
Biochemical and Immunological implications of Lutein and Zeaxanthin

 , , , and
, , , and
Abstract
:1. Introduction
2. Lutein and Zeaxanthin in Different Kingdoms of Life
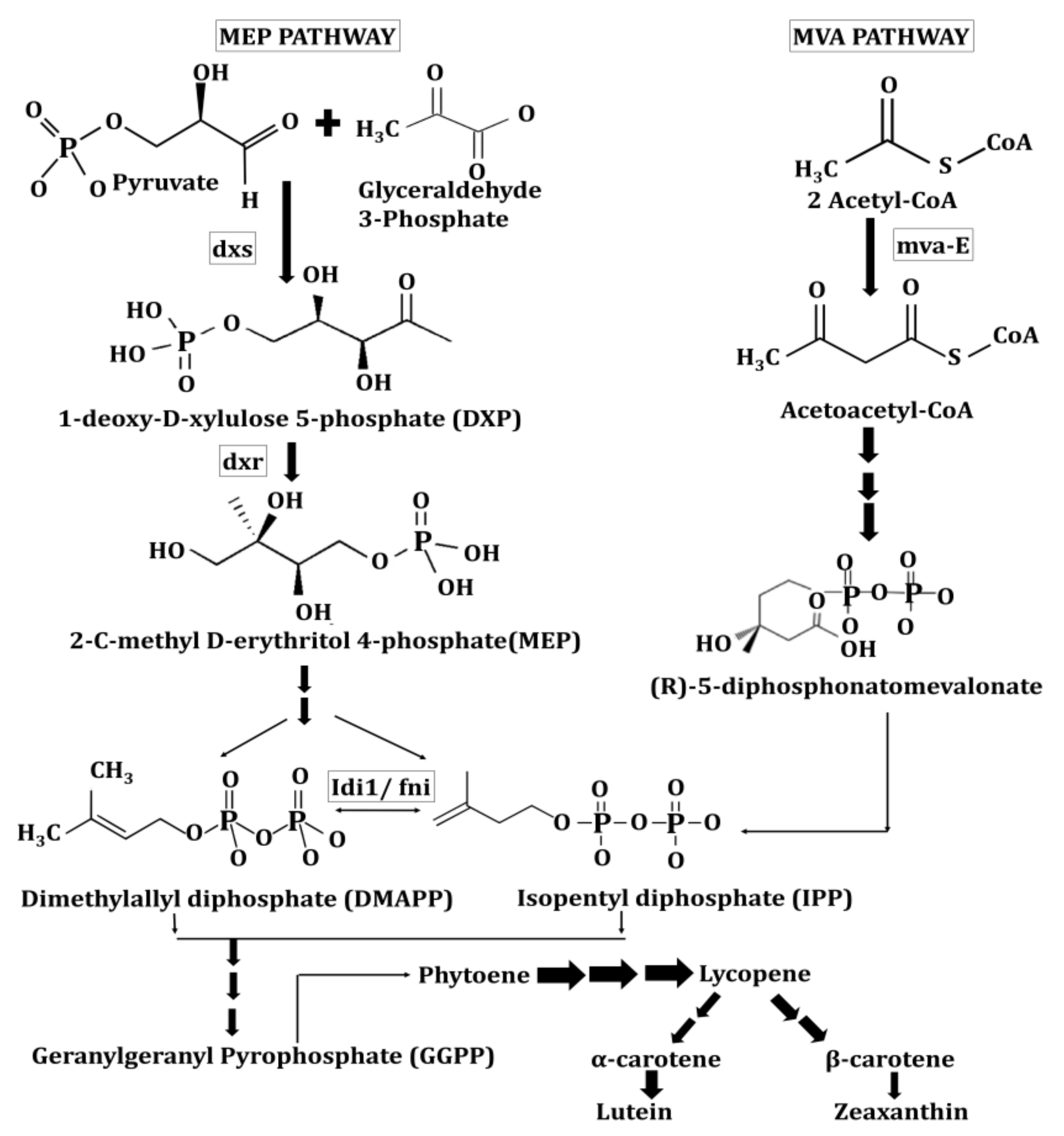
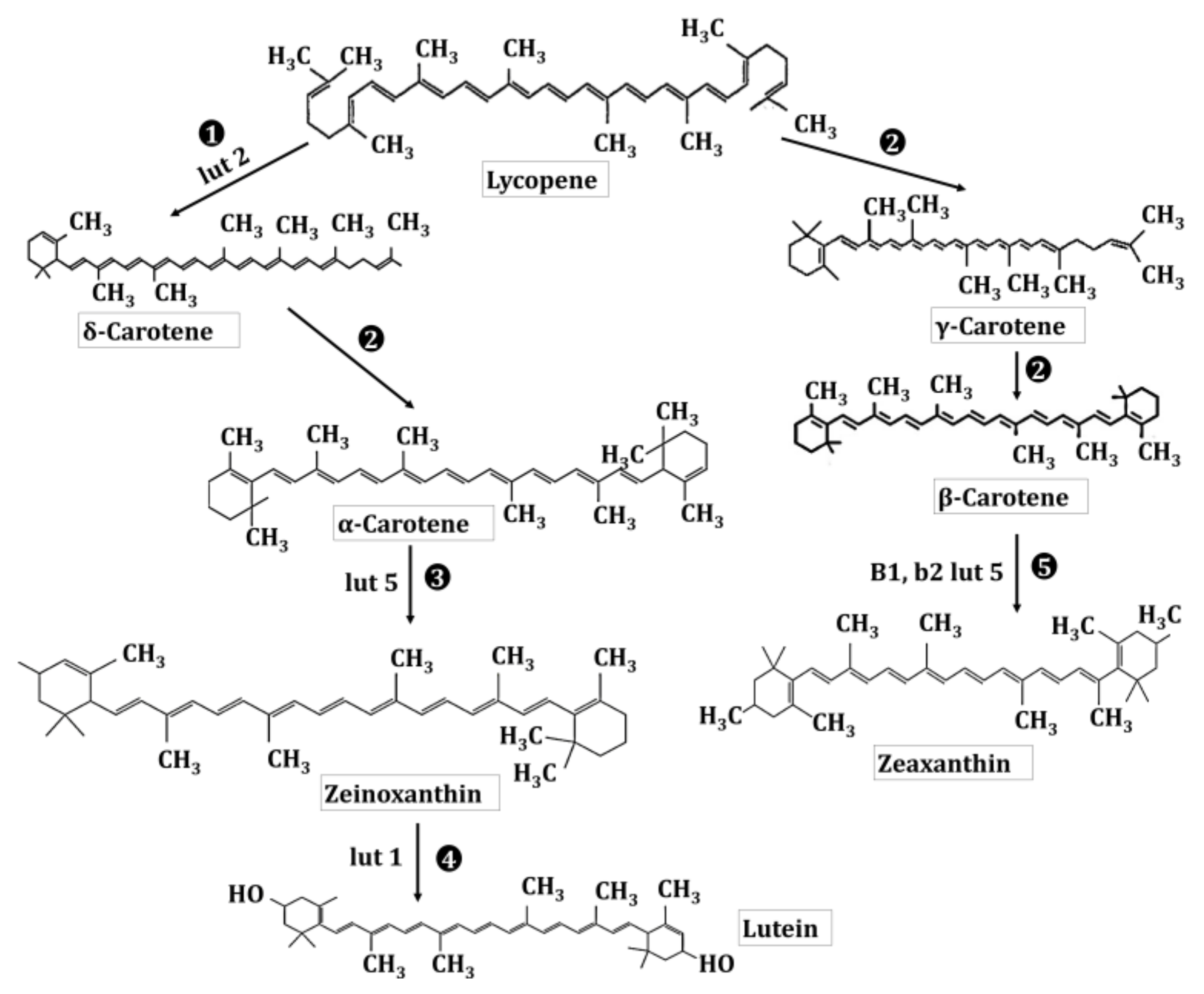
2.1. Biosynthesis and Presence of Lutein and Zeaxanthin in Kingdom Plantae and Animalia

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source | Common Names | Concentration of Lutein (mg/100g) | Concentration of Zeaxanthin (mg/100g) | Cumulative Concentration of Lutein + Zeaxanthin (mg/100g) | General Applications | Reference |
|---|---|---|---|---|---|---|
| Vegetables: | ||||||
| Asparagus officinalis L. | Garden asparagus/Sparrow grass | Cooked asparagus | 0.382–0.653 (in raw asparagus) | Antioxidant property | [36,37] | |
| 0.991 | 0.22 | |||||
| Asparagus acutifolius | Wild asparagus | 0.544–1.913 | N.A | N.A | Nutritional supplements | [38] |
| Brassica oleracea var. italica | Broccoli | Cooked broccoli | 1.079 | Antioxidant property | [39,40] | |
| 3.11–3.960 | 1.73628 | |||||
| Solanum nigrum L. | European black nightshade | 84.38 | N.A | N.A | Nutritional supplements | [41] |
| Hibiscus cannabinus L. | Kenaf | 104.24 | 4.59 | N.A | Antioxidant property | [42] |
| Capsicum annuum | Green chilies | 1.902 | 0.06285 | 13.74 | Antioxidant property | [43] |
| Solanum tuberosum | Potato | 0.1352–0.1524 | 0.0077 | N.A | Nutritional supplement | [44,45] |
| Daucus carota L. | Wild carrot | 40.17 | 0.59 | N.A | Antioxidant property | [37] |
| Lactuca sativa | Lettuce | 3.824 | N.A | 2.313 | Ophthalmological significance | [37] |
| Herbaceous parts | ||||||
| Ocimum basilicum L. | Basil | 4.2–8.3 | 0.2–0.6 | 6.634 ± 0.410 | Nutritional supplements | [46,47] |
| Petroselinum crispum | Parsley | 4.326 | 1.236 | 5.562 | Nutritional supplements | [48] |
| Coriandrum sativum | Coriander | 9.920 | N.A | 4.740 ± 4.36 | Adjuvants in food preparation + antioxidants | [43,49] |
| Mentha spicata | Spearmint | 17.74 | 0.28 | 18.00 | Nutritional supplement | [41,43] |
| Elattaria cardamomum | Green or true cardamom | 0.44 | N.A | 0.35 | Antioxidant property | [50] |
| Fruits | ||||||
| Malpighia emarginata | Barbados Cherry | 9.20 ± 0.23 | 1.14 ± 0.03 | N.A | Antioxidant property | [51] |
| Cucurbita maxima | Pumpkin | 10.62 | 0.278 | N.A | Antioxidant property | [50] |
| Solanum lycopersicum | Tomato | 0.289 | 0.0144 | 94 | Antioxidant property | [37] |
| Citrus X sinensis | Orange | 0.033 | 0.029 | 0.129 | Antioxidant property | [37] |
| Vitis vinifera ‘Red Globe’ | Red grapes | 0.024 | 0.004 | N.A | Antioxidant property | |
| Seed/Grain | ||||||
| Zea mays L. | Corn | 1.47 | 0.01031 | 0.01662–0.02057 | Antioxidant property | [52,53] |
| Triticum durum | Whear | 1.5–4.0 | 0.00049 | N.A | Nutritional supplmenets | [54,55] |
2.2. Prevalence of Lutein and Zeaxanthin in Kingdom Protista- Algae and Fungi
2.3. Biosynthesis of Lutein and Zeaxanthin in Domain Eubacteria
3. Enhancing the Production of Lutein and Zeaxanthin by Genetic Engineering Approach
3.1. Engineering Biosynthetic Pathway of Lutein by Random Mutagenesis Approach
3.2. Engineering Biosynthetic Pathway of Lutein and Zeaxanthin by Gene Assembly Approach
3.3. Enhanced Production of Zeaxanthin by CRISPR/Cas 9 Based Approaches
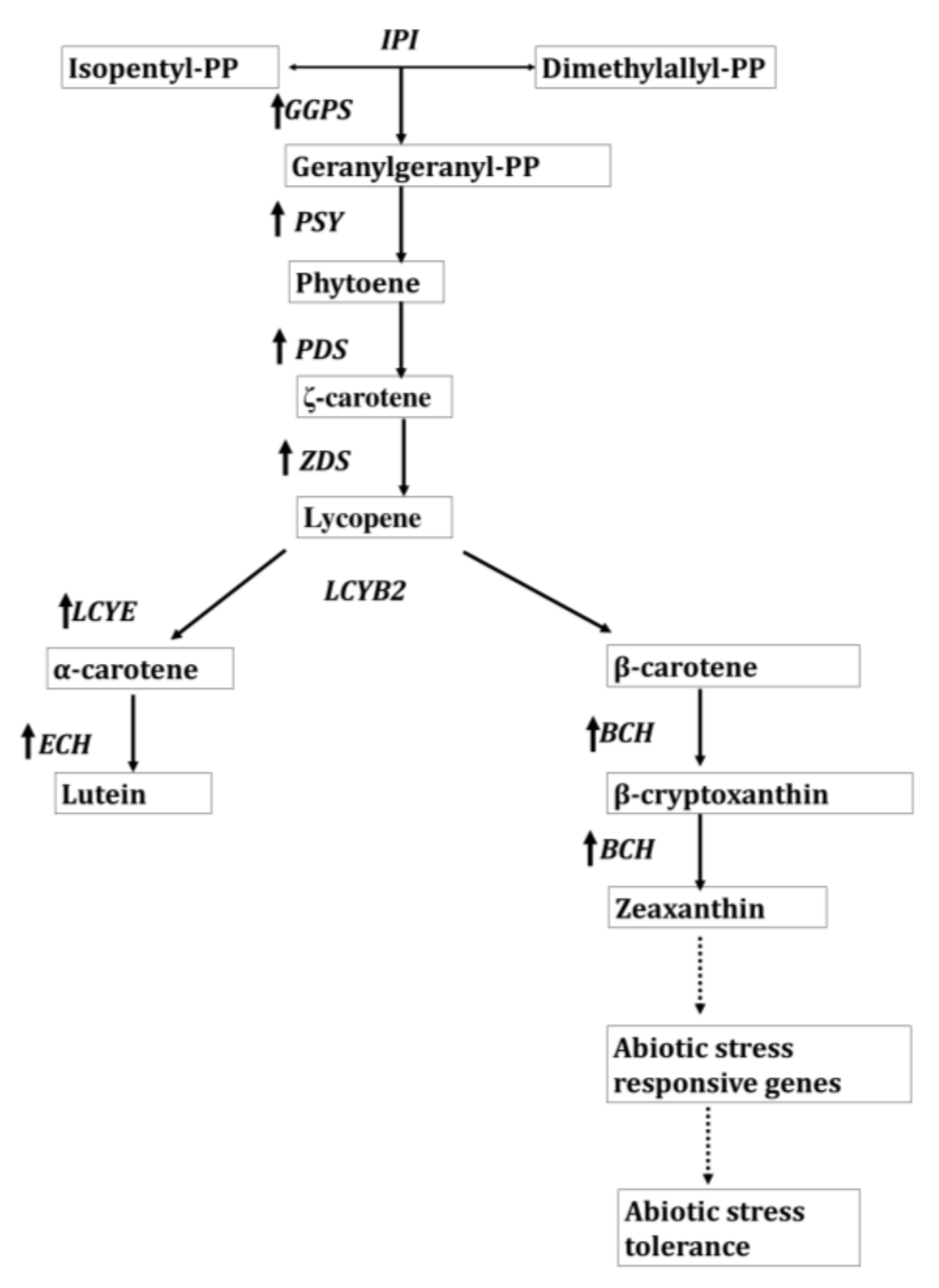
3.4. Regulation of Biosynthetic Pathways of Lutein and Zeaxanthin by Carotenoid Desaturases Cyclases
4. Esterification of Lutein and Zeaxanthin to Enhance the Stability and Bioavailability in Harsh Environmental Conditions
| Ester | Matrix | Results | Application | References |
|---|---|---|---|---|
| Lutein | ||||
| ω-3 polyunsaturated fatty acid esters of lutein | Fish oil | Better anti-oxidant ability and augmenting shelf-life | Oral supplements and nutraceuticals | [142] |
| Lutein esters | Nanoparticles | Improve ocular delivery efficiency | Oral drug formulation and eye-targeted drug delivery system | [143] |
| Microcapsules | Increased half-life of lutein | For feed industry | [144] | |
| Polyvinylpyrrolidone | Improved shelf life and solubility | Antioxidant | [145] | |
| Polyoxyethylene sorbitan monooleate capsules | Improved stability | Ocular delivery system | [135] | |
| Zeaxanthin | ||||
| Zeaxanthin dipalmitate | Sea Buckthorn oil and water emulsion | Increase bioaccesibility | Functional foods and nutraceutics | [146] |
| Zeaxanthin ester | Glycyrrhizic acid, arabinogalactan | Solubility enhancement | Food Industry | [147] |
5. Thermal Degradation Kinetics and Cold Adaptability of Lutein and Zeaxanthin
6. Medicinal Applications of Lutein and Zeaxanthin
6.1. Role of Lutein and Zeaxanthin in Treatment of Cognitive Impairment
6.2. Role of Lutein and Zeaxanthin in Diabetes
6.3. Role of Lutein and Zeaxanthin in Cancer
6.3.1. Treatment of Ovarian Cancer:
6.3.2. Treatment of Colorectal Cancer:
6.3.3. Treatment of Breast cancer
6.4. Role of Lutein and Zeaxanthin in Hepatic Disorders
6.5. Role of Lutein and Zeaxanthin in Pregnancy
6.6. Role of Lutein and Zeaxanthin in Treatment of COVID-19
7. Lutein and Zeaxanthin Mediated Biological Pathways
7.1. Antioxidative Pathway in Macula
Role of Human Macular Carotenoids in Reducing Alu RNA during Dry AMD
7.2. Zeaxanthin Mediated Apoptosis via ROS-Regulated Signaling Pathway in Human Gastric Cancer Cells
8. Industrial Applications of Lutein and Zeaxanthin
8.1. Textile Industry
8.2. Feed Industry
8.3. Cosmetics Industry
9. Future Perspective
10. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Arunkumar, R.; Gorusupudi, A.; Bernstein, P.S. The macular carotenoids: A biochemical overview. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2020, 1865, 158617. [Google Scholar] [CrossRef] [PubMed]
- Takaichi, S. Carotenoids in algae: Distributions, biosyntheses and functions. Mar. Drugs 2011, 9, 1101–1118. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Gan, Z.; Cui, Y.; Shi, C.; Shi, X. Structure and function characterization of the phytoene desaturase related to the lutein biosynthesis in Chlorella protothecoides CS-41. Mol. Biol. Rep. 2012, 40, 3351–3361. [Google Scholar] [CrossRef]
- Bernstein, P.S.; Li, B.; Vachali, P.P.; Gorusupudi, A.; Shyam, R.; Henriksen, B.S.; Nolan, J.M. Lutein, zeaxanthin, and meso-zeaxanthin: The basic and clinical science underlying carotenoid-based nutritional interventions against ocular disease. Prog. Retin. Eye Res. 2016, 50, 34–66. [Google Scholar] [CrossRef] [Green Version]
- Granado, F.; Olmedilla, B.; Blanco, I. Nutritional and clinical relevance of lutein in human health. Br. J. Nutr. 2003, 90, 487–502. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; George, E.W.; Rognon, G.T.; Gorusupudi, A.; Ranganathan, A.; Chang, F.Y.; Bernstein, P.S. Imaging lutein and zeaxanthin in the human retina with confocal resonance Raman microscopy. Proc. Nat. Acad. Sci. USA 2020, 117, 12352–12358. [Google Scholar] [CrossRef] [PubMed]
- Evans, M.; Beck, M.; Elliott, J.; Etheve, S.; Roberts, R.; Schalch, W. Effects of formulation on the bioavailability of lutein and zeaxanthin: A randomized, double-blind, cross-over, comparative, single-dose study in healthy subjects. Europ. J. Nutri. 2013, 52, 1381–1391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maoka, T. Recent progress in structural studies of carotenoids in animals and plants. Arch. Biochem. Biophys. 2009, 483, 191–195. [Google Scholar] [CrossRef] [PubMed]
- Renzi, L.M.; Bovier, E.R.; Hammond, B.R. A role for the macular carotenoids in visual motor response. Nutr. Neurosci. 2013, 16, 262–268. [Google Scholar] [CrossRef] [PubMed]
- Landrum, J.T.; Bone, R.A. Lutein, zeaxanthin, and the macular pigment. Arch. Biochem. Biophys. 2001, 385, 28–40. [Google Scholar] [CrossRef] [PubMed]
- The Global Market for Carotenoids. BCC Research. 2018. Retrieved 28 October 2018. Available online: https://www.bccresearch.com/market-research/food-and-beverage/carotenoids-market-fod025c.html (accessed on 5 September 2021).
- Cordero, B.F.; Obraztsova, I.; Couso, I.; Leon, R.; Vargas, M.A.; Rodriguez, H. Enhancement of lutein production in Chlorella sorokiniana (Chorophyta) by improvement of culture conditions and random mutagenesis. Mar. Drugs 2011, 9, 1607–1624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marino, T.; Casella, P.; Sangiorgio, P.; Verardi, A.; Ferraro, A.; Hristoforou, E.; Molino, A.; Musmarra, D. Natural beta-carotene: A microalgae derivate for nutraceutical applications. Chem. Eng. Trans. 2020, 79, 103–108. [Google Scholar]
- Zeaxanthin Market. Transparency Market Research. 2020. Available online: https://www.transparencymarketresearch.com/zeaxanthin-market (accessed on 5 September 2021).
- Qv, X.V.; Zeng, Z.P.; Jiang, J.G. Preparation of lutein microencapsulation by complex coacervation method and its physicochemical properties and stability. Food Hydrocoll. 2011, 25, 1596–1603. [Google Scholar] [CrossRef]
- Gouveia, L.; Empis, J. Relative stabilities of microalgal carotenoids in microalgal extracts, biomass and fish feed: Effect of storage conditions. Innov. Food Sci. Emerg. Technol. 2003, 4, 227–233. [Google Scholar] [CrossRef]
- Hirschberg, J. Carotenoid biosynthesis in flowering plants. Curr. Opin. Plant Biol. 2001, 4, 210–218. [Google Scholar] [CrossRef]
- Concepcion, R.M.; Boronat, A. Elucidation of the methylerythritol phosphate pathway for isoprenoid biosynthesis in bacteria and plastids. A metabolic milestone achieved through genomics. Plant physiology 2002, 130, 1079–1089. [Google Scholar] [CrossRef] [Green Version]
- Rohmer, M.; Knani, M.; Simonin, P.; Sutter, B.; Sahm, H. Isoprenoid biosynthesis in bacteria: A novel pathway for the early steps leading to isopentenyl diphosphate. Biochem. J. 1993, 295, 517–524. [Google Scholar] [CrossRef]
- Cunningham, F.X.; Lafond, T.P., Jr.; Gantt, E. Evidence of a role for LytB in the nonmevalonate pathway of isoprenoid biosynthesis. J. Bacteriol. 2000, 182, 5841–5848. [Google Scholar] [CrossRef] [Green Version]
- Arigoni, D.; Eisenreich, W.; Latzel, C.; Sagner, S.; Radykewicz, T.; Zenk, M.H.; Bacher, A. Dimethylallyl pyrophosphate is not the committed precursor of isopentenyl pyrophosphate during terpenoid biosynthesis from 1-deoxyxylulose in higher plants. Proc. Nat. Acad. Sci. USA 1999, 96, 1309–1314. [Google Scholar] [CrossRef] [Green Version]
- Jahns, P.; Holzwarth, A.R. The role of the xanthophyll cycle and of lutein in photoprotection of photosystem II. Biochim. Biophys. Acta 2012, 1817, 182–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cunningham, F.X.; Pogson, B.; Sun, Z.; McDonald, Z.; DellaPenna, K.A.; Gantt, E. Functional Analysis of the β and ε Lycopene Cyclase Enzymes of Arabidopsis Reveals a Mechanism for Control of Cyclic Carotenoid Formation. Plant Cell 1996, 8, 1613–1626. [Google Scholar] [CrossRef] [Green Version]
- Bialek-Bylka, G.; Sakano, Y.; Mizoguchi, T.; Shimamura, T.; Phillip, D.; Koyama, Y.; Young, A. Central-Cis isomers of lutein found in the major light-harvesting complex of Photosystem II (LHC IIb) of higher plants. Photosynth. Res. 2004, 56, 255–264. [Google Scholar] [CrossRef]
- Tian, L.; DellaPenna, D. Progress in understanding the origin and functions of carotenoid hydroxylases in plants. Arch. Biochem. Biophys. 2004, 430, 22–29. [Google Scholar] [CrossRef]
- Kim, J.; DellaPenna, D. Defining the primary route for lutein synthesis in plants: The role of Arabidopsis carotenoid beta-ring hydroxylase CYP97A3. Proc. Nat. Acad. Sci. USA 2006, 103, 3474–3479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Havaux, M.; García-Plazaola, J.I. Beyond non-photochemical fluorescence quenching: The overlapping antioxidant functions of zeaxanthin and tocopherols. In Non-Photochemical Quenching and Energy Dissipation in Plants, Algae and Cyanobacteria (Advances in Photosynthesis and Respiration); Demmig-Adams, B., Garab, G., Adams, W.W., III, Govindjee, Eds.; Springer: Dordrecht, The Netherlands, 2014; Volume 40, pp. 583–603. [Google Scholar]
- Calvo, M.M. Lutein: A valuable ingredient of fruit and vegetables. Crit. Rev. Food Sci. Nutr. 2005, 45, 671–696. [Google Scholar] [CrossRef] [PubMed]
- Sommerburg, O.; Keunen, J.E.; Bird, A.C.; van Kuijk, F.J. Fruits and vegetables that are sources for lutein and zeaxanthin: The macular pigment in human eyes. Br. J. Ophthalmol. 1998, 82, 907–910. [Google Scholar] [CrossRef]
- Humphries, J.M.; Khachik, F. Distribution of Lutein, Zeaxanthin, and Related Geometrical Isomers in Fruit, Vegetables, Wheat, and Pasta Products. J. Agric. Food Chem. 2003, 51, 1322–1327. [Google Scholar] [CrossRef]
- Harashima, K.; Ohno, T.; Sawachika, T.; Hidaka, T.; Ohnishi, E. Carotenoids in orange pupae of the swallowtail, Papilio xuthus. Insect Biochem. 1972, 2, 29–48. [Google Scholar] [CrossRef]
- Maoka, T.; Kawase, N.; Hironaka, M.; Nishida, R. Carotenoids of hemipteran insects, from the perspective of chemo-systematic and chemical ecological studies. Biochem. Sys. Ecol. 2021, 95, 104241. [Google Scholar] [CrossRef]
- Czeczuga, B. The presence of carotenoids in various species of Lepidoptera. Biochem. Sys. Ecol. 1986, 14, 345–351. [Google Scholar] [CrossRef]
- Asahina, S. The development of odonatology in the Far Esast. Odonatologica 1974, 3, 5–12. [Google Scholar]
- Maoka, T.; Kawase, N.; Ueda, T.; Nishida, R. Carotenoids of dragonflies, from the perspective of comparative biochemical and chemical ecological studies. Biochem. Sys. Ecol. 2020, 89, 104001. [Google Scholar] [CrossRef]
- Fernández-Cano, M.I.; Arreciado Maramon, A.; Feijoo-Cid, M. The Bachelor’s thesis in nursing: Characteristics and students’ approach and satisfaction. Nurs Educ Pract. 2021, 53, 103067. [Google Scholar] [CrossRef]
- Perry, A.; Rasmussen, H.; Johnson, E. Xanthophyll (lutein, zeaxanthin) content in fruits, vegetables and corn and egg products. J. Food Comp. Anal. 2009, 22, 9–15. [Google Scholar] [CrossRef]
- García-Herrera, P.; Sánchez-Mata, M.C.; Cámara, M.; Tardío, J.; Olmedilla-Alonso, B. Carotenoid content of wild edible young shoots traditionally consumed in Spain (Asparagus acutifolius L., Humulus lupulus L., Bryonia dioica Jacq. and Tamus communis L.). J. Sci. Food Agric. 2014, 94, 1914–1916. [Google Scholar] [CrossRef]
- De Sá, M.C.; Rodriguez-Amaya, D.B. Carotenoid composition of cooked green vegetables from restaurants. Food Chemistry 2003, 83, 595–600. [Google Scholar] [CrossRef]
- US Department of Agriculture, Agricultral Research Service, Nutrient Data Laboratory USDA National Nutrient Database for Standard Reference. Available online: http://www.ars.usda.gov/ba/bhnrc/ndl (accessed on 15 March 2016).
- Lakshminarayana, R.; Raju, M.; Krishnakantha, T.P.; Baskaran, V. Lutein and zeaxanthin in leafy greens and their bioavailability: Olive oil influences the absorption of dietary lutein and its accumulation in adult rats. J. Agric. Food Chem. 2007, 55, 6395–6400. [Google Scholar] [CrossRef] [PubMed]
- Bangera, M.; Sangeetha, R.K.; Vallikannan, B. Provitamin-A and xanthophyll carotenoids in vegetables and food grains of nutritional and medicinal importance. Int. J. Food Sci. Technol. 2011, 46, 315–323. [Google Scholar] [CrossRef]
- Aruna, G.; Baskaran, V. Comparative study on the levels of carotenoids lutein, zeaxanthin and β-carotene in Indian spices of nutritional and medicinal importance. Food Chem. 2010, 123, 404–409. [Google Scholar] [CrossRef]
- Fernandez-Orozco, R.; Gallardo-Guerrero, L.; Hornero-Méndez, D. Carotenoid profiling in tubers of different potato (Solanum sp) cultivars: Accumulation of carotenoids mediated by xanthophyll esterification. Food Chem. 2013, 141, 2864–2872. [Google Scholar] [CrossRef]
- Meléndez-Martínez, A.J.; Mapelli-Brahm, P.; Hornero-Méndez, D.; Vicario, I.M. Chapter 1: Structures, Nomenclature and General chemistry of carotenoids and their esters. In Food Chemistry, Function and Analysis; Mercadante, A.Z., Ed.; Royal Society of Chemistry: London, UK, 2019; pp. 3–50. [Google Scholar]
- Kopsell, D.; Curran-Celentano, J. Carotenoid and Chlorophyll Pigments in Sweet Basil Grown in the Field and Greenhouse. Hortscience 2005, 40, 1230–1233. [Google Scholar] [CrossRef]
- Daly, T.; Jiwan, M.A.; O’Brien, N.M.; Aherne, S.A. Carotenoid content of commonly consumed herbs and assessment of their bioaccessibility using an in vitro digestion model. Plant Foods Hum. Nutr. 2010, 65, 164–169. [Google Scholar] [CrossRef]
- Eisenhauer, B.; Natoli, S.; Liew, G.; Flood, V.M. Lutein and Zeaxanthin Food Sources, Bioavailability and Dietary Variety in Age-Related Macular Degeneration Protection. Nutrients 2017, 9, 120. [Google Scholar] [CrossRef]
- Singh, G.; Kawatra, A.; Sehgal, S. Nutritional composition of selected green leafy vegetables, herbs and carrots. Plant Foods Hum Nutr. 2001, 56, 359–364. [Google Scholar] [CrossRef]
- Aruna, G.; Bangera, M.; Baskaran, V. Lutein content of selected Indian vegetables and vegetable oils determined by HPLC. J. Food Compos. Anal. 2009, 22, 632–636. [Google Scholar] [CrossRef]
- Khonsarn, N.; Lawan, S. Level of Xanthophyll, Lutein and Zeaxanthin in Selected Thai Fruits Determined by HPLC. IPCBEE 2012, 39, 240. [Google Scholar]
- Updike, A.A.; Schwartz, S.J. Thermal processing of vegetables increases cis isomers of lutein and zeaxantin. J. Agric. Food Chem. 2003, 51, 6184–6190. [Google Scholar] [CrossRef] [PubMed]
- Rios, S.; Dias Paes, M.C.; Cardoso, W.S.; Borém, A.; Teixeira, F.F. Color of Corn Grains and Carotenoid Profile of Importance for Human Health. Am. J. Plant Sci. 2014, 5, 857–862. [Google Scholar] [CrossRef] [Green Version]
- Abdel-Aal, E.S.M.; Young, J.C.; Rabalski, I.; Hucl, P.; Frégeau Reid, J. Identification and Quantification of Seed Carotenoids in Selected Wheat Species. J. Agr. Food Chem. 2007, 55, 787–794. [Google Scholar] [CrossRef] [PubMed]
- Hentschel, V.; Kranl, K.; Hollmann, J.; Lindhauer, M.G.; Böhm, V.; Bitsch, R. Spectrophotometric determination of yellow pigment content and evaluation of carotenoids by high-performance liquid chromatography in durum wheat grain. J. Agric. Food Chem. 2002, 50, 6663–6668. [Google Scholar] [CrossRef]
- Del Campo, J.A.; Moreno, J.; Rodríguez, H.; Vargas, M.A.; Rivas, J.; Guerrero, M.G. Carotenoid content of chlorophycean microalgae: Factors determining lutein accumulation in Muriellopsis sp. (Chlorophyta). J. Biotechnol. 2000, 76, 51–59. [Google Scholar] [CrossRef]
- Saha, S.H.; Murray, P.; Ermis, H. Marine Microalgae for Potential Lutein Production. Appl. Sci. 2020, 10, 6457. [Google Scholar] [CrossRef]
- Ho, S.H.; Chan, M.C.; Liu, C.C.; Chen, C.Y.; Lee, W.L.; Lee, D.J.; Chang, J.S. Enhancing lutein productivity of an indigenous microalga Scenedesmus obliquus FSP-3 using light-related strategies. Bioresour. Technol. 2014, 152, 275–282. [Google Scholar] [CrossRef] [PubMed]
- Takaichi, S.; Yokoyama, A.; Mochimaru, M.; Uchida, H.; Murakami, A. Carotenogenesis diversification in phylogenetic lineages of Rhodophyta. J. Phycol. 2016, 52, 329–338. [Google Scholar] [CrossRef]
- Deli, J.; Gonda, S.; Nagy, L.Z.; Szabó, I.; Gulyás-Fekete, G.; Agócs, A.; Vasas, G. Carotenoid composition of three bloom-forming algae species. Food Res. Int. 2014, 65, 215–223. [Google Scholar] [CrossRef]
- García-González, M.; Moreno, J.; Manzano, J.C.; Florêncio, F.J.; Guerrero, M.G. Production of Dunaliella salina biomass rich in 9-cis-β-carotene and lutein in a closed tubular photobioreactor. J. Biotechnol. 2005, 115, 81–90. [Google Scholar] [CrossRef]
- El-Baky, H.A.; El Baz, F.K.; El-Baroty, G.S. Production of carotenoids from marine microalgae and its evaluation as safe food colorant and lowering cholesterol agents. Amer. Eur. J. Agric. Environ. Sci. 2007, 2, 792–800. [Google Scholar]
- Inbaraj, B.S.; Chien, J.T.; Chen, B.H. Improved high performance liquid chromatographic method for determination of carotenoids in the microalga Chlorella pyrenoidosa. J. Chromatogr. A 2006, 1102, 193–199. [Google Scholar] [CrossRef]
- Minhas, A.K.; Hodgson, P.; Barrow, C.J.; Sashidhar, B.; Adholeya, A. The isolation and identification of new microalgal strains producing oil and carotenoid simultaneously with biofuel potential. Bioresour. Technol. 2016, 211, 556–565. [Google Scholar] [CrossRef] [PubMed]
- Gayathri, S.; Rajasree, S.R.R.; Suman, T.Y.; Aranganathan, L.; Thriuganasambandam, R.; Narendrakumar, G. Induction of β, ε-carotene-3, 3′-diol (lutein) production in green algae Chlorella salina with airlift photobioreactor: Interaction of different aeration and light-related strategies. Biomass Convers. Biorefin. 2020, 11, 2003–2011. [Google Scholar] [CrossRef]
- Huang, W.; Lin, Y.; He, M.; Gong, Y.; Huang, J. Induced High-Yield Production of Zeaxanthin, Lutein, and β-Carotene by a Mutant of Chlorella zofingiensis. J. Agric. Food Chem. 2018, 66, 891–897. [Google Scholar] [CrossRef]
- Del Campo, J.A.; Rodríguez, H.; Moreno, J.; Vargas, M.Á.; Rivas, J.; Guerrero, M.G. Accumulation of astaxanthin and lutein in Chlorella zofingiensis (Chlorophyta). Appl. Microbiol. Biotechnol. 2004, 64, 848–854. [Google Scholar] [CrossRef] [PubMed]
- Zia-Ul-Haq, M. Historical and introductory aspects of carotenoids. In Carotenoids: Structure and Function in the Human Body; Zia-Ul-Haq, M., Dewanjee, S., Riaz, M., Eds.; Springer: Cham, Switzerland, 2021; pp. 1–42. [Google Scholar]
- Jin, E.; Feth, B.; Melis, A. A mutant of the green alga Dunaliella salina constitutively accumulates zeaxanthin under all growth conditions. Biotechnol. Bioeng. 2003, 81, 115–124. [Google Scholar] [CrossRef]
- Shi, X.; Zhang, X.; Chen, F. Heterotrophic production of biomass and lutein by Chlorella protothecoides on various nitrogen sources. Enzyme Microb. Technol. 2000, 27, 312–318. [Google Scholar] [CrossRef]
- Piña, F.; Contreras-Porcia, L. Enhancement of Xanthophyll Synthesis in Porphyra/Pyropia Species (Rhodophyta, Bangiales) by Controlled Abiotic Factors: A Systematic Review and Meta-Analysis. Mar. Drugs 2021, 19, 221. [Google Scholar] [CrossRef]
- Koizumi, J.; Takatani, N.; Kobayashi, N.; Mikami, K.; Miyashita, K.; Yamano, Y.; Wada, A.; Maoka, T.; Hosokawa, M. Carotenoid Profiling of a Red Seaweed Pyropia yezoensis: Insights into Biosynthetic Pathways in the Order Bangiales. Mar. Drugs 2018, 16, 426. [Google Scholar] [CrossRef] [Green Version]
- Oh, S.; Lee, E.; Choe, E. Light Effects on Lipid Oxidation, Antioxidants, and Pigments in Dried Laver (Porphyra) during Storage. Food Sci. Biotechnol. 2014, 23, 701–709. [Google Scholar] [CrossRef]
- Esteban, R.; Martínez, B.; Fernández-Marín, B.; Becerril, J.M.; García-Plazaola, J.I. Carotenoid composition in Rhodophyta: Insights into xanthophyll regulation in Corallina elongata. Eur. Jol. Phycol. 2009, 44, 221–230. [Google Scholar] [CrossRef]
- Zia-Ul-Haq, M. Past, present and future of Carotenoids Research. In Carotenoids: Structure and Function in the Human Body; Zia-Ul-Haq, M., Dewanjee, S., Riaz, M., Eds.; Springer: Cham, Switzerland, 2021; pp. 827–854. [Google Scholar]
- Goodwin, T.W.; Jamikorn, M. Studies in Carotenogenesis. Some Observations on Carotenoid Synthesis in Two Varieties of Euglena gracilis. J. Protozool. Res. 1954, 1, 216–219. [Google Scholar] [CrossRef]
- Manikprabhu, D.; Lingappa, K. γ Actinorhodin a natural and attorney source for the synthetic dye to detect acid production of fungi. Saudi J. Biol. Sci. 2013, 20, 163–168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venil, C.K.; Velmurugan, P.; Dufossé, L.; Devi, P.R.; Ravi, A.V. Fungal Pigments: Potential Coloring Compounds for Wide Ranging Applications in Textile Dyeing. J. Fungi 2020, 6, 68. [Google Scholar] [CrossRef]
- Dufosse, L. Microbial production of food grade pigments. Food Technol. Biotechnol. 2006, 44, 313–321. [Google Scholar]
- Mohamed, H.; El-Shanawany, A.R.; Shah, A.M.; Nazir, Y.; Naz, T.; Ullah, S.; Song, Y. Comparative Analysis of Different Isolated Oleaginous Mucoromycota Fungi for Their γ-Linolenic Acid and Carotenoid Production. BioMed. Res. Int 2020, 2020, 1–13. [Google Scholar] [CrossRef]
- Rodríguez-Sáiz, M.; de la Fuente, J.L.; Barredo, J.L. Metabolic engineering of Mucor circinelloides for zeaxanthin production. Methods Mol. Bio. (Clifton, N.J.) 2012, 898, 133–151. [Google Scholar] [CrossRef]
- Vila, E.; Hornero-Méndez, D.; Azziz, G.; Lareo, C.; Saravia, V. Carotenoids from heterotrophic bacteria isolated from Fildes Peninsula, King George Island. Antarctica. Biotechnol. Rep. 2019, 21, e00306. [Google Scholar] [CrossRef]
- Stankovic, I. Zeaxanthin. Chemical and Technical Assessment (CTA), 63rd ed.; Food and Agriculture Organization of the United Nations: Rome, Italy, 2004. [Google Scholar]
- Gierhart, D.L. Production of Zeaxanthin and Zeaxanthin-Containing Compositions. Applied Food Biotechnology, Inc. US Patent 5308759A, 3 May 1994. [Google Scholar]
- Choi, S.K.; Osawa, A.; Maoka, T.; Hattan, J.; Ito, K.; Uchiyama, A.; Suzuki, M.; Shindo, K.; Misawa, N. 3-β-Glucosyl-3’-β-quinovosyl zeaxanthin, a novel carotenoid glycoside synthesized by Escherichia coli cells expressing the Pantoea ananatis carotenoid biosynthesis gene cluster. Appl. Microbiol. Biotechnol. 2013, 97, 8479–8486. [Google Scholar] [CrossRef]
- Fidan, O.; Zhan, J. Discovery and engineering of an endophytic Pseudomonas strain from Taxus chinensis for efficient production of zeaxanthin diglucoside. J. Biol. Eng. 2019, 13, 66. [Google Scholar] [CrossRef] [Green Version]
- Vila, E.; Hornero-Méndez, D.; Lareo, C.; Saravia, V. Biotechnological production of zeaxanthin by an Antarctic Flavobacterium: Evaluation of culture conditions. J. Biotechnol. 2020, 319, 54–60. [Google Scholar] [CrossRef] [PubMed]
- Bhosale, P.; Bernstein, P.S. Beta-carotene production by Flavobacterium multivorum in the presence of inorganic salts and urea. J. Ind. Microbiol. Biotechnol. 2004, 31, 565–571. [Google Scholar] [CrossRef]
- Bhosale, P.; Larson, A.J.; Bernstein, P.S. Factorial analysis of tricarboxylic acid cycle intermediates for optimization of zeaxanthin production from Flavobacterium multivorum. J. Appl. Microbiol. 2004, 96, 623–629. [Google Scholar] [CrossRef] [PubMed]
- Alcantara, S.; Sanchez, S. Influence of carbon and nitrogen sources on Flavobacteriumgrowth and zeaxanthin biosynthesis. J. Ind. Microbiol. Biotechnol. 1999, 23, 697–700. [Google Scholar] [CrossRef] [PubMed]
- Asker, D.; Beppu, T.; Ueda, K. Mesoflavibacter zeaxanthinifaciens: A novel zeaxanthin-producing marine bacterium of the family Flavobacteriaceae. Sys. Appl. Microbiol. 2007, 30, 291–296. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Hwang, Y.M.; Baik, K.S.; Choi, K.S.; Ka, J.O.; Seong, C.N. Mesoflavibacter aestuarii sp., a zeaxanthin-producing marine bacterium isolated from seawater. Int. J. Syst. Evol. 2014, 64(Pt. 6), 1932–1937. [Google Scholar] [CrossRef]
- Hameed, A.; Shahina, M.; Lin, S.Y.; Lai, W.A.; Hsu, Y.H.; Liu, Y.C.; Young, C.C. Aquibacter zeaxanthinifaciens, a zeaxanthin-producing bacterium of the family Flavobacteriaceae isolated from surface seawater, and emended descriptions of the genera Aestuariibaculum and Gaetbulibacter. Int. J. Syst. Evol. 2014, 64 Pt 1, 138–145. [Google Scholar] [CrossRef] [Green Version]
- Misawa, N.; Shimada, H. Metabolic engineering for the production of carotenoids in non-carotenogenic bacteria and yeasts. J. Biotechnol. 1997, 59, 169–181. [Google Scholar] [CrossRef]
- Li, X.R.; Tian, G.Q.; Shen, H.J.; Liu, J.Z. Metabolic engineering of Escherichia coli to produce zeaxanthin. J. Ind. Microbiol. Biotechnol. 2015, 42, 627–636. [Google Scholar] [CrossRef]
- Asker, D.; Beppu, T.; Ueda, K. Nubsella zeaxanthinifaciens gen. nov., sp. nov., a zeaxanthin-producing bacterium of the family Sphingobacteriaceae isolated from freshwater. Int. J. Syst. Evol. 2008, 58 Pt 3, 601–606. [Google Scholar] [CrossRef] [PubMed]
- Hirschberg, J.; Chamovitz, D. Carotenoids in Cyanobacteria. In The Molecular Biology of Cyanobacteria; Kluwer Academic Publishers: Drive Norwell, MA, USA, 1994; pp. 559–579. [Google Scholar]
- Morone, J.; Lopes, G.; Preto, M.; Vasconcelos, V.; Martins, R. Exploitation of Filamentous and Picoplanktonic Cyanobacteria for Cosmetic Applications: Potential to Improve Skin Structure and Preserve Dermal Matrix Components. Mar. Drugs 2020, 18, 486. [Google Scholar] [CrossRef] [PubMed]
- Mora-Pale, J.M.; Perez-Munguia, S.; Gonzalez-Mejia, J.C.; Dordick, J.S.; Barzana, E. The lipase catalyzed hydrolysis of lutein diesters in nonaqueous media is favoured at extremely low water activities. Biotechnol Bioeng. 2007, 98, 535–542. [Google Scholar] [CrossRef]
- Khachik, F. Process for Isolation, Purification and Recrystallization of Lutein from Marigold Oleoresin and Uses Thereof. US Patent 5382714, 17 January 1995. [Google Scholar]
- Xu, X.; Shao, B.; Zhou, D.; Ye, S.; Wang, Y.; Chen, B. Process for the Isolation and Purification of Xanthophylls Crystals from Plant Oleoresin. US Patent 7271298B2, 18 September 2007. [Google Scholar]
- Yen, H.W.; Sun, C.H.; Ma, T.W. The comparison of lutein production by Scenedesmus sp. in the autotrophic and the mixotrophic cultivation. Appl. Biochem. Biotechnol. 2011, 164, 353–361. [Google Scholar] [CrossRef]
- Chen, J.H.; Chen, C.-Y.; Chang, J.-S. Lutein production with wild-type and mutant strains of Chlorella sorokiniana MB-1 under mixotrophic growth. J. Taiwan Inst. Chem. Eng. 2017, 79, 66–73. [Google Scholar] [CrossRef]
- Umeno, D.; Tobias, A.V.; Arnold, F.H. Diversifying carotenoid biosynthetic pathways by directed evolution. Microbiol. Mol. Biol. Rev. 2005, 69, 51–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matthews, P.D.; Wurtzel, E.T. Metabolic engineering of carotenoid accumulation in Escherichia coli by modulation of the isoprenoid precursor pool with expression of deoxyxylulose phosphate synthase. Appl. Microbiol. Biotechnol. 2000, 53, 396–400. [Google Scholar] [CrossRef] [PubMed]
- Nishizaki, T.; Tsuge, K.; Itaya, M.; Doi, N.; Yanagawa, H. Metabolic engineering of carotenoid biosynthesis in Escherichia coli by ordered gene assembly in Bacillus subtilis. Appl. Enviro. Microbiol. 2007, 73, 1355–1361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohamed, A.; Jansson, C. Influence of light on accumulation of photosynthesis-specific transcripts in the cyanobacterium Synechocystis 6803. Plant Mol. Biol. 1989, 13, 693–700. [Google Scholar] [CrossRef]
- Lagarde, D.; Beuf, L.; Vermaas, W. Increased production of zeaxanthin and other pigments by application of genetic engineering techniques to Synechocystis sp. strain PCC 6803. Appl. Environ. Microbiol. 2000, 66, 64–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takaichi, S.; Maoka, T.; Masamoto, K. Myxoxanthophyll in Synechocystis sp. PCC 6803 is myxol 2′-dimethyl-fucoside, (3R,2′S)-myxol 2′-(2,4-di-O-methyl-alpha-L-fucoside), not rhamnoside. Plant Cell Physiol. 2001, 42, 756–762. [Google Scholar] [CrossRef]
- Lehmann, M.; Vamvaka, E.; Torrado, A.; Jahns, P.; Dann, M.; Rosenhammer, L.; Aziba, A.; Leister, D.; Rühle, T. Introduction of the Carotenoid Biosynthesis α-Branch Into Synechocystis sp. PCC 6803 for Lutein Production. Front. Plant Sci. 2021, 12, 699424. [Google Scholar] [CrossRef]
- Baroli, I.; Do, A.D.; Yamane, T.; Niyogi, K.K. Zeaxanthin accumulation in the absence of a functional xanthophyll cycle protects Chlamydomonas reinhardtii from photooxidative stress. Plant Cell 2003, 15, 992–1008. [Google Scholar] [CrossRef] [Green Version]
- Baek, K.; Yu, J.; Jeong, J.; Sim, S.J.; Bae, S.; Jin, E. Photoautotrophic production of macular pigment in a Chlamydomonas reinhardtii strain generated by using DNA-free CRISPR-Cas9 RNP-mediated mutagenesis. Biotechnol. Bioeng. 2018, 115, 719–728. [Google Scholar] [CrossRef]
- Song, I.; Kim, J.; Baek, K.; Choi, Y.; Shin, B.; Jin, E. The generation of metabolic changes for the production of high-purity zeaxanthin mediated by CRISPR-Cas9 in Chlamydomonas reinhardtii. Microb. Cell Fact. 2020, 19, 220. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Kang, C.; Song, X.; Yu, L.; Liu, D.; He, S.; Zhai, H.; Liu, Q. A ζ-carotene desaturase gene, IbZDS, increases β-carotene and lutein contents and enhances salt tolerance in transgenic sweetpotato. Plant Sci. 2017, 262, 39–51. [Google Scholar] [CrossRef]
- Fantini, E.; Falcone, G.; Frusciante, S.; Giliberto, L.; Giuliano, G. Dissection of Tomato Lycopene Biosynthesis through Virus-Induced Gene Silencing. Plant Physiol. 2013, 163, 986–998. [Google Scholar] [CrossRef]
- Cazzonelli, C.I.; Pogson, B.J. Source to sink: Regulation of carotenoid biosynthesis in plants. Trends Plant Sci. 2010, 15, 266–274. [Google Scholar] [CrossRef]
- Blatt, A.; Bauch, M.E.; Pörschke, Y.; Lohr, M. A lycopene β-cyclase/lycopene ε-cyclase/light-harvesting complex-fusion protein from the green alga Ostreococcus lucimarinus can be modified to produce α-carotene and β-carotene at different ratios. Plant J. 2015, 82, 582–595. [Google Scholar] [CrossRef] [PubMed]
- Zeng, J.; Wang, C.; Chen, X.; Zang, M.; Yuan, C.; Wang, X.; Wang, Q.; Li, M.; Li, X.; Chen, L.; et al. The lycopene β-cyclase plays a significant role in provitamin A biosynthesis in wheat endosperm. BMC Plant Biol. 2015, 15, 112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, C.; Hong, Z.; Luyao, X.; Ning, Z.; He, S.; Liu, Q. A lycopene β-cyclase gene, IbLCYB2, enhances carotenoid contents and abiotic stress tolerance in transgenic sweetpotato. Plant Sci. 2018, 272, 243–254. [Google Scholar] [CrossRef] [PubMed]
- Albrecht, M.; Misawa, N.; Sandmann, G. Metabolic engineering of the terpenoid biosynthetic pathway of Escherichia coli for production of the carotenoids beta-carotene and zeaxanthin. Biotechnol. Lett. 1999, 21, 791–795. [Google Scholar] [CrossRef]
- Li, R.; Turner, S.D.; Brautigan, D.L. Xanthophylls lutein and zeaxanthin modify gene expression and induce synthesis of hyaluronan in keratinocyte model of human skin. Biochem. Biophys. Rep. 2015, 4, 52–58. [Google Scholar] [CrossRef] [Green Version]
- Zia-Ul-Haq, M.; Riaz, M.; Ahmedah, H.T. Carotenoids and Periodontal Diseases. In Carotenoids: Structure and Function in the Human Body; Zia-Ul-Haq, M., Dewanjee, S., Riaz, M., Eds.; Springer: Cham, Switzerland, 2021; pp. 715–720. [Google Scholar]
- Yu, B.; Lydiate, D.J.; Young, L.W.; Schäfer, U.A.; Hannoufa, A. Enhancing the carotenoid content of Brassica napus seeds by downregulating lycopene epsilon cyclase. Transgenic Res. 2008, 17, 573–585. [Google Scholar] [CrossRef]
- Atienza, S.G.; Ballesteros, J.; Martín, A.; Hornero-Méndez, D. Genetic variability of carotenoid concentration and degree of esterification among tritordeum (xTritordeum Ascherson et Graebner) and durum wheat accessions. J. Agric. Food Chem. 2007, 55, 4244–4251. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, D.B.; Mercadante, A.Z.; Mariutti, L. Marigold carotenoids: Much more than lutein esters. Food Res. Int. (Ottawa, Ont.) 2019, 119, 653–664. [Google Scholar] [CrossRef] [PubMed]
- Mares, D.J.; Campbell, A.W. Mapping components of flour and noodle colour in Australian wheat. Aust. J. Agric. Res. 2001, 52, 1297–1309 101071/AR01048. [Google Scholar] [CrossRef] [Green Version]
- Bone, R.A.; Landrum, J.T.; Friedes, L.M.; Gomez, C.M.; Kilburn, M.D.; Vidal, E.M.; Wang, W. Distribution of lutein and zeaxanthin stereoisomers in the human retina. Exp. Eye Res. 1997, 64, 211–218. [Google Scholar] [CrossRef] [Green Version]
- Abdel-Aal, E.S.M.; Young, J.C.; Akhtar, H.; Rabalski, I. Stability of lutein in wholegrain bakery products naturally high in lutein or fortified with free lutein. J. Agric. Food. Chem. 2010, 58, 10109–10117. [Google Scholar] [CrossRef]
- Hidalgo, A.; Brandolini, A. Kinetics of carotenoid degradation during the storage of Einkorn (Triticum monococcum L. ssp. monococcum) and bread wheat (Triticum aestivum L. ssp. aestivum) flours. J. Agric. Food Chem. 2008, 56, 11300–11305. [Google Scholar] [CrossRef]
- Zia-Ul-Haq, M.; Riaz, M.; Modhi, A.O. Carotenoids and Bone Health. In Carotenoids: Structure and Function in the Human Body; Zia-Ul-Haq, M., Dewanjee, S., Riaz, M., Eds.; Springer: Cham, Switzerland, 2021; pp. 697–713. [Google Scholar]
- Requena-Ramírez, M.D.; Atienza, S.G.; Hornero-Méndez, D.; Rodríguez-Suárez, C. Mediation of a GDSL Esterase/Lipase in Carotenoid Esterification in Tritordeum Suggests a Common Mechanism of Carotenoid Esterification in Triticeae Species. Front. Plant Sci. 2020, 11, 592515. [Google Scholar] [CrossRef] [PubMed]
- Watkins, J.L.; Li, M.; McQuinn, R.P.; Chan, K.X.; McFarlane, H.E.; Ermakova, M.; Furbank, R.T.; Mares, D.; Dong, C.; Chalmers, K.J.; et al. A GDSL Esterase/Lipase Catalyzes the Esterification of Lutein in Bread Wheat. Plant Cell 2019, 31, 3092–3112. [Google Scholar] [CrossRef] [PubMed]
- Ziegler, J.U.; Wahl, S.; Würschum, T.; Longin, C.F.H.; Carle, R.; Schweiggert, R.M. Lutein and lutein esters in whole grain flours made from 75 genotypes of 5 triticum species grown at multiple sites. J. Agric. Food Chem. 2015, 63, 5061–5071. [Google Scholar] [CrossRef]
- Shangguan, H.; Zhang, S.; Li, X.; Zhou, Q.; Shi, J.; Deng, Q.; Huang, F. Synthesis of lutein esters using a novel biocatalyst of Candida antarctica lipase B covalently immobilized on functionalized graphitic carbon nitride nanosheets. RSC Adv. 2020, 10, 8949–8957. [Google Scholar] [CrossRef]
- Gombač, Z.; Črnivec, I.; Skrt, M.; Istenič, K.; Knafelj, A.K.; Pravst, I.; Ulrih, N.P. Stabilisation of Lutein and Lutein Esters with Polyoxyethylene Sorbitan Monooleate, Medium-Chain Triglyceride Oil and Lecithin. Foods 2021, 10, 500. [Google Scholar] [CrossRef]
- Weller, P.; Breithaupt, D.E. Identification and quantification of zeaxanthin esters in plants using liquid chromatograph-mass spectrometry. J Agric. Food Chem. 2003, 51, 7044–7049. [Google Scholar] [CrossRef]
- Zhou, L.; Leung, I.; Tso, M.O.; Lam, K.W. The identification of dipalmityl zeaxanthin as the major carotenoid in Gou Qi Zi by high pressure liquid chromatography and mass spectrometry. J. Ocul. Pharmacol. Ther. 1999, 15, 557–565. [Google Scholar] [CrossRef] [PubMed]
- Breithaupt, D.E.; Weller, P.; Wolters, M.; Hahn, A. Comparison of plasma responses in human subjects after the ingestion of 3R, 3R′-zeaxanthin dipalmitate from wolfberry (Lycium barbarum) and non-esterified 3R, 3R′-zeaxanthin using chiral high-performance liquid chromatography. Br. J. Nutr. 2004, 91, 707–713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lam, K.W.; But, P. The content of zeaxanthin in Gou Qi Zi, a potential health benefit to improve visual acuity. Food Chem. 1999, 67, 173–176. [Google Scholar] [CrossRef]
- Leung, I.Y.F.; Ngai, J.; Lam, K.W.; Tso, M.O.M. Absorption of zeaxanthin in rats after feeding with purified zeaxanthin or a traditional Chinese medicine, Gou Qi Zi. In Vest. Ophthalmol. Vis. Sci. 1999, 40, S608. [Google Scholar]
- Wingerath, T.; Stahl, W.; Kirsch, D.; Kaufmann, R.; Sies, H. Fruit juice carotenol fatty acid esters and carotenoids as identified by matrix-assisted laser desorption ionization (MALDI) mass spectrometry. J. Agric. Food Chem. 1996, 44, 2006–2013. [Google Scholar] [CrossRef]
- Dey, T.K.; Maiti, I.; Chakraborty, S.; Ghosh, M.; Dhar, P. Enzymatic synthesis of lipophilic lutein-PUFA esters and assessment of their stabilization potential in EPA-DHA rich fish oil matrix. J. food Sci. Technol. 2019, 56, 2345–2354. [Google Scholar] [CrossRef]
- Wu, M.; Feng, Z.; Deng, Y.; Zhong, C.; Liu, Y.; Liu, J.; Zhao, X.; Fu, Y. Liquid antisolvent precipitation: An effective method for ocular targeting of lutein esters. Int. J. Nanomed. 2019, 14, 2667–2681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuang, P.; Zhang, H.; Bajaj, P.R.; Yuan, Q.; Tang, J.; Chen, S.; Sablani, S.S. Physicochemical properties and storage stability of lutein microcapsules prepared with maltodextrins and sucrose by spray drying. J. Food Sci. 2015, 80, E359–E369. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Cheng, H.; Jiang, P.; Yao, Y.; Han, J. Preparation of lutein-loaded particles for improving solubility and stability by Polyvinylpyrrolidone (PVP) as an emulsion-stabilizer. Food Chem. 2014, 156, 123–128. [Google Scholar] [CrossRef] [PubMed]
- Tudor, C.; Bohn, T.; Iddir, M.; Dulf, F.V.; Foc¸san, M.; Rugină, D.O.; Pintea, A. Sea Buckthorn Oil as a Valuable Source of Bioaccessible Xanthophylls. Nutrients 2020, 12, 76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ligia Focsan, A.; Polyakov, N.E.; Kispert, L.D. Supramolecular Carotenoid Complexes of Enhanced Solubility and Stability - The Way of Bioavailability Improvement. Molecules 2019, 24, 3947. [Google Scholar] [CrossRef] [Green Version]
- Aparicio-Ruiz, R.; Mínguez-Mosquera, I.; Rojas, G.; Gandul-Rojas, B. Thermal degradation kinetics of lutein, β-carotene and β-cryptoxanthin in virgin olive oils. J. Food Compos. Anal. 2011, 24, 811–820. [Google Scholar] [CrossRef]
- Verhoeven, A. Sustained energy dissipation in winter evergreens. New Phytol. 2014, 201, 57–65. [Google Scholar] [CrossRef]
- Sofronova, V.E.; Dymova, O.V.; Golovko, T.K.; Chepalov, V.A.; Petrov, K.A. Adaptive changes in pigment complex of Pinus sylvestris needles upon cold acclimation. Russ. J. Plant Physiol. 2016, 63, 433–442. [Google Scholar] [CrossRef]
- Krinsky, N.I.; Johnson, E.J. Carotenoid actions and their relation to health and disease. Mol. Asp. Med. 2005, 26, 459–516. [Google Scholar] [CrossRef]
- Vishwanathan, R.; Iannaccone, A.; Scott, T.M.; Kritchevsky, S.B.; Jennings, B.J.; Carboni, G.; Forma, G.; Satterfield, S.; Harris, T.; Johnson, K.C.; et al. Macular pigment optical density is related to cognitive function in older people. Age Ageing 2014, 43, 271–275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, E.J.; Vishwanathan, R.; Johnson, M.A.; Hausman, D.B.; Davey, A.; Scott, T.M.; Green, R.C.; Miller, L.S.; Gearing, M.; Woodard, J. Relationship between serum and brain carotenoids, α-tocopherol, and retinol concentrations and cognitive performance in the oldest old from the georgia centenarian study. J. Aging Res. 2013, 2013, 951786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feeney, J.; Finucane, C.; Savva, G.M.; Cronin, H.; Beatty, S.; Nolan, J.M.; Kenny, R.A. Low macular pigment optical density is associated with lower cognitive performance in a large, population-based sample of older adults. Neurobiol. Aging 2013, 34, 2449–2456. [Google Scholar] [CrossRef]
- Bovier, E.R.; Renzi, L.M.; Hammond, B.R. A double-blind, placebo-controlled study on the effects of lutein and zeaxanthin on neural processing speed and efficiency. PLoS ONE 2014, 9, e108178. [Google Scholar] [CrossRef]
- Johnson, E.J.; McDonald, K.; Caldarella, S.M. Cognitive findings of an exploratory trial of docosahexaenoic acid and lutein supplementation in older women. Nutr. Neurosci. 2008, 11, 75–83. [Google Scholar] [CrossRef]
- Stahl, W.; Seis, H. Effects of carotenoids and retinoids on gap junctional communication. Biofactors 2001, 15, 95–98. [Google Scholar] [CrossRef] [PubMed]
- Hammond, B.R., Jr.; Wooten, B.R. CFF Thresholds: Relation to macular pigment optical density. Ophthalmic Physiol. Opt. 2005, 25, 315–319. [Google Scholar] [CrossRef] [PubMed]
- Fatani, A.J.; Al-Rejaie, S.S.; Parmar, M.Y.; Ahmed, O.M.; Abuohashish, H.M.; Ahmed, M.M. Lutein attenuates diabetic-induced renal damage via inhibiting oxidative and nitrosative stresses. Progr. Nutr. 2017, 19, 57–66. [Google Scholar]
- Hu, B.J.; Hu, Y.N.; Lin, S.; Ma, W.J.; Li, X.R. Application of Lutein and Zeaxanthin in nonproliferative diabetic retinopathy. Int. J. Ophthalmol. 2011, 4, 303–306. [Google Scholar] [PubMed]
- Mares-Perlman, J.A.; Brady, W.E.; Klein, B.E.K.; Klein, R.; Haus, G.J.; Palta, M.; Ritter, L.L.; Shoff, S.M. Diet and nuclear lens opacities. Am. J. Epidemiol. 1995, 141, 322–334. [Google Scholar] [CrossRef]
- Lyle, B.J.; Mares-Perlman, J.A.; Klein, B.E.K.; Klein, R.; Palta, M.; Bowen, P.E.; Greger, J.L. Serum carotenoids and tocopherols and incidence of age-related nuclear cataract. Am. J. Clin. Nutr. 1999, 69, 272–277. [Google Scholar] [CrossRef]
- Olmedilla, B.; Granado, F.; Blanco, I.; Herrero, C.; Vaquero, M.; Millan, I. Serum status of carotenoids and tocopherols in patients with age-related cataracts: A case-control study. J. Nutr. Health Aging 2002, 6, 66–68. [Google Scholar]
- Slattery, M.L.; Benson, J.; Curtin, K.; Ma, K.; Schaeffer, D.; Potte, R.J.D. Carotenoids and colon cancer. Am. J. Clin. Nutr. 2000, 71, 575–582. [Google Scholar] [CrossRef] [Green Version]
- Le Marchand, L.; Franke, A.A.; Custer, L.; Wilkens, L.R.; Cooney, R.V. Lifestyle and nutritional correlates of cytochrome CYP1A2 activity. Inverse associations with plasma lutein and alphatocopherol. Pharmacogenetics 1997, 7, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.M.; Araki, S.; Kim, D.J.; Park, C.B.; Takasuka, N.; Baba-Toriyama, H.; Ota, T.; Nir, Z.; Khachik, F.; Shimidzu, N.; et al. Chemopreventive effects of carotenoids and curcumins on mouse colon carcinogenesis after 1,2-dimethylhydrazine initiation. Carcinogenesis 1998, 19, 81–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haegele, A.D.; Gillette, C.; O’Neill, C.; Wolfe, P.; Heimendinger, J.; Sedlacek, S.; Thompson, H.J. Plasma xanthophyll carotenoids correlate inversely with indices of oxidative DNA damage and lipid peroxidation. Cancer Epidemiol. Biomark. Prev. 2000, 9, 421–425. [Google Scholar]
- Zhang, M.; Holman, C.D.; Binns, C.W. Intake of specific carotenoids and the risk of epithelial ovarian cancer. Br. J. Nutr. 2007, 98, 187–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gohar, U.F.; Iqbal, I.; Shah, Z.; Mukhtar, H.; Zia-Ul-Haq, M. COVID-19: Recent Developments in Therapeutic Approaches. In Alternative Medicine Interventions for COVID-19; Zia-Ul-Haq, M., Bin-Jumah, M.N., Alothamn, S.I., Henidi, H.A., Eds.; Springer: Cham, Switzerland, 2021; pp. 249–274. [Google Scholar]
- Wang, Q.; Wang, X.; Hernandez, A.; Kim, S.; Evers, B.M. Inhibition of the phosphatidylinositol 3-kinase pathway contributes to HT29 and Caco-2 intestinal cell differentiation. Gastroenterology 2001, 120, 1543–1546. [Google Scholar] [CrossRef]
- Rock, C.L.; Saxe, G.A.; Ruffin, M.T.; August, D.A.; Schottenfeld, D. Carotenoids, vitamin A, and estrogen receptor status in breast cancer. Nutr. Cancer 1996, 25, 281–296. [Google Scholar] [CrossRef]
- Ribaya-Mercado, J.D.; Blumberg, J.B. Lutein and Zeaxanthin and Their Potential Roles in Disease Prevention. J. Am. Coll. Nutr. 2004, 23, 567S–587S. [Google Scholar] [CrossRef]
- Gong, X.; Smith, J.; Swanson, H.; Rubin, L. Carotenoid Lutein Selectively Inhibits Breast Cancer Cell Growth and Potentiates the Effect of Chemotherapeutic Agents through ROS-Mediated Mechanisms. Molecules 2018, 23, 905. [Google Scholar] [CrossRef] [Green Version]
- Park, J.S.; Chew, B.P.; Wong, T.S. Dietary lutein from marigold extract inhibits mammary tumor development in BALB/c mice. J. Nutr. 1998, 128, 1650–1656. [Google Scholar] [CrossRef] [PubMed]
- Gao, B.; Ahmad, M.F.; Nagy, L.E.; Tsukamoto, H. Inflammatory pathways in alcoholic steatohepatitis. J. Hepatol. 2019, 70, 249–259. [Google Scholar] [CrossRef] [Green Version]
- Xiao, J.; Wang, J.; Xing, F.; Han, T.; Jiao, R.; Liong, E.C.; Fung, M.-L.; So, K.-F.; Tipoe, G.L. Zeaxanthin dipalmitate therapeutically improves hepatic functions in an alcoholic fatty liver disease model through modulating MAPK pathway 9, no. 4, Article ID e95214, 2014. PLoS ONE 2014, 9, e95214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chamberlain, S.M.; Hall, J.D.; Patel, J. Protective effects of the carotenoid zeaxanthin in experimental nonalcoholic steatohepatitis. Dig. Dis. Sci. 2009, 54, 1460–1464. [Google Scholar] [CrossRef]
- Zielińska, M.A.; Wesołowska, A.; Pawlus, B.; Hamułka, J. Health Effects of Carotenoids during Pregnancy and Lactation. Nutrients 2017, 9, 838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahmassani, H.A.; Switkowski, K.M.; Scott, T.M.; Johnson, E.J.; Rifas-Shiman, S.L.; Oken, E.; Jacques, P.F. Maternal Intake of Lutein and Zeaxanthin during Pregnancy Is Positively Associated with Offspring Verbal Intelligence and Behavior Regulation in Mid-Childhood in the Project Viva Cohort. J. Nutr. 2021, 151, 615–627. [Google Scholar] [CrossRef]
- Mortensen, A. Supplements. In Carotenoids. Nutrition and Health; Britton, G., Pfander, H., Liaaen-Jensen, S., Eds.; Birkhäuser: Basel, Switzerland, 2009; Volume 5, pp. 67–82. [Google Scholar]
- Brantsæter, A.L.; Haugen, M.; Rasmussen, S.E.; Alexander, J.; Samuelsen, S.O.; Meltzer, H.M. Urine flavonoids and plasma carotenoids in the validation of fruit, vegetable and tea intake during pregnancy in the Norwegian Mother and Child Cohort Study (MoBa). Public Health Nutr. 2007, 10, 838–847. [Google Scholar] [CrossRef] [PubMed]
- Soy, M.; Keser, G.; Atagündüz, P.; Tabak, F.; Atagündüz, I.; Kayhan, S. Cytokine storm in COVID-19: Pathogenesis and overview of anti-inflammatory agents used in treatment. Clin. Rheumatol. 2020, 39, 2085–2094. [Google Scholar] [CrossRef]
- Guzik, B.; Berry, C.; Touyz, R.M.; Kreutz, R.; Wang, D.W.; McInnes, I.B. COVID-19 and the cardiovascular system: Implications for risk assessment, diagnosis, and treatment options. Cardiovasc. Res. 2020, 116, 1666–1687. [Google Scholar] [CrossRef]
- Walston, J.; Xue, Q.; Semba, R.D.; Ferrucci, L.; Cappola, A.R.; Ricks, M.; Guralnik, J.; Fried, L.P. Serum antioxidants, inflammation, and total mortality in older women. Am. J. Epidemiol. 2006, 163, 18–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ozawa, Y.; Sasaki, M.; Takahashi, N.; Kamoshita, M.; Miyake, S.; Tsubota, K. Neuroprotective effects of lutein in the retina. Curr. Pharm. Des. 2012, 18, 51–56. [Google Scholar] [CrossRef] [Green Version]
- Ambati, J.; Anand, A.; Fernandez, S.; Sakurai, E.; Lynn, B.C.; Kuziel, W.A.; Rollins, B.J.; Ambati, B.K. An animal model of age-related macular degeneration in senescent Ccl-2- or Ccr-2-deficient mice. Nat. Med. 2003, 9, 1390–1397. [Google Scholar] [CrossRef] [PubMed]
- Kamoshita, M.; Toda, E.; Osada, H.; Narimatsu, T.; Kobayashi, S.; Tsubota, K.; Ozawa, Y. Lutein acts via multiple antioxidant pathways in the photo-stressed retina. Sci. Rep. 2016, 6, 30226. [Google Scholar] [CrossRef] [PubMed]
- Sachdeva, M.M.; Cano, M.; Handa, J.T. Nrf 2 signaling is impaired in the aging RPE given an oxidative insult. Exp. Eye Res. 2014, 119, 111–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, M.; An, C.; Gao, Y.; Leak, R.K.; Chen, J.; Zhang, F. Emerging roles of Nrf 2 and phase II antioxidant enzymes in neuroprotection. Prog. Neurobiol. 2013, 100, 30–47. [Google Scholar] [CrossRef] [Green Version]
- Itoh, K.; Wakabayashi, N.; Katoh, Y.; Ishii, T.; Igarashi, K.; Engel, J.D.; Yamamoto, M. Keap1 represses nuclear activation of antioxidant responsive elements by Nrf 2 through binding to the amino-terminal Neh2 domain. Genes Dev. 1999, 13, 76–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Z.; Dong, Z.; Liu, H.; Chen, X.; Shi, H.; Fan, Y.; Hou, D.; Zhang, X. Astaxanthin protects ARPE-19 cells from oxidative stress via upregulation of Nrf 2-regulated phase II enzymes through activation of PI3K/Akt. Mol. Vis. 2013, 19, 1656–1666. [Google Scholar]
- Frede, K.; Ebert, F.; Kipp, A.P.; Schwerdtle, T.; Baldermann, S. Lutein Activates the Transcription Factor Nrf2 in Human Retinal Pigment Epithelial Cells. J. Agric. Food Chem. 2017, 65, 5944–5952. [Google Scholar] [CrossRef] [Green Version]
- Sun, J.K.; Glassman, A.R.; Beaulieu, W.T.; Stockdale, C.R.; Bressler, N.M.; Flaxel, C.; Gross, J.G.; Shami, M.; Jampol, L.M. Diabetic Retinopathy Clinical Research Network, Rationale and Application of the Protocol S Anti-Vascular Endothelial Growth Factor Algorithm for Proliferative Diabetic Retinopathy. Ophthalmology 2019, 126, 87–95. [Google Scholar] [CrossRef]
- Keegan, G.; Pardhan, S.; Chichger, H. Lutein and zeaxanthin attenuates VEGF-induced neovascularisation in human retinal microvascular endothelial cells through a Nox4-dependent pathway. Exp. Eye Res. 2020, 197, 108104. [Google Scholar] [CrossRef]
- Serrander, L.; Cartier, L.; Bedard, K.; Banfi, B.; Lardy, B.; Plastre, O.; Sienkiewicz, A.; Forro, L.; Schlegel, W.; Krause, K.H. NOX4 activity is determined by mRNA levels and reveals a unique pattern of ROS generation. Biochem. J. 2007, 406, 105–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaneko, H.; Dridi, S.; Tarallo, V. DICER1 deficit induces Alu RNA toxicity in age-related macular degeneration. Nature 2011, 471, 325–330. [Google Scholar] [CrossRef] [Green Version]
- Halliwell, B.; Clement, M.V.; Long, L.H. Hydrogen peroxide in the human body. FEBS Lett. 2000, 486, 10–13. [Google Scholar] [CrossRef] [Green Version]
- Zareba, M.; Raciti, M.W.; Henry, M.M.; Sarna, T.; Burke, J.M. Oxidative stress in ARPE-19 cultures: Do melanosomes confer cytoprotection? Free Radic. Biol. Med. 2006, 40, 87–100. [Google Scholar] [CrossRef]
- Chong, Y.S.; Mai, C.W.; Leong, C.O.; Wong, L.C. Lutein improves cell viability and reduces Alu RNA accumulation in hydrogen peroxide challenged retinal pigment epithelial cells. Cutan. Ocul. Toxicol. 2018, 37, 52–60. [Google Scholar] [CrossRef] [PubMed]
- Kuo, L.M.; Chen, P.J.; Sung, P.J. The bioactive extract of Pinnigorgia sp induces apoptosis of hepatic stellate cells via ROS-ERK/JNK-Caspase-3 signaling. Mar. Drugs 2018, 16, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krasilnikov, M.; Ivanov, V.N.; Dong, J. ERK and PI3K negatively regulate STAT-transcriptional activities in human melanoma cells: Implications towards sensitization to apoptosis. Oncogene 2003, 22, 4092–4101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poage, M.; Le Martret, B.; Jansen, M.A.; Nugent, G.D.; Dix, P.J. Modification of reactive oxygen species scavenging capacity of chloroplasts through plastid transformation. Plant. Mol. Biol. 2011, 76, 371–384. [Google Scholar] [CrossRef]
- Sheng, Y.N.; Luo, Y.H.; Liu, S.B.; Xu, W.T.; Zhang, Y.; Zhang, T.; Xue, H.; Zuo, W.B.; Li, Y.N.; Wang, C.Y.; et al. Zeaxanthin Induces Apoptosis via ROS-Regulated MAPK and AKT Signaling Pathway in Human Gastric Cancer Cells. OncoTargets Ther. 2020, 13, 10995–11006. [Google Scholar] [CrossRef]
- Wang, K.; Chu, D.; Wu, J.; Zhao, M.; Zhang, M.; Li, B.; Du, W.; Du, J.; Guo, R. Cinobufagin induced cell apoptosis and protective autophagy through the ROS/MAPK signaling pathway. Life Sci. 2019. [Google Scholar] [CrossRef]
- Rapp, L.M.; Maple, S.S.; Choi, J.H. Lutein and zeaxanthin concentrations in rod outer segment membranes from perifoveal and peripheral human retina. Invest. Ophthalmol. Vis. Sci. 2000, 41, 1200–1209. [Google Scholar]
- Dufossé, L.; Galaup, P.; Yaron, A.; Arad, S.M.; Blanc, P.; Murthy, K.N.C.; Ravishankar, G.A. Microorganisms and microalgae as sources of pigments for food use: A scientifc oddity or an industrial reality? Trends Food Sci. Technol. 2005, 16, 389–406. [Google Scholar] [CrossRef]
- Karmakar, A.; Das, A.K.; Ghosh, S.; Sil, P.C. Carotenoids as Coloring Agents. In Carotenoids: Structure and Function in the Human Body; Zia-Ul-Haq, M., Dewanjee, S., Riaz, M., Eds.; Springer: Cham, Switzerland, 2021. [Google Scholar] [CrossRef]
- Saini, R.K.; Keum, Y.S. Progress in microbial carotenoids production. Indian J. Microbiol. 2017, 57, 129–130. [Google Scholar] [CrossRef] [PubMed]
- Lebeau, J.; Venkatachalam, M.; Fouillaud, M.; Petit, T.; Vinale, F.; Dufossé, L.; Yanis, C. Production and new extraction method of polyketide red pigments produced by Ascomycetes fungi from terrestrial and marine habitats. J. Fungi 2017, 3, 34. [Google Scholar] [CrossRef] [Green Version]
- Dayal, R.; Dobhal, P.C. Natural dyes from some Indian plants. Colourage 2001, 48, 33–38. [Google Scholar]
- Adeel, S.; Gulzar, T.; Azeem, M.; Rehman, F.; Saeed, M.; Hanif, I.; Iqbal, N. Appraisal of marigold flower based leutin as natural colourant for textile dyeing under the influence of gamma radiations. Radiat. Phys. Chem. 2017, 130, 35–39. [Google Scholar] [CrossRef]
- Levy, L.; Inventor, W. Trans-Xanthophyll Ester Concentrates of Enhanced Purity and Methods of Making Same. Inexa Industria Extractora, C.A., Assignee. US Patent 6191293B1, 20 February 2001. [Google Scholar]
- Bailey, C.A.; Chen, B.H. Chromatographic analyses of xanthophylls in egg yolks from laying hens fed turf Bermuda grass (Cynodon dactylon) meal. J. Food Sci. 1989, 54, 584–586. [Google Scholar] [CrossRef]
- Hencken, H. Chemical and physiological behavior of feed carotenoids and their effects on pigmentation. Poult. Sci. 1992, 71, 711–717. [Google Scholar] [CrossRef]
- Tyczkowski, J.K.; Hamilton, P.B. Absorption, transport, and digestion in chickens of lutein diester, a carotenoid extracted from marigold (Tagetes erecta) petals. Poult. Sci. 1986, 65, 1526–1531. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Xue, M.; Wang, J.; Wu, X.; Zheng, Y.; Cao, C. Effect of natural xanthophylls on growth performance and body pigmentation of yellow catfish (Pelteobagrus fulvidraco). J. Fish. China 2012, 36, 1102–1110. [Google Scholar] [CrossRef]
- Nelis, J.H.; DeLeenheer, P.A. Microbial sources of carotenoid pigments used in foods and feeds. J. Appl. Bacteriol. 1991, 70, 181–191. [Google Scholar] [CrossRef]
- Tabor, A.; Blair, R. Nutritional Cosmetics: Beauty from within; William Andrew: Norwich, NY, USA, 2009; pp. 185–198. [Google Scholar]
- Bin-Jumah, M.; Alwakeel, S.S.; Moga, M.; Buvnariu, L.; Bigiu, N.; Zia-Ul-Haq, M. Application of Carotenoids in Cosmetics. In Carotenoids: Structure and Function in the Human Body; Zia-Ul-Haq, M., Dewanjee, S., Riaz, M., Eds.; Springer: Cham, Switzerland, 2021. [Google Scholar] [CrossRef]
- Shen, H.J.; Cheng, B.Y.; Zhang, Y.M.; Tang, L.; Li, Z.; Bu, Y.F.; Li, X.R.; Tian, G.Q.; Liu, J.Z. Dynamic control of the mevalonate pathway expression for improved zeaxanthin production in Escherichia coli and comparative proteome analysis. Metab. Eng. 2016, 38, 180–190. [Google Scholar] [CrossRef]
- Du, W.; Song, Y.; Liu, M.; Yang, H.; Zhang, Y.; Fan, Y.; Luo, X.; Li, Z.; Wang, N.; He, H.; et al. Gene expression pattern analysis of a recombinant Escherichia coli strain possessing high growth and lycopene production capability when using fructose as carbon source. Biotechnol. Lett. 2016, 38, 1571–1577. [Google Scholar] [CrossRef] [PubMed]
- Furubayashi, M.; Ikezumi, M.; Takaichi, S.; Maoka, T.; Hemmi, H.; Ogawa, T.; Saito, K.; Tobias, A.V.; Umeno, D. A highly selective biosynthetic pathway to non-natural C 50 carotenoids assembled from moderately selective enzymes. Nat. Commun. 2015, 6, 7534. [Google Scholar] [CrossRef] [PubMed] [Green Version]





| Microalgae | Culture Conditions | Biomass | Lutein Yield | Zeaxanthin Yield | Other Carotenoids | References |
|---|---|---|---|---|---|---|
| Division Chlorophyta | ||||||
| Chlorella pyrenoidosa | Fresh water cultures | N.A | 125034.4 µg/g | 2170.3µg/g | cis isomers of β-carotene, all trans- β -carotene, cisisomers of α-carotene, β –cryptoxanthin, neoxanthin and its cis isomers, neochrome, auroxanthin, violaxanthin and its cis isomers | [63] |
| Chlorella sorokiniana | Fresh water cultures | 1.98 g/L/d | 6490 µg/g) | N.A | Astaxanthin | [64] |
| Chlorella salina | Fresh water cultures | 665.89 mg | 9.73 mg/L/day | N.A | N.A | [65] |
| Chlorella zofingiensis | Fresh water cultures | 7 g/L | 4000 µg/g | 7000 ± 820 µg/g | monoester of astaxanthin and canthaxanthin | [66,67] |
| Chlorella vulgaris | Fresh water cultures | N.A | 3360 µg/g | N.A | Astaxanthin 12.5% TC violaxanthin | [68] |
| Dunaliella salina zea1 (mutant) | Fresh water | N.A | N.A | 200 µg/g | Astaxanthin and violaxanthin | [69] |
| Chlorella protothecoides | Fresh water | 19.6 g/L | 68.42 and 83.81 mg/L | N.A | N.A | [70] |
| Division Rhodophyta | ||||||
| Porphyra acanthophora var. brasiliensis | Saline condition | N.A | 4.16–30.71 µg/g | 4.43–36.31 µg/g | N.A | [71] |
| Pyropia yezoensis: | Marine culture | 3460 570 µg/g | 2.36–17.58 µg/g | α-carotene, α-cryptoxanthin (β,ε-caroten-3′-ol), and zeinoxanthin (β,ε-caroten-3-ol) | [72] | |
| Porphyra sp. | Marine culture | N.A | 430–1117 µg/g | N.A | N.A | [73] |
| Corallina elongata | Marine culture | N.A | 1.3 ± 0.6% | 4.7 ± 0.1% | Anteraxanthin | [74] |
| Rhodymenia corallina. | Marine Culture | N.A | 3.4 ± 0.3 µg/g | N.A | Trans-β-carotene, Cis-β-carotene, | [75] |
| Division Euglenophyta | ||||||
| Euglena gracilis | Fresh water | 0.5 g/L | 700 mg/100g | N.A | Neoxanthin and β-carotene. | [76] |
| Family Name | Microorganism | Major Carotenoid Content | Other Carotenoids | General Applications | References |
|---|---|---|---|---|---|
| Flavobacteriaceae | Flavobacterium sp. P8 strain | 205 μg/g | β-cryptoxanthin and β-carotene | Prevention of macular degeneration | [87] |
| Flavobacterium multivorum | (0.05 μg/mL/h) | β-cryptoxanthin and β-carotene | Antioxidant | [88] | |
| 10.65 ± 0.63 μg/mL | β-cryptoxanthin and β-carotene | For commercialization of zeaxanthin based products | [89] | ||
| Flavobacterium sp. | 1 mg/L | β-cryptoxanthin and β-carotene | Natural food colorant for fish and poultry | [90] | |
| Mesoflavibacter zeaxanthinifaciens | 0.91 mg/g | Yellow carotenoid pigments | For use as a natural colorant in feed industry | [91] | |
| Mesoflavibacter aestuarii sp. | 1200 mg/100 mg | N.A | Antioxidant | [92] | |
| Aquibacter zeaxanthinifaciens | N.A | Some unidentified carotenoids | Antioxidant | [93] | |
| Enterobacteriaceae | E. coli (metabolic engineered strain) | 1.6 mg/g | β-carotene | Antioxidant | [94] |
| E. coli | 11.95 mg/g | β-cryptoxanthin and β-carotene | Antioxidant | [95] | |
| Sphingobacteriaceae | Nubsella zeaxanthinifaciens | 0.8 mg/g | N.A | Natural colorant | [96] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zafar, J.; Aqeel, A.; Shah, F.I.; Ehsan, N.; Gohar, U.F.; Moga, M.A.; Festila, D.; Ciurea, C.; Irimie, M.; Chicea, R. Biochemical and Immunological implications of Lutein and Zeaxanthin. Int. J. Mol. Sci. 2021, 22, 10910. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222010910
Zafar J, Aqeel A, Shah FI, Ehsan N, Gohar UF, Moga MA, Festila D, Ciurea C, Irimie M, Chicea R. Biochemical and Immunological implications of Lutein and Zeaxanthin. International Journal of Molecular Sciences. 2021; 22(20):10910. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222010910
Chicago/Turabian StyleZafar, Javaria, Amna Aqeel, Fatima Iftikhar Shah, Naureen Ehsan, Umar Farooq Gohar, Marius Alexandru Moga, Dana Festila, Codrut Ciurea, Marius Irimie, and Radu Chicea. 2021. "Biochemical and Immunological implications of Lutein and Zeaxanthin" International Journal of Molecular Sciences 22, no. 20: 10910. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222010910