Impact of Selected Signaling Proteins on SNAIL 1 and SNAIL 2 Expression in Ovarian Cancer Cell Lines in Relation to Cells’ Cisplatin Resistance and EMT Markers Level


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
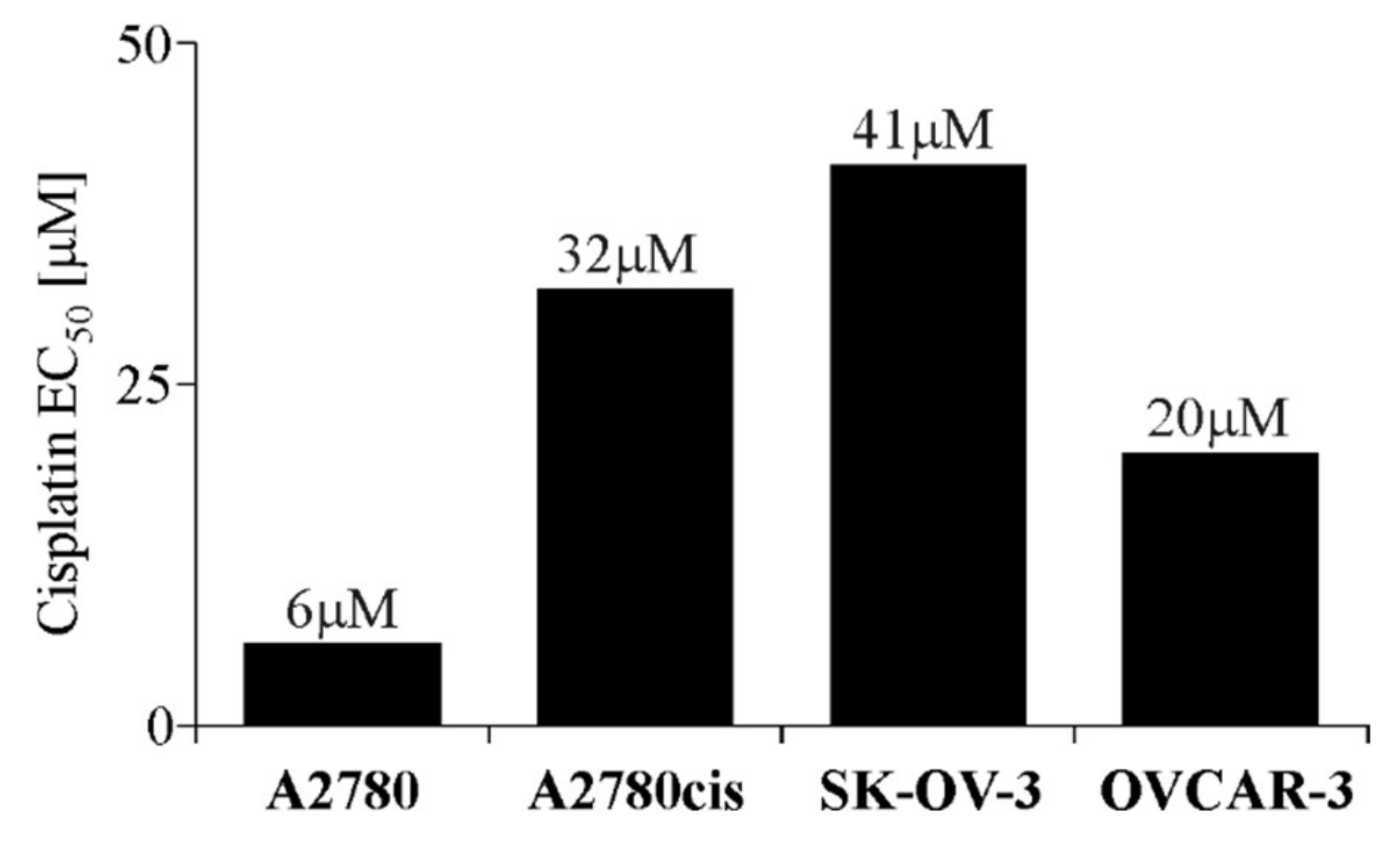
2.1. EC50 Value of Cisplatin for Each Ovarian Cancer Cell Line
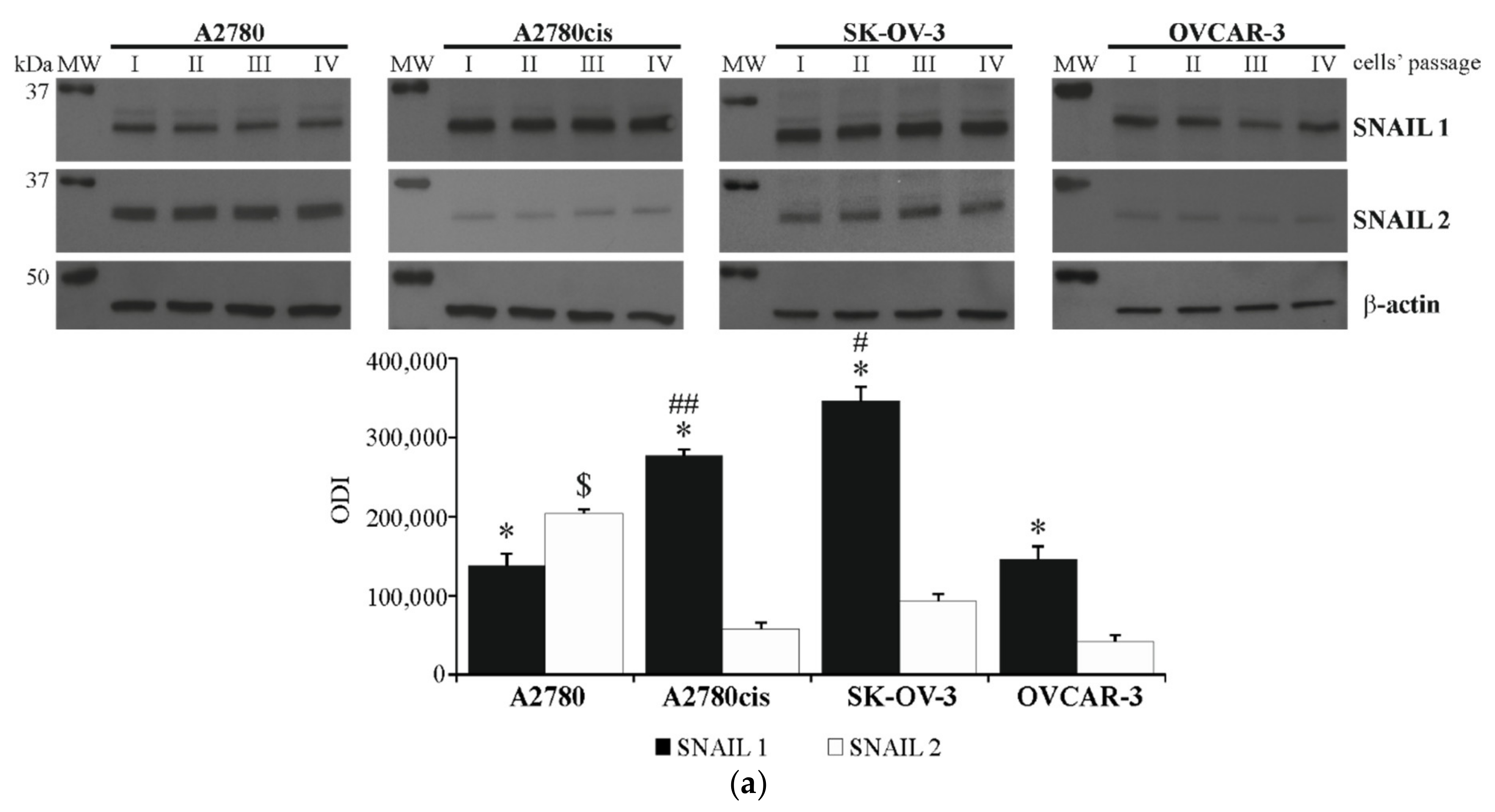
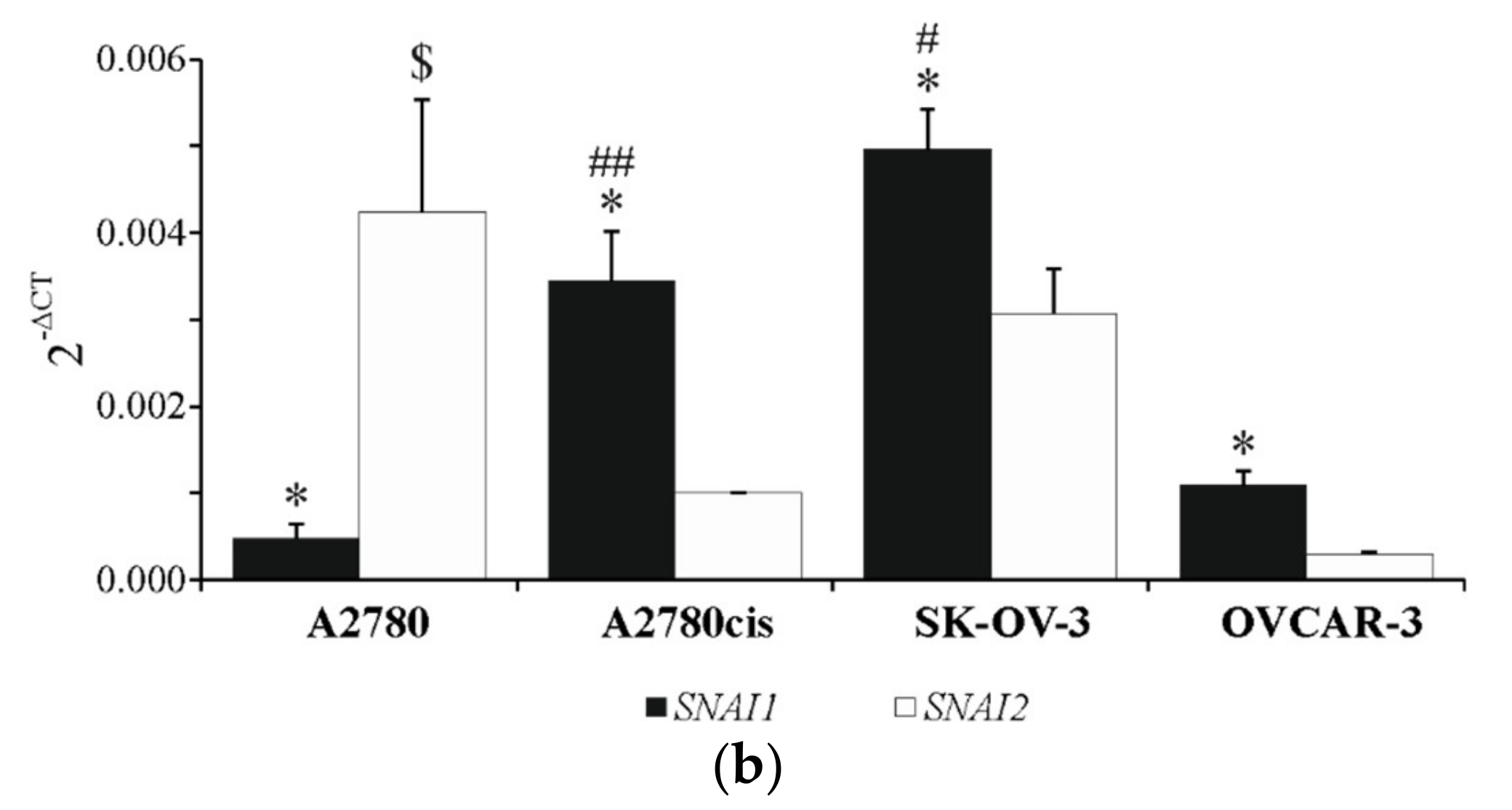
2.2. Basal Expression of SNAIL 1 and SNAIL 2 in Ovarian Cancer Cell Lines
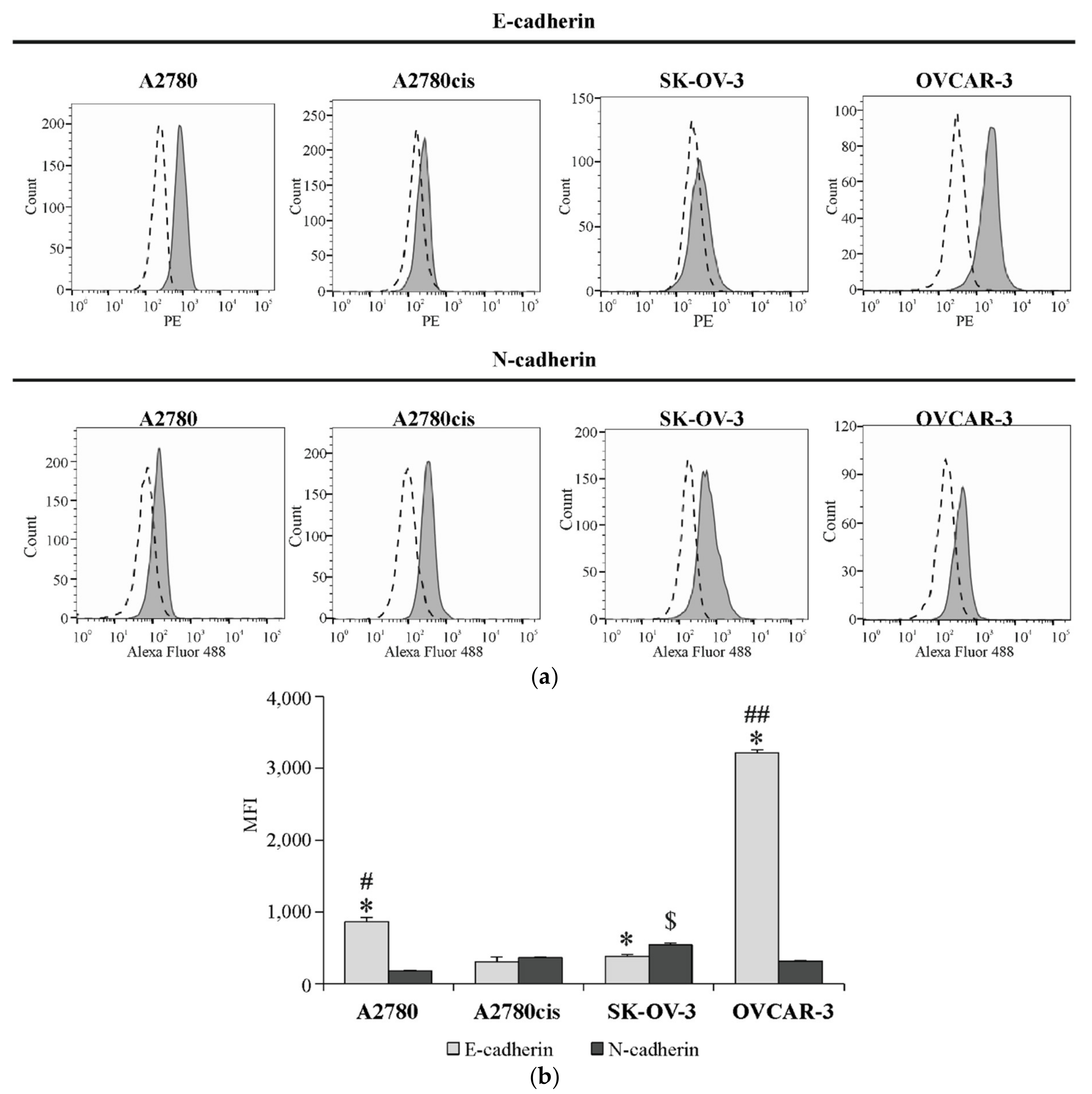
2.3. The Basal Surface Level of E-Cadherin and N-Cadherin on Ovarian Cancer Cell Lines
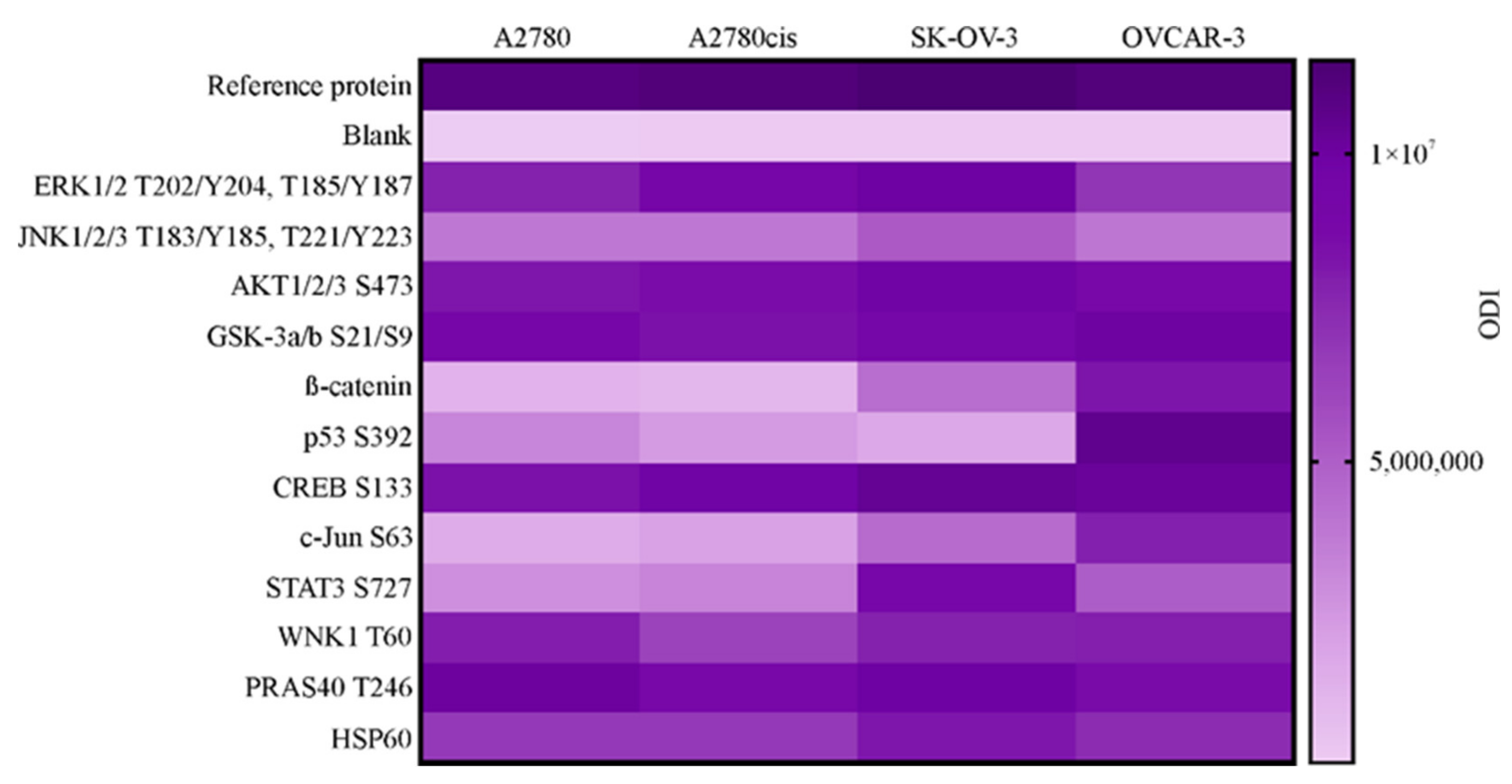
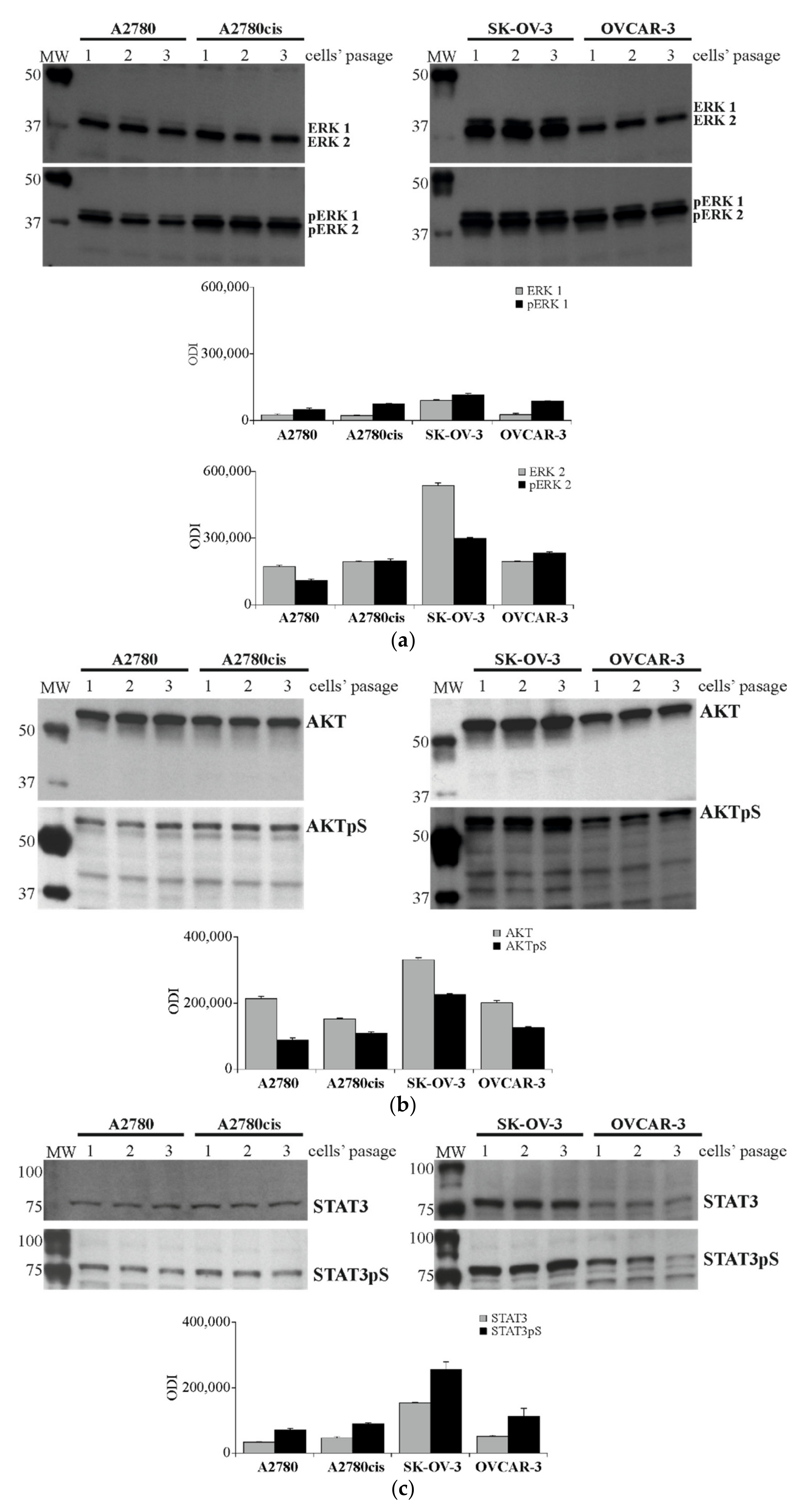
2.4. Profiling the Activity of 43 Proteins in Ovarian Cancer Cell Lines. Determination of Selected Proteins Level and Activity
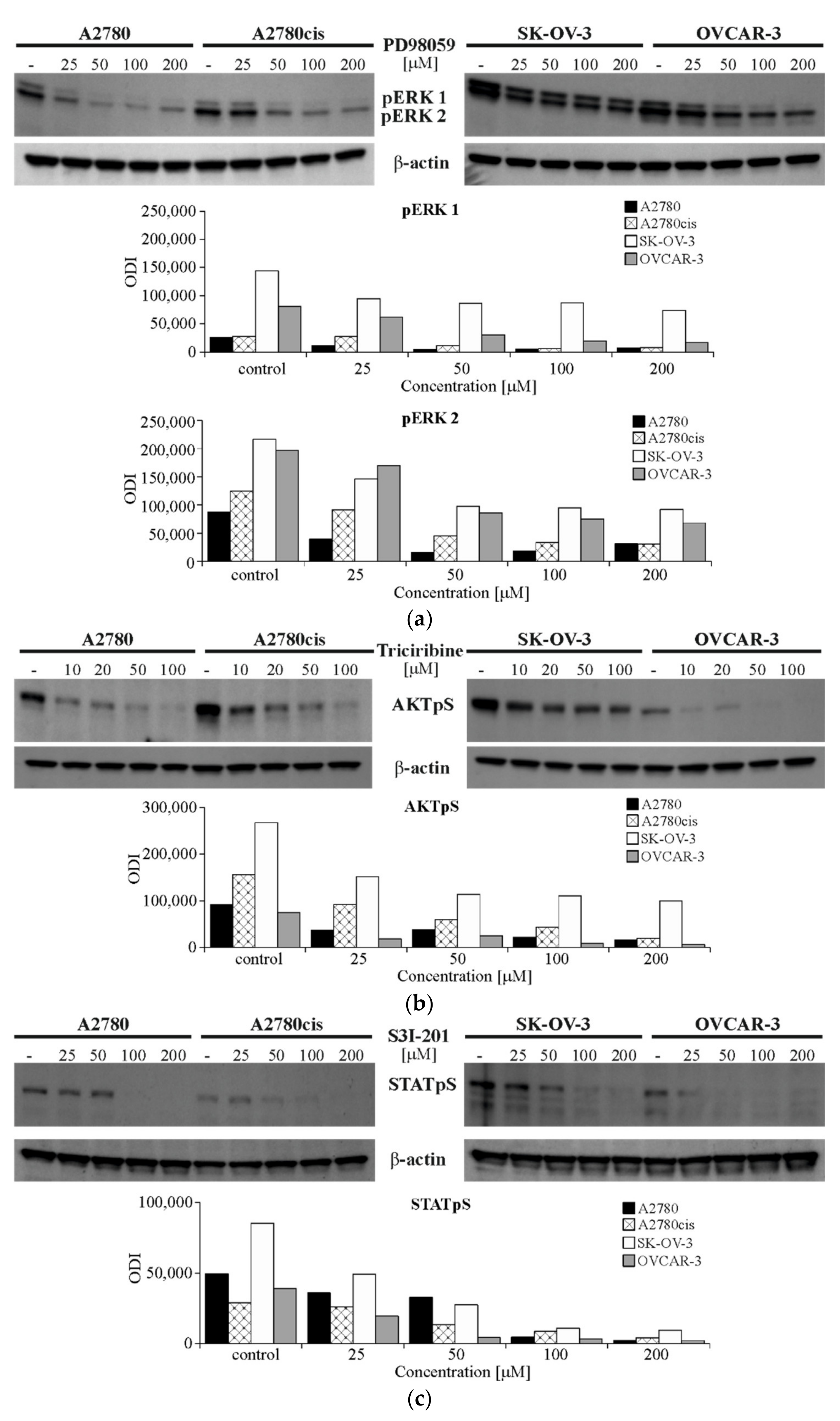
2.5. The Effect of Inhibiting ERK 1/2, AKT and STAT3 Signaling Proteins Activity on the Expression of SNAIL 1 and SNAIL 2 in Ovarian Cancer Cell Lines
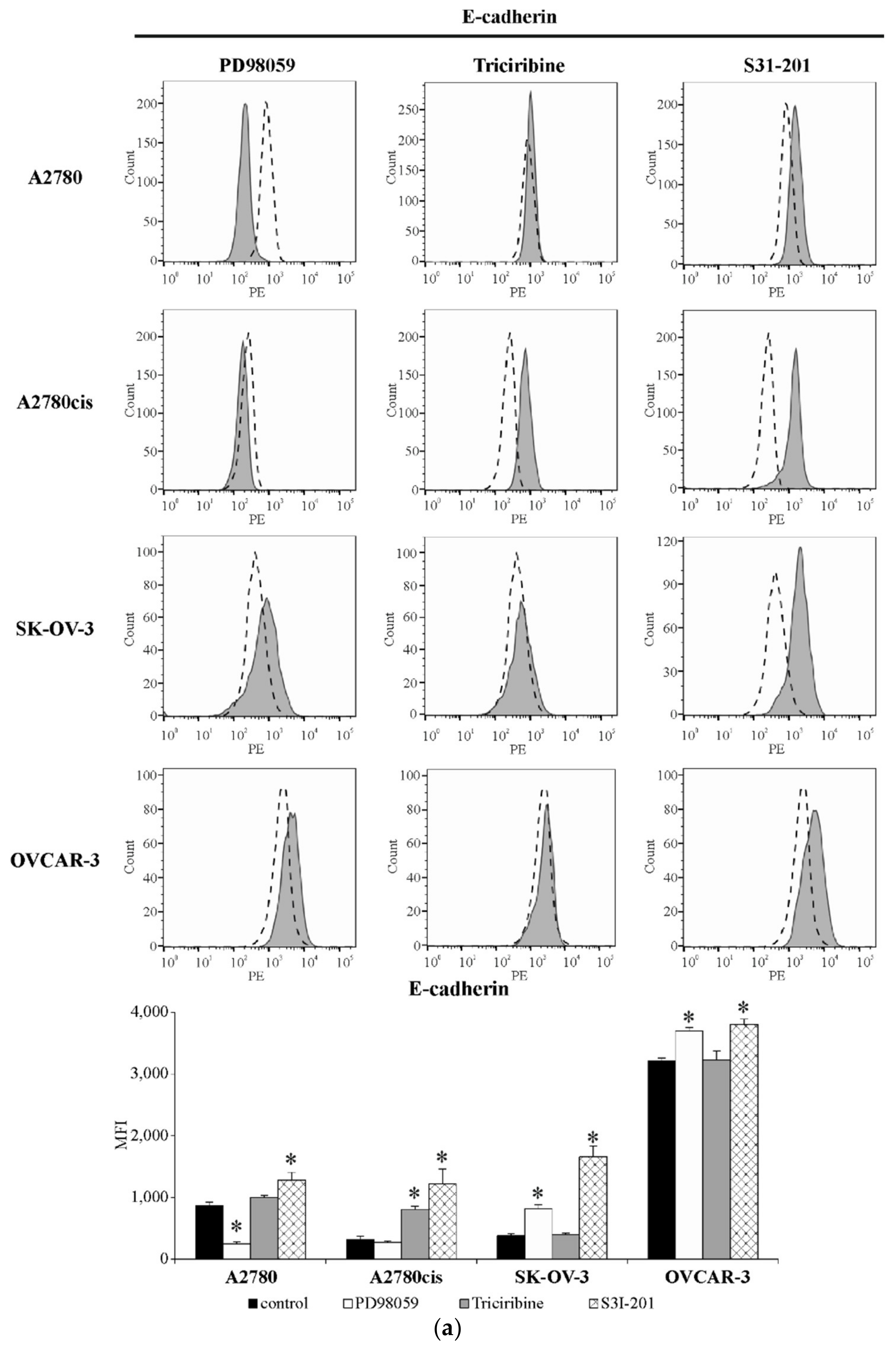
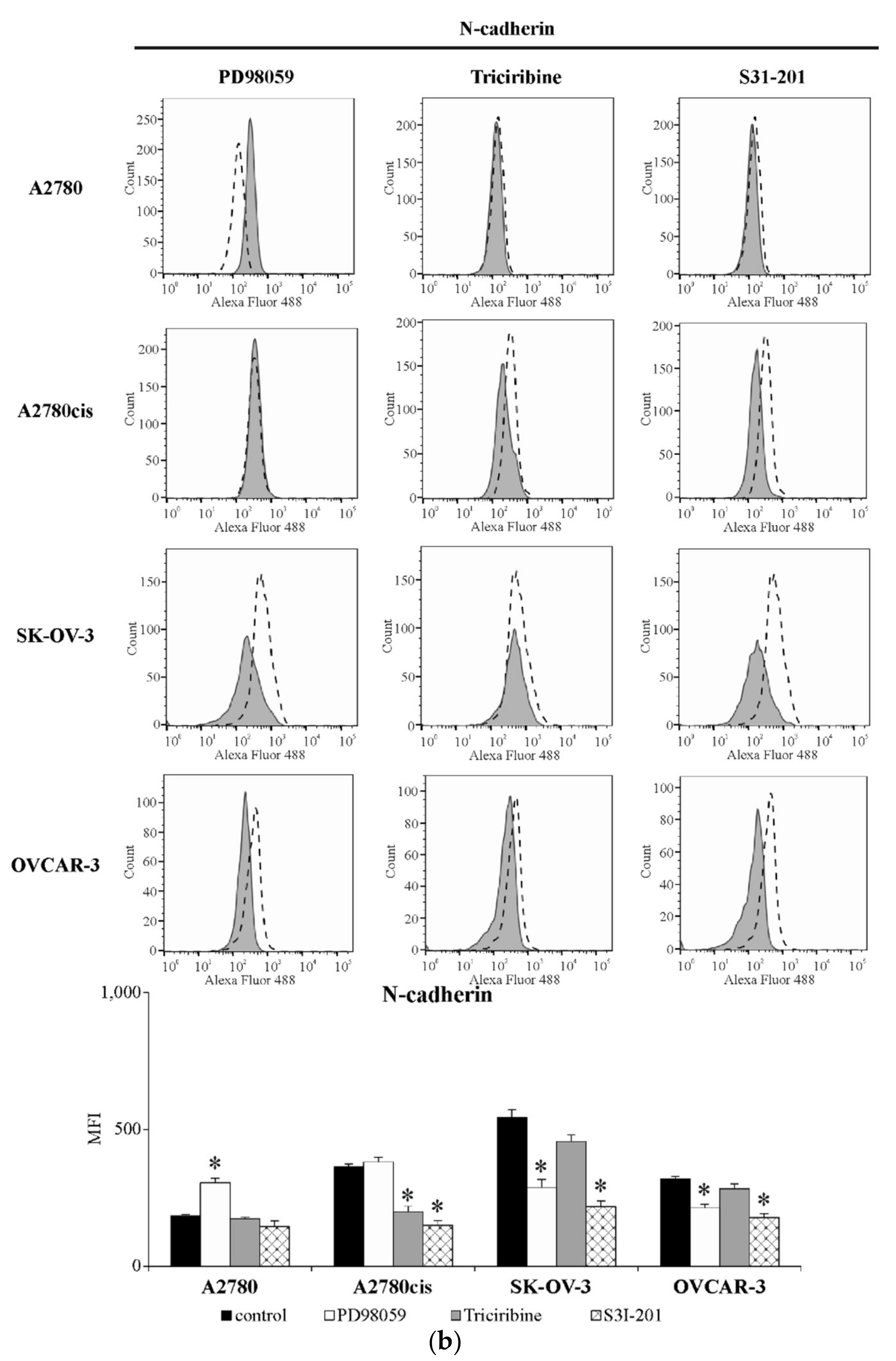
2.6. The Level of E-Cadherin and N-Cadherin in Ovarian Cancer Cell Lines Exposed to the Inhibitors of ERK 1/2, AKT and STAT3
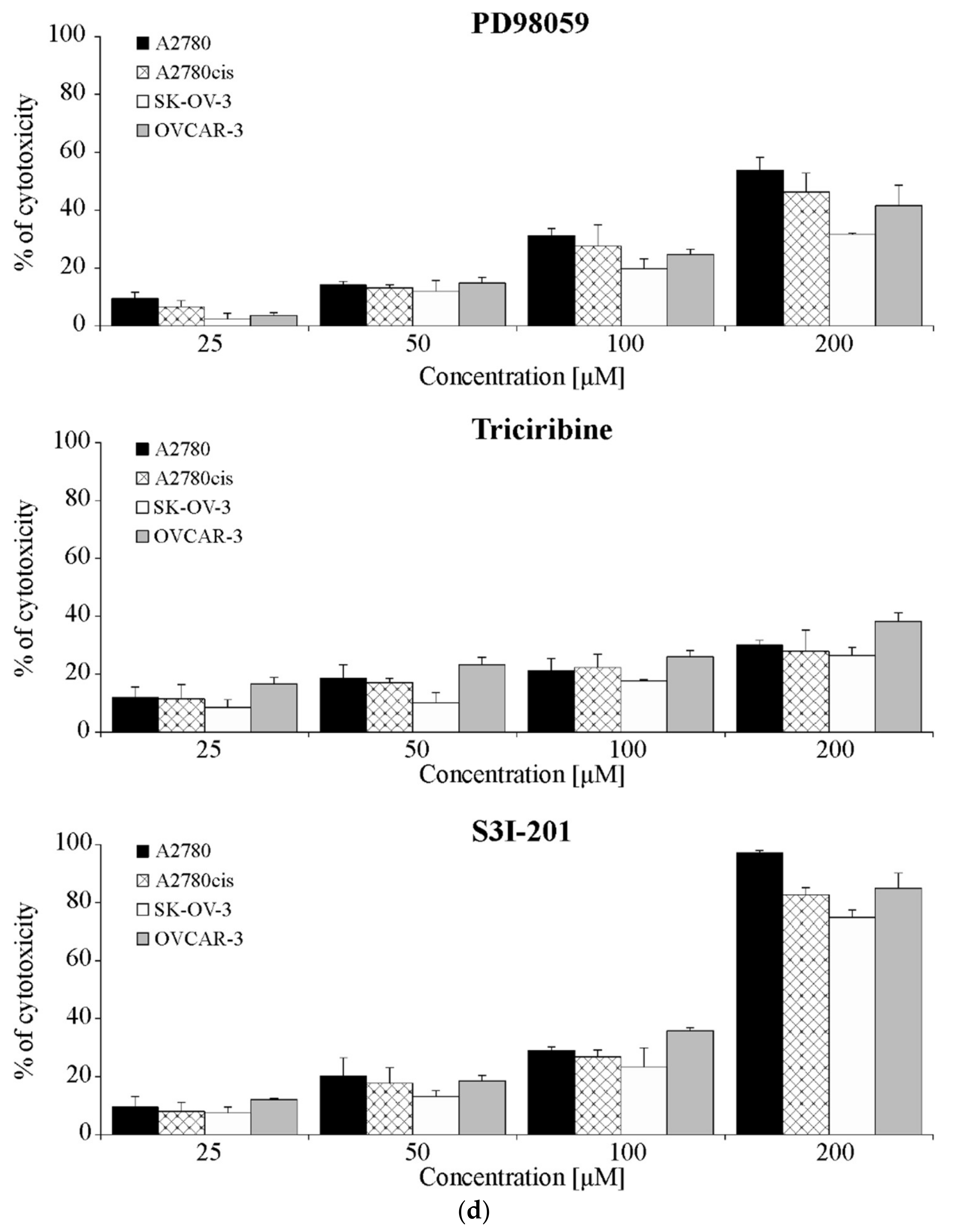
2.7. The Effect of Inhibiting ERK 1/2, AKT and STAT3 Signaling Proteins Activity on A2780, A2780cis, SK-OV-3 and OVCAR-3 Cells Response to Cisplatin Treatment
3. Discussion
4. Materials and Methods
4.1. Reagents and Antibodies
4.2. Cell Lines Culture and Treatment
4.3. Human Phospho-Kinase Array—Proteome Profiling
4.4. MTT Assay
4.5. Immunobloting ECL
4.6. Total RNA Isolation and Real-Time PCR Analysis
4.7. Flow Cytometry
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Memarzadeh, S.; Berek, J.S. Advances in the Management of Epithelial Ovarian Cancer. J. Reprod Med. 2001, 46, 621–629. [Google Scholar] [PubMed]
- Gupta, S.; Nag, S.; Aggarwal, S.; Rauthan, A.; Warrier, N. Maintenance Therapy for Recurrent Epithelial Ovarian Cancer: Current Therapies and Future Perspectives—A Review. J. Ovarian Res. 2019, 12, 103. [Google Scholar] [CrossRef] [PubMed]
- Braga, E.A.; Fridman, M.V.; Kushlinskii, N.E. Molecular Mechanisms of Ovarian Carcinoma Metastasis: Key Genes and Regulatory MicroRNAs. Biochem. Mosc. 2017, 82, 529–541. [Google Scholar] [CrossRef] [PubMed]
- Steeg, P.S. Perspectives on Classic Article: Metastasis Suppressor Genes. J. Natl. Cancer Inst. 2004, 96, E4. [Google Scholar] [CrossRef] [PubMed]
- Kalluri, R.; Weinberg, R.A. The Basics of Epithelial-Mesenchymal Transition. J. Clin. Invest. 2009, 119, 1420–1428. [Google Scholar] [CrossRef] [Green Version]
- Lamouille, S.; Xu, J.; Derynck, R. Molecular Mechanisms of Epithelial-Mesenchymal Transition. Nat. Rev. Mol. Cell Biol. 2014, 15, 178–196. [Google Scholar] [CrossRef] [Green Version]
- Dongre, A.; Weinberg, R.A. New Insights into the Mechanisms of Epithelial-Mesenchymal Transition and Implications for Cancer. Nat. Rev. Mol. Cell Biol. 2019, 20, 69–84. [Google Scholar] [CrossRef]
- Arumugam, T.; Ramachandran, V.; Fournier, K.F.; Wang, H.; Marquis, L.; Abbruzzese, J.L.; Gallick, G.E.; Logsdon, C.D.; McConkey, D.J.; Choi, W. Epithelial to Mesenchymal Transition Contributes to Drug Resistance in Pancreatic Cancer. Cancer Res. 2009, 69, 5820–5828. [Google Scholar] [CrossRef] [Green Version]
- McConkey, D.J.; Choi, W.; Marquis, L.; Martin, F.; Williams, M.B.; Shah, J.; Svatek, R.; Das, A.; Adam, L.; Kamat, A.; et al. Role of Epithelial-to-Mesenchymal Transition (EMT) in Drug Sensitivity and Metastasis in Bladder Cancer. Cancer Metastasis Rev. 2009, 28, 335–344. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Li, H.; Ren, G. Epithelial-Mesenchymal Transition and Drug Resistance in Breast Cancer (Review). Int. J. Oncol. 2015, 47, 840–848. [Google Scholar] [CrossRef] [Green Version]
- Chowanadisai, W.; Messerli, S.M.; Miller, D.H.; Medina, J.E.; Hamilton, J.W.; Messerli, M.A.; Brodsky, A.S. Cisplatin Resistant Spheroids Model Clinically Relevant Survival Mechanisms in Ovarian Tumors. PLoS ONE 2016, 11, e0151089. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashrafizadeh, M.; Zarrabi, A.; Hushmandi, K.; Kalantari, M.; Mohammadinejad, R.; Javaheri, T.; Sethi, G. Association of the Epithelial-Mesenchymal Transition (EMT) with Cisplatin Resistance. Int. J. Mol. Sci 2020, 21, 4002. [Google Scholar] [CrossRef] [PubMed]
- Haslehurst, A.M.; Koti, M.; Dharsee, M.; Nuin, P.; Evans, K.; Geraci, J.; Childs, T.; Chen, J.; Li, J.; Weberpals, J.; et al. EMT Transcription Factors Snail and Slug Directly Contribute to Cisplatin Resistance in Ovarian Cancer. BMC Cancer 2012, 12, 91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brozovic, A. The Relationship between Platinum Drug Resistance and Epithelial-Mesenchymal Transition. Arch. Toxicol. 2017, 91, 605–619. [Google Scholar] [CrossRef] [PubMed]
- Elloul, S.; Elstrand, M.B.; Nesland, J.M.; Tropé, C.G.; Kvalheim, G.; Goldberg, I.; Reich, R.; Davidson, B. Snail, Slug, and Smad-Interacting Protein 1 as Novel Parameters of Disease Aggressiveness in Metastatic Ovarian and Breast Carcinoma. Cancer 2005, 103, 1631–1643. [Google Scholar] [CrossRef] [PubMed]
- Hotz, B.; Arndt, M.; Dullat, S.; Bhargava, S.; Buhr, H.-J.; Hotz, H.G. Epithelial to Mesenchymal Transition: Expression of the Regulators Snail, Slug, and Twist in Pancreatic Cancer. Clin. Cancer Res. 2007, 13, 4769–4776. [Google Scholar] [CrossRef] [Green Version]
- Roy, H.K.; Smyrk, T.C.; Koetsier, J.; Victor, T.A.; Wali, R.K. The Transcriptional Repressor SNAIL Is Overexpressed in Human Colon Cancer. Dig. Dis. Sci. 2005, 50, 42–46. [Google Scholar] [CrossRef]
- Smith, B.N.; Odero-Marah, V.A. The Role of Snail in Prostate Cancer. Cell Adhesion Migr. 2012, 6, 433–441. [Google Scholar] [CrossRef] [Green Version]
- Cano, A.; Neto, M.A. Snail Transcription Factors. In Encyclopedia of Cancer; Schwab, M., Ed.; Springer: Berlin/Heidelberg, Germany, 2011; pp. 3456–3459. ISBN 978-3-642-16483-5. [Google Scholar]
- Kaufhold, S.; Bonavida, B. Central Role of Snail1 in the Regulation of EMT and Resistance in Cancer: A Target for Therapeutic Intervention. J. Exp. Clin. Cancer Res. 2014, 33, 62. [Google Scholar] [CrossRef]
- De Craene, B.; van Roy, F.; Berx, G. Unraveling Signalling Cascades for the Snail Family of Transcription Factors. Cell. Signal. 2005, 17, 535–547. [Google Scholar] [CrossRef]
- Wang, Y.; Shi, J.; Chai, K.; Ying, X.; Zhou, B.P. The Role of Snail in EMT and Tumorigenesis. Curr. Cancer Drug Targets 2013, 13, 963–972. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Li, G.; Zhang, K. Pro-Survival Effects by NF-ΚB, Akt and ERK(1/2) and Anti-Apoptosis Actions by Six1 Disrupt Apoptotic Functions of TRAIL-Dr4/5 Pathway in Ovarian Cancer. Biomed. Pharmacother. 2016, 84, 1078–1087. [Google Scholar] [CrossRef] [PubMed]
- Ghoneum, A.; Said, N. PI3K-AKT-MTOR and NFκB Pathways in Ovarian Cancer: Implications for Targeted Therapeutics. Cancers 2019, 11, 949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duan, Z.; Foster, R.; Bell, D.A.; Mahoney, J.; Wolak, K.; Vaidya, A.; Hampel, C.; Lee, H.; Seiden, M.V. Signal Transducers and Activators of Transcription 3 Pathway Activation in Drug-Resistant Ovarian Cancer. Clin. Cancer Res. 2006, 12, 5055–5063. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurrey, N.K.; Amit, K.; Bapat, S.A. Snail and Slug Are Major Determinants of Ovarian Cancer Invasiveness at the Transcription Level. Gynecol. Oncol. 2005, 97, 155–165. [Google Scholar] [CrossRef] [PubMed]
- Vergara, D.; Merlot, B.; Lucot, J.-P.; Collinet, P.; Vinatier, D.; Fournier, I.; Salzet, M. Epithelial-Mesenchymal Transition in Ovarian Cancer. Cancer Lett. 2010, 291, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Yao, Y.; Sun, L.; Zhou, J.; Miao, M.; Luo, S.; Deng, G.; Li, J.; Wang, J.; Tang, J. Snail Driving Alternative Splicing of CD44 by ESRP1 Enhances Invasion and Migration in Epithelial Ovarian Cancer. Cell. Physiol. Biochem. 2017, 43, 2489–2504. [Google Scholar] [CrossRef]
- Ma, S.; Tan, W.; Du, B.; Liu, W.; Li, W.; Che, D.; Zhang, G. Oridonin Effectively Reverses Cisplatin Drug Resistance in Human Ovarian Cancer Cells via Induction of Cell Apoptosis and Inhibition of Matrix Metalloproteinase Expression. Mol. Med. Rep. 2016, 13, 3342–3348. [Google Scholar] [CrossRef]
- Shinagawa, A.; Yoshida, Y.; Kurokawa, T.; Horiuchi, Y.; Tsuyoshi, H.; Orisaka, M.; Sawamura, Y.; Kleinman, H.K.; Kotsuji, F. The Potent Peptide Antagonist to Angiogenesis, C16Y, and Cisplatin Act Synergistically in the down-Regulation of the Bcl-2/Bax Ratio and the Induction of Apoptosis in Human Ovarian Cancer Cells. Int. J. Oncol. 2011, 39, 1359–1364. [Google Scholar] [CrossRef] [Green Version]
- Beaufort, C.M.; Helmijr, J.C.A.; Piskorz, A.M.; Hoogstraat, M.; Ruigrok-Ritstier, K.; Besselink, N.; Murtaza, M.; van IJcken, W.F.J.; Heine, A.A.J.; Smid, M.; et al. Ovarian Cancer Cell Line Panel (OCCP): Clinical Importance of In Vitro Morphological Subtypes. PLoS ONE 2014, 9, e103988. [Google Scholar] [CrossRef]
- Kielbik, M.; Szulc, I.; Brzezinska, M.; Bednarska, K.; Przygodzka, P.; Sulowska, Z.; Nowak, M.; Klink, M. Nitric Oxide Donors Reduce the Invasion Ability of Ovarian Cancer Cells in Vitro. Anticancer Drugs 2014, 25, 1141–1151. [Google Scholar] [CrossRef] [PubMed]
- Sonego, M.; Pellarin, I.; Costa, A.; Vinciguerra, G.L.R.; Coan, M.; Kraut, A.; D’Andrea, S.; Dall’Acqua, A.; Castillo-Tong, D.C.; Califano, D.; et al. USP1 Links Platinum Resistance to Cancer Cell Dissemination by Regulating Snail Stability. Sci. Adv. 2019, 5, eaav3235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhuo, W.; Wang, Y.; Zhuo, X.; Zhang, Y.; Ao, X.; Chen, Z. Knockdown of Snail, a Novel Zinc Finger Transcription Factor, via RNA Interference Increases A549 Cell Sensitivity to Cisplatin via JNK/Mitochondrial Pathway. Lung Cancer 2008, 62, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Hsu, D.S.-S.; Lan, H.-Y.; Huang, C.-H.; Tai, S.-K.; Chang, S.-Y.; Tsai, T.-L.; Chang, C.-C.; Tzeng, C.-H.; Wu, K.-J.; Kao, J.-Y.; et al. Regulation of Excision Repair Cross-Complementation Group 1 by Snail Contributes to Cisplatin Resistance in Head and Neck Cancer. Clin. Cancer Res. 2010, 16, 4561–4571. [Google Scholar] [CrossRef] [Green Version]
- Escrivà, M.; Peiró, S.; Herranz, N.; Villagrasa, P.; Dave, N.; Montserrat-Sentís, B.; Murray, S.A.; Francí, C.; Gridley, T.; Virtanen, I.; et al. Repression of PTEN Phosphatase by Snail1 Transcriptional Factor during Gamma Radiation-Induced Apoptosis. Mol. Cell. Biol. 2008, 28, 1528–1540. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Wang, C.; Jin, S.; Qu, D.; Ying, H. Expression of NF-ΚB and PTEN in Primary Epithelial Ovarian Carcinoma and the Correlation with Chemoresistance. Int. J. Clin. Exp. Pathol. 2015, 8, 10953–10963. [Google Scholar]
- Gheldof, A.; Berx, G. Chapter Fourteen—Cadherins and Epithelial-to-Mesenchymal Transition. In Progress in Molecular Biology and Translational Science; The Molecular Biology of Cadherins; van Roy, F., Ed.; Academic Press: Cambridge, MA, USA, 2013; Volume 116, pp. 317–336. [Google Scholar]
- Peinado, H.; Olmeda, D.; Cano, A. Snail, Zeb and BHLH Factors in Tumour Progression: An Alliance against the Epithelial Phenotype? Nat. Rev. Cancer 2007, 7, 415–428. [Google Scholar] [CrossRef]
- Yang, J.; Xing, H.; Hong, L.; Lu, D.; Li, B.; Tang, J.; Min, J. STAT3-Mediated Twist1 Upregulation Contributes to Epithelial-Mesenehymal Transition in Cisplatin Resistant Ovarian Cancer. Int. J. Clin. Exp. Med. 2018, 11, 6749–6757. [Google Scholar]
- Rosso, M.; Majem, B.; Devis, L.; Lapyckyj, L.; Besso, M.J.; Llauradó, M.; Abascal, M.F.; Matos, M.L.; Lanau, L.; Castellví, J.; et al. E-Cadherin: A Determinant Molecule Associated with Ovarian Cancer Progression, Dissemination and Aggressiveness. PLoS ONE 2017, 12, e0184439. [Google Scholar] [CrossRef] [Green Version]
- Ruibin, J.; Guoping, C.; Zhiguo, Z.; Maowei, N.; Danying, W.; Jianguo, F.; Linhui, G. Establishment and Characterization of a Highly Metastatic Ovarian Cancer Cell Line. Biomed. Res. Int. 2018, 2018. [Google Scholar] [CrossRef]
- Wang, L.; Zhou, S.; Guo, B. Vitamin D Suppresses Ovarian Cancer Growth and Invasion by Targeting Long Non-Coding RNA CCAT2. Int. J. Mol. Sci. 2020, 21, 2334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dobbin, Z.C.; Landen, C.N. The Importance of the PI3K/AKT/MTOR Pathway in the Progression of Ovarian Cancer. Int. J. Mol. Sci. 2013, 14, 8213–8227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, R.; Chen, X.; Chen, L.; Wan, F.; Chen, K.; Sun, Y.; Zhu, X. STAT3 Signaling in Ovarian Cancer: A Potential Therapeutic Target. J. Cancer 2020, 11, 837–848. [Google Scholar] [CrossRef] [Green Version]
- Yu, H.; Zhang, L.; Liu, P. CXCR7 Signaling Induced Epithelial-Mesenchymal Transition by AKT and ERK Pathways in Epithelial Ovarian Carcinomas. Tumor Biol. 2015, 36, 1679–1683. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.-Y.; Zhang, Y.-Y.; Zhu, B.-L.; Feng, F.-Z.; Zhang, H.-T.; Yan, H.; Zhou, B. MiR-203a-3p Regulates the Biological Behaviors of Ovarian Cancer Cells through Mediating the Akt/GSK-3β/Snail Signaling Pathway by Targeting ATM. J. Ovarian Res. 2019, 12, 60. [Google Scholar] [CrossRef]
- Liu, S.; Sun, J.; Cai, B.; Xi, X.; Yang, L.; Zhang, Z.; Feng, Y.; Sun, Y. NANOG Regulates Epithelial-Mesenchymal Transition and Chemoresistance through Activation of the STAT3 Pathway in Epithelial Ovarian Cancer. Tumor Biol. 2016, 37, 9671–9680. [Google Scholar] [CrossRef]
- Wang, J.; Zhou, J.-Y.; Wu, G.S. ERK-Dependent MKP-1-Mediated Cisplatin Resistance in Human Ovarian Cancer Cells. Cancer Res. 2007, 67, 11933–11941. [Google Scholar] [CrossRef] [Green Version]
- Ali, A.Y.; Kim, J.-Y.; Pelletier, J.-F.; Vanderhyden, B.C.; Bachvarov, D.R.; Tsang, B.K. Akt Confers Cisplatin Chemoresistance in Human Gynecological Carcinoma Cells by Modulating PPM1D Stability. Mol. Carcinog. 2015, 54, 1301–1314. [Google Scholar] [CrossRef]
- Zhang, X.; Liu, P.; Zhang, B.; Wang, A.; Yang, M. Role of STAT3 Decoy Oligodeoxynucleotides on Cell Invasion and Chemosensitivity in Human Epithelial Ovarian Cancer Cells. Cancer Genet. Cytogenet. 2010, 197, 46–53. [Google Scholar] [CrossRef]
- Lee, S.; Rauch, J.; Kolch, W. Targeting MAPK Signaling in Cancer: Mechanisms of Drug Resistance and Sensitivity. Int. J. Mol. Sci. 2020, 21, 1102. [Google Scholar] [CrossRef] [Green Version]
- Song, M.; Bode, A.M.; Dong, Z.; Lee, M.-H. AKT as a Therapeutic Target for Cancer. Cancer Res. 2019, 79, 1019–1031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Latifi, A.; Abubaker, K.; Castrechini, N.; Ward, A.C.; Liongue, C.; Dobill, F.; Kumar, J.; Thompson, E.W.; Quinn, M.A.; Findlay, J.K.; et al. Cisplatin Treatment of Primary and Metastatic Epithelial Ovarian Carcinomas Generates Residual Cells with Mesenchymal Stem Cell-like Profile. J. Cell. Biochem. 2011, 112, 2850–2864. [Google Scholar] [CrossRef] [PubMed]
- Cheng, S.; Guo, J.; Yang, Q.; Yang, X. Crk-like Adapter Protein Regulates CCL19/CCR7-Mediated Epithelial-to-Mesenchymal Transition via ERK Signaling Pathway in Epithelial Ovarian Carcinomas. Med. Oncol. 2015, 32, 47. [Google Scholar] [CrossRef] [PubMed]
- Sawada, K.; Mitra, A.K.; Radjabi, A.R.; Bhaskar, V.; Kistner, E.O.; Tretiakova, M.; Jagadeeswaran, S.; Montag, A.; Becker, A.; Kenny, H.A.; et al. Loss of E-Cadherin Promotes Ovarian Cancer Metastasis via A5-Integrin, Which Is a Therapeutic Target. Cancer Res. 2008, 68, 2329–2339. [Google Scholar] [CrossRef] [Green Version]
- Olea-Flores, M.; Zuñiga-Eulogio, M.D.; Mendoza-Catalán, M.A.; Rodríguez-Ruiz, H.A.; Castañeda-Saucedo, E.; Ortuño-Pineda, C.; Padilla-Benavides, T.; Navarro-Tito, N. Extracellular-Signal Regulated Kinase: A Central Molecule Driving Epithelial–Mesenchymal Transition in Cancer. Int. J. Mol. Sci. 2019, 20, 2885. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Y.; Zhao, X.; Xiao, Q.; Liu, Q.; Ding, K.; Yu, F.; Zhang, R.; Zhu, T.; Ge, G. Snail and Slug Mediate Tamoxifen Resistance in Breast Cancer Cells through Activation of EGFR–ERK Independent of Epithelial–Mesenchymal Transition. J. Mol. Cell Biol. 2014, 6, 352–354. [Google Scholar] [CrossRef] [Green Version]
- Yin, S.; Wang, P.; Yang, L.; Liu, Y.; Wang, Y.; Liu, M.; Qi, Z.; Meng, J.; Shi, T.-Y.; Yang, G.; et al. Wip1 Suppresses Ovarian Cancer Metastasis through the ATM/AKT/Snail Mediated Signaling. Oncotarget 2016, 7, 29359–29370. [Google Scholar] [CrossRef] [Green Version]
- Deng, J.; Bai, X.; Feng, X.; Ni, J.; Beretov, J.; Graham, P.; Li, Y. Inhibition of PI3K/Akt/MTOR Signaling Pathway Alleviates Ovarian Cancer Chemoresistance through Reversing Epithelial-Mesenchymal Transition and Decreasing Cancer Stem Cell Marker Expression. BMC Cancer 2019, 19, 618. [Google Scholar] [CrossRef]
- Liu, L.; Wu, N.; Wang, Y.; Zhang, X.; Xia, B.; Tang, J.; Cai, J.; Zhao, Z.; Liao, Q.; Wang, J. TRPM7 Promotes the Epithelial–Mesenchymal Transition in Ovarian Cancer through the Calcium-Related PI3K/AKT Oncogenic Signaling. J. Exp. Clin. Cancer Res. 2019, 38, 106. [Google Scholar] [CrossRef]
- Ma, Y.; Zhang, X.; Xu, X.; Shen, L.; Yao, Y.; Yang, Z.; Liu, P. STAT3 Decoy Oligodeoxynucleotides-Loaded Solid Lipid Nanoparticles Induce Cell Death and Inhibit Invasion in Ovarian Cancer Cells. PLoS ONE 2015, 10, e0124924. [Google Scholar] [CrossRef] [Green Version]
- Liang, F.; Ren, C.; Wang, J.; Wang, S.; Yang, L.; Han, X.; Chen, Y.; Tong, G.; Yang, G. The Crosstalk between STAT3 and P53/RAS Signaling Controls Cancer Cell Metastasis and Cisplatin Resistance via the Slug/MAPK/PI3K/AKT-Mediated Regulation of EMT and Autophagy. Oncogenesis 2019, 8, 59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kong, G.; Jiang, Y.; Sun, X.; Cao, Z.; Zhang, G.; Zhao, Z.; Zhao, Y.; Yu, Q.; Cheng, G. Irisin Reverses the IL-6 Induced Epithelial-Mesenchymal Transition in Osteosarcoma Cell Migration and Invasion through the STAT3/Snail Signaling Pathway. Oncol. Rep. 2017, 38, 2647–2656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, L.; Wang, Y.; Zhen, Y.; Li, D.; He, X.; Yang, H.; Zhang, H.; Liu, Q. Piperine Inhibits Colorectal Cancer Migration and Invasion by Regulating STAT3/Snail-Mediated Epithelial-Mesenchymal Transition. Biotechnol. Lett. 2020. [Google Scholar] [CrossRef] [PubMed]
- Schmittgen, T.D.; Livak, K.J. Analyzing Real-Time PCR Data by the Comparative C(T) Method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]













Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kielbik, M.; Szulc-Kielbik, I.; Klink, M. Impact of Selected Signaling Proteins on SNAIL 1 and SNAIL 2 Expression in Ovarian Cancer Cell Lines in Relation to Cells’ Cisplatin Resistance and EMT Markers Level. Int. J. Mol. Sci. 2021, 22, 980. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22020980
Kielbik M, Szulc-Kielbik I, Klink M. Impact of Selected Signaling Proteins on SNAIL 1 and SNAIL 2 Expression in Ovarian Cancer Cell Lines in Relation to Cells’ Cisplatin Resistance and EMT Markers Level. International Journal of Molecular Sciences. 2021; 22(2):980. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22020980
Chicago/Turabian StyleKielbik, Michal, Izabela Szulc-Kielbik, and Magdalena Klink. 2021. "Impact of Selected Signaling Proteins on SNAIL 1 and SNAIL 2 Expression in Ovarian Cancer Cell Lines in Relation to Cells’ Cisplatin Resistance and EMT Markers Level" International Journal of Molecular Sciences 22, no. 2: 980. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22020980