Effects of 6,8-Diprenylgenistein on VEGF-A-Induced Lymphangiogenesis and Lymph Node Metastasis in an Oral Cancer Sentinel Lymph Node Animal Model




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
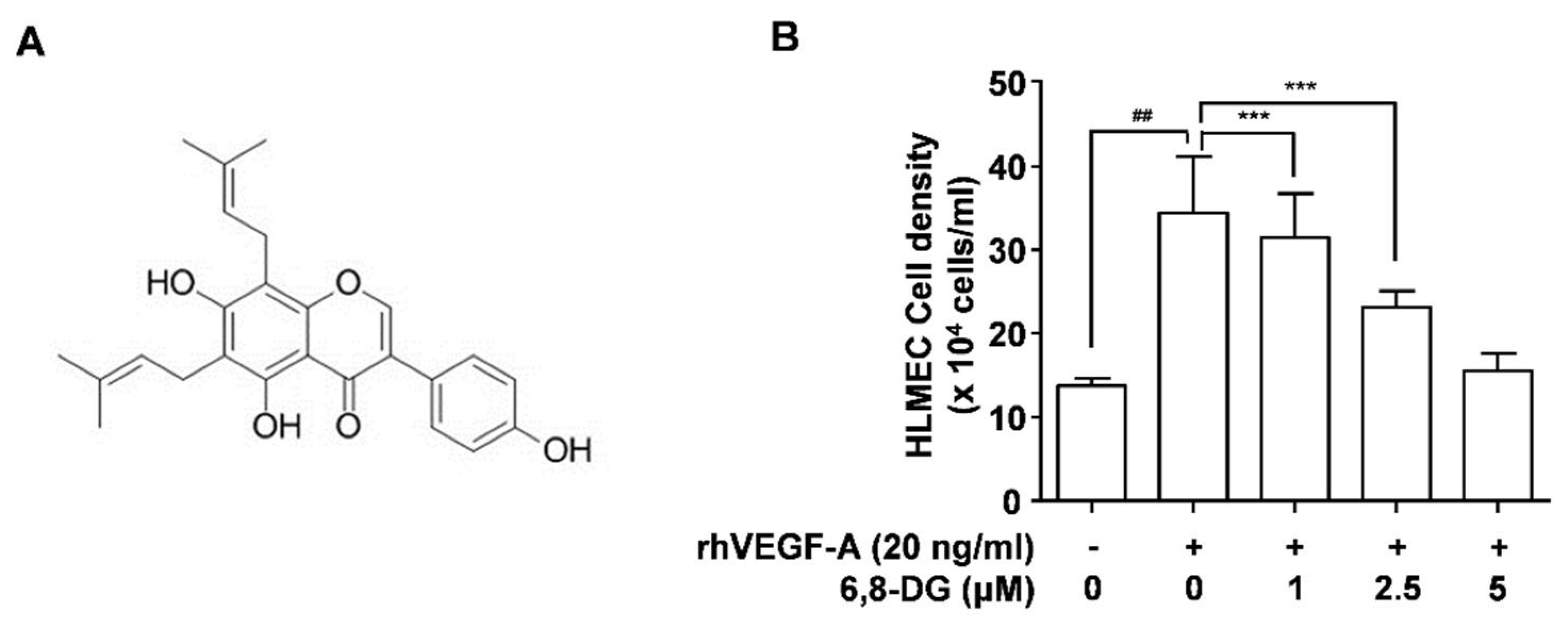
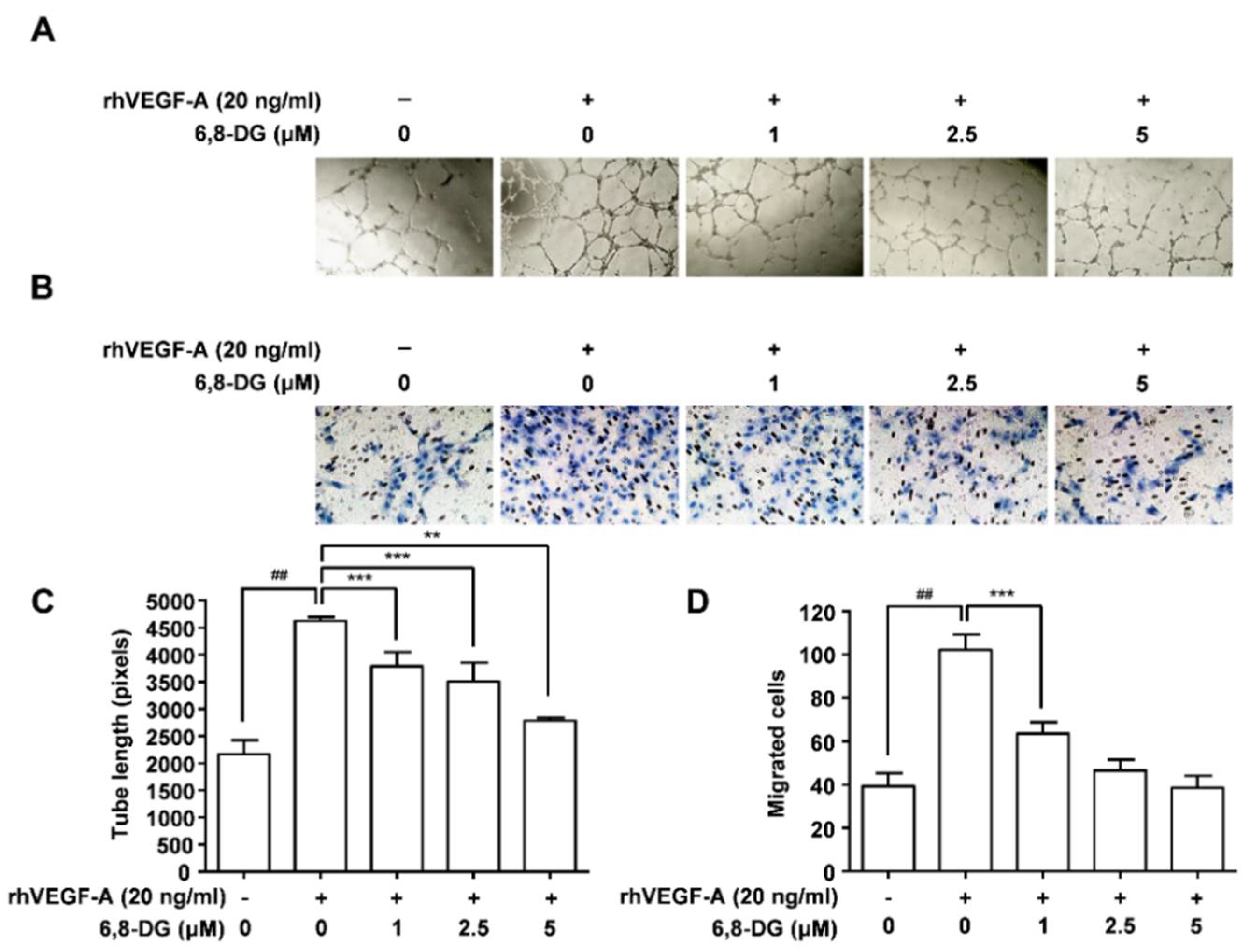
2.1. Effect of 6,8-Diprenylgenestein on VEGF-A-Induced Proliferation, Tube Formation, and Migration of HLMECs
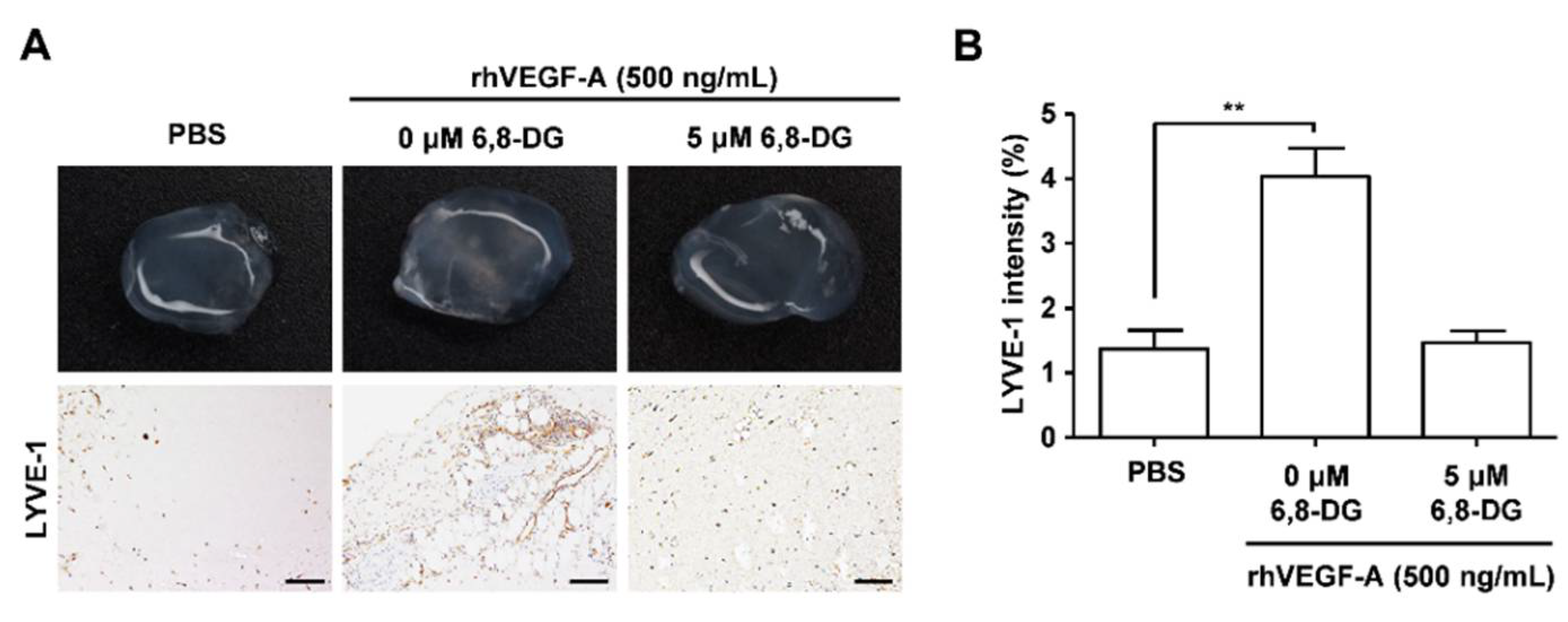
2.2. Effects of 6,8-Diprenylgenistein on rhVEGF-A-Induced Lymphangiogenesis In Vivo in a Matrigel Plug
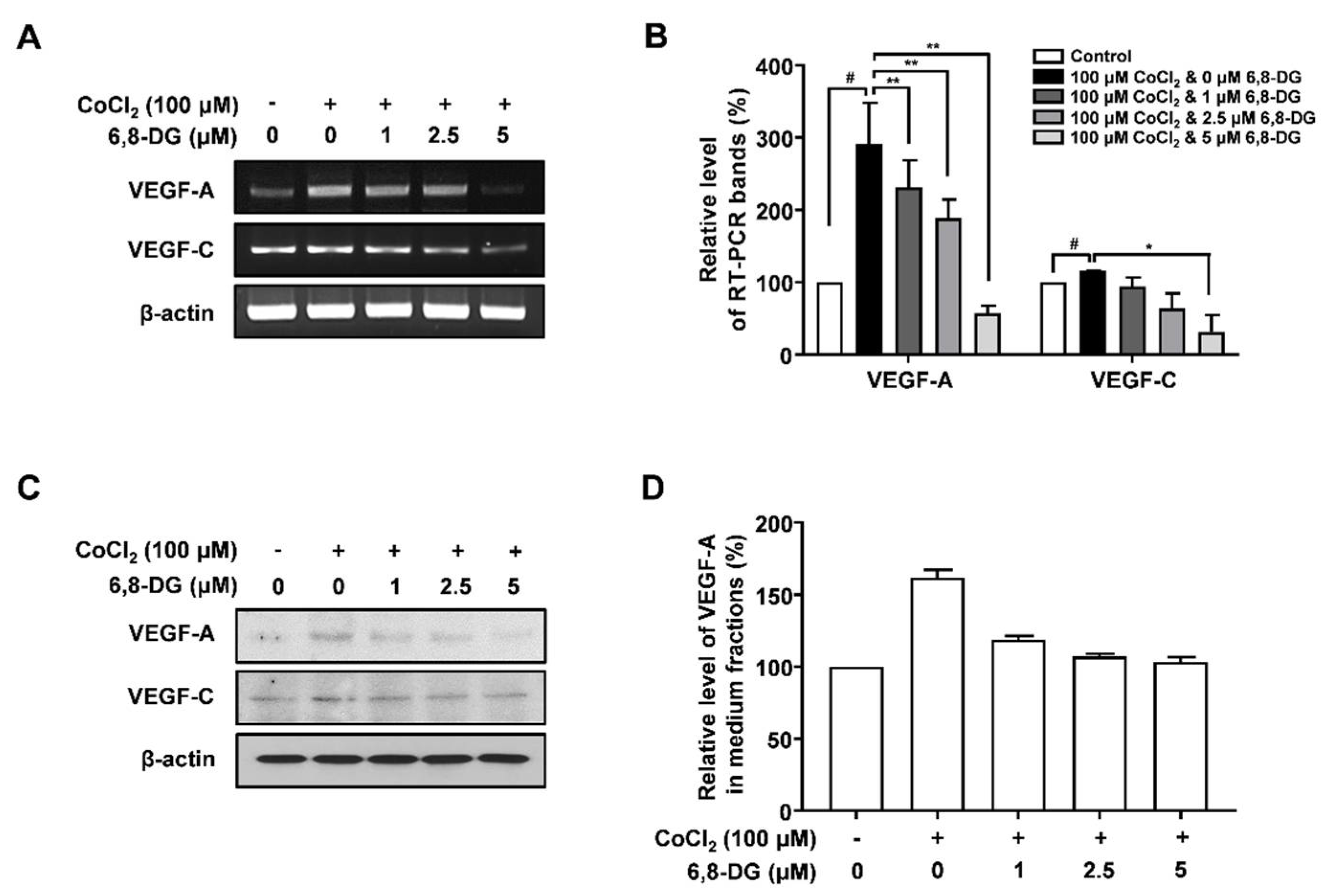
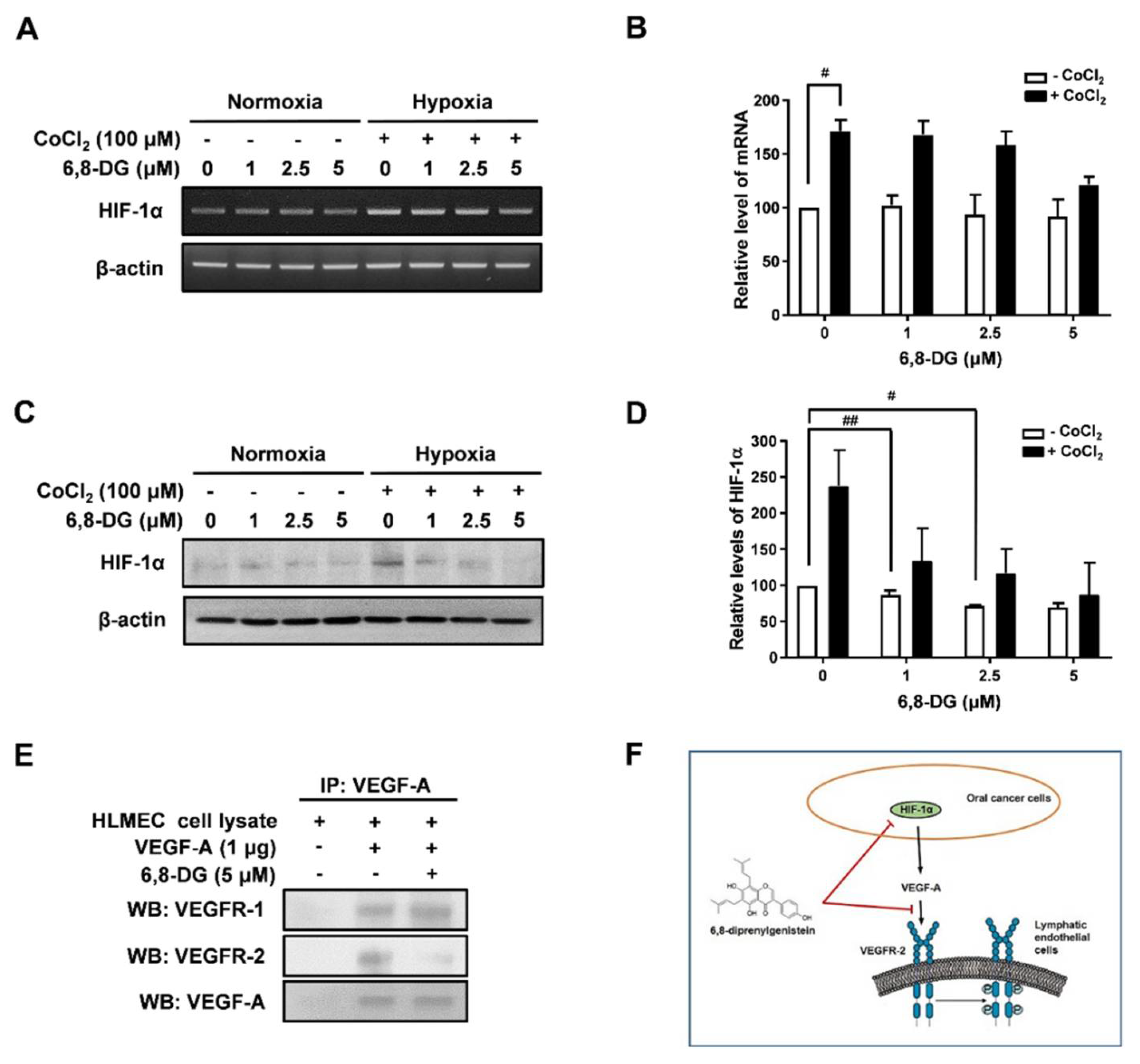
2.3. Effects of 6,8-Diprenylgenistein on Expression of Lymphangiogenic Factors in CoCl2-Treated SCCVII Cells
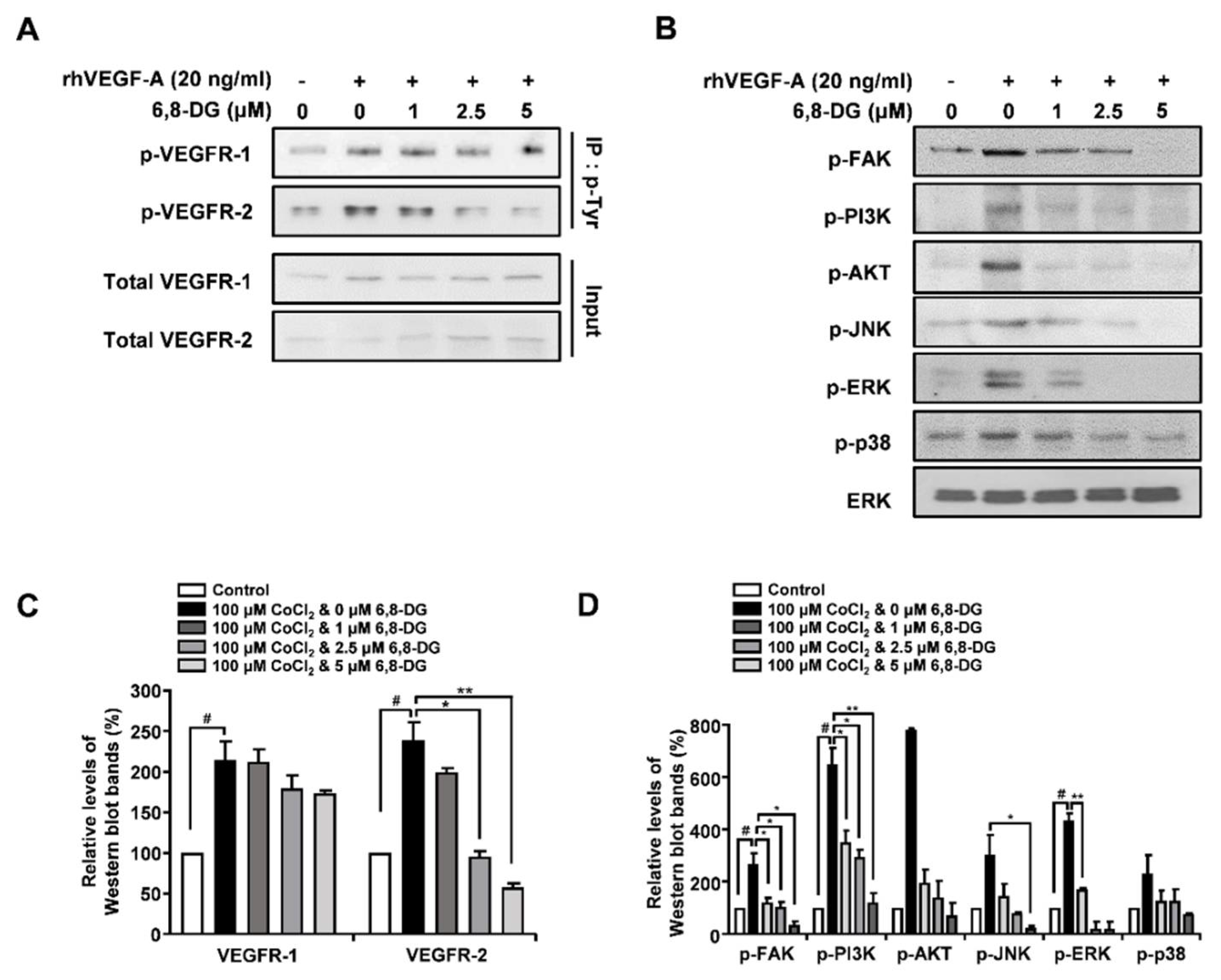
2.4. Effects of 6,8-Diprenylgenistein on Activation of VEGF Receptor-2, and Lymphangiogenesis-Related Downstream Signaling Factors in rhVEGF-A-Treated HLMECs
2.5. Effects of 6,8-Diprenylgenistein on Oral Cancer Related Lymphangiogenesis
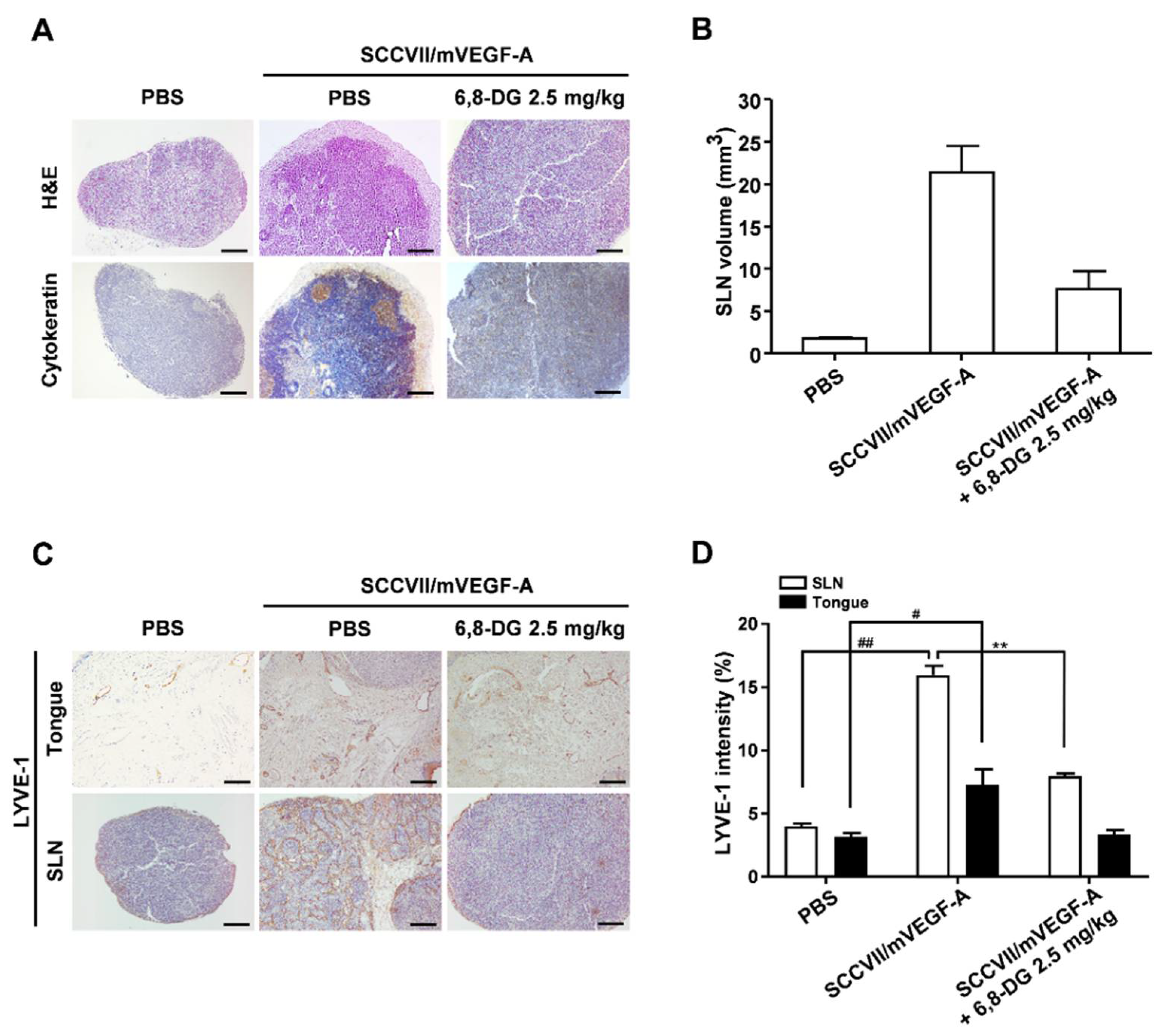
2.6. Effect of 6,8-Diprenylgenistein on Lymphangiogenesis and Lymph Node Metastasis in a VEGF-A-Induced OCSLN Animal Model
3. Discussion
4. Materials and Methods
4.1. Cell Lines and Compound
4.2. Proliferation, Migration, and Tube Formation Assay
4.3. In Vivo Matrigel Plug Assay
4.4. Reverse Transcription-Polymerase Chain Reaction (RT-PCR) Analysis
4.5. Western Blot and Immunoprecipitation Analysis
4.6. ELISA Assay
4.7. Co-Immunoprecipitation Assay
4.8. Animals and Study Design
4.9. Immunohistochemical Analysis
4.10. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, W.M.; Zhao, Z.L.; Ma, S.R.; Yu, G.T.; Liu, B.; Zhang, L.; Zhang, W.F.; Kulkarni, A.B.; Sun, Z.J.; Zhao, Y.F. Epidermal growth factor receptor inhibition reduces angiogenesis via hypoxia-inducible factor-1α and notch1 in head neck squamous cell carcinoma. PLoS ONE 2015, 10, e0119723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ozasa, R.; Ohno, J.; Iwahashi, T.; Taniguchi, K. Tumor-induced lymphangiogenesis in cervical lymph nodes in oral melanoma-bearing mice. J. Exp. Clin. Cancer Res. 2012, 31, 83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wakisaka, N.; Hasegawa, Y.; Yoshimoto, S.; Miura, K.; Shiotani, A.; Yokoyama, J.; Sugasawa, M.; Moriyama-Kita, M.; Endo, K.; Yoshizaki, T. Primary tumor-secreted lymphangiogenic factors induce pre-metastatic lymphvascular niche formation at sentinel lymph nodes in oral squamous cell carcinoma. PLoS ONE 2015, 10, e0144056. [Google Scholar] [CrossRef] [PubMed]
- Duong, T.; Koopman, P.; Francois, M. Tumor lymphangiogenesis as a potential therapeutic target. J. Oncol. 2012, 2012, 204946. [Google Scholar] [CrossRef] [Green Version]
- Hwang-Bo, J.; Park, J.H.; Bae, M.G.; Chung, I.S. Recombinant canstatin inhibits VEGF-A-induced lymphangiogenesis and metastasis in an oral squamous cell carcinoma SCC-VII animal model. Cancer Med. 2016, 5, 2977–2988. [Google Scholar] [CrossRef]
- Christiansen, A.; Detmar, M. Lymphangiogenesis and cancer. Genes Cancer 2011, 2, 1146–1158. [Google Scholar] [CrossRef] [Green Version]
- Hwang-Bo, J.; Bae, M.G.; Park, J.H.; Chung, I.S. 3-O-Acetyloleanolic acid inhibits VEGF-A-induced lymphangiogenesis and lymph node metastasis in an oral cancer sentinel lymph node animal model. BMC Cancer 2018, 18, 714. [Google Scholar] [CrossRef]
- Miyahara, M.; Tanuma, J.; Sugihara, K.; Semba, I. Tumor lymphangiogenesis correlates with lymph node metastasis and clinicopathologic parameters in oral squamous cell carcinoma. Cancer 2007, 110, 1287–1294. [Google Scholar] [CrossRef]
- Dadras, S.S.; Paul, T.; Bertoncini, J.; Brown, L.F.; Muzikansky, A.; Jackson, D.G.; Ellwanger, U.; Garbe, C.; Mihm, M.C.; Detmar, M. Tumor lymphangiogenesis: A novel prognostic indicator for cutaneous melanoma metastasis and survival. Am. J. Pathol. 2003, 162, 1951–1960. [Google Scholar] [CrossRef]
- Renyi-Vamos, F.; Tovari, J.; Fillinger, J.; Timar, J.; Paku, S.; Kenessey, I.; Ostoros, G.; Agocs, L.; Soltesz, I.; Dome, B. Lymphangiogenesis correlates with lymph node metastasis, prognosis, and angiogenic phenotype in human non-small cell lung cancer. Clin. Cancer Res. 2005, 11, 7344–7353. [Google Scholar] [CrossRef] [Green Version]
- Mumprecht, V.; Detmar, M. Lymphangiogenesis and cancer metastasis. J. Cell Mol. Med. 2009, 13, 1405–1416. [Google Scholar] [CrossRef] [PubMed]
- Hartiala, P.; Saarikko, A. Lymphangiogenesis and lymphangiogenic growth factors. J. Reconstr. Microsurg. 2016, 32, 10–15. [Google Scholar] [PubMed] [Green Version]
- Ji, R.C. Lymph node lymphangiogenesis: A new concept for modulating tumor metastasis and inflammatory process. Histol. Histopathol. 2009, 24, 377–384. [Google Scholar]
- Hwang-Bo, J.; Yoo, K.H.; Park, J.H.; Jeong, H.S.; Chung, I.S. Recombinant canstatin inhibits angiopoietin-1-induced angiogenesis and lymphangiogenesis. Int. J. Cancer 2012, 131, 298–309. [Google Scholar] [CrossRef] [PubMed]
- Hirakawa, S.; Kodama, S.; Kunstfeld, R.; Kajiya, K.; Brown, L.F.; Detmar, M. VEGF-A induces tumor and sentinel lymph node lymphangiogenesis and promotes lymphatic metastasis. J. Exp. Med. 2005, 201, 1089–1099. [Google Scholar] [CrossRef] [Green Version]
- Shin, J.W.; Huggenberger, R.; Detmar, M. Transcriptional profiling of VEGF-A and VEGF-C target genes in lymphatic endothelium reveals endothelial-specific molecule-1 as a novel mediator of lymphangiogenesis. Blood 2008, 112, 2318–2326. [Google Scholar] [CrossRef] [Green Version]
- Ferrara, N.; Gerber, H.P.; LeCouter, J. The biology of VEGF and its receptors. Nat. Med. 2003, 9, 669–676. [Google Scholar] [CrossRef]
- Melincovici, C.S.; Boşca, A.B.; Şuşman, S.; Mărginean, M.; Mihu, C.; Istrate, M.; Moldovan, I.M.; Roman, A.L.; Mihu, C.M. Vascular endothelial growth factor (VEGF)—Key factor in normal and pathological angiogenesis. Rom. J. Morphol. Embryol. 2018, 59, 455–467. [Google Scholar]
- Spagnuolo, C.; Russo, G.L.; Orhan, I.E.; Habtemariam, S.; Daglia, M.; Sureda, A.; Nabavi, S.F.; Devi, K.P.; Loizzo, M.R.; Tundis, R.; et al. Genistein and cancer: Current status, challenges, and future directions. Adv. Nutr. 2015, 6, 408–419. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.R.; Gugger, E.T.; Tanaka, T.; Guo, Y.; Blackburn, G.L.; Clinton, S.K. Soybean phytochemicals inhibit the growth of transplantable human prostate carcinoma and tumor angiogenesis in mice. J. Nutr. 1999, 129, 1628–1635. [Google Scholar] [CrossRef] [Green Version]
- Fassina, G.; Venè, R.; Morini, M.; Minghelli, S.; Benelli, R.; Noonan, D.M.; Albini, A. Mechanisms of inhibition of tumor angiogenesis and vascular tumor growth by epigallocatechin-3-gallate. Clin. Cancer Res. 2004, 10, 4865–4873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, F.Y.; Chiang, E.P.; Shih, C.J. Green tea catechin inhibits ephrin-A1-mediated cell migration and angiogenesis of human umbilical vein endothelial cells. J. Nutr. Biochem. 2007, 18, 391–399. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.S.; Kim, I.A.; Ko, Y.J.; Jeong, J.A.; Kim, J.E.; Song, B.J.; Lim, W.B.; Lim, H.S.; Park, J.I.; Kim, S.Y.; et al. Methanol extract of leaves from Cudrania tricuspidata effects in HT-29 colorectal adenocarcinoma. Kor. J. Oral Maxillofac. Pathol. 2009, 33, 19–26. [Google Scholar]
- Kwon, S.B.; Kim, M.J.; Yang, J.M.; Lee, H.P.; Hong, J.T.; Jeong, H.S.; Kim, E.S.; Yoon, D.Y. Cudrania tricuspidata stem extract induces apoptosis via the extrinsic pathway in SiHa cervical cancer cells. PLoS ONE 2016, 11, e0150235. [Google Scholar] [CrossRef] [PubMed]
- Cho, E.J.; Yokozawa, T.; Rhyu, D.Y.; Kim, H.Y.; Shibahara, N.; Park, J.C. The inhibitory effects of 12 medicinal plants and their component compounds on lipid peroxidation. Am. J. Chin. Med. 2003, 31, 907–917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jo, Y.H.; Choi, K.M.; Liu, Q.; Kim, S.B.; Ji, H.J.; Kim, M.; Shin, S.K.; Do, S.G.; Shin, E.; Jung, G.; et al. Anti-obesity effect of 6,8-diprenylgenistein, an isoflavonoid of Cudrania tricuspidata fruits in high-fat diet-induced obese mice. Nutrients 2015, 7, 10480–10490. [Google Scholar] [CrossRef]
- Ahn, S.J.; Park, S.N.; Lee, Y.J.; Cho, E.J.; Lim, Y.K.; Li, X.M.; Choi, M.H.; Seo, Y.W.; Kook, J.K. In vitro antimicrobial activities of 1-methoxyficifolinol, licorisoflavan A, and 6,8-diprenylgenistein against Streptococcus mutans. Caries Res. 2015, 49, 78–89. [Google Scholar] [CrossRef]
- Vadlapatla, R.K.; Vadlapudi, A.D.; Mitra, A.K. Hypoxia-inducible factor-1 (HIF-1): A potential target for intervention in ocular neovascular diseases. Curr. Drug Targets 2013, 14, 919–935. [Google Scholar] [CrossRef]
- Roh, J.L.; Cho, K.J.; Kwon, G.Y.; Ryu, C.H.; Chang, H.W.; Choi, S.H.; Nam, S.Y.; Kim, S.Y. The prognostic value of hypoxia markers in T2-staged oral tongue cancer. Oral Oncol. 2009, 45, 63–68. [Google Scholar] [CrossRef]
- Borenstein, X.; Fiszman, G.L.; Blidner, A.; Vanzulli, S.I.; Jasnis, M.A. Functional changes in murine mammary cancer cells elicited by CoCl2-induced hypoxia. Nitric Oxide 2010, 23, 234–241. [Google Scholar] [CrossRef]
- Tsuzuki, T.; Okada, H.; Cho, H.; Tsuji, S.; Nishigaki, A.; Yasuda, K.; Kanzaki, H. Hypoxic stress simultaneously stimulates vascular endothelial growth factor via hypoxia-inducible factor-1 and inhibits stromal cell-derived factor-1 in human endometrial stromal cells. Hum. Reprod. 2012, 27, 523–530. [Google Scholar] [CrossRef] [PubMed]
- Huusko, J.; Merentie, M.; Dijkstr, M.H.; Ryhänen, M.M.; Karvinen, H.; Rissanen, T.T.; Vanwildemeersch, M.; Hedman, M.; Lipponen, J.; Heinonen, S.E.; et al. The effects of VEGF-R1 and VEGF-R2 ligands on angiogenic responses and left ventricular function in mice. Cardiovasc. Res. 2010, 86, 122–130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ji, R.C. Hypoxia and lymphangiogenesis in tumor microenvironment and metastasis. Cancer Lett. 2014, 346, 6–16. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Wang, N.; Han, S.; Wang, D.; Mo, S.; Yu, L.; Huang, H.; Tsui, K.; Shen, J.; Chen, J. Dietary compound isoliquiritigenin inhibits breast cancer neoangiogenesis via VEGF/VEGFR-2 signaling pathway. PLoS ONE 2013, 8, e68566. [Google Scholar] [CrossRef] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bae, M.G.; Hwang-Bo, J.; Lee, D.Y.; Lee, Y.-H.; Chung, I.S. Effects of 6,8-Diprenylgenistein on VEGF-A-Induced Lymphangiogenesis and Lymph Node Metastasis in an Oral Cancer Sentinel Lymph Node Animal Model. Int. J. Mol. Sci. 2021, 22, 770. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22020770
Bae MG, Hwang-Bo J, Lee DY, Lee Y-H, Chung IS. Effects of 6,8-Diprenylgenistein on VEGF-A-Induced Lymphangiogenesis and Lymph Node Metastasis in an Oral Cancer Sentinel Lymph Node Animal Model. International Journal of Molecular Sciences. 2021; 22(2):770. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22020770
Chicago/Turabian StyleBae, Mun Gyeong, Jeon Hwang-Bo, Dae Young Lee, Youn-Hyung Lee, and In Sik Chung. 2021. "Effects of 6,8-Diprenylgenistein on VEGF-A-Induced Lymphangiogenesis and Lymph Node Metastasis in an Oral Cancer Sentinel Lymph Node Animal Model" International Journal of Molecular Sciences 22, no. 2: 770. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22020770