Comparison of the Translational Potential of Human Mesenchymal Progenitor Cells from Different Bone Entities for Autologous 3D Bioprinted Bone Grafts

 ,
,  , and
, and
Abstract
:1. Introduction
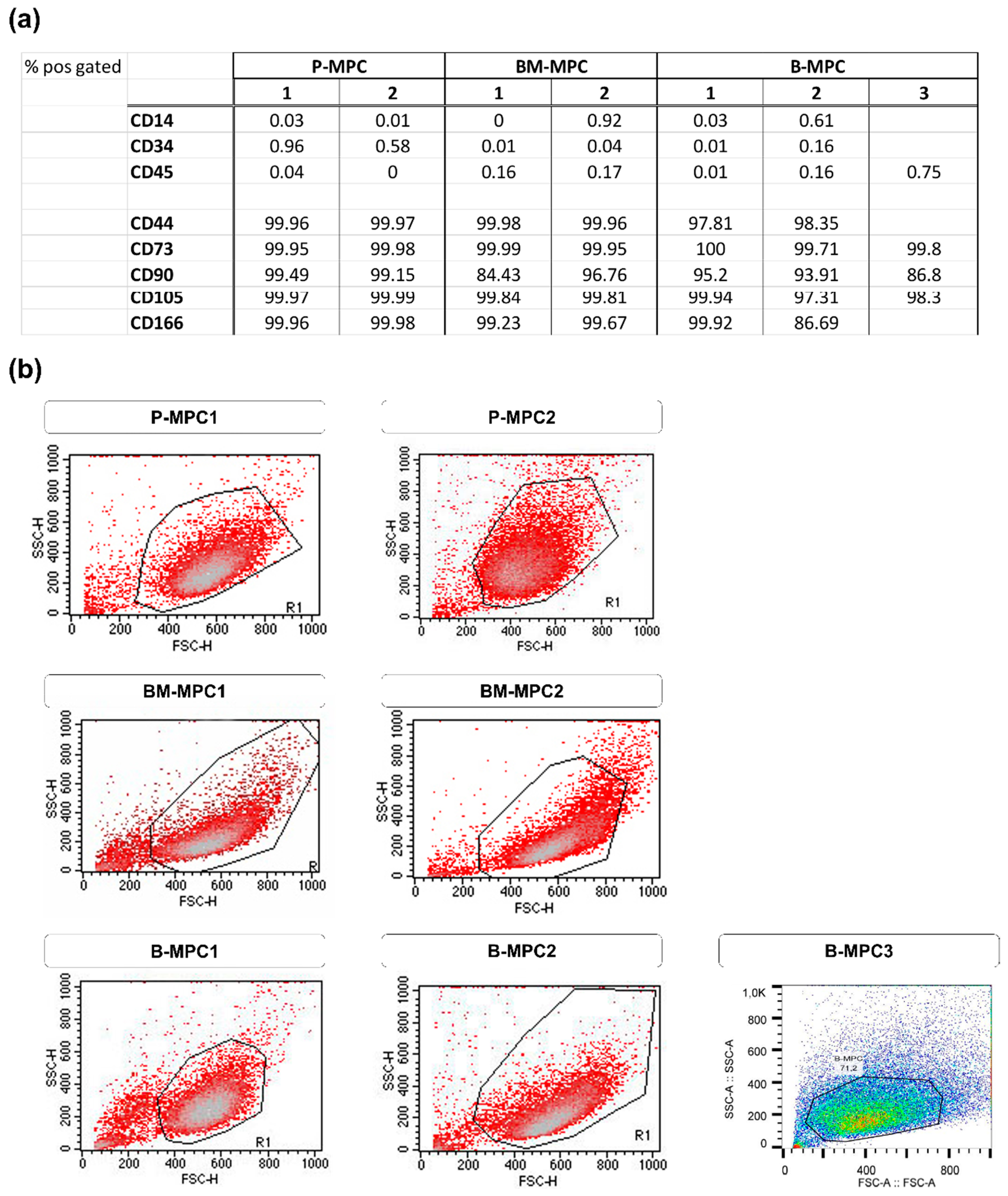
2. Results
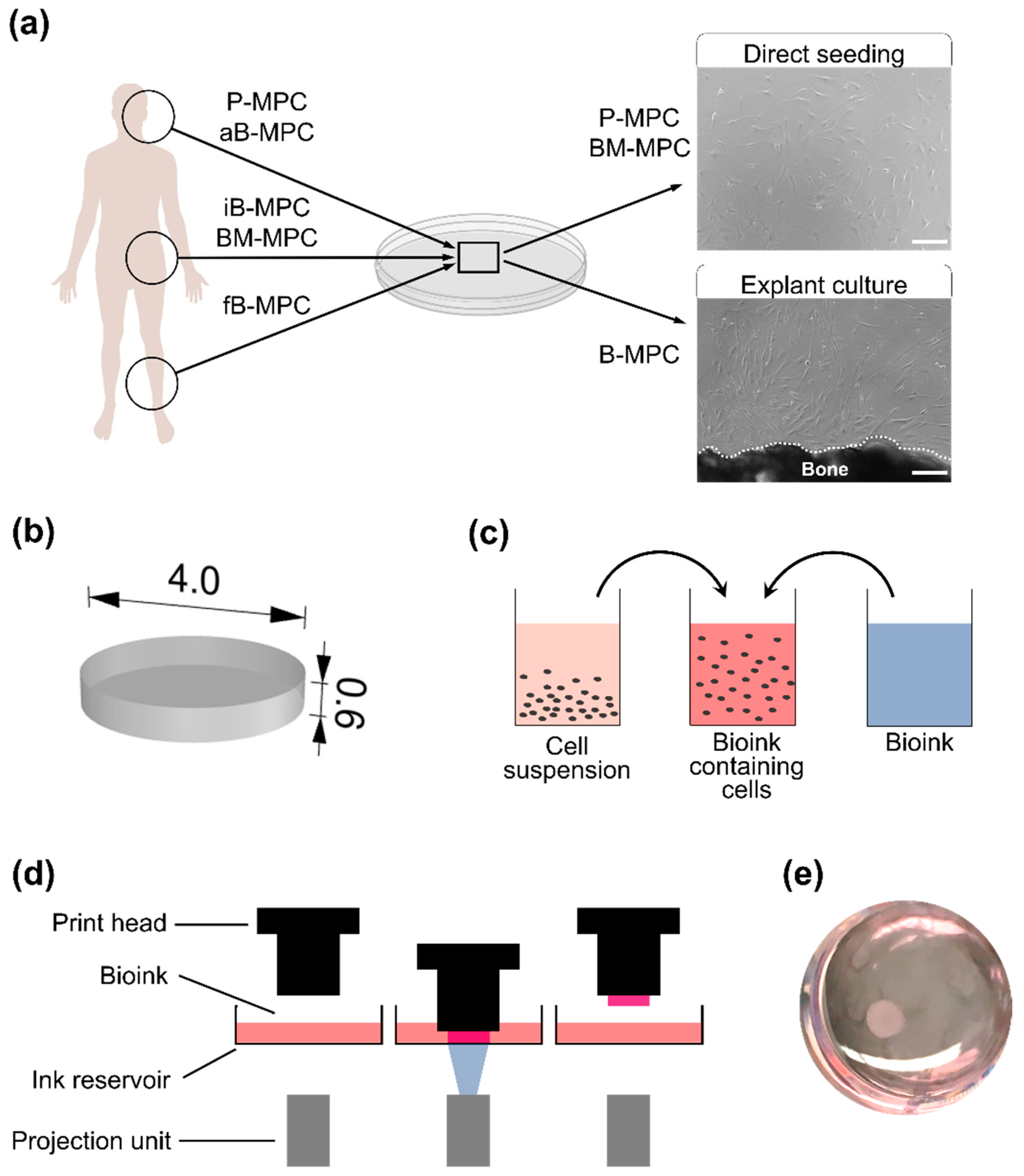
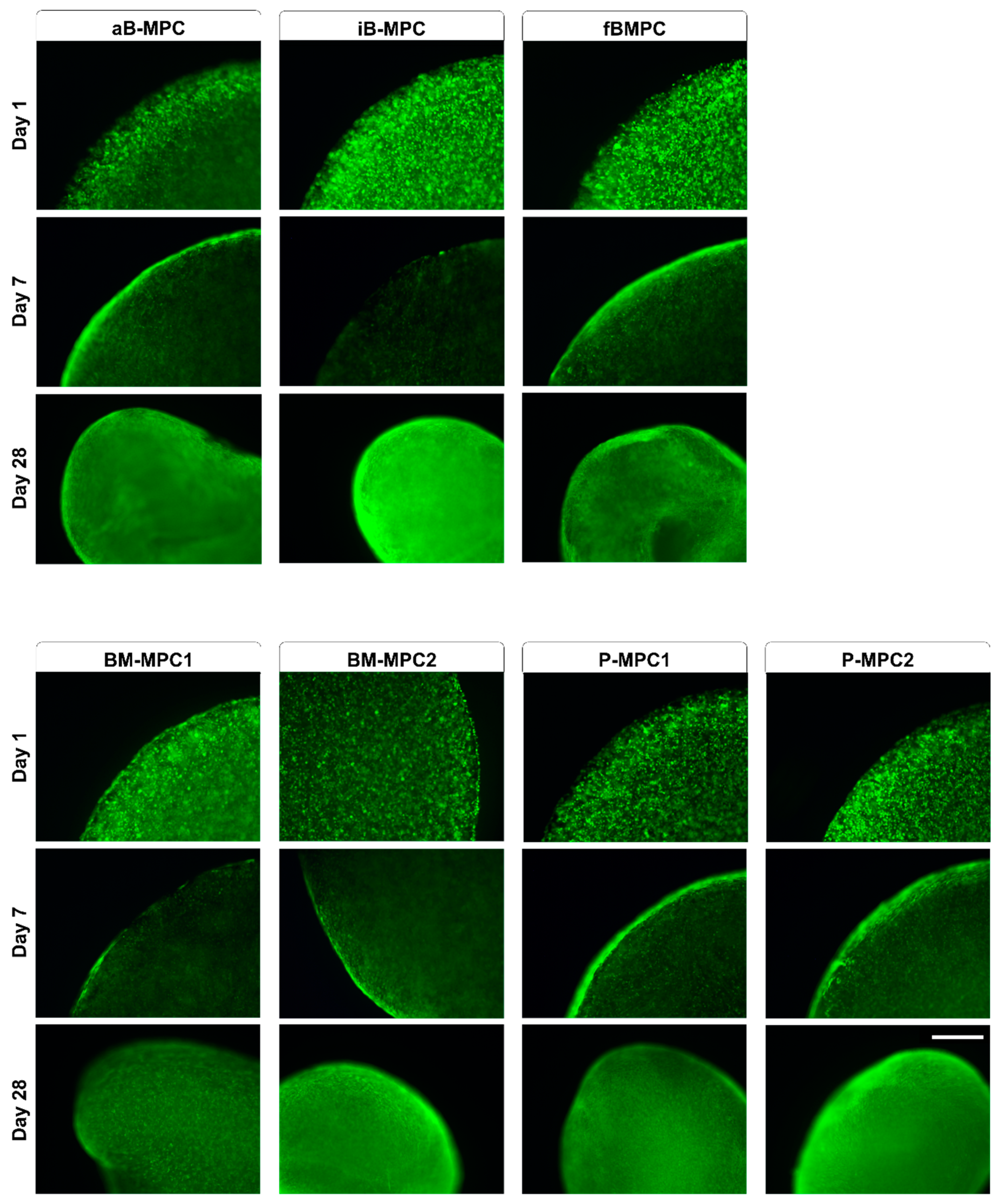
2.1. Bioprinting and Cultivation of Printed Constructs
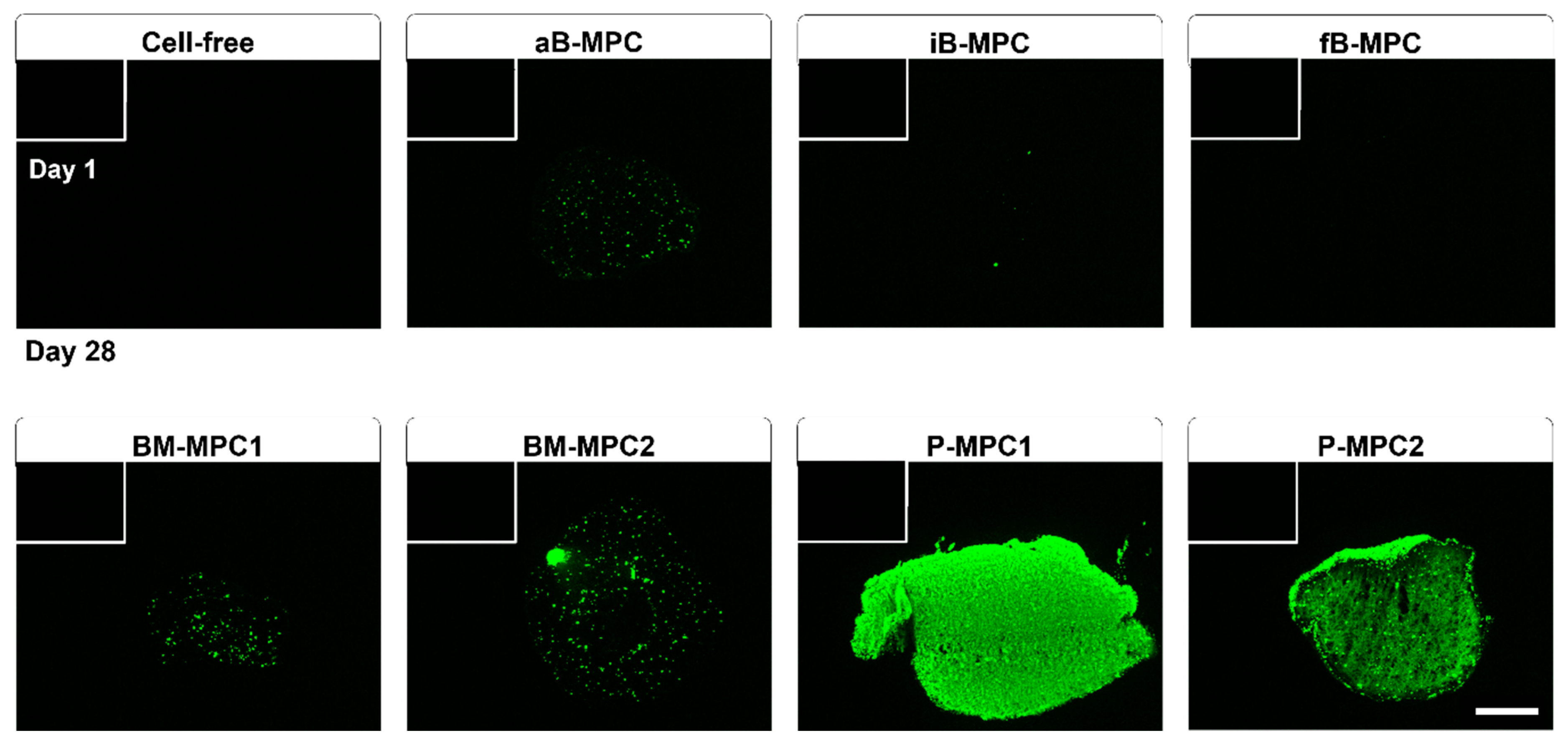
2.2. Viability and Metabolic Activity
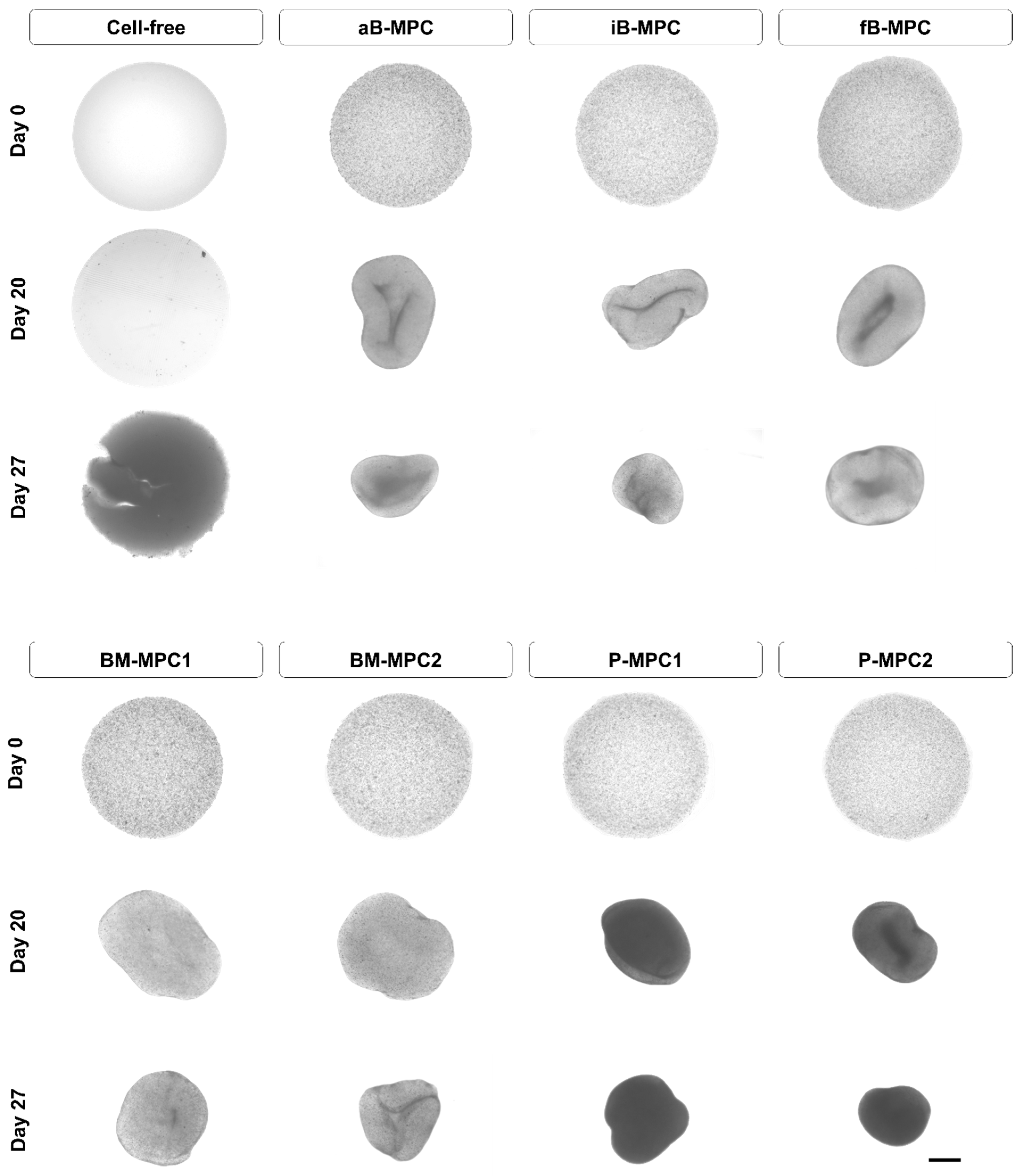
2.3. Mineralization
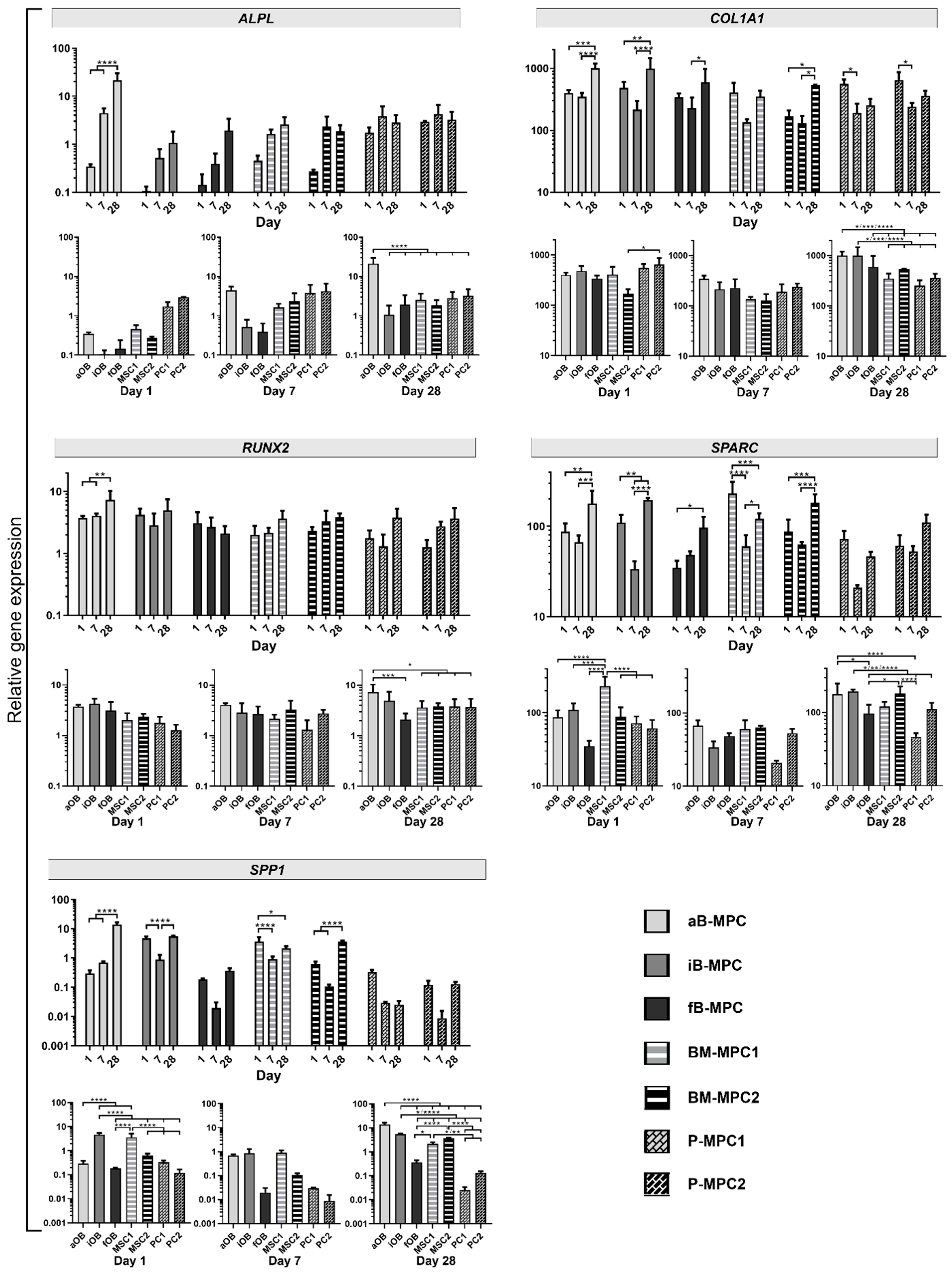
2.4. Gene Expression Analysis
3. Discussion
4. Materials and Methods
4.1. Photoink Synthesis
4.2. Cell Isolation and Culture
4.2.1. Ethical Statement
4.2.2. Bone Marrow MPCs
4.2.3. Bone-Derived MPCs
4.2.4. Periosteum-Derived MPCs
4.3. Bioprinting
4.4. Bioprint Cultivation
4.5. Assessment of Viability
4.5.1. alamarBlueTM Assay
4.5.2. Live/Dead Staining
4.6. Mineralization
4.7. Real-Time PCR
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| 3D | Three-dimensional |
| 1H-NMR | Proton nuclear magnetic resonance |
| a | Alveolar bone |
| AB | alamarBlueTM |
| B-MPC | Bone-derived mesenchymal progenitor cell |
| BM-MPC | Bone-marrow-derived mesenchymal progenitor cell |
| CAD | Computer-aided design |
| CO2 | Carbon dioxide |
| DMEM | Dulbecco’s modified Eagle’s medium |
| ECM | Extracellular matrix |
| EDTA | Ethylenediaminetetraacetic acid |
| f | Fibular bone |
| FCS | Fetal calf serum |
| FGF | Fibroblast growth factor |
| GelMA | Methacrylated gelatin |
| HEPES | Hydroxyethyl piperazineethanesulfonic acid |
| i | Iliac-crest bone |
| kDa | Kilodalton |
| LAP | Lithium phenyl-2,4,6-trimethylbenzoylphosphinate |
| min | Minute |
| mL | Milliliter |
| MHz | Megahertz |
| MPC | Mesenchymal progenitor cell |
| mRNA | Messenger ribonucleic acid |
| µL | Microliter |
| nm | Nanometer |
| PBS | Phosphate-buffered saline |
| PI/FDA | Propidium iodide/fluorescein diacetate |
| P-MPC | Periosteum-derived mesenchymal progenitor cell |
| RGD | Tripeptide Arg-Gly-Asp |
| STL | Stereolithography |
Appendix A



References
- Reichert, J.C.; Saifzadeh, S.; Wullschleger, M.E.; Epari, D.R.; Schütz, M.A.; Duda, G.N.; Schell, H.; Van Griensven, M.; Redl, H.; Hutmacher, D.W. The challenge of establishing preclinical models for segmental bone defect research. Biomater 2009, 30, 2149–2163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rendenbach, C.; Hölterhoff, N.; Hischke, S.; Kreutzer, K.; Smeets, R.; Assaf, A.T.; Heiland, M.; Wikner, J. Free flap surgery in Europe: An interdisciplinary survey. Int. J. Oral Maxillofac. Surg. 2018, 47, 676–682. [Google Scholar] [CrossRef] [PubMed]
- Homma, Y.; Zimmermann, G.; Hernigou, P. Cellular therapies for the treatment of non-union: The past, present and future. Injury 2013, 44, S46–S49. [Google Scholar] [CrossRef]
- McKay, W.F.; Peckham, S.M.; Badura, J.M. A comprehensive clinical review of recombinant human bone morphogenetic protein-2 (INFUSE® Bone Graft). Int. Orthop. 2007, 31, 729–734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beck-Broichsitter, B.E.; Garling, A.; Koehne, T.; Barvencik, F.; Smeets, R.; Mehl, C.; Jeschke, A.; Wiltfang, J.; Becker, S.T. 3D-tracking the regenerative potential of the mandible with micro-CTs. Oral Maxillofac. Surg. 2015, 19, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Rendenbach, C.; Steffen, C.; Hanken, H.; Schluermann, K.; Henningsen, A.; Beck-Broichsitter, B.; Kreutzer, K.; Heiland, M.; Precht, C. Complication rates and clinical outcomes of osseous free flaps: A retrospective comparison of CAD/CAM versus conventional fixation in 128 patients. Int. J. Oral Maxillofac. Surg. 2019, 48, 1156–1162. [Google Scholar] [CrossRef] [PubMed]
- Rendenbach, C.; Rashad, A.; Hansen, L.; Kohlmeier, C.; Dyck, M.L.; Suling, A.; Assaf, A.T.; Amling, M.; Heiland, M.; Wikner, J.; et al. Functional donor site morbidity longer than one year after fibula free flap: A prospective biomechanical analysis. Microsurgery 2018, 38, 395–401. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, T.; Khademhosseini, A.; Langer, R. Chasing the Paradigm: Clinical Translation of 25 Years of Tissue Engineering. Tissue Eng. Part A 2019, 25, 679–687. [Google Scholar] [CrossRef] [PubMed]
- Baltazar, T.; Merola, J.; Catarino, C.; Xie, C.B.; Kirkiles-Smith, N.C.; Lee, V.; Hotta, S.; Dai, G.; Xu, X.; Ferreira, F.C.; et al. Three Dimensional Bioprinting of a Vascularized and Perfusable Skin Graft Using Human Keratinocytes, Fibroblasts, Pericytes, and Endothelial Cells. Tissue Eng. Part A 2020, 26, 227–238. [Google Scholar] [CrossRef] [PubMed]
- Lam, T.; Ruppelt, A.; Thomas, A.; Amler, A.-K.; Noichl, B.P.; Lauster, R.; Kloke, L. Bioprinting Perfusion-Enabled Liver Equivalents for Advanced Organ-on-a-Chip Applications. Genes 2018, 9, 176. [Google Scholar] [CrossRef] [Green Version]
- Rathan, S.; Dejob, L.; Schipani, R.; Haffner, B.; Möbius, M.E.; Kelly, D.J. Fiber Reinforced Cartilage ECM Functionalized Bioinks for Functional Cartilage Tissue Engineering. Adv. Healthc. Mater. 2019, 8, e1801501. [Google Scholar] [CrossRef] [PubMed]
- Lam, T.; Dehne, T.; Krüger, J.P.; Hondke, S.; Endres, M.; Thomas, A.; Lauster, R.; Sittinger, M.; Kloke, L. Photopolymerizable gelatin and hyaluronic acid for stereolithographic 3D bioprinting of tissue-engineered cartilage. J. Biomed. Mater. Res. Part. B Appl. Biomater. 2019, 107, 2649–2657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Genova, T.; Roato, I.; Carossa, M.; Motta, C.; Cavagnetto, D.; Mussano, F. Advances on Bone Substitutes through 3D Bioprinting. Int. J. Mol. Sci. 2020, 21, 7012. [Google Scholar] [CrossRef]
- Orciani, M.; Fini, M.; Di Primio, R.; Mattioli-Belmonte, M. Biofabrication and Bone Tissue Regeneration: Cell Source, Approaches, and Challenges. Front. Bioeng. Biotechnol. 2017, 5, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Midha, S.; Dalela, M.; Sybil, D.; Patra, P.; Mohanty, S. Advances in three-dimensional bioprinting of bone: Progress and challenges. J. Tissue Eng. Regen. Med. 2019, 13, 925–945. [Google Scholar] [CrossRef]
- Scognamiglio, C.; Soloperto, A.; Ruocco, G.; Cidonio, G. Bioprinting stem cells: Building physiological tissues one cell at a time. Am. J. Physiol. Cell Physiol. 2020, 319, C465–C480. [Google Scholar] [CrossRef]
- Neumann, K.; Dehne, T.; Endres, M.; Erggelet, C.; Kaps, C.; Ringe, J.; Sittinger, M. Chondrogenic differentiation capacity of human mesenchymal progenitor cells derived from subchondral cortico-spongious bone. J. Orthop. Res. 2008, 26, 1449–1456. [Google Scholar] [CrossRef]
- Leberfinger, A.N.; Ravnic, D.J.; Dhawan, A.; Ozbolat, I.T. Concise Review: Bioprinting of Stem Cells for Transplantable Tissue Fabrication. Stem Cells Transl. Med. 2017, 6, 1940–1948. [Google Scholar] [CrossRef] [Green Version]
- Secunda, R.; Vennila, R.; Mohanashankar, A.M.; Rajasundari, M.; Jeswanth, S.; Surendran, R. Isolation, expansion and characterisation of mesenchymal stem cells from human bone marrow, adipose tissue, umbilical cord blood and matrix: A comparative study. Cytotechnology 2015, 67, 793–807. [Google Scholar] [CrossRef]
- El-Jawhari, J.J.; Kleftouris, G.; El-Sherbiny, Y.; Saleeb, H.; West, R.M.; Jones, E.; Giannoudis, P.V. Defective Proliferation and Osteogenic Potential with Altered Immunoregulatory phenotype of Native Bone marrow-Multipotential Stromal Cells in Atrophic Fracture Non-Union. Sci. Rep. 2019, 9, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Ferretti, C.; Mattioli-Belmonte, M. Periosteum derived stem cells for regenerative medicine proposals: Boosting current knowledge. World J. Stem Cells 2014, 6, 266–277. [Google Scholar] [CrossRef]
- Rampersad, S.N. Multiple Applications of Alamar Blue as an Indicator of Metabolic Function and Cellular Health in Cell Viability Bioassays. Sensors 2012, 12, 12347–12360. [Google Scholar] [CrossRef] [PubMed]
- Garant, P.R. Oral Cells and Tissues; Quintessence Publishing Company: Chicago, IL, USA, 2003. [Google Scholar]
- Schroeder, H.E. The Periodontium. The Handbook of Microscopic Anatomy; Springer: Berlin/Heidelberg, Germany, 1986; Volume 5. [Google Scholar]
- Kuhn, L.T. Bone mineralization. In Encyclopedia of Materials: Science and Technology, 2nd ed.; Buschow, K.H.J., Cahn, R.W., Flemings, M.C., Ilschner, B., Kramer, E.J., Mahajan, S., Veyssière, P., Eds.; Elsevier: Amsterdam, The Netherlands, 2001; pp. 787–794. [Google Scholar]
- Orimo, H. The Mechanism of Mineralization and the Role of Alkaline Phosphatase in Health and Disease. J. Nippon. Med. Sch. 2010, 77, 4–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.S.; Kim, M.E.; Seon, J.K.; Kang, J.Y.; Yoon, T.R.; Park, Y.-D.; Kim, H.K. Bone-forming peptide-3 induces osteogenic differentiation of bone marrow stromal cells via regulation of the ERK1/2 and Smad1/5/8 pathways. Stem Cell Res. 2018, 26, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Kirkham, G.R.; Cartmell, S.H. Genes and Proteins Involved in the Regulation of Osteogenesis. In Topics in Tissue Engineering; Ashammakhi, N., Reis, R.L., Chiellini, E., Eds.; Oulu University: Oulu, Finland, 2007; Volume 3. [Google Scholar]
- Atala, A.; Yoo, J.J. Essentials of 3D Biofabrication and Translation; Academic Press: Cambridge, MA, USA, 2015. [Google Scholar]
- Sodek, J.; McKee, M.D. Molecular and cellular biology of alveolar bone. Periodontol. 2000 2000, 24, 99–126. [Google Scholar] [CrossRef]
- Altmann, B.; Löchner, A.; Swain, M.; Kohal, R.-J.; Giselbrecht, S.; Gottwald, E.; Steinberg, T.; Tomakidi, P. Differences in morphogenesis of 3D cultured primary human osteoblasts under static and microfluidic growth conditions. Biomaterials 2014, 35, 3208–3219. [Google Scholar] [CrossRef]
- Wein, M.; Huelter-Hassler, D.; Nelson, K.; Fretwurst, T.; Nahles, S.; Finkenzeller, G.; Altmann, B.; Steinberg, T. Differential osteopontin expression in human osteoblasts derived from iliac crest and alveolar bone and its role in early stages of angiogenesis. J. Bone Miner. Metab. 2018, 37, 105–117. [Google Scholar] [CrossRef]
- McBeth, C.; Lauer, J.; Ottersbach, M.; Campbell, J.; Sharon, A.; Sauer-Budge, A.F. 3D bioprinting of GelMA scaffolds triggers mineral deposition by primary human osteoblasts. Biofabrication 2017, 9, 015009. [Google Scholar] [CrossRef]
- Park, J.Y.; Choi, J.-C.; Shim, J.-H.; Lee, J.-S.; Park, H.; Kim, S.W.; Doh, J.; Cho, D.-W. A comparative study on collagen type I and hyaluronic acid dependent cell behavior for osteochondral tissue bioprinting. Biofabrication 2014, 6, 035004. [Google Scholar] [CrossRef]
- Liu, Y.; Chan-Park, M.B. A biomimetic hydrogel based on methacrylated dextran-graft-lysine and gelatin for 3D smooth muscle cell culture. Biomaterials 2010, 31, 1158–1170. [Google Scholar] [CrossRef]
- Amler, A.-K.; Thomas, A.; Tüzüner, S.; Lam, T.; Geiger, M.-A.; Kreuder, A.-E.; Palmer, C.; Nahles, S.; Lauster, R.; Kloke, L. A 3D bioprinted human jawbone model. Sci. Rep. Under Review.
- Pattappa, G.; Heywood, H.K.; De Bruijn, J.D.; Lee, D.A. The metabolism of human mesenchymal stem cells during proliferation and differentiation. J. Cell. Physiol. 2011, 226, 2562–2570. [Google Scholar] [CrossRef]
- Long, F. Building strong bones: Molecular regulation of the osteoblast lineage. Nat. Rev. Mol. Cell Biol. 2012, 13, 27–38. [Google Scholar] [CrossRef] [PubMed]
- Miron, R.J.; Zhang, Y. Osteoinduction: A Review of Old Concepts with New Standards. J. Dent. Res. 2012, 91, 736–744. [Google Scholar] [CrossRef] [PubMed]
- Ducy, P.; Starbuck, M.; Priemel, M.; Shen, J.; Pinero, G.; Geoffroy, V.; Amling, M.; Karsenty, G. A Cbfa1-dependent genetic pathway controls bone formation beyond embryonic development. Genes Dev. 1999, 13, 1025–1036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Czekanska, E.M.; Stoddart, M.J.; Ralphs, J.R.; Richards, R.G.; Hayes, J.S. A phenotypic comparison of osteoblast cell lines versus human primary osteoblasts for biomaterials testing. J. Biomed. Mater. Res. Part. A 2014, 102, 2636–2643. [Google Scholar] [CrossRef] [PubMed]
- Stich, S.; Loch, A.; Leinhase, I.; Neumann, K.; Kaps, C.; Sittinger, M.; Ringe, J. Human periosteum-derived progenitor cells express distinct chemokine receptors and migrate upon stimulation with CCL2, CCL25, CXCL8, CXCL12, and CXCL13. Eur. J. Cell Biol. 2008, 87, 365–376. [Google Scholar] [CrossRef] [PubMed]
- Almela, T.; Al-Sahaf, S.; Bolt, R.; Brook, I.M.; Moharamzadeh, K. Characterization of Multilayered Tissue-Engineered Human Alveolar Bone and Gingival Mucosa. Tissue Eng. Part. C Methods 2018, 24, 99–107. [Google Scholar] [CrossRef]
- Wein, M.; Fretwurst, T.; Nahles, S.; Duttenhoefer, F.; Tomakidi, P.; Steinberg, T.; Nelson, K. Pilot investigation of the molecular discrimination of human osteoblasts from different bone entities. J. Cranio-Maxillofac. Surg. 2015, 43, 1487–1493. [Google Scholar] [CrossRef] [PubMed]
- Thitiset, T.; Damrongsakkul, S.; Bunaprasert, T.; Leeanansaksiri, W.; Honsawek, S. Development of Collagen/Demineralized Bone Powder Scaffolds and Periosteum-Derived Cells for Bone Tissue Engineering Application. Int. J. Mol. Sci. 2013, 14, 2056–2071. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Y.-X.; Ringe, J.; Liang, Z.; Loch, A.; Chen, L.; Sittinger, M. Osteogenic potential of human periosteum-derived progenitor cells in PLGA scaffold using allogeneic serum. J. Zhejiang Univ. Sci. B 2006, 7, 817–824. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ringe, J.; Leinhase, I.; Stich, S.; Loch, A.; Neumann, K.; Haisch, A.; Häup, T.; Manz, R.; Kaps, C.; Sittinger, M. Human mastoid periosteum-derived stem cells: Promising candidates for skeletal tissue engineering. J. Tissue Eng. Regen. Med. 2008, 2, 136–146. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.; Tate, M.L.K. Concise Review: The Periosteum: Tapping into a Reservoir of Clinically Useful Progenitor Cells. Stem Cells Transl. Med. 2012, 1, 480–491. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.-K.; Nakata, H.; Yamamoto, M.; Miyasaka, M.; Kasugai, S.; Kuroda, S. Osteogenic Potential of Mouse Periosteum-Derived Cells Sorted for CD90 In Vitro and In Vivo. Stem Cells Transl. Med. 2016, 5, 227–234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kreuz, P.C.; Steinwachs, M.; Erggelet, C.; Krause, S.J.; Ossendorf, C.; Maier, D.; Ghanem, N.; Uhl, M.; Haag, M. Classification of graft hypertrophy after autologous chondrocyte implantation of full-thickness chondral defects in the knee. Osteoarthr. Cartil. 2007, 15, 1339–1347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trautvetter, W.; Kaps, C.; Schmelzeisen, R.; Sauerbier, S.; Sittinger, M. Tissue-Engineered Polymer-Based Periosteal Bone Grafts for Maxillary Sinus Augmentation: Five-Year Clinical Results. J. Oral Maxillofac. Surg. 2011, 69, 2753–2762. [Google Scholar] [CrossRef] [PubMed]
- Murphy, S.V.; Atala, A. 3D bioprinting of tissues and organs. Nat. Biotechnol. 2014, 32, 773–785. [Google Scholar] [CrossRef]
- Kaempfen, A.; Todorov, A.; Güven, S.; Largo, R.D.; Claude, J.; Scherberich, A.; Martin, I.; Schaefer, D.J. Engraftment of Prevascularized, Tissue Engineered Constructs in a Novel Rabbit Segmental Bone Defect Model. Int. J. Mol. Sci. 2015, 16, 12616–12630. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Zhou, Y.; Yu, N.; Ma, H.; Wang, K.; Liu, J.; Zhang, W.; Cai, Z.; He, Y. Construction of vascularized tissue-engineered bone with polylysine-modified coral hydroxyapatite and a double cell-sheet complex to repair a large radius bone defect in rabbits. Acta Biomater. 2019, 91, 82–98. [Google Scholar] [CrossRef]
- Thomas, A.; Orellano, I.; Lam, T.; Noichl, B.; Geiger, M.-A.; Amler, A.-K.; Kreuder, A.-E.; Palmer, C.; Duda, G.; Lauster, R.; et al. Vascular bioprinting with enzymatically degradable bioinks via multi-material projection-based stereolithography. Acta Biomater. 2020, 117, 121–132. [Google Scholar] [CrossRef]
- Chiesa, I.; De Maria, C.; Lapomarda, A.; Fortunato, G.M.; Montemurro, F.; Di Gesù, R.; Tuan, R.S.; Vozzi, G.; Gottardi, R. Endothelial cells support osteogenesis in an in vitro vascularized bone model developed by 3D bioprinting. Biofabrication 2020, 12, 025013. [Google Scholar] [CrossRef] [PubMed]
- Mikos, A.G.; Herring, S.W.; Ochareon, P.; Elisseeff, J.; Lu, H.H.; Kandel, R.; Schoen, F.J.; Toner, M.; Mooney, D.; Atala, A.; et al. Engineering Complex Tissues. Tissue Eng. 2006, 12, 3307–3339. [Google Scholar] [CrossRef] [PubMed]
- Du, M.; Chen, B.; Meng, Q.; Liu, S.; Zheng, X.; Zhang, C.; Wang, H.; Li, H.; Wang, N.; Dai, J. 3D bioprinting of BMSC-laden methacrylamide gelatin scaffolds with CBD-BMP2-collagen microfibers. Biofabrication 2015, 7, 044104. [Google Scholar] [CrossRef] [PubMed]
- Koolen, M.; Longoni, A.; Van Der Stok, J.; Van Der Jagt, O.; Gawlitta, D.; Weinans, H. Complete regeneration of large bone defects in rats with commercially available fibrin loaded with BMP-2. Eur. Cells Mater. 2019, 38, 94–105. [Google Scholar] [CrossRef] [PubMed]
- Ringe, J.; Strassburg, S.; Neumann, K.; Endres, M.; Notter, M.; Burmester, G.-R.; Kaps, C.; Sittinger, M. Towards in situ tissue repair: Human mesenchymal stem cells express chemokine receptors CXCR1, CXCR2 and CCR2, and migrate upon stimulation with CXCL8 but not CCL2. J. Cell. Biochem. 2007, 101, 135–146. [Google Scholar] [CrossRef] [PubMed]
- Endres, M.; Andreas, K.; Kalwitz, G.; Freymann, U.; Neumann, K.; Ringe, J.; Sittinger, M.; Häupl, T.; Kaps, C. Chemokine profile of synovial fluid from normal, osteoarthritis and rheumatoid arthritis patients: CCL25, CXCL10 and XCL1 recruit human subchondral mesenchymal progenitor cells. Osteoarthr. Cartil. 2010, 18, 1458–1466. [Google Scholar] [CrossRef] [Green Version]
- Daly, A.C.; Pitacco, P.; Nulty, J.; Cunniffe, G.M.; Kelly, D.J. 3D printed microchannel networks to direct vascularisation during endochondral bone repair. Biomaterials 2018, 162, 34–46. [Google Scholar] [CrossRef]
- Daly, A.C.; Cunniffe, G.M.; Sathy, B.N.; Jeon, O.; Alsberg, E.; Kelly, D.J. 3D Bioprinting of Developmentally Inspired Templates for Whole Bone Organ Engineering. Adv. Healthc. Mater. 2016, 5, 2353–2362. [Google Scholar] [CrossRef]
- Van Den Bulcke, A.I.; Bogdanov, B.; De Rooze, N.; Schacht, E.H.; Cornelissen, M.; Berghmans, H. Structural and Rheological Properties of Methacrylamide Modified Gelatin Hydrogels. Biomacromolecules 2000, 1, 31–38. [Google Scholar] [CrossRef]
- Shirahama, H.; Lee, B.H.; Tan, L.P.; Cho, N.-J. Precise Tuning of Facile One-Pot Gelatin Methacryloyl (GelMA) Synthesis. Sci. Rep. 2016, 6, 31036. [Google Scholar] [CrossRef] [Green Version]
- Majima, T.; Schnabel, W.; Weber, W. Phenyl-2, 4, 6-trimethylbenzoylphosphinates as water-soluble photoinitiators. Generation and reactivity of O Ṗ (C6H5)(O−) radical anions. Die Makromol. Chem. 1991, 192, 2307–2315. [Google Scholar] [CrossRef]
- Fairbanks, B.D.; Schwartz, M.P.; Bowman, C.N.; Anseth, K.S. Photoinitiated polymerization of PEG-diacrylate with lithium phenyl-2,4,6-trimethylbenzoylphosphinate: Polymerization rate and cytocompatibility. Biomaterials 2009, 30, 6702–6707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ullah, M.; Eucker, J.; Sittinger, M.; Ringe, J. Mesenchymal stem cells and their chondrogenic differentiated and dedifferentiated progeny express chemokine receptor CCR9 and chemotactically migrate toward CCL25 or serum. Stem Cell Res. Ther. 2013, 4, 99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trentz, O.A.; Arikketh, D.; Sentilnathan, V.; Hemmi, S.; Handschin, A.E.; De Rosario, B.; Mohandas, P.V.A. Surface proteins and osteoblast markers: Characterization of human adipose tissue-derived osteogenic cells. Eur. J. Trauma Emerg. Surg. 2010, 36, 457–463. [Google Scholar] [CrossRef] [PubMed]
- Kruger, J.P.; Enz, A.; Hondke, S.; Wichelhaus, A.; Endres, M.; Mittlmeier, T. Proliferation, migration and differentiation potential of human mesenchymal progenitor cells derived from osteoarthritic subchondral cancellous bone. J. Stem Cells Regen. Med. 2018, 14, 45–52. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Construct | Cell Type | Biopsy Site | Biopsy Method | Expansion Method |
|---|---|---|---|---|
| Cell-free | – | – | – | – |
| aB-MPC | Bone-derived MPC | Alveolar bone | Bone explantation | Explant outgrowth |
| iB-MPC | Iliac crest | |||
| fB-MPC | Fibula | |||
| BM-MPC1 | Bone marrow MPC | Iliac crest bone marrow | Fine needle aspiration | Direct seeding |
| BM-MPC2 | ||||
| P-MPC1 | Periosteal MPC | Mastoid | Periosteum explantation | Seeding after tissue digestion |
| P-MPC2 |
| Gene | Accession Number | Description | For | Rev |
|---|---|---|---|---|
| ALPL | NM_000478 | Alkaline phosphatase | cccacttcatctggaaccgc | ccgtggtcaattctgcctcc |
| COL1A1 | NM_000088 | Collagen type I alpha 1 chain | gccgtgacctcaagatgtg | gccgaaccagacatgcctc |
| RUNX2 | NM_001015051 | Runt-related transcription factor 2 | tcacaaatcctccccaagtagc | ggcgggacacctactctcatac |
| SPARC | NM_003118 | Secreted protein acidic and cysteine rich | gcagaagctgcgggtgaagaa | ctcgaaaaagcgggtggtgc |
| SPP1 | NM_000582 | Secreted phosphoprotein 1 | cactgattttcccacggacct | ccattcaactcctcgctttcc |
| TBP | NM_003194 | TATA-box binding protein | ccttgtgctcacccaccaac | tcgtcttcctgaatccctttagaatag |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Amler, A.-K.; Dinkelborg, P.H.; Schlauch, D.; Spinnen, J.; Stich, S.; Lauster, R.; Sittinger, M.; Nahles, S.; Heiland, M.; Kloke, L.; et al. Comparison of the Translational Potential of Human Mesenchymal Progenitor Cells from Different Bone Entities for Autologous 3D Bioprinted Bone Grafts. Int. J. Mol. Sci. 2021, 22, 796. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22020796
Amler A-K, Dinkelborg PH, Schlauch D, Spinnen J, Stich S, Lauster R, Sittinger M, Nahles S, Heiland M, Kloke L, et al. Comparison of the Translational Potential of Human Mesenchymal Progenitor Cells from Different Bone Entities for Autologous 3D Bioprinted Bone Grafts. International Journal of Molecular Sciences. 2021; 22(2):796. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22020796
Chicago/Turabian StyleAmler, Anna-Klara, Patrick H. Dinkelborg, Domenic Schlauch, Jacob Spinnen, Stefan Stich, Roland Lauster, Michael Sittinger, Susanne Nahles, Max Heiland, Lutz Kloke, and et al. 2021. "Comparison of the Translational Potential of Human Mesenchymal Progenitor Cells from Different Bone Entities for Autologous 3D Bioprinted Bone Grafts" International Journal of Molecular Sciences 22, no. 2: 796. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22020796