | 0.75 (p < 0.0001) | 0.57 (p < 0.0001) |
| SNCAtv2 and SNCA126 | 0.81 (p < 0.0001) | 0.81 (p < 0.0001) |
| SNCAtv3 and SNCA126 | 0.81 (p < 0.0001) | 0.81 (p < 0.0001) |
| Primer Name | Primer Sequence (5′–3′) | Primer Comb 1 | Transcript 2 |
|---|---|---|---|
| SNCA-1U * | ATC CAG GAA CAG CTG TCT TC | 1U + 4L | SNCAtv3 |
| SNCA-2aU * | TTC AAG CCT TCT GCC TTT CC | 2aU + 4L | SNCAtv1 |
| SNCA-2bU * | AGT CGG AGT TGT GGA GAA GCA | 2bU + 4L | SNCAtv2 |
| SNCA-4L | ACC ACT GCT CCT CCA ACA T | ||
| SNCA-3/5U * | CTC TAT GTA GTG GCT GAG AA | 3/5U + 6L | SNCA126 |
| SNCA-4U | GTG CAT GGT GTG GCA ACA | ||
| SNCA-5/7L * | ATA CCC TTC CTT GCC CAA C | 3U + 5/7L | SNCA112 |
| SNCA-6L | GAG CAC TTG TAC AGG ATG G | ||
| NSE_6U | TGT ATC GCC ACA TTG CTC AG | ||
| NSE_7L1 | ACT GGG AGG ATC ATG AAC TC | 6U + 7L1 3 | |
| NSE_7L2 | ATG GCA TCC CGA AAG CTC TCA | 6U + 7L2 3 | |
| SYPex2U | GCT TTG TGA AGG TGC TGC AAT | 2U + 4L1 3 | |
| SYPex3U | TCT TCG CCA TCT TCG CCT TTG | 3U + 4L1 3 | |
| SYPex4L1 | TGC ATC AAA GTA CAC TTG GTG CA | ||
| PBGD_U1 | ACA CAC AGC CTA CTT TCC AAG | ||
| PBGD_L1 | TCA ATG TTG CCA CCA CAC TGT | ||
| beta-actin U2 | TCT ACA ATG AGC TGC GTG TG | ||
| beta-actin L3 | TAG ATG GGC ACA GTG TGG GT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marsal-García, L.; Urbizu, A.; Arnaldo, L.; Campdelacreu, J.; Vilas, D.; Ispierto, L.; Gascón-Bayarri, J.; Reñé, R.; Álvarez, R.; Beyer, K. Expression Levels of an Alpha-Synuclein Transcript in Blood May Distinguish between Early Dementia with Lewy Bodies and Parkinson’s Disease. Int. J. Mol. Sci. 2021, 22, 725. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22020725
Marsal-García L, Urbizu A, Arnaldo L, Campdelacreu J, Vilas D, Ispierto L, Gascón-Bayarri J, Reñé R, Álvarez R, Beyer K. Expression Levels of an Alpha-Synuclein Transcript in Blood May Distinguish between Early Dementia with Lewy Bodies and Parkinson’s Disease. International Journal of Molecular Sciences. 2021; 22(2):725. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22020725
Chicago/Turabian StyleMarsal-García, Laura, Aintzane Urbizu, Laura Arnaldo, Jaume Campdelacreu, Dolores Vilas, Lourdes Ispierto, Jordi Gascón-Bayarri, Ramón Reñé, Ramiro Álvarez, and Katrin Beyer. 2021. "Expression Levels of an Alpha-Synuclein Transcript in Blood May Distinguish between Early Dementia with Lewy Bodies and Parkinson’s Disease" International Journal of Molecular Sciences 22, no. 2: 725. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22020725