
Characteristics and Therapeutic Potential of Human Amnion-Derived Stem Cells

Abstract
:1. Introduction
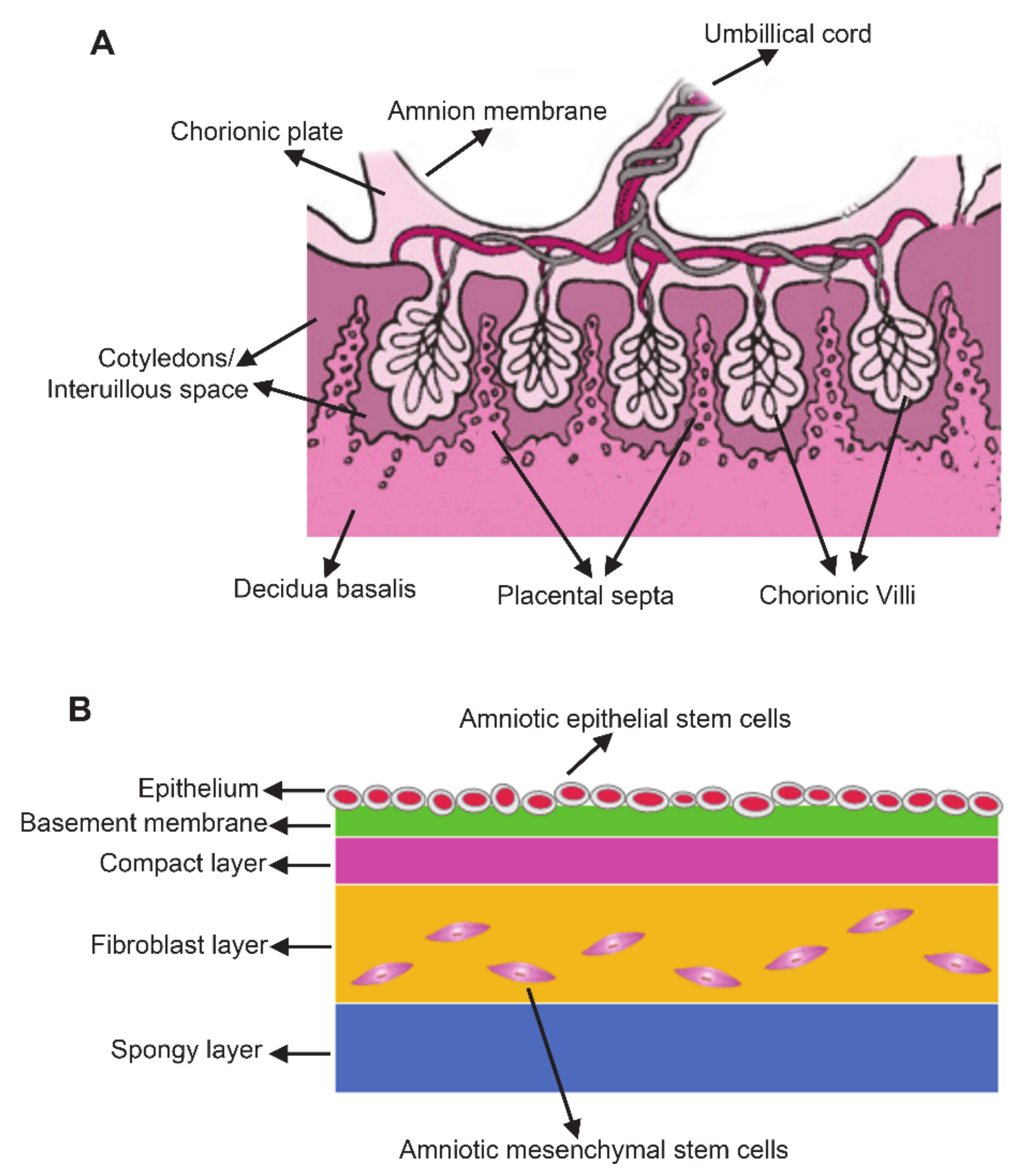
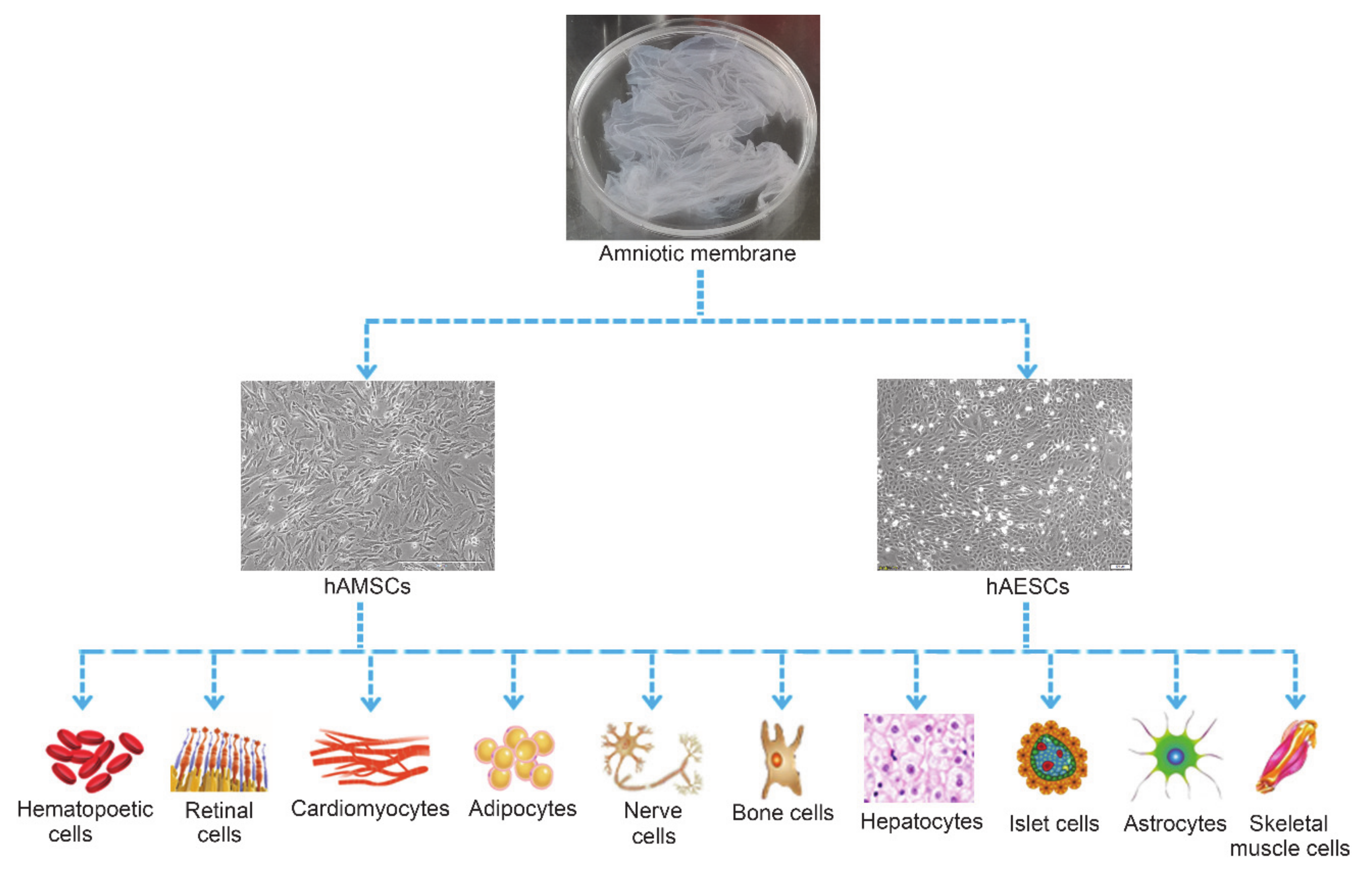
2. Characteristics of Human Amnion-Derived Stem Cells
2.1. Advantages of hADSCs over Other Stem Cells
2.2. The Molecular Markers of hADSCs
2.3. High Pluripotency of hADSCs
2.4. Immunosuppressive and Immunomodulatory Effects of hADSCs
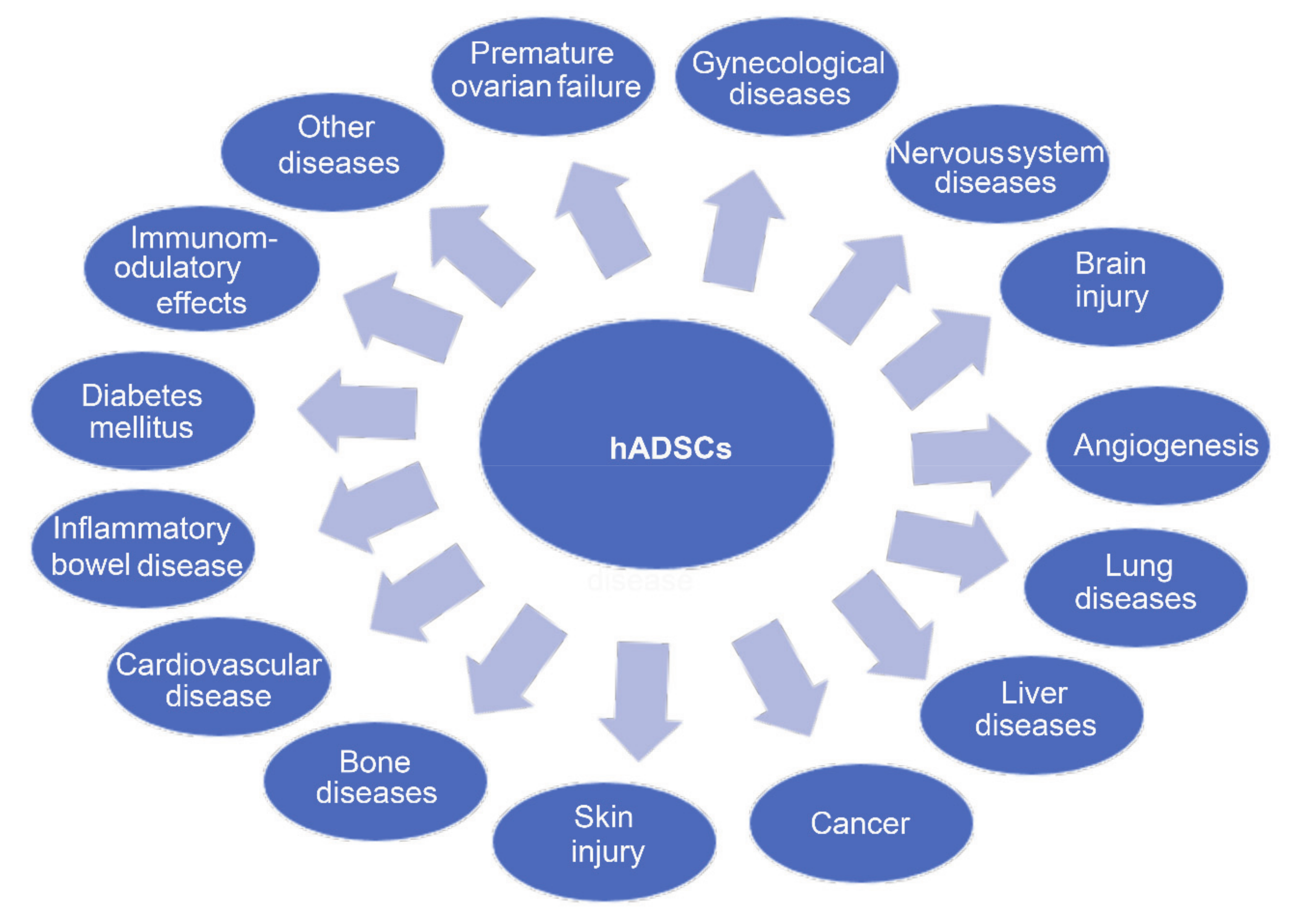
3. Cell-Based Therapy with Human Amnion-Derived Stem Cells
3.1. Gynecological Diseases
3.2. Nervous System Diseases
3.3. Brain Injury
3.4. Lung Diseases
3.5. Liver Diseases
3.6. Cancer
3.7. Skin Diseases
3.8. Bone Diseases
3.9. Cardiovascular Disease
3.10. Inflammatory Bowel Diseases
3.11. Kidney Disease
3.12. Diabetes Mellitus
3.13. Autoimmune Diseases
3.14. Other Diseases
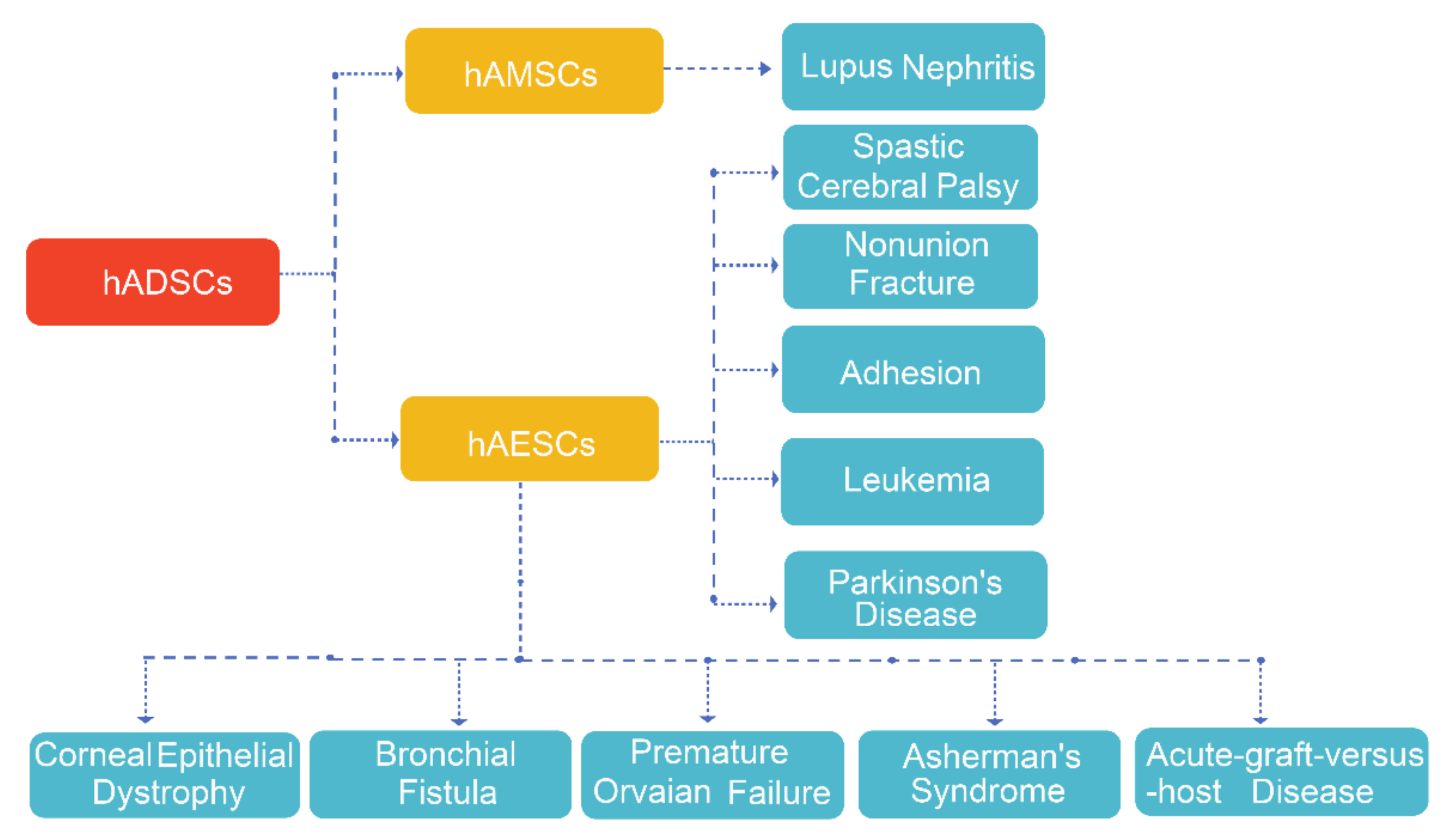
4. Clinical Trials with hAMSCs and hAESCs
5. hADSCs Clinical Applications
5.1. The Possible Therapeutic Mechanisms of hADSCs in Treating Diseases
5.2. The Challenges of hADSCs Applications
6. Conclusions and Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| hADSCs | human amnion-derived stem cells |
| PSCs | pluripotent stem cells |
| ESCs | embryonic stem cells |
| iPSCs | induced pluripotent stem cells |
| ASCs | adult stem cells |
| hAMSCs | human amnion mesenchymal stem cells |
| hAESCs | human amniotic epithelial stem cells |
| HLA-ABC | histocompatibility class I antigen |
| HLA-DR | histocompatibility class II antigen |
| POF | Premature ovarian failure |
| GC | granulosa cell |
| SCI | Spinal Cord Injury |
| AD | Alzheimer’s Disease |
| ASD | Autism spectrum disorder |
| MPS1 | Mucopolysaccharidosis type 1 |
| PD | Parkinson’s disease |
| BDNF | brain derived neurotrophic factor |
| ICH | Intracerebral hemorrhage |
| EAE | Encephalomyelitis |
| BPD | bronchopulmonary dysplasia |
| CF | cystic fibrosis |
| COPD | Chronic obstructive pulmonary disease |
| ALI | Acute liver injury |
| HLCs | hepatocyte-like cells |
| KCs | Kupffer cells |
| ECM | extracellular matrix |
| HSCs | hepatic stellate cells |
| α-SMA | αlpha-smooth muscle actin |
| MSUD | Maple syrup urine disease |
| BCAA | branched-chain amino acids |
| TE | tissue-engineered |
| OA | Osteoarthritis |
| CAI | collagen-induced arthritis |
| DM | diabetes mellitus |
| BM-MSCs | bone marrow mesenchymal stem cells |
| HAM | Human amniotic membrane |
| AS | Atherosclerosis |
| MI | Myocardial infarction |
| VEGF | vascular endothelial growth factor |
| HF | Heart failure |
| IBD | inflammatory bowel disease |
| CKF | chronic kidney failure |
| AKI | Acute kidney injury |
| ICs | islet cells |
| PBMC | peripheral blood mononuclear cell |
| MS | Multiple sclerosis |
| EAU | experimental autoimmune uveitis |
| SLE | Systemic lupus erythematosus |
| PE | Pre-eclampsia |
| NPC | Niemann–Pick disease type C1 |
| VML | volumetric muscle loss |
References
- De los Angeles, A.; Ferrari, F.; Xi, R.; Fujiwara, Y.; Benvenisty, N.; Deng, H.; Hochedlinger, K.; Jaenisch, R.; Lee, S.; Leitch, H.G.; et al. Hallmarks of pluripotency. Nature 2015, 525, 469–478. [Google Scholar] [CrossRef] [PubMed]
- Yagi, M.; Yamanaka, S.; Yamada, Y. Epigenetic foundations of pluripotent stem cells that recapitulate in vivo pluripotency. Lab. Investig. 2017, 97, 1133–1141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, N.H.; Hwang, K.A.; Kim, S.U.; Kim, Y.B.; Hyun, S.H.; Jeung, E.B.; Choi, K.C. Potential antitumor therapeutic strategies of human amniotic membrane and amniotic fluid-derived stem cells. Cancer Gene Ther. 2012, 19, 517–522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, C.-Y.; Liu, J.; Wan, F.; Tian, M.; Zhang, Y.-L.; He, Q.-H.; Si, Y.-C. Effects of Tianqijiangtang capsule on survival, self-renewal and differentiation of hippocampal neural stem cells of embryonic rats cultured in high glucose medium. Am. J. Transl. Res. 2019, 11, 5560–5572. [Google Scholar]
- Wang, C.; Spradling, A.C. An abundant quiescent stem cell population in Malpighian tubules protects principal cells from kidney stones. Elife 2020, 9, e54096. [Google Scholar] [CrossRef]
- Loukogeorgakis, S.P.; de Coppi, P. Concise review: Amniotic fluid stem cells: The known, the unknown, and potential regenerative medicine applications. Stem Cells 2017, 35, 1663–1673. [Google Scholar] [CrossRef] [Green Version]
- Vidane, A.S.; Souza, A.F.; Sampaio, R.V.; Bressan, F.F.; Pieri, N.C.; Martins, D.S.; Meirelles, F.V.; Miglino, M.A.; Ambrósio, C.E. Cat amniotic membrane multipotent cells are nontumorigenic and are safe for use in cell transplantation. Stem Cells Cloning 2014, 7, 71–78. [Google Scholar]
- Parolini, O.; Alviano, F.; Bagnara, G.P.; Bilic, G.; Buhring, H.J.; Evangelista, M.; Hennerbichler, S.; Liu, B.; Magatti, M.; Mao, N.; et al. Concise review: Isolation and characterization of cells from human term placenta: Outcome of the first international Workshop on Placenta Derived Stem Cells. Stem Cells 2008, 26, 300–311. [Google Scholar] [CrossRef] [Green Version]
- Xu, H.; Zhang, J.; Tsang, K.S.; Yang, H.; Gao, W.-Q. Therapeutic potential of human amniotic epithelial cells on injuries and disorders in the central nervous system. Stem Cells Int. 2019, 2019, 5432301. [Google Scholar] [CrossRef] [Green Version]
- Blum, B.; Benvenisty, N. The tumorigenicity of human embryonic stem cells. Adv. Cancer Res. 2008, 100, 133–158. [Google Scholar]
- Takahashi, K.; Yamanaka, S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell 2006, 126, 663–676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ankrum, J.A.; Ong, J.F.; Karp, J.M. Mesenchymal stem cells: Immune evasive, not immune privileged. Nat. Biotechnol. 2014, 32, 252–260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nauta, A.J.; Westerhuis, G.; Kruisselbrink, A.B.; Lurvink, E.G.A.; Willemze, R.; Fibbe, W.E. Donor-derived mesenchymal stem cells are immunogenic in an allogeneic host and stimulate donor graft rejection in a nonmyeloablative setting. Blood 2006, 108, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Blokzijl, F.; de Ligt, J.; Jager, M.; Sasselli, V.; Roerink, S.; Sasaki, N.; Huch, M.; Boymans, S.; Kuijk, E.; Prins, P.; et al. Tissue-specific mutation accumulation in human adult stem cells during life. Nature 2016, 538, 260–264. [Google Scholar] [CrossRef] [PubMed]
- Weeden, C.E.; Asselin-Labat, M.-L. Mechanisms of DNA damage repair in adult stem cells and implications for cancer formation. Biochim. Biophys. Acta Mol. Basis Dis. 2018, 1864, 89–101. [Google Scholar] [CrossRef] [PubMed]
- Abbasi-Kangevari, M.; Ghamari, S.-H.; Safaeinejad, F.; Bahrami, S.; Niknejad, H. Potential therapeutic features of human amniotic mesenchymal stem cells in multiple sclerosis: Immunomodulation, inflammation suppression, angiogenesis promotion, oxidative stress inhibition, neurogenesis induction, MMPs regulation, and remyelination stimulation. Front. Immunol. 2019, 10, 238. [Google Scholar]
- Magatti, M.; Vertua, E.; Cargnoni, A.; Silini, A.; Parolini, O. The immunomodulatory properties of amniotic cells: The two sides of the coin. Cell Transplant. 2018, 27, 31–44. [Google Scholar] [CrossRef] [Green Version]
- Muttini, A.; Barboni, B.; Valbonetti, L.; Russo, V.; Maffulli, N. Amniotic epithelial stem cells: Salient features and possible therapeutic role. Sports Med. Arthrosc. Rev. 2018, 26, 70–74. [Google Scholar] [CrossRef]
- Umezawa, A.; Hasegawa, A.; Inoue, M.; Tanuma-Takahashi, A.; Kajiwara, K.; Makino, H.; Chikazawa, E.; Okamoto, A. Amnion-derived cells as a reliable resource for next-generation regenerative medicine. Placenta 2019, 84, 50–56. [Google Scholar] [CrossRef]
- Pogozhykh, O.; Prokopyuk, V.; Figueiredo, C.; Pogozhykh, D. Placenta and placental derivatives in regenerative therapies: Experimental studies, history, and prospects. Stem Cells Int. 2018, 2018, 4837930. [Google Scholar] [CrossRef]
- Mathew, S.A.; Naik, C.; Cahill, P.A.; Bhonde, R.R. Placental mesenchymal stromal cells as an alternative tool for therapeutic angiogenesis. Cell. Mol. Life Sci. 2020, 77, 253–265. [Google Scholar] [CrossRef] [PubMed]
- Farhadihosseinabadi, B.; Farahani, M.; Tayebi, T.; Jafari, A.; Biniazan, F.; Modaresifar, K.; Moravvej, H.; Bahrami, S.; Redl, H.; Tayebi, L.; et al. Amniotic membrane and its epithelial and mesenchymal stem cells as an appropriate source for skin tissue engineering and regenerative medicine. Artif. Cells Nanomed. Biotechnol. 2018, 46, 431–440. [Google Scholar] [CrossRef]
- Liu, Q.W.; Liu, Q.Y.; Li, J.Y.; Wei, L.; Ren, K.K.; Zhang, X.C.; Ding, T.; Xiao, L.; Zhang, W.J.; Wu, H.Y.; et al. Therapeutic efficiency of human amniotic epithelial stem cell-derived functional hepatocyte-like cells in mice with acute hepatic failure. Stem Cell Res. Ther. 2018, 9, 321. [Google Scholar] [CrossRef] [PubMed]
- Li, J.Y.; Ren, K.K.; Zhang, W.J.; Xiao, L.; Wu, H.Y.; Liu, Q.Y.; Ding, T.; Zhang, X.C.; Nie, W.J.; Ke, Y.; et al. Human amniotic mesenchymal stem cells and their paracrine factors promote wound healing by inhibiting heat stress-induced skin cell apoptosis and enhancing their proliferation through activating PI3K/AKT signaling pathway. Stem Cell Res. Ther. 2019, 10, 247. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.-W.; Li, J.-Y.; Zhang, X.-C.; Liu, Y.; Liu, Q.-Y.; Xiao, L.; Zhang, W.-J.; Wu, H.-Y.; Deng, K.-Y.; Xin, H.-B. Human amniotic mesenchymal stem cells inhibit hepatocellular carcinoma in tumour-bearing mice. J. Cell. Mol. Med. 2020, 24, 10525–10541. [Google Scholar] [CrossRef]
- Miki, T. A rational strategy for the use of amniotic epithelial stem cell therapy for liver diseases. Stem Cells Transl. Med. 2016, 5, 405–409. [Google Scholar] [CrossRef] [Green Version]
- Insausti, C.L.; Blanquer, M.; Bleda, P.; Iniesta, P.; Majado, M.J.; Castellanos, G.; Moraleda, J.M. The amniotic membrane as a source of stem cells. Histol. Histopathol. 2010, 25, 91–98. [Google Scholar]
- Chiavegato, A.; Bollini, S.; Pozzobon, M.; Callegari, A.; Gasparotto, L.; Taiani, J.; Piccoli, M.; Lenzini, E.; Gerosa, G.; Vendramin, I.; et al. Human amniotic fluid-derived stem cells are rejected after transplantation in the myocardium of normal, ischemic, immuno-suppressed or immuno-deficient rat. J. Mol. Cell. Cardiol. 2007, 42, 746–759. [Google Scholar] [CrossRef]
- Srinivasan, R.C.; Strom, S.C.; Gramignoli, R. Effects of cryogenic storage on human amnion epithelial cells. Cells 2020, 9, 1696. [Google Scholar] [CrossRef]
- Murphy, S.; Rosli, S.; Acharya, R.; Mathias, L.; Lim, R.; Wallace, E.; Jenkin, G. Amnion epithelial cell isolation and characterization for clinical use. Curr. Protoc. Stem Cell Biol. 2010, 13, 1E.6.1–1E.6.25. [Google Scholar] [CrossRef]
- Banas, R.A.; Trumpower, C.; Bentlejewski, C.; Marshall, V.; Sing, G.; Zeevi, A. Immunogenicity and immunomodulatory effects of amnion-derived multipotent progenitor cells. Hum. Immunol. 2008, 69, 321–328. [Google Scholar] [CrossRef] [PubMed]
- Wolbank, S.; Peterbauer, A.; Fahrner, M.; Hennerbichler, S.; van Griensven, M.; Stadler, G.; Redl, H.; Gabriel, C. Dose-dependent immunomodulatory effect of human stem cells from amniotic membrane: A comparison with human mesenchymal stem cells from adipose tissue. Tissue Eng. 2007, 13, 1173–1183. [Google Scholar] [CrossRef] [PubMed]
- Pratama, G.; Vaghjiani, V.; Tee, J.Y.; Liu, Y.H.; Chan, J.; Tan, C.; Murthi, P.; Gargett, C.; Manuelpillai, U. Changes in culture expanded human amniotic epithelial cells: Implications for potential therapeutic applications. PLoS ONE 2011, 6, e26136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Coppi, P.; Bartsch, G.; Siddiqui, M.M.; Xu, T.; Santos, C.C.; Perin, L.; Mostoslavsky, G.; Serre, A.C.; Snyder, E.Y.; Yoo, J.J.; et al. Isolation of amniotic stem cell lines with potential for therapy. Nat. Biotechnol. 2007, 25, 100–106. [Google Scholar] [CrossRef]
- Yang, P.J.; Yuan, W.X.; Liu, J.; Li, J.Y.; Tan, B.; Qiu, C.; Zhu, X.L.; Qiu, C.; Lai, D.M.; Guo, L.H.; et al. Biological characterization of human amniotic epithelial cells in a serum-free system and their safety evaluation. Acta Pharmacol. Sin. 2018, 39, 1305–1306. [Google Scholar] [CrossRef]
- Miki, T.; Strom, S.C. Amnion-derived pluripotent/multipotent stem cells. Stem Cell Rev. 2006, 2, 133–142. [Google Scholar] [CrossRef]
- Ding, C.; Li, H.; Wang, Y.; Wang, F.; Wu, H.; Chen, R.; Lv, J.; Wang, W.; Huang, B. Different therapeutic effects of cells derived from human amniotic membrane on premature ovarian aging depend on distinct cellular biological characteristics. Stem Cell Res. Ther. 2017, 8, 173. [Google Scholar] [CrossRef] [Green Version]
- Wu, Q.; Fang, T.; Lang, H.; Chen, M.; Shi, P.; Pang, X.; Qi, G. Comparison of the proliferation, migration and angiogenic properties of human amniotic epithelial and mesenchymal stem cells and their effects on endothelial cells. Int. J. Mol. Med. 2017, 39, 918–926. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Castro, I.L.; Garcia-Lopez, G.; Avila-Gonzalez, D.; Flores-Herrera, H.; Molina-Hernandez, A.; Portillo, W.; Ramon-Gallegos, E.; Diaz, N.F. Markers of pluripotency in human amniotic epithelial cells and their differentiation to progenitor of cortical neurons. PLoS ONE 2015, 10, e0146082. [Google Scholar] [CrossRef] [Green Version]
- Evron, A.; Goldman, S.; Shalev, E. Human amniotic epithelial cells cultured in substitute serum medium maintain their stem cell characteristics for up to four passages. Int. J. Stem Cells 2011, 4, 123–132. [Google Scholar] [CrossRef] [Green Version]
- Miki, T.; Lehmann, T.; Cai, H.; Stolz, D.B.; Strom, S.C. Stem cell characteristics of amniotic epithelial cells. Stem Cells 2005, 23, 1549–1559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, X.; Gao, F.; Wu, Y.; Sun, R.; Guan, W.; Tian, X. Isolation and biological characteristics of sheep amniotic epithelial cells. Cytotechnology 2019, 71, 539–551. [Google Scholar] [CrossRef] [PubMed]
- Koike, C.; Zhou, K.; Takeda, Y.; Fathy, M.; Okabe, M.; Yoshida, T.; Nakamura, Y.; Kato, Y.; Nikaido, T. Characterization of amniotic stem cells. Cell. Reprogram. 2014, 16, 298–305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bačenková, D.; Rosocha, J.; Tóthová, T.; Rosocha, L.; Šarisský, M. Isolation and basic characterization of human term amnion and chorion mesenchymal stromal cells. Cytotherapy 2011, 13, 1047–1056. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Prado, S.; Muiños-López, E.; Hermida-Gómez, T.; Rendal-Vázquez, M.E.; Fuentes-Boquete, I.; de Toro, F.J.; Blanco, F.J. Multilineage differentiation potential of cells isolated from the human amniotic membrane. J. Cell. Biochem. 2010, 111, 846–857. [Google Scholar] [CrossRef] [Green Version]
- Sugiura, R.; Ohnishi, S.; Ohara, M.; Ishikawa, M.; Miyamoto, S.; Onishi, R.; Yamamoto, K.; Kawakubo, K.; Kuwatani, M.; Sakamoto, N. Effects of human amnion-derived mesenchymal stem cells and conditioned medium in rats with sclerosing cholangitis. Am. J. Transl. Res. 2018, 10, 2102–2114. [Google Scholar] [PubMed]
- Borghesi, J.; Ferreira Lima, M.; Mario, L.C.; da Anunciação, d.A.A.R.; Silveira Rabelo, A.C.; da Silva, G.K.C.M.; Assunpção Fernandes, F.; Miglino, M.A.; Oliveira Carreira, A.C.; Oliveira Favaron, P. Canine amniotic membrane mesenchymal stromal/stem cells: Isolation, characterization and differentiation. Tissue Cell 2019, 58, 99–106. [Google Scholar] [CrossRef]
- Mihu, C.M.; Rus Ciucă, D.; Soritău, O.; Suşman, S.; Mihu, D. Isolation and characterization of mesenchymal stem cells from the amniotic membrane. Rom. J. Morphol. Embryol. 2009, 50, 73–77. [Google Scholar]
- Seo, M.-S.; Park, S.-B.; Kim, H.-S.; Kang, J.-G.; Chae, J.-S.; Kang, K.-S. Isolation and characterization of equine amniotic membrane-derived mesenchymal stem cells. J. Vet. Sci. 2013, 14, 151–159. [Google Scholar] [CrossRef] [Green Version]
- Nogami, M.; Tsuno, H.; Koike, C.; Okabe, M.; Yoshida, T.; Seki, S.; Matsui, Y.; Kimura, T.; Nikaido, T. Isolation and characterization of human amniotic mesenchymal stem cells and their chondrogenic differentiation. Transplantation 2012, 93, 1221–1228. [Google Scholar] [CrossRef] [Green Version]
- Díaz-Prado, S.; Muiños-López, E.; Hermida-Gómez, T.; Rendal-Vázquez, M.E.; Fuentes-Boquete, I.; de Toro, F.J.; Blanco, F.J. Isolation and characterization of mesenchymal stem cells from human amniotic membrane. Tissue Eng. Part C Methods 2011, 17, 49–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, M.S.; Hwang, S.M.; Tsai, Y.L.; Cheng, F.C.; Lee, J.L.; Chang, Y.J. Clonal amniotic fluid-derived stem cells express characteristics of both mesenchymal and neural stem cells. Biol. Reprod. 2006, 74, 545–551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bollini, S.; Pozzobon, M.; Nobles, M.; Riegler, J.; Dong, X.; Piccoli, M.; Chiavegato, A.; Price, A.N.; Ghionzoli, M.; Cheung, K.K.; et al. In vitro and in vivo cardiomyogenic differentiation of amniotic fluid stem cells. Stem Cell Rev. 2011, 7, 364–380. [Google Scholar] [CrossRef]
- Pisciotta, A.; Riccio, M.; Carnevale, G.; Lu, A.; de Biasi, S.; Gibellini, L.; la Sala, G.B.; Bruzzesi, G.; Ferrari, A.; Huard, J.; et al. Stem cells isolated from human dental pulp and amniotic fluid improve skeletal muscle histopathology in mdx/SCID mice. Stem Cell Res. Ther. 2015, 6, 156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, D.; Yan, K.; Zhou, J.; Xu, T.; Xu, M.; Lin, J.; Bai, J.; Ge, G.; Hu, D.; Si, W.; et al. Myogenic differentiation of human amniotic mesenchymal cells and its tissue repair capacity on volumetric muscle loss. J. Tissue Eng. 2019, 10, 2041731419887100. [Google Scholar] [CrossRef] [Green Version]
- Ditadi, A.; de Coppi, P.; Picone, O.; Gautreau, L.; Smati, R.; Six, E.; Bonhomme, D.; Ezine, S.; Frydman, R.; Cavazzana-Calvo, M.; et al. Human and murine amniotic fluid c-Kit+Lin− cells display hematopoietic activity. Blood 2009, 113, 3953–3960. [Google Scholar] [CrossRef] [Green Version]
- Perin, L.; Sedrakyan, S.; da Sacco, S.; de Filippo, R. Characterization of human amniotic fluid stem cells and their pluripotential capability. Methods Cell Biol. 2008, 86, 85–99. [Google Scholar]
- Rota, C.; Imberti, B.; Pozzobon, M.; Piccoli, M.; de Coppi, P.; Atala, A.; Gagliardini, E.; Xinaris, C.; Benedetti, V.; Fabricio, A.S.; et al. Human amniotic fluid stem cell preconditioning improves their regenerative potential. Stem Cells Dev. 2012, 21, 1911–1923. [Google Scholar] [CrossRef] [Green Version]
- Ghaderi, S.; Soheili, Z.S.; Ahmadieh, H.; Davari, M.; Jahromi, F.S.; Samie, S.; Rezaie-Kanavi, M.; Pakravesh, J.; Deezagi, A. Human amniotic fluid promotes retinal pigmented epithelial cells’ trans-differentiation into rod photoreceptors and retinal ganglion cells. Stem Cells Dev. 2011, 20, 1615–1625. [Google Scholar] [CrossRef]
- Decembrini, S.; Cananzi, M.; Gualdoni, S.; Battersby, A.; Allen, N.; Pearson, R.A.; Ali, R.R.; de Coppi, P.; Sowden, J.C. Comparative analysis of the retinal potential of embryonic stem cells and amniotic fluid-derived stem cells. Stem Cells Dev. 2011, 20, 851–863. [Google Scholar] [CrossRef]
- Vacz, G.; Cselenyak, A.; Cserep, Z.; Benko, R.; Kovacs, E.; Pankotai, E.; Lindenmair, A.; Wolbank, S.; Schwarz, C.M.; Horvathy, D.B.; et al. Effects of amniotic epithelial cell transplantation in endothelial injury. Interv. Med. Appl. Sci. 2016, 8, 164–171. [Google Scholar] [CrossRef] [PubMed]
- Jiang, F.; Ma, J.; Liang, Y.; Niu, Y.; Chen, N.; Shen, M. Amniotic mesenchymal stem cells can enhance angiogenic capacity via MMPs in vitro and in vivo. BioMed Res. Int. 2015, 2015, 324014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.-W.; Zhang, H.-Z.; Kim, C.E.; An, H.S.; Kim, J.-M.; Kim, M.H. Amniotic mesenchymal stem cells have robust angiogenic properties and are effective in treating hindlimb ischaemia. Cardiovasc. Res. 2012, 93, 525–534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Danieli, P.; Malpasso, G.; Ciuffreda, M.C.; Cervio, E.; Calvillo, L.; Copes, F.; Pisano, F.; Mura, M.; Kleijn, L.; de Boer, R.A.; et al. Conditioned medium from human amniotic mesenchymal stromal cells limits infarct size and enhances angiogenesis. Stem Cells Transl. Med. 2015, 4, 448–458. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.G.; Choi, O.H. Neovascularization in a mouse model via stem cells derived from human fetal amniotic membranes. Heart Vessel. 2011, 26, 196–205. [Google Scholar] [CrossRef]
- König, J.; Huppertz, B.; Desoye, G.; Parolini, O.; Fröhlich, J.D.; Weiss, G.; Dohr, G.; Sedlmayr, P.; Lang, I. Amnion-derived mesenchymal stromal cells show angiogenic properties but resist differentiation into mature endothelial cells. Stem Cells Dev. 2012, 21, 1309–1320. [Google Scholar] [CrossRef] [Green Version]
- Yuan, Z.; Bian, Y.; Ma, X.; Tang, Z.; Chen, N.; Shen, M. LncRNA H19 knockdown in human amniotic mesenchymal stem cells suppresses angiogenesis by associating with EZH2 and activating vasohibin-1. Stem Cells Dev. 2019, 28, 781–790. [Google Scholar] [CrossRef]
- Tang, Z.; Tan, J.; Yuan, X.; Zhou, Q.; Yuan, Z.; Chen, N.; Shen, M. Circular RNA-ABCB10 promotes angiogenesis induced by conditioned medium from human amnion-derived mesenchymal stem cells via the microRNA-29b-3p/vascular endothelial growth factor A axis. Exp. Ther. Med. 2020, 20, 2021–2030. [Google Scholar]
- Tang, Z.; Wu, X.; Hu, L.; Xiao, Y.; Tan, J.; Zuo, S.; Shen, M.; Yuan, X. Circ-100290 positively regulates angiogenesis induced by conditioned medium of human amnion-derived mesenchymal stem cells through miR-449a/eNOS and miR-449a/VEGFA axes. Int. J. Biol. Sci. 2020, 16, 2131–2144. [Google Scholar] [CrossRef]
- Rossi, D.; Pianta, S.; Magatti, M.; Sedlmayr, P.; Parolini, O. Characterization of the conditioned medium from amniotic membrane cells: Prostaglandins as key effectors of its immunomodulatory activity. PLoS ONE 2012, 7, e46956. [Google Scholar] [CrossRef]
- Bulati, M.; Miceli, V.; Gallo, A.; Amico, G.; Carcione, C.; Pampalone, M.; Conaldi, P.G. The immunomodulatory properties of the human amnion-derived mesenchymal stromal/stem cells are induced by INF-γ produced by activated lymphomonocytes and are mediated by cell-to-cell contact and soluble factors. Front. Immunol. 2020, 11, 54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dabrowski, F.A.; Burdzinska, A.; Kulesza, A.; Chlebus, M.; Kaleta, B.; Borysowski, J.; Zolocinska, A.; Paczek, L.; Wielgos, M. Mesenchymal stem cells from human amniotic membrane and umbilical cord can diminish immunological response in an in vitro allograft model. Gynecol. Obstet. Investig. 2017, 82, 267–275. [Google Scholar] [CrossRef] [PubMed]
- Meesuk, L.; Tantrawatpan, C.; Kheolamai, P.; Manochantr, S. The immunosuppressive capacity of human mesenchymal stromal cells derived from amnion and bone marrow. Biochem. Biophys. Rep. 2016, 8, 34–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morandi, F.; Marimpietri, D.; Görgens, A.; Gallo, A.; Srinivasan, R.C.; El-Andaloussi, S.; Gramignoli, R. Human amnion epithelial cells impair T Cell proliferation: The role of HLA-G and HLA-E molecules. Cells 2020, 9, 2123. [Google Scholar] [CrossRef]
- Carvajal, H.G.; Suárez-Meade, P.; Borlongan, C.V. Amnion-derived stem cell transplantation: A novel treatment for neurological disorders. Brain Circ. 2016, 2, 1–7. [Google Scholar]
- Han, K.; Lee, J.E.; Kwon, S.J.; Park, S.Y.; Shim, S.H.; Kim, H.; Moon, J.H.; Suh, C.S.; Lim, H.J. Human amnion-derived mesenchymal stem cells are a potential source for uterine stem cell therapy. Cell Prolif. 2008, 41, 709–725. [Google Scholar] [CrossRef]
- Ling, L.; Feng, X.; Wei, T.; Wang, Y.; Wang, Y.; Wang, Z.; Tang, D.; Luo, Y.; Xiong, Z. Human amnion-derived mesenchymal stem cell (hAD-MSC) transplantation improves ovarian function in rats with premature ovarian insufficiency (POI) at least partly through a paracrine mechanism. Stem Cell Res. Ther. 2019, 10, 46. [Google Scholar] [CrossRef] [Green Version]
- Feng, X.; Ling, L.; Zhang, W.; Liu, X.; Wang, Y.; Luo, Y.; Xiong, Z. Effects of human amnion-derived mesenchymal stem cell (hAD-MSC) transplantation in situ on primary ovarian insufficiency in SD rats. Reprod. Sci. 2020, 27, 1502–1512. [Google Scholar] [CrossRef]
- Ding, C.; Zou, Q.; Wang, F.; Wu, H.; Chen, R.; Lv, J.; Ling, M.; Sun, J.; Wang, W.; Li, H.; et al. Human amniotic mesenchymal stem cells improve ovarian function in natural aging through secreting hepatocyte growth factor and epidermal growth factor. Stem Cell Res. Ther. 2018, 9, 55. [Google Scholar] [CrossRef] [Green Version]
- Liu, R.; Zhang, X.; Fan, Z.; Wang, Y.; Yao, G.; Wan, X.; Liu, Z.; Yang, B.; Yu, L. Human amniotic mesenchymal stem cells improve the follicular microenvironment to recover ovarian function in premature ovarian failure mice. Stem Cell Res. Ther. 2019, 10, 299. [Google Scholar] [CrossRef]
- Fouad, H.; Sabry, D.; Elsetohy, K.; Fathy, N. Therapeutic efficacy of amniotic membrane stem cells and adipose tissue stem cells in rats with chemically induced ovarian failure. J. Adv. Res. 2016, 7, 233–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ling, L.; Feng, X.; Wei, T.; Wang, Y.; Wang, Y.; Zhang, W.; He, L.; Wang, Z.; Zeng, Q.; Xiong, Z. Effects of low-intensity pulsed ultrasound (LIPUS)-pretreated human amnion-derived mesenchymal stem cell (hAD-MSC) transplantation on primary ovarian insufficiency in rats. Stem Cell Res. Ther. 2017, 8, 283. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Bu, S.; Sun, J.; Xu, M.; Yao, X.; He, K.; Lai, D. Paracrine effects of human amniotic epithelial cells protect against chemotherapy-induced ovarian damage. Stem Cell Res. Ther. 2017, 8, 270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Q.; Xu, M.; Yao, X.; Li, T.; Wang, Q.; Lai, D. Human amniotic epithelial cells inhibit granulosa cell apoptosis induced by chemotherapy and restore the fertility. Stem Cell Res. Ther. 2015, 6, 152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, F.; Wang, L.; Yao, X.; Lai, D.; Guo, L. Human amniotic epithelial cells can differentiate into granulosa cells and restore folliculogenesis in a mouse model of chemotherapy-induced premature ovarian failure. Stem Cell Res. Ther. 2013, 4, 124. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Zhang, Q.; Sun, J.; Lai, D. Human amniotic epithelial cells improve fertility in an intrauterine adhesion mouse model. Stem Cell Res. Ther. 2019, 10, 257. [Google Scholar] [CrossRef] [Green Version]
- Ouyang, X.; You, S.; Zhang, Y.; Zhang, C.; Zhang, G.; Shao, X.; He, F.; Hu, L. Transplantation of human amnion epithelial cells improves endometrial regeneration in rat model of intrauterine adhesions. Stem Cells Dev. 2020, 29, 1346–1362. [Google Scholar] [CrossRef]
- Gan, L.; Duan, H.; Xu, Q.; Tang, Y.-Q.; Li, J.-J.; Sun, F.-Q.; Wang, S. Human amniotic mesenchymal stromal cell transplantation improves endometrial regeneration in rodent models of intrauterine adhesions. Cytotherapy 2017, 19, 603–616. [Google Scholar] [CrossRef]
- Motedayyen, H.; Rezaei, A.; Zarnani, A.-H.; Tajik, N. Human amniotic epithelial cells inhibit activation and pro-inflammatory cytokines production of naive CD4+ T cells from women with unexplained recurrent spontaneous abortion. Reprod. Biol. 2018, 18, 182–188. [Google Scholar] [CrossRef]
- Sebastià-Alcácer, V.; Alcanyis-Alberola, M.; Giner-Pascual, M.; Gomez-Pajares, F. Are the characteristics of the patient with a spinal cord injury changing? Spinal Cord 2014, 52, 29–33. [Google Scholar] [CrossRef]
- Zhou, H.-L.; Fang, H.; Luo, H.-T.; Ye, M.-H.; Yu, G.-Y.; Zhang, Y.; Mao, G.-H.; Gao, Z.-Y.; Cheng, Z.-J.; Zhu, X.-G. Intravenous administration of human amniotic mesenchymal stem cells improves outcomes in rats with acute traumatic spinal cord injury. Neuroreport 2020, 31, 730–736. [Google Scholar] [CrossRef] [PubMed]
- Sankar, V.; Muthusamy, R. Role of human amniotic epithelial cell transplantation in spinal cord injury repair research. Neuroscience 2003, 118, 11–17. [Google Scholar] [CrossRef]
- Wu, Z.Y.; Hui, G.Z.; Lu, Y.; Wu, X.; Guo, L.H. Transplantation of human amniotic epithelial cells improves hindlimb function in rats with spinal cord injury. Chin. Med. J. 2006, 119, 2101–2107. [Google Scholar] [CrossRef] [PubMed]
- Roh, D.H.; Seo, M.S.; Choi, H.S.; Park, S.B.; Han, H.J.; Beitz, A.J.; Kang, K.S.; Lee, J.H. Transplantation of human umbilical cord blood or amniotic epithelial stem cells alleviates mechanical allodynia after spinal cord injury in rats. Cell Transplant. 2013, 22, 1577–1590. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Bai, C.; Zheng, D.; Li, C.; Zhang, W.; Li, M.; Guan, W.; Ma, Y. Combination of melatonin and Wnt-4 promotes neural cell differentiation in bovine amniotic epithelial cells and recovery from spinal cord injury. J. Pineal Res. 2016, 60, 303–312. [Google Scholar] [CrossRef]
- Ibach, B.; Haen, E. Acetylcholinesterase inhibition in Alzheimer’s Disease. Curr. Pharm. Des. 2004, 10, 231–251. [Google Scholar] [CrossRef]
- Jiao, H.; Shi, K.; Zhang, W.; Yang, L.; Yang, L.; Guan, F.; Yang, B. Therapeutic potential of human amniotic membrane-derived mesenchymal stem cells in APP transgenic mice. Oncol. Lett. 2016, 12, 1877–1883. [Google Scholar] [CrossRef] [Green Version]
- Zheng, X.Y.; Wan, Q.Q.; Zheng, C.Y.; Zhou, H.L.; Dong, X.Y.; Deng, Q.S.; Yao, H.; Fu, Q.; Gao, M.; Yan, Z.J.; et al. Amniotic mesenchymal stem cells decrease Aβ deposition and improve memory in APP/PS1 transgenic mice. Neurochem. Res. 2017, 42, 2191–2207. [Google Scholar] [CrossRef]
- Xue, S.-R.; Chen, C.-F.; Dong, W.-L.; Hui, G.-Z.; Liu, T.-J.; Guo, L.-H. Intracerebroventricular transplantation of human amniotic epithelial cells ameliorates spatial memory deficit in the doubly transgenic mice coexpressing APPswe and PS1ΔE9-deleted genes. Chin. Med. J. 2011, 124, 2642–2648. [Google Scholar]
- Xue, S.; Chen, C.; Dong, W.; Hui, G.; Liu, T.; Guo, L. Therapeutic effects of human amniotic epithelial cell transplantation on double-transgenic mice co-expressing APPswe and PS1ΔE9-deleted genes. Sci. China Life Sci. 2012, 55, 132–140. [Google Scholar] [CrossRef] [Green Version]
- Ferdousi, F.; Kondo, S.; Sasaki, K.; Uchida, Y.; Ohkohchi, N.; Zheng, Y.W.; Isoda, H. Microarray analysis of verbenalin-treated human amniotic epithelial cells reveals therapeutic potential for Alzheimer’s Disease. Aging 2020, 12, 5516–5538. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.A.; Peñagarikano, O.; Belgard, T.G.; Swarup, V.; Geschwind, D.H. The emerging picture of autism spectrum disorder: Genetics and pathology. Annu. Rev. Pathol. 2015, 10, 111–144. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Cai, Y.; Xiao, R.; Zhong, H.; Li, X.; Guo, L.; Xu, H.; Fan, X. Human amniotic epithelial cell transplantation promotes neurogenesis and ameliorates social deficits in BTBR mice. Stem Cell Res. Ther. 2019, 10, 153. [Google Scholar] [CrossRef] [PubMed]
- Bie, H.; Yin, J.; He, X.; Kermode, A.R.; Goddard-Borger, E.D.; Withers, S.G.; James, M.N. Insights into mucopolysaccharidosis I from the structure and action of α-L-iduronidase. Nat. Chem. Biol. 2013, 9, 739–745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguez, N.S.; Yanuaria, L.; Parducho, K.M.R.; Garcia, I.M.; Varghese, B.A.; Grubbs, B.H.; Miki, T. Liver-directed human amniotic epithelial cell transplantation improves systemic disease phenotype in Hurler syndrome mouse model. Stem Cells Transl. Med. 2017, 6, 583–1594. [Google Scholar] [CrossRef] [Green Version]
- Olson, L. Regeneration in the adult central nervous system: Experimental repair strategies. Nat. Med. 1997, 3, 1329–1335. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Guo, L.; Ahn, H.S.; Kim, M.H.; Kim, S.-W. Amniotic mesenchymal stem cells display neurovascular tropism and aid in the recovery of injured peripheral nerves. J. Cell. Mol. Med. 2014, 18, 1028–1034. [Google Scholar] [CrossRef]
- Yang, X.; Song, L.; Wu, N.; Liu, Z.; Xue, S.; Hui, G. An experimental study on intracerebroventricular transplantation of human amniotic epithelial cells in a rat model of Parkinson’s disease. Neurol. Res. 2010, 32, 1054–1059. [Google Scholar] [CrossRef]
- Wei, L.; Wei, Z.Z.; Jiang, M.Q.; Mohamad, O.; Yu, S.P. Stem cell transplantation therapy for multifaceted therapeutic benefits after stroke. Prog. Neurobiol. 2017, 157, 49–78. [Google Scholar] [CrossRef]
- Kaneko, Y.; Hayashi, T.; Yu, S.; Tajiri, N.; Bae, E.C.; Solomita, M.A.; Chheda, S.H.; Weinbren, N.L.; Parolini, O.; Borlongan, C.V. Human amniotic epithelial cells express melatonin receptor MT1, but not melatonin receptor MT2: A new perspective to neuroprotection. J. Pineal Res. 2011, 50, 272–280. [Google Scholar] [CrossRef]
- Evans, M.A.; Lim, R.; Kim, H.A.; Chu, H.X.; Gardiner-Mann, C.V.; Taylor, K.W.E.; Chan, C.T.; Brait, V.H.; Lee, S.; Dinh, Q.N.; et al. Acute or delayed systemic administration of human amnion epithelial cells improves outcomes in experimental stroke. Stroke 2018, 49, 700–709. [Google Scholar] [CrossRef] [PubMed]
- Faezi, M.; Nasseri Maleki, S.; Aboutaleb, N.; Nikougoftar, M. The membrane mesenchymal stem cell derived conditioned medium exerts neuroprotection against focal cerebral ischemia by targeting apoptosis. J. Chem. Neuroanat. 2018, 94, 21–31. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Miao, Z.-N.; Xu, Y.-Y.; Zheng, S.-Y.; Qin, M.-D.; Gu, Y.-Z.; Zhang, X.-G. Transplantation of human amniotic mesenchymal stem cells in the treatment of focal cerebral ischemia. Mol. Med. Rep. 2012, 6, 625–630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nazarinia, D.; Aboutaleb, N.; Gholamzadeh, R.; Nasseri Maleki, S.; Mokhtari, B.; Nikougoftar, M. Conditioned medium obtained from human amniotic mesenchymal stem cells attenuates focal cerebral ischemia/reperfusion injury in rats by targeting mTOR pathway. J. Chem. Neuroanat. 2019, 102, 101707. [Google Scholar] [CrossRef]
- Aboutaleb, N.; Faezi, M.; Maleki, S.N.; Nazarinia, D.; Razavi Tousi, S.M.T.; Hashemirad, N. Conditioned medium obtained from mesenchymal stem cells attenuates focal cerebral ischemia reperfusion injury through activation of ERK1/ERK2-BDNF signaling pathway. J. Chem. Neuroanat. 2019, 97, 87–98. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Wu, J.; Huang, Q.; Hou, Y.; Jiang, Z.; Zang, S.; Guo, L. Human amniotic epithelial cells ameliorate behavioral dysfunction and reduce infarct size in the rat middle cerebral artery occlusion model. Shock 2008, 29, 603–611. [Google Scholar] [CrossRef] [PubMed]
- Tao, J.; Ji, F.; Liu, B.; Wang, F.; Dong, F.; Zhu, Y. Improvement of deficits by transplantation of lentiviral vector-modified human amniotic mesenchymal cells after cerebral ischemia in rats. Brain Res. 2012, 1448, 1–10. [Google Scholar] [CrossRef]
- Zhou, H.; Zhang, H.; Yan, Z.; Xu, R. Transplantation of human amniotic mesenchymal stem cells promotes neurological recovery in an intracerebral hemorrhage rat model. Biochem. Biophys. Res. Commun. 2016, 475, 202–208. [Google Scholar] [CrossRef]
- Shu, J.; He, X.; Li, H.; Liu, X.; Qiu, X.; Zhou, T.; Wang, P.; Huang, X. The beneficial effect of human amnion mesenchymal cells in inhibition of inflammation and induction of neuronal repair in EAE mice. J. Immunol. Res. 2018, 2018, 5083797. [Google Scholar] [CrossRef] [Green Version]
- Yawno, T.; Sabaretnam, T.; Li, J.; McDonald, C.; Lim, R.; Jenkin, G.; Wallace, E.M.; Miller, S.L. Human amnion epithelial cells protect against white matter brain injury after repeated endotoxin exposure in the preterm ovine fetus. Cell Transplant. 2017, 26, 541–553. [Google Scholar] [CrossRef]
- Van den Heuij, L.G.; Fraser, M.; Miller, S.L.; Jenkin, G.; Wallace, E.M.; Davidson, J.O.; Lear, C.A.; Lim, R.; Wassink, G.; Gunn, A.J.; et al. Delayed intranasal infusion of human amnion epithelial cells improves white matter maturation after asphyxia in preterm fetal sheep. J. Cereb. Blood Flow Metab. 2019, 39, 223–239. [Google Scholar] [CrossRef] [PubMed]
- Leaw, B.; Zhu, D.; Tan, J.; Muljadi, R.; Saad, M.I.; Mockler, J.C.; Wallace, E.M.; Lim, R.; Tolcos, M. Human amnion epithelial cells rescue cell death via immunomodulation of microglia in a mouse model of perinatal brain injury. Stem Cell Res. Ther. 2017, 8, 46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pischiutta, F.; Brunelli, L.; Romele, P.; Silini, A.; Sammali, E.; Paracchini, L.; Marchini, S.; Talamini, L.; Bigini, P.; Boncoraglio, G.B.; et al. Protection of brain injury by amniotic mesenchymal stromal cell-secreted metabolites. Crit. Care Med. 2016, 44, e1118–e1131. [Google Scholar] [CrossRef] [PubMed]
- Cui, P.; Xin, H.; Yao, Y.; Xiao, S.; Zhu, F.; Gong, Z.; Tang, Z.; Zhan, Q.; Qin, W.; Lai, Y.; et al. Human amnion-derived mesenchymal stem cells alleviate lung injury induced by white smoke inhalation in rats. Stem Cell Res. Ther. 2018, 9, 101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.Y.; Burgess, J.K.; Wang, Y.; Kable, E.P.W.; Weiss, D.J.; Chan, H.-K.; Chrzanowski, W. Atomized human amniotic mesenchymal stromal cells for direct delivery to the airway for treatment of lung injury. J. Aerosol Med. Pulm. Drug Deliv. 2016, 29, 514–524. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.L.; Tan, Y.Z.; Muljadi, R.; Chan, S.T.; Lau, S.N.; Mockler, J.C.; Wallace, E.M.; Lim, R. Amnion epithelial cells promote lung repair via lipoxin A4. Stem Cells Transl. Med. 2017, 6, 1085–1095. [Google Scholar] [CrossRef]
- Lim, R.; Malhotra, A.; Tan, J.; Chan, S.T.; Lau, S.; Zhu, D.; Mockler, J.C.; Wallace, E.M. First-in-human administration of allogeneic amnion cells in premature infants with bronchopulmonary dysplasia: A safety study. Stem Cells Transl. Med. 2018, 7, 628–635. [Google Scholar] [CrossRef] [Green Version]
- Malhotra, A.; Lim, R.; Mockler, J.C.; Wallace, E.M. Two-year outcomes of infants enrolled in the first-in-human study of amnion cells for bronchopulmonary dysplasia. Stem Cells Transl. Med. 2020, 9, 289–294. [Google Scholar] [CrossRef] [Green Version]
- Zhu, D.; Tan, J.; Maleken, A.S.; Muljadi, R.; Chan, S.T.; Lau, S.N.; Elgass, K.; Leaw, B.; Mockler, J.; Chambers, D.; et al. Human amnion cells reverse acute and chronic pulmonary damage in experimental neonatal lung injury. Stem Cell Res. Ther. 2017, 8, 257. [Google Scholar] [CrossRef] [Green Version]
- Zhu, D.; Kusuma, G.; Schwab, R.; Chan, S.; Tan, J.; Saad, M.I.; Leeman, K.; Kim, C.; Wallace, E.; Lim, R. Prematurity negatively impacts regenerative properties of human amnion epithelial cells in the context of lung repair. Clin. Sci. 2020, 134, 2665–2679. [Google Scholar] [CrossRef]
- Li, Z.; Gong, X.; Li, D.; Yang, X.; Shi, Q.; Ju, X. Intratracheal transplantation of amnion-derived mesenchymal stem cells ameliorates hyperoxia-induced neonatal hyperoxic lung injury via aminoacyl-peptide hydrolase. Int. J. Stem Cells 2020, 13, 221–236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farrow, N.; Cmielewski, P.; Donnelley, M.; Rout-Pitt, N.; Moodley, Y.; Bertoncello, I.; Parsons, D. Epithelial disruption: A new paradigm enabling human airway stem cell transplantation. Stem Cell Res. Ther. 2018, 9, 153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moodley, Y.; Ilancheran, S.; Samuel, C.; Vaghjiani, V.; Atienza, D.; Williams, E.D.; Jenkin, G.; Wallace, E.; Trounson, A.; Manuelpillai, U. Human amnion epithelial cell transplantation abrogates lung fibrosis and augments repair. Am. J. Respir. Crit. Care Med. 2010, 182, 643–651. [Google Scholar] [CrossRef]
- Carbone, A.; Castellani, S.; Favia, M.; Diana, A.; Paracchini, V.; di Gioia, S.; Seia, M.; Casavola, V.; Colombo, C.; Conese, M. Correction of defective CFTR/ENaC function and tightness of cystic fibrosis airway epithelium by amniotic mesenchymal stromal (stem) cells. J. Cell. Mol. Med. 2014, 18, 1631–1643. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.L.; Lau, S.N.; Leaw, B.; Nguyen, H.P.T.; Salamonsen, L.A.; Saad, M.I.; Chan, S.T.; Zhu, D.; Krause, M.; Kim, C.; et al. Amnion epithelial cell-derived exosomes restrict lung injury and enhance endogenous lung repair. Stem Cells Transl. Med. 2018, 7, 180–196. [Google Scholar] [CrossRef] [Green Version]
- Royce, S.G.; Patel, K.P.; Mao, W.; Zhu, D.; Lim, R.; Samuel, C.S. Serelaxin enhances the therapeutic effects of human amnion epithelial cell-derived exosomes in experimental models of lung disease. Br. J. Pharmacol. 2019, 176, 2195–2208. [Google Scholar] [CrossRef]
- Zhang, S.; Jiang, W.; Ma, L.; Liu, Y.; Zhang, X.; Wang, S. Nrf2 transfection enhances the efficacy of human amniotic mesenchymal stem cells to repair lung injury induced by lipopolysaccharide. J. Cell. Biochem. 2017, 119, 1627–1636. [Google Scholar] [CrossRef]
- Cargnoni, A.; Romele, P.; Signoroni, P.B.; Farigu, S.; Magatti, M.; Vertua, E.; Toschi, I.; Cesari, V.; Silini, A.R.; Stefani, F.R.; et al. Amniotic MSCs reduce pulmonary fibrosis by hampering lung B-cell recruitment, retention, and maturation. Stem Cells Transl. Med. 2020, 9, 1023–1035. [Google Scholar] [CrossRef]
- Vosdoganes, P.; Hodges, R.J.; Lim, R.; Westover, A.J.; Acharya, R.Y.; Wallace, E.M.; Moss, T.J.M. Human amnion epithelial cells as a treatment for inflammation-induced fetal lung injury in sheep. Am. J. Obstet. Gynecol. 2011, 205, 156.e26–156.e33. [Google Scholar] [CrossRef]
- Geng, L.; Chen, Z.; Ren, H.; Niu, X.; Yu, X.; Yan, H. Effects of an early intervention using human amniotic epithelial cells in a COPD rat model. Pathol. Res. Pract. 2016, 212, 1027–1033. [Google Scholar] [CrossRef] [Green Version]
- Hua, D.; Ju, Z.; Gan, X.; Wang, Q.; Luo, C.; Gu, J.; Yu, Y. Human amniotic mesenchymal stromal cells alleviate acute liver injury by inhibiting the pro-inflammatory response of liver resident macrophage through autophagy. Ann. Transl. Med. 2019, 7, 392. [Google Scholar] [CrossRef] [PubMed]
- Kuk, N.; Hodge, A.; Sun, Y.; Correia, J.; Alhomrani, M.; Samuel, C.; Moore, G.; Lim, R.; Sievert, W. Human amnion epithelial cells and their soluble factors reduce liver fibrosis in murine non-alcoholic steatohepatitis. J. Gastroenterol. Hepatol. 2019, 34, 1441–1449. [Google Scholar] [CrossRef] [PubMed]
- Coronado, R.E.; Somaraki-Cormier, M.; Ong, J.L.; Halff, G.A. Hepatocyte-like cells derived from human amniotic epithelial, bone marrow, and adipose stromal cells display enhanced functionality when cultured on decellularized liver substrate. Stem Cell Res. 2019, 38, 101471. [Google Scholar] [CrossRef] [PubMed]
- Vaghjiani, V.; Vaithilingam, V.; Saraswati, I.; Sali, A.; Murthi, P.; Kalionis, B.; Tuch, B.E.; Manuelpillai, U. Hepatocyte-like cells derived from human amniotic epithelial cells can be encapsulated without loss of viability or function in vitro. Stem Cells Dev. 2014, 23, 866–876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marongiu, F.; Gramignoli, R.; Dorko, K.; Miki, T.; Ranade, A.R.; Serra, M.P.; Doratiotto, S.; Sini, M.; Sharma, S.; Mitamura, K.; et al. Hepatic differentiation of amniotic epithelial cells. Hepatology 2011, 53, 1719–1729. [Google Scholar] [CrossRef] [Green Version]
- Tabatabaei, M.; Mosaffa, N.; Ghods, R.; Nikoo, S.; Kazemnejad, S.; Khanmohammadi, M.; Mirzadegan, E.; Mahmoudi, A.R.; Bolouri, M.R.; Falak, R.; et al. Vaccination with human amniotic epithelial cells confer effective protection in a murine model of Colon adenocarcinoma. Int. J. Cancer 2018, 142, 1453–1466. [Google Scholar] [CrossRef] [Green Version]
- Hodge, A.; Lourensz, D.; Vaghjiani, V.; Nguyen, H.; Tchongue, J.; Wang, B.; Murthi, P.; Sievert, W.; Manuelpillai, U. Soluble factors derived from human amniotic epithelial cells suppress collagen production in human hepatic stellate cells. Cytotherapy 2014, 16, 1132–1144. [Google Scholar] [CrossRef] [PubMed]
- Manuelpillai, U.; Tchongue, J.; Lourensz, D.; Vaghjiani, V.; Samuel, C.S.; Liu, A.; Williams, E.D.; Sievert, W. Transplantation of human amnion epithelial cells reduces hepatic fibrosis in immunocompetent CCl₄-treated mice. Cell Transplant. 2010, 19, 1157–1168. [Google Scholar] [CrossRef] [Green Version]
- Kubo, K.; Ohnishi, S.; Hosono, H.; Fukai, M.; Kameya, A.; Higashi, R.; Yamada, T.; Onishi, R.; Yamahara, K.; Takeda, H.; et al. Human amnion-derived mesenchymal stem cell transplantation ameliorates liver fibrosis in rats. Transplant. Direct 2015, 1, e16. [Google Scholar] [CrossRef]
- Fu, Q.; Ohnishi, S.; Sakamoto, N. Conditioned medium from human amnion-derived mesenchymal stem cells regulates activation of primary hepatic stellate cells. Stem Cells Int. 2018, 2018, 4898152. [Google Scholar] [CrossRef]
- Lee, P.-H.; Tu, C.-T.; Hsiao, C.-C.; Tsai, M.-S.; Ho, C.-M.; Cheng, N.-C.; Hung, T.-M.; Shih, D.T.-B. Antifibrotic activity of human placental amnion membrane-derived CD34+ mesenchymal stem/progenitor cell transplantation in mice with thioacetamide-induced liver injury. Stem Cells Transl. Med. 2016, 5, 1473–1484. [Google Scholar] [CrossRef] [Green Version]
- Lim, R.; Hodge, A.; Moore, G.; Wallace, E.M.; Sievert, W. A pilot study evaluating the safety of intravenously administered human amnion epithelial cells for the treatment of hepatic fibrosis. Front. Pharmacol. 2017, 8, 549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, D.; Jiang, M.; Miao, D. Transplanted human amniotic membrane-derived mesenchymal stem cells ameliorate carbon tetrachloride-induced liver cirrhosis in mouse. PLoS ONE 2011, 6, e16789. [Google Scholar] [CrossRef] [PubMed]
- Pietrosi, G.; Fernández-Iglesias, A.; Pampalone, M.; Ortega-Ribera, M.; Lozano, J.J.; García-Calderó, H.; Abad-Jordà, L.; Conaldi, P.G.; Parolini, O.; Vizzini, G.; et al. Human amniotic stem cells improve hepatic microvascular dysfunction and portal hypertension in cirrhotic rats. Liver Int. 2020, 40, 2500–2514. [Google Scholar] [CrossRef]
- Lin, J.S.; Zhou, L.; Sagayaraj, A.; Jumat, N.H.; Choolani, M.; Chan, J.K.; Biswas, A.; Wong, P.C.; Lim, S.G.; Dan, Y.Y. Hepatic differentiation of human amniotic epithelial cells and in vivo therapeutic effect on animal model of cirrhosis. J. Gastroenterol. Hepatol. 2015, 30, 1673–1682. [Google Scholar] [CrossRef] [PubMed]
- Skvorak, K.J.; Dorko, K.; Marongiu, F.; Tahan, V.; Hansel, M.C.; Gramignoli, R.; Arning, E.; Bottiglieri, T.; Gibson, K.M.; Strom, S.C. Improved amino acid, bioenergetic metabolite and neurotransmitter profiles following human amnion epithelial cell transplant in intermediate maple syrup urine disease mice. Mol. Genet. Metab. 2013, 109, 132–138. [Google Scholar] [CrossRef]
- Skvorak, K.J.; Dorko, K.; Marongiu, F.; Tahan, V.; Hansel, M.C.; Gramignoli, R.; Gibson, K.M.; Strom, S.C. Placental stem cell correction of murine intermediate maple syrup urine disease. Hepatology 2013, 57, 1017–1023. [Google Scholar] [CrossRef] [Green Version]
- Sagar, J.; Chaib, B.; Sales, K.; Winslet, M.; Seifalian, A. Role of stem cells in cancer therapy and cancer stem cells: A review. Cancer Cell Int. 2007, 7, 9. [Google Scholar] [CrossRef] [Green Version]
- Qiao, L.; Xu, Z.; Zhao, T.; Zhao, Z.; Shi, M.; Zhao, R.C.; Ye, L.; Zhang, X. Suppression of tumorigenesis by human mesenchymal stem cells in a hepatoma model. Cell Res. 2008, 18, 500–507. [Google Scholar] [CrossRef] [Green Version]
- Magatti, M.; De Munari, S.; Vertua, E.; Parolini, O. Amniotic membrane-derived cells inhibit proliferation of cancer cell lines by inducing cell cycle arrest. J. Cell. Mol. Med. 2012, 16, 2208–2218. [Google Scholar] [CrossRef]
- Rolfo, A.; Giuffrida, D.; Giuffrida, M.C.; Todros, T.; Calogero, A.E. New perspectives for prostate cancer treatment: In vitro inhibition of LNCaP and PC3 cell proliferation by amnion-derived mesenchymal stromal cells conditioned media. Aging Male 2014, 17, 94–101. [Google Scholar] [CrossRef] [PubMed]
- Jiao, H.; Guan, F.; Yang, B.; Li, J.; Song, L.; Hu, X.; Du, Y. Human amniotic membrane derived-mesenchymal stem cells induce C6 glioma apoptosis in vivo through the Bcl-2/caspase pathways. Mol. Biol. Rep. 2012, 39, 467–473. [Google Scholar] [CrossRef]
- Dhiman, N.; Shagaghi, N.; Bhave, M.; Sumer, H.; Kingshott, P.; Rath, S.N. Indirect co-culture of lung carcinoma cells with hyperthermia-treated mesenchymal stem cells influences tumor spheroid growth in a collagen-based 3-dimensional microfluidic model. Cytotherapy 2020, 23, 25–36. [Google Scholar] [CrossRef]
- Bonomi, A.; Silini, A.; Vertua, E.; Signoroni, P.B.; Coccè, V.; Cavicchini, L.; Sisto, F.; Alessandri, G.; Pessina, A.; Parolini, O. Human amniotic mesenchymal stromal cells (hAMSCs) as potential vehicles for drug delivery in cancer therapy: An in vitro study. Stem Cell Res. Ther. 2015, 6, 155. [Google Scholar] [CrossRef] [Green Version]
- Niknejad, H.; Khayat-Khoei, M.; Peirovi, H.; Abolghasemi, H. Human amniotic epithelial cells induce apoptosis of cancer cells: A new anti-tumor therapeutic strategy. Cytotherapy 2014, 16, 33–40. [Google Scholar] [CrossRef]
- Di Germanio, C.; Bernier, M.; Petr, M.; Mattioli, M.; Barboni, B.; de Cabo, R. Conditioned medium derived from rat amniotic epithelial cells confers protection against inflammation, cancer, and senescence. Oncotarget 2016, 7, 39051–39064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bu, S.; Zhang, Q.; Wang, Q.; Lai, D. Human amniotic epithelial cells inhibit growth of epithelial ovarian cancer cells via TGFbeta1-mediated cell cycle arrest. Int. J. Oncol. 2017, 51, 1405–1414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.W.; Zhang, H.Z.; Guo, L.; Kim, J.M.; Kim, M.H. Amniotic mesenchymal stem cells enhance wound healing in diabetic NOD/SCID mice through high angiogenic and engraftment capabilities. PLoS ONE 2012, 7, e41105. [Google Scholar] [CrossRef]
- Gao, S.; Chen, T.; Hao, Y.; Zhang, F.; Tang, X.; Wang, D.; Wei, Z.; Qi, J. Exosomal miR-135a derived from human amnion mesenchymal stem cells promotes cutaneous wound healing in rats and fibroblast migration by directly inhibiting LATS2 expression. Stem Cell Res. Ther. 2020, 11, 56. [Google Scholar] [CrossRef]
- Tuca, A.-C.; Ertl, J.; Hingerl, K.; Pichlsberger, M.; Fuchs, J.; Wurzer, P.; Pfeiffer, D.; Bubalo, V.; Parvizi, D.; Kamolz, L.-P.; et al. Comparison of Matrigel and Matriderm as a carrier for human amnion-derived mesenchymal stem cells in wound healing. Placenta 2016, 48, 99–103. [Google Scholar] [CrossRef]
- Zhao, B.; Liu, J.Q.; Zheng, Z.; Zhang, J.; Wang, S.Y.; Han, S.C.; Zhou, Q.; Guan, H.; Li, C.; Su, L.L.; et al. Human amniotic epithelial stem cells promote wound healing by facilitating migration and proliferation of keratinocytes via ERK, JNK and AKT signaling pathways. Cell Tissue Res. 2016, 365, 85–99. [Google Scholar] [CrossRef]
- Jin, E.; Kim, T.-H.; Han, S.; Kim, S.-W. Amniotic epithelial cells promote wound healing in mice through high epithelialization and engraftment. J. Tissue Eng. Regen. Med. 2016, 10, 613–622. [Google Scholar] [CrossRef] [PubMed]
- Zhou, A.; Zheng, X.; Yu, L.; Quan, M.; Shao, X.; Jiang, Z. Mechanisms of human amniotic epithelial cell transplantation in treating stage III pressure ulcer in a rat model. Exp. Ther. Med. 2015, 10, 2161–2168. [Google Scholar] [CrossRef] [Green Version]
- Zhao, B.; Zhang, Y.; Han, S.; Zhang, W.; Zhou, Q.; Guan, H.; Liu, J.; Shi, J.; Su, L.; Hu, D. Exosomes derived from human amniotic epithelial cells accelerate wound healing and inhibit scar formation. J. Mol. Histol. 2017, 48, 121–132. [Google Scholar] [CrossRef] [PubMed]
- Fatimah, S.S.; Chua, K.; Tan, G.C.; Azmi, T.I.; Tan, A.E.; Abdul Rahman, H. Organotypic culture of human amnion cells in air-liquid interface as a potential substitute for skin regeneration. Cytotherapy 2013, 15, 1030–1041. [Google Scholar] [CrossRef]
- Jiang, L.W.; Chen, H.; Lu, H. Using human epithelial amnion cells in human de-epidermized dermis for skin regeneration. J. Dermatol. Sci. 2016, 81, 26–34. [Google Scholar] [CrossRef]
- Yu, S.C.; Xu, Y.Y.; Li, Y.; Xu, B.; Sun, Q.; Li, F.; Zhang, X.G. Construction of tissue engineered skin with human amniotic mesenchymal stem cells and human amniotic epithelial cells. Eur. Rev. Med. Pharmacol. Sci. 2015, 19, 4627–4635. [Google Scholar]
- Li, H.; Chu, Y.; Zhang, Z.; Zhang, G.; Jiang, L.; Wu, H.; Liu, S.; Yu, C.; Jin, Y. Construction of bilayered tissue-engineered skin with human amniotic mesenchymal cells and human amniotic epithelial cells. Artif. Organs 2012, 36, 911–919. [Google Scholar] [CrossRef]
- He, D.; Zhao, F.; Jiang, H.; Kang, Y.; Song, Y.; Lin, X.; Shi, P.; Zhang, T.; Pang, X. LOXL2 from human amniotic mesenchymal stem cells accelerates wound epithelialization by promoting differentiation and migration of keratinocytes. Aging 2020, 12, 12960–12986. [Google Scholar] [CrossRef]
- Sato, C.; Yamamoto, Y.; Funayama, E.; Furukawa, H.; Oyama, A.; Murao, N.; Hosono, H.; Kawakubo, K.; Sakamoto, N.; Ohnishi, S. Conditioned medium obtained from amnion-derived mesenchymal stem cell culture prevents activation of keloid fibroblasts. Plast. Reconstr. Surg. 2018, 141, 390–398. [Google Scholar] [CrossRef]
- Zhao, B.; Liu, J.Q.; Yang, C.; Zheng, Z.; Zhou, Q.; Guan, H.; Su, L.L.; Hu, D.H. Human amniotic epithelial cells attenuate TGF-beta1-induced human dermal fibroblast transformation to myofibroblasts via TGF-beta1/Smad3 pathway. Cytotherapy 2016, 18, 1012–1024. [Google Scholar] [CrossRef] [PubMed]
- Pan, C.; Lang, H.; Zhang, T.; Wang, R.; Lin, X.; Shi, P.; Zhao, F.; Pang, X. Conditioned medium derived from human amniotic stem cells delays H2O2-induced premature senescence in human dermal fibroblasts. Int. J. Mol. Med. 2019, 44, 1629–1640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, C.; Yuchi, H.; Sun, L.; Zhou, X.; Lin, J. Human amnion-derived mesenchymal stem cells protect against UVA irradiation-induced human dermal fibroblast senescence, in vitro. Mol. Med. Rep. 2017, 16, 2016–2022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imai, Y.; Yamahara, K.; Hamada, A.; Fujimori, Y.; Yamanishi, K. Human amnion-derived mesenchymal stem cells ameliorate imiquimod-induced psoriasiform dermatitis in mice. J. Dermatol. 2019, 46, 276–278. [Google Scholar] [CrossRef]
- Shu, J.; Pan, L.; Huang, X.; Wang, P.; Li, H.; He, X.; Cai, Z. Transplantation of human amnion mesenchymal cells attenuates the disease development in rats with collagen-induced arthritis. Clin. Exp. Rheumatol. 2015, 33, 484–490. [Google Scholar]
- Topoluk, N.; Steckbeck, K.; Siatkowski, S.; Burnikel, B.; Tokish, J.; Mercuri, J. Amniotic mesenchymal stem cells mitigate osteoarthritis progression in a synovial macrophage-mediated in vitro explant coculture model. J. Tissue Eng. Regen. Med. 2018, 12, 1097–1110. [Google Scholar] [CrossRef]
- Bian, Y.; Ma, X.; Wang, R.; Yuan, H.; Chen, N.; Du, Y. Human amnion-derived mesenchymal stem cells promote osteogenesis of human bone marrow mesenchymal stem cells against glucolipotoxicity. FEBS Open Bio 2019, 9, 74–81. [Google Scholar] [CrossRef] [Green Version]
- Yin, L.; Zhou, Z.-X.; Shen, M.; Chen, N.; Jiang, F.; Wang, S.-L. The human amniotic mesenchymal stem cells (hAMSCs) improve the implant osseointegration and bone regeneration in maxillary sinus floor elevation in rabbits. Stem Cells Int. 2019, 2019, 9845497. [Google Scholar] [CrossRef] [Green Version]
- Jiang, F.; Zhang, W.; Zhou, M.; Zhou, Z.; Shen, M.; Chen, N.; Jiang, X. Human amniotic mesenchymal stromal cells promote bone regeneration via activating endogenous regeneration. Theranostics 2020, 10, 6216–6230. [Google Scholar] [CrossRef]
- Wang, G.; Zhao, F.; Yang, D.; Wang, J.; Qiu, L.; Pang, X. Human amniotic epithelial cells regulate osteoblast differentiation through the secretion of TGFβ1 and microRNA-34a-5p. Int. J. Mol. Med. 2018, 41, 791–799. [Google Scholar] [CrossRef]
- Zhang, Z.; Zeng, L.; Yang, J.; Guo, L.; Hou, Q.; Zhu, F. Amniotic membrane-derived stem cells help repair osteochondral defect in a weight-bearing area in rabbits. Exp. Ther. Med. 2017, 14, 187–192. [Google Scholar] [CrossRef] [Green Version]
- Si, J.; Dai, J.; Zhang, J.; Liu, S.; Gu, J.; Shi, J.; Shen, S.G.F.; Guo, L. Comparative investigation of human amniotic epithelial cells and mesenchymal stem cells for application in bone tissue engineering. Stem Cells Int. 2015, 2015, 565732. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Liu, Z.; Tang, Y.; Fan, Q.; Feng, W.; Luo, C.; Dai, G.; Ge, Z.; Zhang, J.; Zou, G.; et al. Three-dimensional silk fibroin scaffolds enhance the bone formation and angiogenic differentiation of human amniotic mesenchymal stem cells: A biocompatibility analysis. Acta Biochim. Biophys. Sin. 2020, 52, 590–602. [Google Scholar] [CrossRef]
- Muiños-López, E.; Hermida-Gómez, T.; Fuentes-Boquete, I.; de Toro-Santos, J.; Blanco, F.J.; Díaz-Prado, S.M. Human amniotic mesenchymal stromal cells as favorable source for cartilage repair. Tissue Eng. Part A 2017, 23, 901–912. [Google Scholar] [CrossRef]
- Zhang, J.; Liu, Z.; Li, Y.; You, Q.; Yang, J.; Jin, Y.; Zou, G.; Tang, J.; Ge, Z.; Liu, Y. FGF-2-induced human amniotic mesenchymal stem cells seeded on a human acellular amniotic membrane scaffold accelerated tendon-to-bone healing in a rabbit extra-articular model. Stem Cells Int. 2020, 2020, 4701476. [Google Scholar] [CrossRef] [Green Version]
- Wei, X.; Sun, G.; Zhao, X.; Wu, Q.; Chen, L.; Xu, Y.; Pang, X.; Qi, G. Human amnion mesenchymal stem cells attenuate atherosclerosis by modulating macrophage function to reduce immune response. Int. J. Mol. Med. 2019, 44, 1425–1435. [Google Scholar] [CrossRef]
- Kim, S.-W.; Zhang, H.-Z.; Kim, C.-E.; Kim, J.-M.; Kim, M.H. Amniotic mesenchymal stem cells with robust chemotactic properties are effective in the treatment of a myocardial infarction model. Int. J. Cardiol. 2013, 168, 1062–1069. [Google Scholar] [CrossRef]
- Kim, P.J.; Mahmoudi, M.; Ge, X.; Matsuura, Y.; Toma, I.; Metzler, S.; Kooreman, N.G.; Ramunas, J.; Holbrook, C.; McConnell, M.V.; et al. Direct evaluation of myocardial viability and stem cell engraftment demonstrates salvage of the injured myocardium. Circ. Res. 2015, 116, e40–e50. [Google Scholar] [CrossRef] [Green Version]
- Tsuji, H.; Miyoshi, S.; Ikegami, Y.; Hida, N.; Asada, H.; Togashi, I.; Suzuki, J.; Satake, M.; Nakamizo, H.; Tanaka, M.; et al. Xenografted human amniotic membrane-derived mesenchymal stem cells are immunologically tolerated and transdifferentiated into cardiomyocytes. Circ. Res. 2010, 106, 1613–1623. [Google Scholar] [CrossRef] [Green Version]
- Takov, K.; He, Z.; Johnston, H.E.; Timms, J.F.; Guillot, P.V.; Yellon, D.M.; Davidson, S.M. Small extracellular vesicles secreted from human amniotic fluid mesenchymal stromal cells possess cardioprotective and promigratory potential. Basic Res. Cardiol. 2020, 115, 26. [Google Scholar] [CrossRef]
- Fang, C.-H.; Jin, J.; Joe, J.-H.; Song, Y.-S.; So, B.-I.; Lim, S.M.; Cheon, G.J.; Woo, S.-K.; Ra, J.-C.; Lee, Y.-Y.; et al. In vivo differentiation of human amniotic epithelial cells into cardiomyocyte-like cells and cell transplantation effect on myocardial infarction in rats: Comparison with cord blood and adipose tissue-derived mesenchymal stem cells. Cell Transplant. 2012, 21, 1687–1696. [Google Scholar] [CrossRef] [PubMed]
- Naseroleslami, M.; Aboutaleb, N.; Mokhtari, B. Amniotic membrane mesenchymal stem cells labeled by iron oxide nanoparticles exert cardioprotective effects against isoproterenol (ISO)-induced myocardial damage by targeting inflammatory MAPK/NF-κB pathway. Drug Deliv. Transl Res. 2020, 11, 242–254. [Google Scholar] [CrossRef] [PubMed]
- Razavi Tousi, S.M.T.; Faghihi, M.; Nobakht, M.; Molazem, M.; Kalantari, E.; Darbandi Azar, A.; Aboutaleb, N. Improvement of heart failure by human amniotic mesenchymal stromal cell transplantation in rats. J. Tehran Heart Cent. 2016, 11, 123–138. [Google Scholar]
- Nasseri Maleki, S.; Aboutaleb, N.; Nazarinia, D.; Allahverdi Beik, S.; Qolamian, A.; Nobakht, M. Conditioned medium obtained from human amniotic membrane-derived mesenchymal stem cell attenuates heart failure injury in rats. Iran. J. Basic Med. Sci. 2019, 22, 1253–1258. [Google Scholar]
- Miyamoto, S.; Ohnishi, S.; Onishi, R.; Tsuchiya, I.; Hosono, H.; Katsurada, T.; Yamahara, K.; Takeda, H.; Sakamoto, N. Therapeutic effects of human amnion-derived mesenchymal stem cell transplantation and conditioned medium enema in rats with trinitrobenzene sulfonic acid-induced colitis. Am. J. Transl. Res. 2017, 9, 940–952. [Google Scholar]
- Legaki, E.; Roubelakis, M.G.; Theodoropoulos, G.E.; Lazaris, A.; Kollia, A.; Karamanolis, G.; Marinos, E.; Gazouli, M. Therapeutic potential of secreted molecules derived from human amniotic fluid mesenchymal stem/stroma cells in a mice model of colitis. Stem Cell Rev. Rep. 2016, 12, 604–612. [Google Scholar] [CrossRef]
- Onishi, R.; Ohnishi, S.; Higashi, R.; Watari, M.; Yamahara, K.; Okubo, N.; Nakagawa, K.; Katsurada, T.; Suda, G.; Natsuizaka, M.; et al. Human amnion-derived mesenchymal stem cell transplantation ameliorates dextran sulfate sodium-induced severe colitis in rats. Cell Transplant. 2015, 24, 2601–2614. [Google Scholar] [CrossRef] [Green Version]
- Ono, M.; Ohnishi, S.; Honda, M.; Ishikawa, M.; Hosono, H.; Onishi, R.; Nakagawa, K.; Takeda, H.; Sakamoto, N. Effects of human amnion-derived mesenchymal stromal cell transplantation in rats with radiation proctitis. Cytotherapy 2015, 17, 1545–1559. [Google Scholar] [CrossRef] [Green Version]
- Otagiri, S.; Ohnishi, S.; Miura, A.; Hayashi, H.; Kumagai, I.; Ito, Y.M.; Katsurada, T.; Nakamura, S.; Okamoto, R.; Yamahara, K.; et al. Evaluation of amnion-derived mesenchymal stem cells for treatment-resistant moderate Crohn’s disease: Study protocol for a phase I/II, dual-centre, open-label, uncontrolled, dose-response trial. BMJ Open Gastroenterol. 2018, 5, e000206. [Google Scholar] [CrossRef] [PubMed]
- Cetinkaya, B.; Unek, G.; Kipmen-Korgun, D.; Koksoy, S.; Korgun, E.T. Effects of human placental amnion derived mesenchymal stem cells on proliferation and apoptosis mechanisms in chronic kidney disease in the rat. Int. J. Stem Cells 2019, 12, 151–161. [Google Scholar] [CrossRef] [PubMed]
- Ren, Y.; Chen, Y.; Zheng, X.; Wang, H.; Kang, X.; Tang, J.; Qu, L.; Shao, X.; Wang, S.; Li, S.; et al. Human amniotic epithelial cells ameliorate kidney damage in ischemia-reperfusion mouse model of acute kidney injury. Stem Cell Res. Ther. 2020, 11, 410. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Hua, R.; Gong, Z.; Shang, B.; Huang, Y.; Guo, L.; Liu, T.; Xue, J. Human amniotic epithelial cells inhibit CD4+ T cell activation in acute kidney injury patients by influencing the miR-101-c-Rel-IL-2 pathway. Mol. Immunol. 2017, 81, 76–84. [Google Scholar] [CrossRef] [PubMed]
- Fijany, A.; Sayadi, L.R.; Khoshab, N.; Banyard, D.A.; Shaterian, A.; Alexander, M.; Lakey, J.R.; Paydar, K.Z.; Evans, G.R.; Widgerow, A.D. Mesenchymal stem cell dysfunction in diabetes. Mol. Biol. Rep. 2019, 46, 1459–1475. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, A.M. Highlighting diabetes mellitus. Arterioscler. Thromb. Vasc. Biol. 2018, 38, e1–e8. [Google Scholar] [CrossRef] [Green Version]
- Luo, Y.; Cheng, Y.-W.; Yu, C.-Y.; Liu, R.-M.; Zhao, Y.-J.; Chen, D.-X.; Zhong, J.-J.; Xiao, J.-H. Effects of hyaluronic acid on differentiation of human amniotic epithelial cells and cell-replacement therapy in type 1 diabetic mice. Exp. Cell Res. 2019, 384, 111642. [Google Scholar] [CrossRef]
- Zou, G.; Liu, T.; Guo, L.; Huang, Y.; Feng, Y.; Duan, T. MicroRNA-32 silences WWP2 expression to maintain the pluripotency of human amniotic epithelial stem cells and β islet-like cell differentiation. Int. J. Mol. Med. 2018, 41, 1983–1991. [Google Scholar] [CrossRef]
- Okere, B.; Alviano, F.; Costa, R.; Quaglino, D.; Ricci, F.; Dominici, M.; Paolucci, P.; Bonsi, L.; Iughetti, L. In vitro differentiation of human amniotic epithelial cells into insulin-producing 3D spheroids. Int. J. Immunopathol. Pharmacol. 2015, 28, 390–402. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.; Park, S.; Kang, H.M.; Ahn, C.W.; Kwon, H.C.; Song, J.H.; Lee, Y.J.; Lee, K.H.; Yang, H.; Baek, S.Y.; et al. Human insulin secreted from insulinogenic xenograft restores normoglycemia in type 1 diabetic mice without immunosuppression. Cell Transplant. 2012, 21, 2131–2147. [Google Scholar] [CrossRef] [Green Version]
- Lebreton, F.; Lavallard, V.; Bellofatto, K.; Bonnet, R.; Wassmer, C.H.; Perez, L.; Kalandadze, V.; Follenzi, A.; Boulvain, M.; Kerr-Conte, J.; et al. Insulin-producing organoids engineered from islet and amniotic epithelial cells to treat diabetes. Nat. Commun. 2019, 10, 4491. [Google Scholar] [CrossRef] [Green Version]
- McDonald, C.A.; Payne, N.L.; Sun, G.; Moussa, L.; Siatskas, C.; Lim, R.; Wallace, E.M.; Jenkin, G.; Bernard, C.C.A. Immunosuppressive potential of human amnion epithelial cells in the treatment of experimental autoimmune encephalomyelitis. J. Neuroinflamm. 2015, 12, 112. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.H.; Vaghjiani, V.; Tee, J.Y.; To, K.; Cui, P.; Oh, D.Y.; Manuelpillai, U.; Toh, B.-H.; Chan, J. Amniotic epithelial cells from the human placenta potently suppress a mouse model of multiple sclerosis. PLoS ONE 2012, 7, e35758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Qiu, C.; Zhang, Z.; Yuan, W.; Ge, Z.; Tan, B.; Yang, P.; Liu, J.; Zhu, X.; Qiu, C.; et al. Subretinal transplantation of human amniotic epithelial cells in the treatment of autoimmune uveitis in rats. Cell Transplant. 2018, 27, 1504–1514. [Google Scholar] [CrossRef] [PubMed]
- Tan, B.; Yuan, W.; Li, J.; Yang, P.; Ge, Z.; Liu, J.; Qiu, C.; Zhu, X.; Qiu, C.; Lai, D.; et al. Therapeutic effect of human amniotic epithelial cells in murine models of Hashimoto’s thyroiditis and Systemic lupus erythematosus. Cytotherapy 2018, 20, 1247–1258. [Google Scholar] [CrossRef]
- Pianta, S.; Magatti, M.; Vertua, E.; Bonassi Signoroni, P.; Muradore, I.; Nuzzo, A.M.; Rolfo, A.; Silini, A.; Quaglia, F.; Todros, T.; et al. Amniotic mesenchymal cells from pre-eclamptic placentae maintain immunomodulatory features as healthy controls. J. Cell. Mol. Med. 2016, 20, 157–169. [Google Scholar] [CrossRef] [PubMed]
- Miceli, V.; Pampalone, M.; Vella, S.; Carreca, A.P.; Amico, G.; Conaldi, P.G. Comparison of immunosuppressive and angiogenic properties of human amnion-derived mesenchymal stem cells between 2D and 3D culture systems. Stem Cells Int. 2019, 2019, 7486279. [Google Scholar] [CrossRef]
- Hong, S.-B.; Seo, M.-S.; Park, S.-B.; Seo, Y.-J.; Kim, J.-S.; Kang, K.-S. Therapeutic effects of human amniotic epithelial stem cells in Niemann-Pick type C1 mice. Cytotherapy 2012, 14, 630–638. [Google Scholar] [CrossRef]
- Navas, A.; Magaña-Guerrero, F.S.; Domínguez-López, A.; Chávez-García, C.; Partido, G.; Graue-Hernández, E.O.; Sánchez-García, F.J.; Garfias, Y. Anti-inflammatory and anti-fibrotic effects of human amniotic membrane mesenchymal stem cells and their potential in corneal repair. Stem Cells Transl. Med. 2018, 7, 906–917. [Google Scholar] [CrossRef]
- Lv, X.; Guo, Q.; Han, F.; Chen, C.; Ling, C.; Chen, W.; Li, B. Electrospun poly(l-lactide)/poly(ethylene glycol) scaffolds seeded with human amniotic mesenchymal stem cells for urethral epithelium repair. Int. J. Mol. Sci. 2016, 17, 1262. [Google Scholar] [CrossRef] [Green Version]
- Qian, C.; Meng, Q.; Lu, J.; Zhang, L.; Li, H.; Huang, B. Human amnion mesenchymal stem cells restore spermatogenesis in mice with busulfan-induced testis toxicity by inhibiting apoptosis and oxidative stress. Stem Cell Res. Ther. 2020, 11, 290. [Google Scholar] [CrossRef]
- Baker, E.K.; Malhotra, A.; Lim, R.; Jacobs, S.E.; Hooper, S.B.; Davis, P.G.; Wallace, E.M. Human amnion cells for the prevention of bronchopulmonary dysplasia: A protocol for a phase I dose escalation study. BMJ Open 2019, 9, e026265. [Google Scholar] [CrossRef] [Green Version]
- Phan, T.G.; Ma, H.; Lim, R.; Sobey, C.G.; Wallace, E.M. Phase 1 trial of amnion cell therapy for ischemic stroke. Front. Neurol. 2018, 9, 198. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| hADSCs | Epithelial Markers | Mesenchymal Stem Cell Markers | Pluripotent Markers | Hematopoietic Marker | The Major Histocompatibility Complex and Their Co-Stimulatory Molecules | References | |
|---|---|---|---|---|---|---|---|
| Positive | Positive | Positive | Negative | Positive | Negative | ||
| hAESCs | Cytokeratin, E-cadherin, CD49f, CD326, | CD29, CD166, | OCT4, NANOG, SSEA4, TRA-1-60 | CD34, CD45 | HLA-DR, HLA-DQ | Yang et al. [35] | |
| CK19 | CD29, CD44, CD73, CD90, CD105, | SSEA-4, SOX2, OCT-4 | CD31, CD34, CD45, CD49d | HLA-DR | Wu et al. [38] | ||
| CK7, E-cadherin | CD29, CD73, CD105 | OCT4, NANOG, SSEA4 | CD34, CD45 | HLA-ABC | HLA-DR, CD80, CD86, CD40 | Liu et al. [23] | |
| E-cadherin | OCT4, SOX2, NANOG, TFE3, KLF4, SSEA3, SSEA4, TRA-1-60, REX1 | Castro et al. [39] | |||||
| E-cadherin, CD49f, CK7, EpCAM | CD44, CD90, CD105, CD146, PDGFR-b, CD29 | CD45 | HLA-A-B-C | HLA-DP-DQ-DR, CD80, CD86, CD40 | Pratama et al. [33] | ||
| Cytokeratin | SSEA3, SSEA4, TRA-1-60, Oct-4 | CD34 | Evron et al. [40] | ||||
| CD29, CD73 | CD34, CD45 | HLA-ABC | HLA-A2, HLA-DQ, HLA-DR | Murphy et al. [30] | |||
| CD9, CD10, CD29, CD104, CD49f, CD105, CD44 | CD34, CD45 | HLA-ABC | HLA-DR, CD80, CD86, CD40 | Banas et al. [31] | |||
| SSEA-3, SSEA-4, TRA 1-60, TRA 1-81 | CD34 | Miki et al. [41] | |||||
| Cytokeratin, E-cadherin | CD29, CD166, CD90, | OCT4, NANOG, TRA 1-60, SOX2 | CD34, CD45 | HLA-DR, HLA-DQ | Yang et al. [35] | ||
| CK19, E-cadherin | CD29, CD44, CD90 | OCT4, SOX2, SSEA4, | CD34 | Wu et al. [42] | |||
| E-cadherin, | CD9, CD24, | SSEA-3, SSEA-4, TRA 1-60, TRA 1-81, Oct-4, Nanog | CD34 | Miki et al. [36] | |||
| CD73, CD29, | Oct3/4, Sox2, Klf4, SSEA4, c-Myc. | CD34, CD45 | Koike et al. [43] | ||||
| hAMSCs | CD29, CD44, CD49d, CD73, CD90, CD105 | SSEA-4, SOX2, OCT-4 | CD31, CD34, CD45 | HLA-DR | Wu et al. [38] | ||
| CD29, CD73, CD90, CD105 | OCT4, NANOG, SSEA4 | CD34, CD45, CD133 | HLA-ABC | HLA-DR, CD80, CD86, CD40 | Liu and Li et al. [24,25] | ||
| CD44, CD90, CD105, CD146 | Oct-3/4 | CD45, CD34 | HLA-ABC | HLA-DR | Bačenková et al. [44] | ||
| CD90, CD44, CD73, CD166, CD105, CD29 | SSEA-4, STRO-1 | CD34, CD45 | Prado and Sugiura et al. [45,46] | ||||
| CD29, CD44, CD73, CD90, CD105 | SSEA-4, Oct4 | CD34, CD45, CD133 | HLA-ABC | HLA-DR | Coppi et al. [34] | ||
| CD105, CD117, | CD34 | HLA-ABC | HLA-DR | Borghesi et al. [47] | |||
| CD29, CD105 | Oct-3/4, SSEA-4, SOX2, NANOG, Rex-1 | HLA-A, HLA-DQB1 | Mihu et al. [48] | ||||
| CD44, CD90, CD105. CD90 | CD31, CD34 | Seo et al. [49] | |||||
| CD44, CD73, CD90, CD105, CD29, CD49f, CD271 | Oct3/4, Sox2, Klf4,SSEA4, Nanog, TRA1-60 | CD34, CD45 | Koike et al. [43] | ||||
| CD44, CD73, CD90, CD105, Vimentin | OCT3/4, C-MYC, SOX2, NANOG, SSEA-3, SSEA-4 | CD34, CD45 | HLA-DR | Nogami et al. [50] | |||
| Title | NCT Number | Condition or Disease | Phase | Enrollment | Injection Site | Dose | Outcome Measures | Status | Locations | |
|---|---|---|---|---|---|---|---|---|---|---|
| hAMSCs | Allogeneic Amniotic Mesenchymal Stem Cell Therapy for Lupus Nephritis | NCT04318600 | Lupus Nephritis | Phase 1 | 16 | Peripheral intravenous | 1 × 106/kg | Incidence of Adverse Events; 2. Changes in 24h urine protein quantification before and after treatment; 3. Changes in eGFR before and after treatment; 4. Changes in SLEDAI score before and after treatment. | Completed | China |
| hAESCs | Effect of Human Amniotic Epithelial Cells on Children With Spastic Cerebral Palsy | NCT03107975 | Spastic Cerebral Palsy | Phase 1 | 10 | Gross Motor Function Measure-66; 2. Fine Motor Function Measure; 3. Modified Ashworth Scale; 4. Gesell Developmental Scales. | Unknown | China | ||
| Treatment of Nonunion of Limb Fracture With Human Amniotic Epithelial Cells | NCT03031509 | Nonunion Fracture | Phase 1 Phase 2 | 36 | Nonunion site | 50 million | time for bone union; 2. pain in fracture site; 3. limb functional score | Not yet recruiting | China | |
| Transplantation of Tissue Cultured Human Amniotic Epithelial Cells Onto Damaged Ocular Surfaces | NCT00344708 | Corneal Epithelial Dystrophy | 0 | Ocular surface healing. | Not Applicable | United States | ||||
| Human Amniotic Epithelial Cells for Treatment of Bronchial Fistula | NCT02959333 | Bronchial Fistula | Phase 1 | 10 | 3–5 × 107/5 mL | median time for fistula closure; 2. Complication | Unknown | China | ||
| A Therapeutic Trial of Human Amniotic Epithelial Cells Transplantation for Primary Ovarian Insufficiency Patients | NCT02912104 | Primary Ovarian Insufficiency;Premature Ovarian Failure;Infertility | Phase 1 | 2 | Bilateral ovarian artery infusion | 2 × 107/5 mL | bilateral ovarian artery infusion; 2. Hormonal Assessment serum estrogen level will be measured; 3. Hormonal Assessment serum AMH level will be measured; 4. Pregnancy rate will be measured. | Recruiting | China | |
| Human Amniotic Epithelial Cells Prevent Acute Graft-versus-host Disease After Hematopoietic Stem Cell Transplantation | NCT03764228 | Acute-graft-versus-host Disease | Not Applicable | 27 | 1 × 106, 2 × 106, 5 × 106 cell/kg | Occurrence of adverse events; 2. Occurrence of Graft-versus-host disease after hAECs infusion | Recruiting | China | ||
| Human Amniotic Epithelial Cells Treatment for Ovarian Insufficiency | NCT03207412 | Premature Ovarian Failure | Not Applicable | 20 | bilateral ovarian tissue | 200 million | Changes of follicle stimulating hormone; 2. antral follicle count | Unknown | China | |
| Human Amniotic Epithelial Cells for Asherman’s Syndrome | NCT03223454 | Asherman’s Syndrome | Phase 1 | 50 | uterine cavity | 100 million | Menstrual blood volume; 2. Endometrial thickness; 3. Uterine volume; 4. Ongoing pregnancy rate | Not yet recruiting | China | |
| Human Amniotic Epithelial Cell in Treatment of Refractory Severe Intrauterine Adhesion | NCT03381807 | Intrauterine Adhesion | Early Phase 1 | 20 | uterine cavity | 100 million | changes of Endometrial thickness; 2. changes of Menstrual blood volume; 3. pregnancy rate | Not yet recruiting | China | |
| hAECs Are Preliminarily Applied in Allogeneic Hematopoietic Stem Cell Transplantation | NCT03759899 | Leukemia | 30 | Cell counts of T cells, NK cells, B cells and monocytes. | Recruiting | China | ||||
| Stereotactic Transplantation of hAESCs for Parkinson’s Disease | NCT04414813 | Parkinson’s Disease | Early Phase 1 | 3 | Stereotactic | 50 million | Number of Participants with adverse event and serious adverse event; 2. Changes in Unified Parkinson’s Disease Rating Scale; 3. Changes in the Hoehn and Yahr scales; 4. Changes in Parkinson’s Disease Questionnaire; 5. Changes in the Schwab and England scores. | Not yet recruiting | China |
| Diseases | Biological Functions of hAESCs and hAMSCs | References | |
|---|---|---|---|
| Gynecological diseases | Premature ovarian failure |
| [76,77,78] |
| Intrauterine adhesion | [86,87,88] | ||
| Recurrent spontaneous abortion | [89] | ||
| Nervous system diseases | Spinal Cord Injury |
| [91,92,93,94,95] |
| Alzheimer’s Disease | [97,98,99,100,101] | ||
| Autism spectrum disorder | [24,107] | ||
| Parkinson’s disease | [108] | ||
| Stroke | [109,110,111,112,113,114,115,116,117] | ||
| Intracerebral hemorrhage | [118,119] | ||
| Perinatal brain injury | [120,123] | ||
| Lung diseases | bronchopulmonary dysplasia |
| [133,134,138] |
| pulmonary fibrosis | [133,134,135,136,137] | ||
| Liver diseases | Acute liver injury |
| [23,141,142,143,144,145] |
| Liver fibrosis | [147,148] | ||
| Liver cirrhosis | [153,154,155] | ||
| Cancer | hepatocellular carcinoma |
| [25,159] |
| Hematopoietic cancer | [160] | ||
| prostate cancer | [161] | ||
| Glioma | [162] | ||
| epithelial ovarian cancer | [167] | ||
| colon cancer | [146] | ||
| Skin diseases | skin injury |
| [24,171,179] |
| Keloid | [180,181,182,183,184] | ||
| Bone diseases | Osteoarthritis |
| [185,186,187,188,189,190,191] |
| Cardiovascular disease | myocardial infarction |
| [200,201,202] |
| heart failure | [203,204] | ||
| Inflammatory bowel diseases | Crohn’s disease |
| [205,206,207] |
| Ulcerative Colitis | [206,207,208,209] | ||
| Kidney disease | chronic kidney failure |
| [210,211,212] |
| Diabetes mellitus | type 1 diabetes |
| [1,214,218] |
| type 2 diabetes | [1,214,218] | ||
| Autoimmune diseases | Multiple sclerosis |
| [220,221] |
| experimental autoimmune uveitis | [222] | ||
| hashimoto’s thyroiditis | [223] | ||
| systemic lupus erythematosus | [223,224] | ||
| Pre-eclampsia | [224] | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Q.-W.; Huang, Q.-M.; Wu, H.-Y.; Zuo, G.-S.-L.; Gu, H.-C.; Deng, K.-Y.; Xin, H.-B. Characteristics and Therapeutic Potential of Human Amnion-Derived Stem Cells. Int. J. Mol. Sci. 2021, 22, 970. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22020970
Liu Q-W, Huang Q-M, Wu H-Y, Zuo G-S-L, Gu H-C, Deng K-Y, Xin H-B. Characteristics and Therapeutic Potential of Human Amnion-Derived Stem Cells. International Journal of Molecular Sciences. 2021; 22(2):970. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22020970
Chicago/Turabian StyleLiu, Quan-Wen, Qi-Ming Huang, Han-You Wu, Guo-Si-Lang Zuo, Hao-Cheng Gu, Ke-Yu Deng, and Hong-Bo Xin. 2021. "Characteristics and Therapeutic Potential of Human Amnion-Derived Stem Cells" International Journal of Molecular Sciences 22, no. 2: 970. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22020970